

Abstract
Adapting to exploit new food sources may be essential, particularly in winter, when the impact of food limitation on survival of individuals is critical. One of the most important additional sources of food for birds in human settlements is birdfeeders. At a large spatial scale, we experimentally provided birdfeeders with four different kinds of food to analyze exploitation and use of a novel food supply provided by humans. Nine species started foraging at the new birdfeeders. The species that exploited the new feeders the fastest was the great tit. Use of novel food sources was faster in urban habitats and the presence of other feeders reduced the time until a new feeder was located. Urbanization may be associated with behavioural skills, technical innovations and neophilia resulting in faster discovery of new food sources. This process is accelerated by the experience of feeder use in the vicinity, with a strong modifying effect of the number of domestic cats.
Food availability is an important environmental cue when animals decide how much to invest in maintenance and reproduction, ultimately affecting population size1,2. In seasonal environments, winter is a critical period for survival for many bird species3,4,5,6. Therefore, adaptations that allow discovery and use of ephemeral food should be favoured by natural selection7,8. Thus the ability to develop novel behaviour innovations should facilitate exploitation of novel food resources and hence increase the prospects of survival in the new environment9,10. Birdfeeders have increased in numbers in recent decades, and now constitute a massive food source in many parts of the temperate zone. They are directly associated with human settlements across a range of surrounding habitats varying from rural to highly urbanized2,9,11. In fact, many bird species are able to exploit this novel food source in urban and suburban environments, where they thrive in close proximity of humans12,13,14.
For economic, social and biodiversity reasons people often change the presence and the abundance of artificial feeders during winter15. In winter when the time budget of especially small birds is very tight, opportunities to find a rich new food source can provide a large advantage in terms of intra- and interspecific competition and hence survival prospects15,16. Therefore, exploratory behaviour should be an adaptive trait especially in situations of limiting resources and starvation8. On the other hand, dispersal skills of small passerines, both at a local and a regional scale, and the search for novel food is reduced by both intra- and interspecific competition17. However, predation is a stronger selective agent than competition affecting winter mortality of small birds18,19,20,21. The effects of presence of predators or cues left by predators may modify the use of foraging and roosting sites in winter22,23,24.
In summary, discovery of novel food resources may be important for survival, and as a consequence it will promote individual traits linked with personality, enabling use of novel food sources8. The ability to find and explore novel food sources provided by humans would be important for adaptation to novel habitats, and for understanding urbanization processes of animals9,25.
To the best of our knowledge, there is a lack of tests of how fast wintering birds recognize novel food sources, especially those provided in birdfeeders. Such studies have general implications, because winter bird feeding is one of the most widespread direct interactions between humans and nature with important social and environmental consequences2,26,27. Exploitation of such novel feeding sites may allow tests of differences in feeder exploitation between rural and urban habitats2,28. We conducted a manipulative experiment with birdfeeder locations recording the species and the sex at the start of use of novel food sources29. New birdfeeders should be visited faster in urban than in rural habitats because population density of birds is higher and hence intraspecific competition for resources more intense. Moreover, discovery behaviour may be modified by risk of predation30. Finally, discovery behaviour may depend on prior experience and hence the number of birdfeeders provided by humans if prior exposure to feeders causes a phenotypically plastic reduction in neophobia.
Results
The wintering bird community around experimental birdfeeders contained 925 individuals belonging to 43 species (6.7, SD = 2.88; range 1–13). In 89 cases (64.5% of all 138 trials), a total of nine species visited experimental birdfeeders during 120 minutes and started to forage (Fig. 1). The proportion of species that discovered new food sources differed significantly from the species composition in the wintering bird community (Fig. 1; χ2 = 53.33, df = 8, P < 0.001). The bird species most frequently using the new food source was the great tit (64.0% of all cases). There were no significant differences between the frequencies of bird species first using the new food source in rural and urban environments (χ2 = 4.21, df = 3, P = 0.24). Male great tits were the most frequent users of new feeders (80%, χ2 = 4.14, df = 1, P = 0.04), despite an even sex ratio of this species in the study area (sex ratio: 92 females, 106 males; P = 0.39).
Figure 1. Abundance and frequency of first birds visiting a novel food source for different bird species.
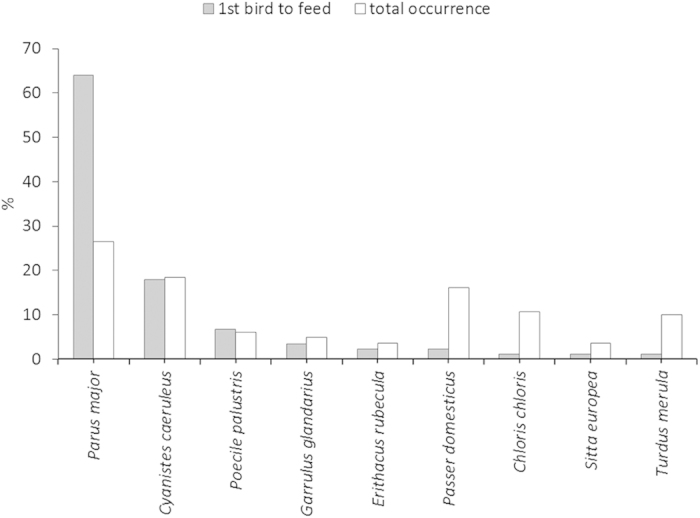
Sample sizes were 89 and 467 individuals, respectively, for the first bird and for the entire wintering bird community.
