

Abstract
Rheumatoid arthritis (RA) is an autoimmune disease which has a significant socio-economic impact. The aim of the current study was to investigate eight candidate RA susceptibility loci to identify the associated variants in Egyptian population. Eight single nucleotide polymorphisms (SNPs) (MTHFR—C677T and A1298C, TGFβ1 T869C, TNFB A252G, and VDR—ApaI, BsmI, FokI, and TaqI) were tested by genotyping patients with RA (n = 105) and unrelated controls (n = 80). Associations were tested using multiplicative, dominant, recessive, and co-dominant models. Also, the linkage disequilibrium (LD) between the VDR SNPs was measured to detect any indirect association. By comparing RA patients with controls (TNFB, BsmI, and TaqI), SNPs were associated with RA using all models. MTHFR C677T was associated with RA using all models except the recessive model. TGFβ1 and MTHFR A1298C were associated with RA using the dominant and the co-dominant models. The recessive model represented the association for ApaI variant. There were no significant differences for FokI and the presence of RA disease by the used models examination. For LD results, There was a high D′ value between BsmI and FokI (D′ = 0.91), but the r2 value between them was poor. All the studied SNPs may contribute to the susceptibility of RA disease in Egyptian population except for FokI SNP.
Introduction
Rheumatoid arthritis (RA) is an autoimmune disease which is considered genetically complex. RA is the leading cause of bone loss and chronic inflammation of the joints, most prominently in white populations. The prevalence of the disease in women is twice that in men. RA attacks the body most often at the age of 40. Over the past 40 years, statistical geneticists have facilitated the discovery of RA biomarkers. Multiple methods have been developed to detect the association between the examined SNPs and disease susceptibility. Although HLA-DRB1 was considered the most widely studied and the most associated gene with RA susceptibility, SNPs at other gene loci may contribute to the disease. The genetic etiology of the disease is still an open question [1]. The examined genes in our study were selected for their critical role in immunogenetics and the contradictory results of their SNPs in the association with RA susceptibility in different populations.
The most popular statistical models for measuring the association between a genotype and a phenotype are multiplicative, dominant, recessive, and co-dominant models. These models differ in the identification of the exposed group and the unexposed group in cases and controls as shown in Table 1. For the multiplicative model, analysis should be done using alleles instead of genotypes [2, 3].
Table 1. The Exposed Group and the Unexposed Group in the Four Used Models.
| Model | Exposed | Unexposed |
|---|---|---|
| Multiplicative | Major Allele | Minor Allele |
| Dominant | Major Homozygote | Minor Homozygote & Heterozygote |
| Recessive | Major Homozygote & Heterozygote | Minor Homozygote |
| Co-dominant | Major Homozygote | Minor Homozygote or Heterozygote |
The co-dominant model is the only one comparing the three categorical genotypes without assuming any relationship between disease and genotypes. So, the co-dominant model has two degrees of freedom (DF). All other models have one DF. The one DF tests are more popular than the two DF tests because of the simplicity of the one DF tests implementation through the 2x2 contingency tables. Also, the one DF tests have higher statistical power than the two DF tests [4].
In our study, the examined eight SNPs are located within four genes which are methylene tetrahydrofolate reductase (MTHFR), transforming growth factor beta (TGFβ1), tumor necrosis factor beta (TNFB), and vitamin D receptor (VDR). The C677T and A1298C are common polymorphisms in the MTHFR gene [5]. The MTHFR gene is a candidate biomarker for RA susceptibility. Homocysteine has been recorded at high levels in RA patients, which is related to the MTHFR gene [6]. The allele (677T) has been found to exhibit lower MTHFR enzyme activity and has been implicated in average elevated levels of homocysteine [7]. The 1298C (minor) allele has a lower effect on MTHFR enzyme than the 677T [8, 9].
The T869C is a common polymorphism within the TGFβ1 gene [10]. The TGFβ1 gene is a strong candidate biomarker for RA susceptibility. The TGFβ1 protein has been found in the synovial fluid of RA patients [11]. The T869C polymorphism is associated with the soluble TGFβ1 serum levels [12].
The A252G polymorphism is located at position 1069 of intron 1 of the TNFB gene [13]. The TNFB is considered as proinflammatory immunostimulatory cytokine. The TNFB cytokine has been detected in nine RA patients (four synovial fluid /serum pairs, three synovial fluid and two sera) out of 27 examined RA patients [14]. The A252G polymorphism influences adhesion molecules and cytokines from different types of leukocytes [15].
ApaI, BsmI, FokI, and TaqI are common polymorphisms within the VDR gene [16]. The VDR protein, through the vitamin D endocrine system, has been implicated in the metabolic pathways involved in the immune response. It plays an important role in absorption of calcium, promoting monocyte differentiation, inhibiting lymphocyte proliferation and secretion of cytokines, such as interleukin (IL)-2, interferon gamma (IFNγ), and IL-12 [17, 18].
There are two objectives of our present case-control study. The first objective was to study the direct association between each studied SNP and RA susceptibility. The second objective was to measure the linkage among the four VDR SNPs and/or the two MTHFR SNPs to detect any indirect association in case of no direct association. The studied parameters are linkage disequilibrium (LD) and odds ratio (OR). These two parameters could be measured using case-control samples.
Patients and Methods
Ethics Statement
The study was approved by the Ethical Committee of Faculty of Medicine, Cairo University, and an oral and written informed consent was obtained from all participants.
Patients
In total, 185 subjects were enrolled in the case-control study: 105 RA patients (89 women and 16 men) and 80 unrelated ethnically matched healthy controls (69 women and 11 men). All subjects in our analysis were Egyptians and were recruited from Rheumatology Department and Outpatient Clinics of Cairo University Hospitals (Kasr El-Aini hospital). RA patients were diagnosed by physician investigators and followed the 1987 American College of Rheumatology (ACR) criteria [19]. DAS28 (Disease Activity Score in 28 Joints), which is a validated score for established RA, was used as a measure for disease activity. No signs of RA, such as joint stiffness in the morning, positive rheumatoid factor (RF) and citrulline antibody or the findings of rheumatoid nodules were observed in controls. Patients with other autoimmune diseases or inflammatory disorders unrelated to RA were not included.
Molecular Genetic Methods
DNA was extracted from peripheral blood using a QIAamp DNA Blood Mini Kit (Qiagen, Valencia, CA, USA) according to the manufacturer's protocol to be used for genotyping of the eight SNPs MTHFR C677T, MTHFR A1298C, TGFβ1 T869C, TNFB A252G, ApaI, BsmI, FokI, and TaqI.