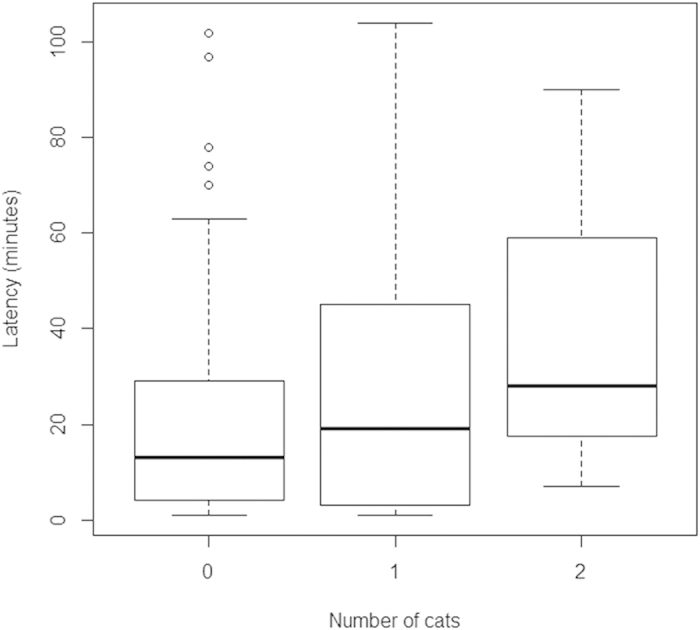
For all species in GAMM models, the urban environment was correlated with a decrease in time spent using the new source of food. The number of bird feeding stations and species richness of other birds that were seedeaters was positively correlated with the speed of use of the new food source, while the presence of cats was a cause of slower use of feeders (Table 1, Fig. 2).
Table 1. Differences in time when the first bird arrived at a new feeder in relation to environment, temperature, number of cats, dogs, humans, corvids, bird species richness and number of feeders. Results of GAMM: R-sq. (adj) = 0.295, scale est = 10.588, n = 89, df = 9. Random effects (SD): intercept = 0.513, residual = 3.254.
| Variable | Estimate | SE | t | P |
|---|---|---|---|---|
| Intercept | 4.277 | 0.368 | 11.614 | <0.001 |
| Environment (urban) | −0.856 | 0.231 | −3.699 | 0 .001 |
| Temperature | 0.004 | 0.019 | 0.234 | 0.816 |
| No. cats | 0.386 | 0.171 | 2.256 | 0.027 |
| No. dogs | 0.156 | 0.134 | 1.159 | 0.250 |
| No. human walkers | −0.001 | 0.046 | −0.022 | 0.983 |
| No. feeders | −0.189 | 0.063 | −3.011 | 0.003 |
| Bird species richness | −0.098 | 0.051 | −1.928 | 0.057 |
| No. corvids | 0.112 | 0.137 | 0.816 | 0.417 |
Figure 2. Differences in average time taken for birds to use a novel source of food (expressed in minutes) on y-axis, related to the presence and number of cats (x-axis) in the study area.
The total occurrence of different bird species at feeders was strongly positively correlated with the frequency of technical innovations (log-transformed data, F = 14.03, df = 1, 6, r2 = 0.70, P = 0.0096, estimate (SE) = 0.669 (0.178)) after adjusting for body mass (log-transformed: F = 5.19, df = 1, 6, r2 = 0.46, P = 0.063, estimate (SE) = −0.439 (0.193)). This suggests that there is a cognitive basis for exploitation of bird feeders linked to the frequency of technical innovations31.
Discussion
Faster detection and exploitation of novel food sources was recorded in urban habitat and increased by the number birdfeeders already present. Additionally, the presence of domestic cats negatively affected time required for discovery of new birdfeeders, suggesting that detection of novel food sources is affected by the risk of predation when searching for food. Finally, in the majority of cases the first species that discovered the novel food was the great tit, especially males, in a proportion larger than predicted from its abundance in the wintering bird community. The great tit is recognized across Europe as a species that often and regularly uses birdfeeders32, although varying among years and seasons, and even individually33. Therefore, great tits were used in many experiments on use of artificial food sources and the spread of social information34,35. Here we have shown that this species is not only a numerous visitor at feeders, but also the first to discover novel resources in the majority of cases. Male great tits generally take greater risks than females36, explaining why males were disproportionately frequent as the discoverers of novel food sources. Latency of some species is also potentially explained by the spectrum of diet, and therefore less attractiveness of novel food provided by humans11.We have shown that early exploitation of bird feeders is peculiar to specific species and even sexes. Indeed the frequency of occurrence at feeders was strongly predicted by a high frequency of technical innovations for a given body size. Because innovations by birds are more common in species with a relatively large brain size, we suggest that the abundance of birds frequenting novel feeders indirectly has an underlying neural basis8,31.