MTHFR C677T genotyping
One set of forward 5'-CAT CCC TAT TGG CAG GTT AC-3' and reverse 5'-GAC GGT GCG GTG AGA GTG- 3' primers were used for the amplification of a fragment of 265 bp, and then the amplified fragments were digested with the HinfI enzyme. The PCR profile was: initial denaturation at 95°C for 5 min, denaturation at 94°C for 30 sec, annealing at 59°C for 30 sec, extension at 72°C for 30 sec for 35 cycles and followed at 72°C for 10 min. At position 677 (rs1801133) of the MTHFR gene, the C wild base, replaced by the T base, produces a cut site for the HinfI enzyme, which cuts the amplicons into two fragments of 171 and 94 bp. Then, the CC genotype would be reflected by a single band of 265 bp (uncut), the CT genotype by three bands of 265, 171 and 94 bp, and the TT genotypes by two bands of 171 and 94 bp.
MTHFR A1298C genotyping
One set of forward 5'-CTT TGG GGA GCT GAA GGA CTA CTA C-3' and reverse 5'-CAC TTT GTG ACC ATT CCG GTT TG-3' primers was used for the amplification of a fragment of 241 bp and then the amplified fragment was digested with the MboII enzyme. The PCR profile was: initial denaturation at 95°C for 5 min, denaturation at 94°C for 30 sec, annealing at 51°C for 30 sec, extension at 72°C for 30 sec for 35 cycles and followed at 72°C for 10 min. At position 1298 (rs1801131) of the MTHFR gene, the transversion of the wild A base, to C base produces a cut site for the MboII enzyme, which cuts the PCR product into two fragments of 211 and 30 bp. Then, the AA genotype results in a single band of 241 bp (uncut), the AC genotype produces three bands of 241, 211 and 30 bp, and the CC genotype produces two bands of 211 and 30 bp. The digestion of 10 μl of PCR products was carried out with 1.5 U of the MboII restriction enzyme in 37°C for two hours.
TGFβ1 T869C genotyping
DNA was genotyped by specific primers: 5'-TTCCCTCGAGGCCCTCCTA-3' and 5'-GCCGCAGCTTGGACAGGATC-3' to amplify a fragment of the TGFβ1 gene (rs1982073), with denaturation at 96°C for 10 min, followed by 35 cycles at 96°C for 75 sec, 62°C for 75 sec, 73°C for 75 sec, and a final extension at 73°C for five min. MspA1I (New England Biolabs, Hitchin, UK) digestion of the 294 bp fragments at 37°C for 3 hours resulted in fragments of the T allele of 161, 67, 40, and 26 bp, and the C allele of 149, 67, 40, 26, and 12 bp. The samples were then analyzed by electrophoresis on 4% agarose gel stained with ethidium bromide and the genotypes were determined.
TNFB A252G genotyping
Genotypes for TNFB (rs909253) were determined by polymerase chain reaction-restriction fragment length polymorphism (PCR-RFLP). Specific oligonucleotide primers were used: 5'-CCGTGCTTCGTGCTTTGGACTA-3' and 5'-AGAGGGGTGGATGCTTGGGTTC-3', 782 bp fragments were amplified for the first intron of the TNFB gene. PCR products were digested with the NcoI restriction enzyme and analyzed on 2% agarose gel. The TNFB digested product generated fragments of 586 and 196 bp or 782 bp for TNFB*1 or TNFB*2 homozygous individuals, respectively. For heterozygous individuals, three fragments (196, 586 and 782 bp) are detected.
VDR (ApaI, BsmI, FokI, and TaqI) genotyping
The VDR ApaI (rs7975232), the BsmI (rs1544410), the FokI (rs2228570), and the TaqI (rs731236) SNPs were detected by PCR–RFLP according to the manufacturer’s instructions (New England BioLabs, Ipswich, USA). DNA digested fragments were separated in 3% agarose gels and visualized by ethidium bromide staining. Primer sequences and conditions for PCR–RFLP analyses were presented in S1 Table, as described earlier [20].
Materials
The marker checks, the normalized D (D′), and the correlation between pair of SNPs (r2) were measured using Haploview 4.2 (from Daly Lab at the Broad Institute, Cambridge, MA 02141, USA) [21]. The odds ratio (OR), its confidence interval (CI), and Pearson chi square (χ2) test for goodness of fit were measured using SNPAnalyzer 2.0 (from Bioinformatics Unit, ISTECH Inc., Republic of Korea) [22, 23]. The statistical power was measured using Genetic Power Calculator (from the Purcell lab, http://pngu.mgh.harvard.edu/~purcell/gpc/) [24].
Statistical Methods
The used bi-allelic marker checks through this study were a) genotype percentage, b) minor allele frequency, c) heterozygosity, and d) Hardy-Weinberg equilibrium P-value. The association between the eight genetic polymorphisms and susceptibility to RA was assessed by the ORs with their corresponding 95% CI under four genetic models including the multiplicative model, the dominant model, the recessive model, and the co-dominant model. A two-sided p-value less than 0.01 was considered statistically significant. The input parameters used in calculating power could be listed as high risk allele frequency, disease prevalence in the general population (1%), genotypic relative risks, type I error rate (0.05), no. of cases (105), and no. of controls (80). Also, the indirect association between any of the eight genetic polymorphisms and susceptibility to RA was detected by D′ and r2 which are the most common measures of LD. An indirect association was considered when D′ value was more than or equal 0.8 and r2 value was more than or equal 0.8 between a directly associated SNP and an unassociated SNP.
The flow chart shown in Fig 1 illustrated the proposed association scheme. The LD should be measured between bi-allelic SNPs that lie on the same chromosome. So, LD could be measured between MTHFR (C677T and A1298C) SNPs to detect any indirect variant. Also, the VDR (ApaI, BsmI, FokI, and TaqI) SNPs were appropriate for LD study as they were all located on chromosome 12.
Fig 1. Flow Chart of the Proposed Study.

The eight SNPs were genotyped to detect the association with RA susceptibility either directly or indirectly. The SNP that fails to associate with the phenotype directly will undergo an LD study to detect any indirect association as a surrogate for direct disease association.
Results
Eighty six percent of control group were females. Eighty five percent of RA patients were females. The mean age of RA women ± standard deviation (SD) was 40.84 ± 11.01 years. The mean age of RA men ± SD was 52.25 ± 13.46 years. The average disease duration of RA women ± SD was 6.24 ± 4.17 years. The average disease duration of RA men ± SD was 2.85 ± 4.38 years.
We conducted a check for conformance with HWE, MAF, and the percentage of individual successfully genotyped for each SNP. The minimum accepted genotype percentage was 75%. The markers that were significantly deviated from HWE (HW p-value < 0.005) were excluded. The minimum accepted MAF was 0.01. The results of the marker checks stage are shown in Table 2. All the SNPs for all individuals were fully genotyped and all SNPs passed the marker checks stage. Also, information about each SNP (ID, physical position, chromosome no., major allele, and minor allele) was provided in Table 2. At last, a data set including 1480 SNPs corresponding to 185 uncorrelated individuals was utilized in our study. The power results of the study were shown in S2 Table.
Table 2. Marker Checks of the Studied SNPs.
| MTHFR C677T | MTHFR A1298C | TGFβ1 869C | TNFB A252G | ApaI | BsmI | FokI | TaqI | |
|---|---|---|---|---|---|---|---|---|
| ID | rs1801133 | rs1801131 | rs1982073 | rs909253 | rs7975232 | rs1544410 | rs2228570 | rs731236 |
| Position | 11856378 | 11854476 | 41858921 | 31540313 | 48238837 | 48239835 | 48272895 | 48238757 |
| Chromosome | 1 | 1 | 19 | 6 | 12 | 12 | 12 | 12 |
| Alleles Major:Minor | C:T | A:C | T:C | A:G | G:T | G:A | C:T | T:C |
| Genotype % | 100% | 100% | 100% | 100% | 100% | 100% | 100% | 100% |
| MAF | 0.254 | 0.416 | 0.473 | 0.343 | 0.341 | 0.4 | 0.219 | 0.395 |
| ObsHET | 0.378 | 0.465 | 0.546 | 0.459 | 0.422 | 0.476 | 0.362 | 0.465 |
| PredHET | 0.379 | 0.486 | 0.499 | 0.451 | 0.449 | 0.48 | 0.342 | 0.478 |
| HW P-value | 1 | 0.6329 | 0.2667 | 0.9567 | 0.4773 | 0.9934 | 0.5924 | 0.8002 |
Table 3 represented the association between the examined SNPs and RA disease. A graphical representation of the association results for the studied SNPs was shown in Fig 2. The red color in Fig 2 demonstrated a statistically significant SNP. From Table 3 and Fig 2, TNFB, BsmI, and TaqI showed significant association with RA susceptibility with the four used models. MTHFR C677T expressed significant association with RA susceptibility with all models except the recessive model. TGFβ1 and MTHFR A1298C showed significant association with RA susceptibility with the dominant and co-dominant models. ApaI imposed significant association with RA susceptibility with only the recessive model. FokI did not show any significant association with RA directly with any of the used models.
Table 3. Case-Control Study—SNP Analysis.
| MTHFR C677T | MTHFR A1298C | TGFβ1 T869C | TNFB A252G | ApaI | BsmI | FokI | TaqI | ||
|---|---|---|---|---|---|---|---|---|---|
| Multiplicative Model | P | 0.001 | 0.348 | 0.233 | 6.435E-05 | 0.151 | 2.711E-04 | 0.307 | 6.588E-04 |
| χ2 | 10.825 | 0.88 | 1.423 | 15.97 | 2.063 | 13.26 | 1.044 | 11.602 | |
| OR (95% CI) | 0.433(0.2610.718) | 1.221(0.8051.852) | 0.778(0.5151.176) | 2.424(1.5633.759) | 0.725(0.468–1.125) | 2.188(1.4313.346) | 0.769(0.4641.274) | 2.082(1.3613.183) | |
| Dominant Model | P | 1.975E-04 | 0.005 | 0.003 | 2.634E-04 | 0.974 | 0.002 | 0.545 | 0.007 |
| χ2 | 13.854 | 8.017 | 8.747 | 13.314 | 0.001 | 9.483 | 0.367 | 7.356 | |
| OR (95% CI) | 0.315(0.1690.584) | 2.503(1.3164.763) | 0.364(0.1840.72) | 3.131(1.6795.838) | 0.99(0.5521.777) | 2.704(1.4225.139) | 0.832(0.4581.51) | 2.366(1.2614.437) | |
| Recessive Model | P | 0.474 | 0.072 | 0.266 | 0.006 | 0.005 | 0.005 | 0.235 | 0.005 |
| χ2 | 0.513 | 3.247 | 1.239 | 7.668 | 7.936 | 8.004 | 1.411 | 8.004 | |
| OR (95% CI) | 0.638(0.1852.199) | 0.482(0.216–1.077) | 1.505(0.731–3.102) | 3.808(1.405–10.321) | 0.224(0.073–0.684) | 3.167(1.388–7.226) | 0.209(0.025–1.771) | 3.167(1.388–7.226) | |
| Co-dominant Model | P | 9.054E-04 | 2.153E-04 | 0.001 | 2.895E-04 | 0.011 | 0.001 | 0.483 | 0.003 |
| χ2 | 14.014 | 16.887 | 13.015 | 16.294 | 8.961 | 13.157 | 1.453 | 11.474 |
Fig 2. Association Analyses for Examined SNPs with RA Disease.

(a) Multiplicative model. (b) Dominant model. (c) Recessive model. (d) Co-dominant model. The horizontal line in each model represents the significance level of the P value (0.01). The figure was generated using the SNPAnalyzer 2.0 program.
Genotype frequencies for each polymorphism for patients and controls were presented in Fig 3. From Table 3 and Fig 3, the genotypes/alleles that increases or decreases the susceptibility for RA could be addressed. Carriers of MTHFR C677T (CC) genotype had a decreased risk of RA. The (CT) genotype and (T) allele of the C677T variant was significantly associated with increased risk of RA. Patients bearing MTHFR A1298C (AA) genotype showed a trend to have an increased risk for RA. The (AC) genotype of the A1298C had a protective role against RA. Carriers of TGFβ1 (TC) genotype had an increased risk of RA. The OR under the dominant model provided an evidence of association for TGFβ1 with RA showing protective role for the (CC) genotype. Carriers of TNFB (AA) genotype and (A) allele had an increased risk of RA whereas individuals harboring the TNFB (AG) and (GG) genotypes were refractory to the disease. The (TT) genotype of the ApaI variant was significantly associated with increased risk of RA. The (GG) genotype of the BsmI predisposed its carrier to RA while the (AA) genotype and (A) allele of the BsmI had a protective role against RA. The (TT) genotype and (T) allele of the TaqI biomarker showed statistical influence in RA patients whereas the (CC) genotype of the TaqI seemed to be protective to RA. There was no direct association between FokI polymorphism and RA due to the comparable frequencies between cases and controls.
Fig 3. Genotype Distributions in RA Patients and Controls.

For the eight SNPs, the genotype frequencies were illustrated as bar charts for cases and controls.
FokI was the only variant that was not associated with RA susceptibility directly. LD between the VDR SNPs was measured to detect whether FokI variant was associated with RA indirectly. LD results for all VDR SNPs were shown in Fig 4. The VDR SNPs were shown in the order in which they appear on the genome. Each D′ value and r2 value in the plot were multiplied by 100. Generally, the nearer SNPs tend to have high D′ values while the SNPs that are farther apart tend to have lower D′ values. The four VDR SNPs were in close proximity. Despite these proximities, the LD results were poor. Only two D′ values were greater than 0.8 which were between (FokI, ApaI) and BsmI. All the r2 values were poor. While the D′ value between FokI and BsmI equaled 0.91, but the r2 value between them equaled 0.15. This was due to one SNP being much rarer than the other. So, the SNPs could not substitute each other. At last, both D′ and r2 values must be specified to take the correct decision.
Fig 4. Pairwise LD Plot for the VDR SNPs.
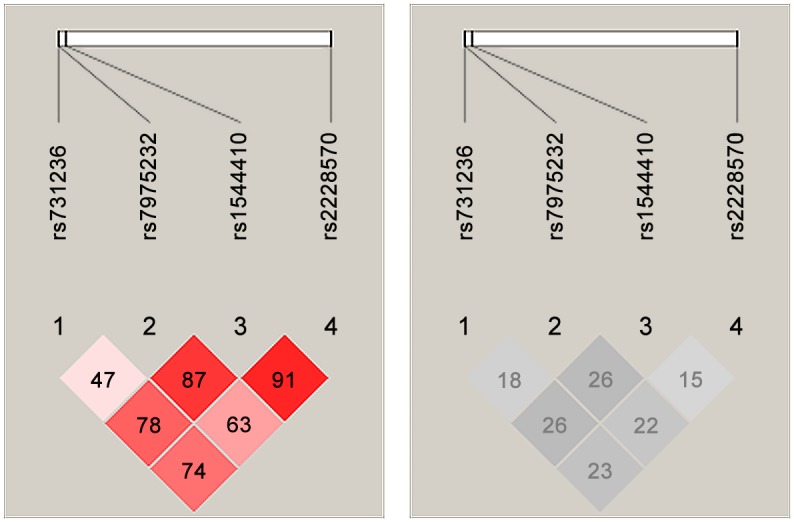
The ID of each SNP was taken as reference. The rs731236, rs7975232, rs1544410, and rs2228570 correspond to TaqI, ApaI, BsmI, and FokI respectively. (a) D′ values. (b) r2 values. The plot was generated using the Haploview 4.2 program.
Discussion
RA is an autoimmune chronic disease that affects body’s joints and bones. RA pathogenesis is an active area of research including several genes. MTHFR, TGFβ1, TNFB, and VDR genes have generated great interest in RA pathogenesis [1, 25–27].
In the present study, the distribution of genotypes and alleles of eight SNPs was used to examine the association with RA susceptibility in the Egyptian population (105 RA patients and 80 healthy controls). The examined SNPs belonged to four genes (MTHFR, TGFβ1, TNFB, and VDR). Seven SNPs were considered candidates for RA susceptibility which are TNFB (rs909253), BsmI (rs1544410), TaqI (rs731236), MTHFR C677T (rs1801133), TGFβ1 (rs1982073), MTHFR A1298C (rs1801131), and ApaI (rs7975232). There was no proof of association for FokI (rs2228570) with RA.
The association between RA and the studied polymorphisms has been examined in several studies. Contradictory results had arisen due to different populations, the age of the subjects, and the sample sizes of these studies. The genetic characteristics of the modern Egyptian population are a mixture of European, Middle Eastern, and African populations [28]. This issue could explain the agreement/disagreement of our results with published data of other populations. Table 4 showed the influential genotype/allele in case of the presence of an association for the studied SNP with RA in the corresponding population. The conflicting results in Table 4 might be due to the small sample size of most of the included studies.
Table 4. Association Status with RA of Our Study and Previous Studies.
| Population | No. of cases | No. of controls | Genotype | Genotype Frequency (cases—controls) | Allele | Allele Frequency (cases—controls) | Association with RA | Reference | |
|---|---|---|---|---|---|---|---|---|---|
| MTHFR C677T | Egyptian (our study) | 105 | 80 | CC | 46–57 | Protective | |||
| CT | 51–19 | T | 67–27 | Susceptible | |||||
| Jewish | 93 | 377 | No association | [29] | |||||
| Italian | 217 | 251 | No association | [5] | |||||
| African-American | 138 | 53 | No association | [7] | |||||
| Caucasian | 393 | 50 | No association | [7] | |||||
| Turkish | 147 | 150 | T | 51–32 | Susceptible | [30] | |||
| MTHFR A1298C | Egyptian (our study) | 105 | 80 | AA | 46–19 | Susceptible | |||
| AC | 35–51 | Protective | |||||||
| Jewish | 93 | 377 | CC | 23–48 | Susceptible | [29] | |||
| Italian | 217 | 251 | CC | 26–12 | Susceptible | [5] | |||
| African-American | 138 | 53 | No association | [7] | |||||
| Caucasian | 393 | 50 | No association | [7] | |||||
| TGFβ1 T869C | Egyptian (our study) | 105 | 80 | TC | 69–32 | Susceptible | |||
| CC | 18–29 | Protective | |||||||
| Japanese (Nagoya) | 155 | 110 | T | 160–101 | Susceptible | [31] | |||
| Japanese (Tsukuba) | 137 | 225 | No association | [10] | |||||
| Chinese | 76 | 100 | CC | 11–33 | C | 63–113 | Protective | [33] | |
| Chinese | 105 | 100 | CC | 17–34 | C | 86–114 | Protective | [34] | |
| Egyptian (Zagazig) | 160 | 168 | T | 215–200 | Susceptible | [32] | |||
| Turkish | 131 | 133 | No association | [35] | |||||
| Caucasian (New Zealand) | 117 | 140 | No association | [36] | |||||
| Caucasian (UK) | 395 | 401 | No association | [37] | |||||
| Korean | 143 | 148 | No association | [38] | |||||
| ApaI | Egyptian (our study) | 105 | 80 | aa (TT) | 20–4 | Susceptible | |||
| Spanish | 120 | 200 | No association | [51] | |||||
| Egyptian (Mansoura) | 128 | 150 | aa (TT) | 26–10 | Susceptible | [57] | |||
| Tunisian | 106 | 153 | No association | [60] | |||||
| BsmI | Egyptian (our study) | 105 | 80 | bb (GG) | 48–19 | Susceptible | |||
| BB (AA) | 10–20 | B (A) | 57–81 | Protective | |||||
| Spanish | 120 | 200 | No association | [51] | |||||
| Korean | 157 | 211 | No association | [52] | |||||
| French | 96 | 96 | No association | [53] | |||||
| Hungarian | 64 | 40 | No association | [54] | |||||
| Turkish | 98 | 122 | No association | [55] | |||||
| Tunisian | 108 | 152 | No association | [56] | |||||
| Egyptian (Mansoura) | 128 | 150 | BB (AA) | 13–36 | B (A) | 78–146 | Protective | [57] | |
| bb (GG) | 63–40 | Susceptible | |||||||
| Egyptian (Zagazig) | 200 | 150 | No association | [58] | |||||
| German | 62 | 40 | No association | [59] | |||||
| FokI | Egyptian (our study) | 105 | 80 | No association | |||||
| French | 100 | 100 | FF (CC) | 45–30 | F (C) | 133–108 | Susceptible | [53] | |
| Turkish | 98 | 122 | No association | [55] | |||||
| Tunisian | 108 | 152 | FF (CC) | 49–46 | F (C) | 147–164 | Susceptible | [56] | |
| Egyptian (Mansoura) | 128 | 150 | No association | [57] | |||||
| German | 62 | 40 | No association | [59] | |||||
| Canadian | 448 | 704 | Ff (CT) | 243–308 | Susceptible | [62] | |||
| Indian | 112 | 125 | No association | [61] | |||||
| TaqI | Egyptian (our study) | 105 | 80 | TT (TT) | 48–21 | T (T) | 143–81 | Susceptible | |
| tt (CC) | 10–20 | Protective | |||||||
| Spanish | 120 | 200 | No association | [51] | |||||
| Korean | 157 | 120 | No association | [52] | |||||
| French | 95 | 95 | No association | [53] | |||||
| Turkish | 98 | 122 | No association | [55] | |||||
| Egyptian (Mansoura) | 128 | 150 | TT (TT) | 64–39 | T (T) | 179–152 | Susceptible | [57] | |
| tt (CC) | 13–37 | Protective | |||||||
| German | 62 | 40 | No association | [59] | |||||
| Tunisian | 106 | 153 | No association | [60] | |||||
| TNFB A252G | Egyptian (our study) | 105 | 80 | AA | 57–22 | A | 156–87 | Susceptible | |
| AG, GG | 42–43, 6–15 | Protective | |||||||
| Belgian | 77 | 58 | No association | ||||||
| Latvian (juvenile RA) | 128 | 114 | No association | ||||||
| Spanish | 60 | 102 | No association | ||||||
| Japanese | 60 | 103 | No association | ||||||
| Caucasian (UK) | 388 | 399 | GG | 77–47 | G | 337–283 | Susceptible | ||
| Portuguese (White) | 388 | 269 | AA | 197–117 | A | 547–346 | Susceptible | ||
| Saudi Arabian | 106 | 126 | AA, GG | 32–15, 27–18 | Susceptible | ||||
| AG | 47–93 | Protective | |||||||
| Tunisian | 108 | 226 | G | 84–131 | Susceptible | ||||
| AA | 37–115 | Protective |
The MTHFR A1298C was verified as a biomarker for RA disease in Jewish and Italian populations [5, 29]. The current results in this article supported the association of A1298C with RA susceptibility. This agreement may be due to these studied populations represent Mediterranean populations. Contradictory results were shown in American population with Caucasian and African ethnicities [7]. The negative findings found in the Americans might be due to the enrichment of the flour products in the US with folic acid since 1998 [1, 5]. The allele (1298C) was found to exhibit lower MTHFR enzyme activity, hyperhomocysteinemia, and decreased folate levels.
The MTHFR C677T was highly suggested for association with RA cases in Italian population [5]. This study demonstrated the association of MTHFR C677T with RA susceptibility in Egyptian population which had been addressed in Turkish population [30]. These similarities might be explained as Egypt and Turkey are Middle Eastern countries.
There were controversial results of TGFβ1 T869C for the susceptibility to RA disease in different populations. The association between T869C variant and RA was confirmed in Japanese (Nagoya), Chinese and Egyptian populations [31–34]. Other results did not show association between RA and T869C in Caucasian (New Zealand and UK), Turkish, Japanese (Tsukuba) and Korean populations [10, 35–38]. Chang et al. conducted a meta-analysis on seven studies and resulted in contradictory outcomes. They concluded that T869C was associated with RA in Asian patients but not in non-Asian patients [39]. This conclusion was confirmed by Zhang et al. [40]. The (TT) genotype represented a risk factor for RA while (CC) genotype or (C) allele seemed to be protective to RA through a meta-analysis conducted by Zhou et al. [41]. TGFβ1 results differed in our study and the study of [32] for the Egyptian population. This variation in results might be due to the sample sizes (our study: cases 105, controls 80 while Hussein et al.: cases 160, controls 168) or the age of patients (our study: 42.71 ± 12.07 years while Hussein et al.: 47.3 ± 9.3 years) or the disease duration (our study: 5.72 ± 4.35 years while Hussein et al.: 10.23 ± 7.5 years).
The results of the association between RA susceptibility and TNFB A252G have proven conflicting in different populations. By analyzing the possible influence of A252G on the susceptibility of RA in Belgian, Japanese, Latvian (juvenile RA), and Spanish populations, the results did not show any significant association [42–45]. The association between A252G variant and RA was confirmed in Caucasian (UK), white Portuguese, Saudi Arabian, and Tunisian populations [13, 46–48]. The current results for TNFB were consistent with the findings in Portuguese (White).
HLA-DRB1 alleles were associated with RA susceptibility in the Egyptian population [49]. TNFB and HLA-DRB1 are in LD, as they are located in the major histocompatibility complex (MHC) region (about 1000 kb apart from each other) [50]. So, the association between TNFB and RA susceptibility in the Egyptian population might be due to the LD between TNFB and HLA-DRB1.
The VDR polymorphisms and RA susceptibility showed unclear relations. There were no associations between VDR SNPs and RA in Spanish (BsmI, ApaI, TaqI), Korean (BsmI, TaqI), French (BsmI, TaqI), Hungarian (BsmI), Turkish (BsmI, TaqI, FokI), Tunisian (ApaI, BsmI, TaqI), Egyptian (Mansoura) (FokI), Egyptian (Zagazig) (BsmI), Indian (FokI), and German (BsmI, TaqI, and FokI) populations [51–61]. Other results showed association between VDR polymorphisms and RA susceptibility in French (FokI), Tunisian (FokI), Egyptian (Mansoura) (BsmI, TaqI, ApaI), and Canadian (North American Natives) (FokI) populations [53, 56, 57, 62].
For VDR gene polymorphisms, our results confirmed the previous results of Mosaad et al. [57]. These two studies supported the confirmation of the major role of (BsmI, TaqI, and ApaI) polymorphisms in RA susceptibility in the Egyptian population. BsmI results differed in our study and the study of [58] for the Egyptian population. This variation may be due to the studied gender (our study: both females & males, Hussein et al.: female only), the sample sizes (our study: cases 105, controls 80 while Hussein et al.: cases 200, controls 150) or the age of patients (our study: 42.71 ± 12.07 years while Hussein et al.: 57.3 ± 3.9 years) or the disease duration (our study: 5.72 ± 4.35 years while Hussein et al.: 1.25 ± 0.78 years).
Conclusions
Direct associations between TNFB, BsmI, TaqI, MTHFR (C677T, A1298C), TGFβ1, and ApaI polymorphisms and RA susceptibility have been demonstrated in this study. In addition to the absence of a confirmed direct functional effect of FokI polymorphism, our results indicate that FokI have no role in RA susceptibility indirectly through the poor r2 values between FokI and all the VDR SNPs. Further studies with extended sample sizes (from the same population) are necessary to overcome the lack in power results and confirm our results in the Egyptian population. In addition, further investigations of other polymorphisms and its association with RA susceptibility may be helpful to clarify the pathogenesis of the disease.
Supporting Information
(TIF)
The table was generated using the Genetic Power Calculator program.
(TIF)
Data Availability
All relevant data are within the paper and its Supporting Information files.
Funding Statement
The authors received no specific funding for this work.
References
- 1. Saad MN, Mabrouk MS, Eldeib AM, Shaker OG. Identification of rheumatoid arthritis biomarkers based on single nucleotide polymorphisms and haplotype blocks: a systematic review and meta-analysis. J Adv Res. 2015. 10.1016/j.jare.2015.01.008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Lewis CM. Genetic association studies: design, analysis and interpretation. Brief Bioinform. 2002;3(2): 146–153. [DOI] [PubMed] [Google Scholar]
- 3. Lewis CM, Knight J. Introduction to genetic association studies. Cold Spring Harb Protoc. 2012;2012(3): 297–306. 10.1101/pdb.top068163 [DOI] [PubMed] [Google Scholar]
- 4. The basics of genetic association analysis In: Laird NM, Lange C, editors. The fundamentals of modern statistical genetics: Springer; 2010. pp. 99–108. [Google Scholar]
- 5. Rubini M, Padovan M, Baricordi O, Fotinidi M, Govoni M, Trotta F. The c.1298A>C polymorphism in the methylenetetrahydrofolate reductase gene is associated with rheumatoid arthritis susceptibility in Italian patients. Clin Exp Rheumatol. 2008;26(1): 163 [PubMed] [Google Scholar]
- 6. Palomino-Morales R, Gonzalez-Juanatey C, Vazquez-Rodriguez TR, Rodriguez L, Miranda-Filloy JA, Fernandez-Gutierrez B, et al. A1298C polymorphism in the MTHFR gene predisposes to cardiovascular risk in rheumatoid arthritis. Arthritis Res Ther. 2010;12(2): R71 10.1186/ar2989 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Hughes LB, Beasley TM, Patel H, Tiwari HK, Morgan SL, Baggott JE, et al. Racial or ethnic differences in allele frequencies of single-nucleotide polymorphisms in the methylenetetrahydrofolate reductase gene and their influence on response to methotrexate in rheumatoid arthritis. Ann Rheum Dis. 2006;65(9): 1213–1218. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Brambila-Tapia AJ, Duran-Gonzalez J, Sandoval-Ramirez L, Mena JP, Salazar-Paramo M, Gamez-Nava JI, et al. MTHFR C677T, MTHFR A1298C, and OPG A163G polymorphisms in Mexican patients with rheumatoid arthritis and osteoporosis. Dis Markers. 2012;32(2): 109–114. 10.3233/DMA-2011-0868 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Lee YH, Song GG. Associations between the C677T and A1298C polymorphisms of MTHFR and the efficacy and toxicity of methotrexate in rheumatoid arthritis: a meta-analysis. Clin Drug Investig. 2010;30(2): 101–108. 10.2165/11531070-000000000-00000 [DOI] [PubMed] [Google Scholar]
- 10. Kobayashi T, Murasawa A, Ito S, Yamamoto K, Komatsu Y, Abe A, et al. Cytokine gene polymorphisms associated with rheumatoid arthritis and periodontitis in Japanese adults. J Periodontol. 2009;80(5): 792–799. 10.1902/jop.2009.080573 [DOI] [PubMed] [Google Scholar]
- 11. Menegatti E, Davit A, Francica S, Berardi D, Rossi D, Baldovino S, et al. Genetic factors associated with rheumatoid arthritis and systemic vasculitis: evaluation of a panel of polymorphisms. Dis Markers. 2009;27(5): 217–223. 10.3233/DMA-2009-0666 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Munoz-Valle JF, Torres-Carrillo NM, Guzman-Guzman IP, Torres-Carrillo N, Ruiz-Quezada SL, Palafox-Sanchez CA, et al. The functional class evaluated in rheumatoid arthritis is associated with soluble TGF-beta1 serum levels but not with G915C (Arg25Pro) TGF-beta1 polymorphism. Rheumatol Int. 2012;32(2): 367–372. 10.1007/s00296-010-1624-x [DOI] [PubMed] [Google Scholar]
- 13. Al-Rayes H, Al-Swailem R, Albelawi M, Arfin M, Al-Asmari A, Tariq M. TNF-alpha and TNF-beta gene polymorphism in Saudi rheumatoid arthritis patients. Clin Med Insights Arthritis Musculoskelet Disord. 2011;4: 55–63. 10.4137/CMAMD.S6941 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Steiner G, Tohidast-Akrad M, Witzmann G, Vesely M, Studnicka-Benke A, Gal A, et al. Cytokine production by synovial T cells in rheumatoid arthritis. Rheumatology (Oxford). 1999;38(3): 202–213. [DOI] [PubMed] [Google Scholar]
- 15. Li N, Liu R, Zhai H, Li L, Yin Y, Zhang J, et al. Polymorphisms of the LTA gene may contribute to the risk of myocardial infarction: a meta-analysis. PLoS One. 2014;9(3): e92272 10.1371/journal.pone.0092272 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Saad MN, Mabrouk MS, Eldeib AM, Shaker OG. Vitamin D receptor gene polymorphisms in rheumatoid arthritis patients associating osteoporosis. 7th Cairo International Biomedical Engineering Conference; Cairo, Egypt: IEEE; 2014. pp. 75–78.
- 17. Uitterlinden AG, Fang Y, van Meurs JB, van Leeuwen H, Pols HA. Vitamin D receptor gene polymorphisms in relation to vitamin D related disease states. J Steroid Biochem Mol Biol. 2004;89–90(1–5): 187–193. [DOI] [PubMed] [Google Scholar]
- 18. Valdivielso JM, Fernandez E. Vitamin D receptor polymorphisms and diseases. Clin Chim Acta. 2006;371(1–2): 1–12. [DOI] [PubMed] [Google Scholar]
- 19. Arnett FC, Edworthy SM, Bloch DA, McShane DJ, Fries JF, Cooper NS, et al. The American Rheumatism Association 1987 revised criteria for the classification of rheumatoid arthritis. Arthritis Rheum. 1988;31(3): 315–324. [DOI] [PubMed] [Google Scholar]
- 20. Mostowska A, Lianeri M, Wudarski M, Olesinska M, Jagodzinski PP. Vitamin D receptor gene BsmI, FokI, ApaI and TaqI polymorphisms and the risk of systemic lupus erythematosus. Mol Biol Rep. 2013;40(2): 803–810. 10.1007/s11033-012-2118-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Barrett JC, Fry B, Maller J, Daly MJ. Haploview: analysis and visualization of LD and haplotype maps. Bioinformatics. 2005;21(2): 263–265. [DOI] [PubMed] [Google Scholar]
- 22. Yoo J, Seo B, Kim Y. SNPAnalyzer: a web-based integrated workbench for single-nucleotide polymorphism analysis. Nucleic Acids Res. 2005;33(Web Server issue): W483–488. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Yoo J, Lee Y, Kim Y, Rha SY, Kim Y. SNPAnalyzer 2.0: a web-based integrated workbench for linkage disequilibrium analysis and association analysis. BMC Bioinformatics. 2008;9: 290 10.1186/1471-2105-9-290 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Purcell S, Cherny SS, Sham PC. Genetic Power Calculator: design of linkage and association genetic mapping studies of complex traits. Bioinformatics. 2003;19(1): 149–150. [DOI] [PubMed] [Google Scholar]
- 25. McInnes IB, Schett G. The pathogenesis of rheumatoid arthritis. N Engl J Med. 2011;365(23): 2205–2219. 10.1056/NEJMra1004965 [DOI] [PubMed] [Google Scholar]
- 26. Chatzikyriakidou A, Voulgari PV, Lambropoulos A, Drosos AA. Genetics in rheumatoid arthritis beyond HLA genes: what meta-analyses have shown? Semin Arthritis Rheum. 2013;43(1): 29–38. 10.1016/j.semarthrit.2012.12.003 [DOI] [PubMed] [Google Scholar]
- 27. Saad MN, Mabrouk MS, Eldeib AM, Shaker OG. Effect of MTHFR, TGFβ1, and TNFB polymorphisms on osteoporosis in rheumatoid arthritis patients. Gene. 2015. 10.1016/j.gene.2015.05.037 [DOI] [PubMed] [Google Scholar]
- 28. Manni F, Leonardi P, Barakat A, Rouba H, Heyer E, Klintschar M, et al. Y-chromosome analysis in Egypt suggests a genetic regional continuity in Northeastern Africa. Hum Biol. 2002;74(5): 645–658. [DOI] [PubMed] [Google Scholar]
- 29. Berkun Y, Levartovsky D, Rubinow A, Orbach H, Aamar S, Grenader T, et al. Methotrexate related adverse effects in patients with rheumatoid arthritis are associated with the A1298C polymorphism of the MTHFR gene. Ann Rheum Dis. 2004;63(10): 1227–1231. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Inanir A, Yigit S, Tekcan A, Tural S, Kismali G. IL-4 and MTHFR gene polymorphism in rheumatoid arthritis and their effects. Immunol Lett. 2013;152(2): 104–108. 10.1016/j.imlet.2013.05.004 [DOI] [PubMed] [Google Scholar]
- 31. Sugiura Y, Niimi T, Sato S, Yoshinouchi T, Banno S, Naniwa T, et al. Transforming growth factor β1 gene polymorphism in rheumatoid arthritis. Ann Rheum Dis. 2002;61(9): 826–828. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Hussein YM, Mohamed RH, El-Shahawy EE, Alzahrani SS. Interaction between TGF-beta1 (869C/T) polymorphism and biochemical risk factor for prediction of disease progression in rheumatoid arthritis. Gene. 2014;536(2): 393–397. 10.1016/j.gene.2013.11.042 [DOI] [PubMed] [Google Scholar]
- 33. Zhu Y, Liu F, Zhang F, Yao DF, Chen XD. Correlation of transforming growth factor β1 gene T869C polymorphism and rheumatoid arthritis in Chinese population. Jiangsu Med J. 2006;12: 1112–1114. [Google Scholar]
- 34. Wang YP, Xiao LS, Zhou Y, Fei Y, Li JP. Gene polymorphisms of transforming growth factor-β1 with rheumatoid arthritis. Chin J Public Health. 2007;11: 1405–1406. [Google Scholar]
- 35. Alayli G, Kara N, Tander B, Canturk F, Gunes S, Bagci H. Association of transforming growth factor beta1 gene polymorphism with rheumatoid arthritis in a Turkish population. Joint Bone Spine. 2009;76(1): 20–23. 10.1016/j.jbspin.2008.02.012 [DOI] [PubMed] [Google Scholar]
- 36. Pokorny V, Chau J, Wu L, Yeoman S, Black P, McQueen F, et al. Transforming growth factor beta 1 gene (HSTGFB1) nucleotide T869C (codon 10) polymorphism is not associated with prevalence or severity of rheumatoid arthritis in a Caucasian population. Ann Rheum Dis. 2003;62(9): 907–908. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Panoulas VF, Douglas KM, Smith JP, Stavropoulos-Kalinoglou A, Metsios GS, Nightingale P, et al. Transforming growth factor-beta1 869T/C, but not interleukin-6 -174G/C, polymorphism associates with hypertension in rheumatoid arthritis. Rheumatology (Oxford). 2009;48(2): 113–118. [DOI] [PubMed] [Google Scholar]
- 38. Kim SY, Han SW, Kim GW, Lee JM, Kang YM. TGF-beta1 polymorphism determines the progression of joint damage in rheumatoid arthritis. Scand J Rheumatol. 2004;33(6): 389–394. [DOI] [PubMed] [Google Scholar]
- 39. Chang W-W, Su H, He L, Zhao K-F, Wu J-L, Xu Z-W. Association between transforming growth factor-β1 T869C polymorphism and rheumatoid arthritis: a meta-analysis. Rheumatology (Oxford). 2010;49(4): 652–656. [DOI] [PubMed] [Google Scholar]
- 40. Zhang L, Yan JW, Wang YX, Wan YN, Li JP, Liu P, et al. Association of TGF-beta1 +869C/T promoter polymorphism with susceptibility to autoimmune diseases: a meta-analysis. Mol Biol Rep. 2013;40(8): 4811–4817. 10.1007/s11033-013-2577-4 [DOI] [PubMed] [Google Scholar]
- 41. Zhou TB, Zhao HL, Fang SL, Drummen GP. Association of transforming growth factor-beta1 T869C, G915C, and C509T gene polymorphisms with rheumatoid arthritis risk. J Recept Signal Transduct Res. 2014;34(6): 469–475. 10.3109/10799893.2014.919594 [DOI] [PubMed] [Google Scholar]
- 42. Vandevyver C, Raus P, Stinissen P, Philippaerts L, Cassiman JJ, Raus J. Polymorphism of the tumour necrosis factor beta gene in multiple sclerosis and rheumatoid arthritis. Eur J Immunogenet. 1994;21(5): 377–382. [DOI] [PubMed] [Google Scholar]
- 43. Zake LN, Cimdina I, Rumba I, Dabadghao P, Sanjeevi CB. Major histocompatibility complex class I chain related (MIC) A gene, TNFa microsatellite alleles and TNFB alleles in juvenile idiopathic arthritis patients from Latvia. Hum Immunol. 2002;63(5): 418–423. [DOI] [PubMed] [Google Scholar]
- 44. Vinasco J, Beraun Y, Nieto A, Fraile A, Mataran L, Pareja E, et al. Polymorphism at the TNF loci in rheumatoid arthritis. Tissue Antigens. 1997;49(1): 74–78. [DOI] [PubMed] [Google Scholar]
- 45. Takeuchi F, Nabeta H, Hong GH, Kawasugi K, Mori M, Matsuta K, et al. The genetic contribution of the TNFa11 microsatellite allele and the TNFb + 252*2 allele in Japanese RA. Clin Exp Rheumatol. 2005;23(4): 494–498. [PubMed] [Google Scholar]
- 46. Panoulas VF, Nikas SN, Smith JP, Douglas KM, Nightingale P, Milionis HJ, et al. Lymphotoxin 252A>G polymorphism is common and associates with myocardial infarction in patients with rheumatoid arthritis. Ann Rheum Dis. 2008;67(11): 1550–1556. 10.1136/ard.2007.082594 [DOI] [PubMed] [Google Scholar]
- 47. Santos MJ, Fernandes D, Caetano-Lopes J, Perpetuo IP, Vidal B, Canhao H, et al. Lymphotoxin-alpha 252 A>G polymorphism: a link between disease susceptibility and dyslipidemia in rheumatoid arthritis? J Rheumatol. 2011;38(7): 1244–1249. 10.3899/jrheum.101170 [DOI] [PubMed] [Google Scholar]
- 48. Karray EF, Bendhifallah I, Benabdelghani K, Hamzaoui K, Zakraoui L. Tumor necrosis factor gene polymorphisms and susceptibility to rheumatoid arthritis in regional Tunisian population. J Infect Dis Immun. 2011;3(2): 30–35. [Google Scholar]
- 49. Farouk HM, Mansour HE, Rahman SA, Mostafa AA, Shamy HA, Zarouk WA. Effect of the human leukocyte antigen HLA-DRB1 and anti-cyclic citrullinated peptide on the outcome of rheumatoid arthritis patients. Braz J Med Biol Res. 2009;42(9): 831–838. [DOI] [PubMed] [Google Scholar]
- 50. Newton JL, Harney SM, Wordsworth BP, Brown MA. A review of the MHC genetics of rheumatoid arthritis. Genes Immun. 2004;5(3): 151–157. [DOI] [PubMed] [Google Scholar]
- 51. Garcia-Lozano JR, Gonzalez-Escribano MF, Valenzuela A, Garcia A, Nunez-Roldan A. Association of vitamin D receptor genotypes with early onset rheumatoid arthritis. Eur J Immunogenet. 2001;28(1): 89–93. [DOI] [PubMed] [Google Scholar]
- 52. Lee CK, Hong JS, Cho YS, Yoo B, Kim GS, Moon HB. Lack of relationship between vitamin D receptor polymorphism and bone erosion in rheumatoid arthritis. J Korean Med Sci. 2001;16(2): 188–192. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53. Maalej A, Petit-Teixeira E, Michou L, Rebai A, Cornelis F, Ayadi H. Association study of VDR gene with rheumatoid arthritis in the French population. Genes Immun. 2005;6(8): 707–711. [DOI] [PubMed] [Google Scholar]
- 54. Rass P, Pakozdi A, Lakatos P, Zilahi E, Sipka S, Szegedi G, et al. Vitamin D receptor gene polymorphism in rheumatoid arthritis and associated osteoporosis. Rheumatol Int. 2006;26(11): 964–971. [DOI] [PubMed] [Google Scholar]
- 55. Ates Ö, Dolek B, Dalyan L, Topal-Sarikaya A. Vitamin D receptor gene polymorphisms in rheumatoid arthritis. Turk J Rheumatol. 2011;26(2): 145–149. [Google Scholar]
- 56. Karray EF, Ben Dhifallah I, Ben Abdelghani K, Ben Ghorbel I, Khanfir M, Houman H, et al. Associations of vitamin D receptor gene polymorphisms FokI and BsmI with susceptibility to rheumatoid arthritis and Behcet's disease in Tunisians. Joint Bone Spine. 2012;79(2): 144–148. 10.1016/j.jbspin.2011.06.003 [DOI] [PubMed] [Google Scholar]
- 57. Mosaad YM, Hammad EM, Fawzy Z, Abdal Aal IA, Youssef HM, ElSaid TO, et al. Vitamin D receptor gene polymorphism as possible risk factor in rheumatoid arthritis and rheumatoid related osteoporosis. Hum Immunol. 2014;75(5): 452–461. 10.1016/j.humimm.2014.02.009 [DOI] [PubMed] [Google Scholar]
- 58. Hussien YM, Shehata A, Karam RA, Alzahrani SS, Magdy H, El-Shafey AM. Polymorphism in vitamin D receptor and osteoprotegerin genes in Egyptian rheumatoid arthritis patients with and without osteoporosis. Mol Biol Rep. 2013;40(5): 3675–3680. 10.1007/s11033-012-2443-9 [DOI] [PubMed] [Google Scholar]
- 59. Goertz B, Fassbender WJ, Williams JC, Marzeion AM, Bretzel RG, Stracke H, et al. Vitamin D receptor genotypes are not associated with rheumatoid arthritis or biochemical parameters of bone turnover in German RA patients. Clin Exp Rheumatol. 2003;21(3): 333–339. [PubMed] [Google Scholar]
- 60. Tizaoui K, Kaabachi W, Ouled Salah M, Ben Amor A, Hamzaoui A, Hamzaoui K. Vitamin D receptor TaqI and ApaI polymorphisms: a comparative study in patients with behcet's disease and rheumatoid arthritis in Tunisian population. Cell Immunol. 2014;290(1): 66–71. 10.1016/j.cellimm.2014.05.002 [DOI] [PubMed] [Google Scholar]
- 61. Shukla S, Tripathi AK, Tripathi JK, Indurkar M, Chauhan UK. Role of PTPN22 and VDR gene polymorphisms in susceptibility to rheumatoid arthritis: a study from central India. Adv Genomics Genet. 2014;4: 79–85. [Google Scholar]
- 62. Hitchon CA, Sun Y, Robinson DB, Peschken CA, Bernstein CN, Siminovitch KA, et al. Vitamin D receptor polymorphism rs2228570 (Fok1) is associated with rheumatoid arthritis in North American natives. J Rheumatol. 2012;39(9): 1792–1797. 10.3899/jrheum.120387 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
(TIF)
The table was generated using the Genetic Power Calculator program.
(TIF)
Data Availability Statement
All relevant data are within the paper and its Supporting Information files.