One the most interesting results were the differences between urban and rural habitats in time required to find novel food sources. Such differences can be the result of bird density and feeder density as already known food sources, but also skills of particular individual in exploratory behavior8. Food is not presented in exploration trials, so the motivation is assumed to be information gathering16,37. The costs of exploration may be the time, energy and attention diverted from other activities or risks, or indeed the potential unknown dangers of the novel object or the environment itself. We found empirical support for this suggestion because the latency until detection of novel food increased in the presence of a larger number of predators, domestic cats. The number of cats in human settlements is increasing, and they have a disproportionately large negative impact on biodiversity38 that accounts for a large fraction of mortality in wildlife including birds39. Urban habitats can support more risky and exploratory individuals than rural habitats9,13,24,40 because humans provide refuges against predators that generally avoid human proximity41 Species that readily taste new foods and/or develop novel foraging techniques are more likely to survive and to reproduce in a novel environment than individuals belonging to a more stereotyped species with less exploratory behavior10. We did not find a significant effect of temperature on discovery of birdfeeders as in other studies42, although the effect of weather is probably more important in the use of already known places than when searching for new sites15,16,43.
Two broad conclusions can be drawn from our field experiments. First, exploratory behaviour and search for novel food is influenced by multiple sources of variation such as rural vs. urban habitats, the presence of other birdfeeders provided earlier, and the presence of domestic cats. Second, there is significant intra- and interspecific variation in search for novel food sources, and part of this variation is due to sex, and an ability to innovate by particular bird species. This implies that a range of different factors contribute to recruitment of birds to novel sources of food and hence for the diversity of the winter bird community in urban environments.
Methods
Field methods
Data were collected during December 2013-February 2014 in eight cities and nearby rural areas across Poland (for more details on cities and map of location, see ). In total 138 experimental trials (80 and 58 in rural and urban areas, respectively) were carried out during days (1–4 hrs after sunrise) with favourable weather conditions (no snow or rain, strong wind). The site of the experiment was chosen randomly. To attract as many species as possible, in each trial one birdfeeder contained at the bottom four different trays (changed randomly for each trial) with four different kinds of food (contain carbohydrates and lipids): animal fat, dry fruits of rowanberry, sunflowers and millet seeds. Birdfeeders of a single model for all trials were used across the entire country, each being in the shape of a small house with a roof, and a 1.20 m pole that was dug into the ground (grass/soil) and was transported by car to the site of the experiment.
Before starting observations of activity at the birdfeeder, to quantify the composition of the local wintering bird community we recorded birds at three points at distances of 100 m from the feeder, located at virtual triangle tops with the birdfeeder in the middle, birds were counted using the point-counting method with 5-minute observations at each point. Data from point-counts were summarized and used to describe the winter bird community around feeders. Additionally at the start of the experiment temperature was also noted, and during the first 15 minutes of feeder watching we recorded the number of cats and dogs, as well as human walkers within a distance of 100 m (Table 2). At a radius of 100 m from experimental birdfeeders we also recorded all other feeders provided by humans because they potentially attract birds in the vicinity. Experimental birdfeeders were provided at a distance larger than 50 m from already existing (i.e. non-experimental) birdfeeders provided by humans.
Table 2. Characteristic of explanatory variables. Data are presented as means ± SE.
| Variable | Urban n = 58 | Rural n = 80 |
|---|---|---|
| No. cats | 0.38 ± 0.08 | 0.15 ± 0.04 |
| No. dogs | 0.81 ± 0.14 | 0.25 ± 0.06 |
| No. human walkers | 8.84 ± 2.41 | 0.84 ± 0.19 |
| No. feeders | 0.41 ± 0.08 | 2.03 ± 0.28 |
When the new experimental birdfeeder was provided at a specific site, it was observed for 120 minutes from a distance (e. g. from a parked car with good visibility) noting when and which bird species first started to explore food in the feeder, and latency time (minutes since the start of the experiment) of this first bird was noted. Immediately, at least after 10 min when the first bid used the experimental birdfeeder, or if no birds arrived at the birdfeeders following an experimental trial of 120 min, the experiment was terminated and the observer with the experimental feeder moved to another place, located at least 2 km from the previous one. All sampled sites, studied in the proximity of the eight Polish cities, were classified as ‘urban’ or ‘rural’ following the main landscape. During observations birds were sexed from a distance with binoculars, although sufficient data on sex was only collected for the great tit Parus major. Bird species richness was calculated as the sum of all seed-eating bird species recorded in each sampled site.
The methods were carried out in accordance with the approved guidelines and Polish national law. Moreover, because is not really experimental study using animals, but only observations from some distance, additional approval by the local ethical committee was not required.
Statistics
The sampled sites were treated as statistically independent observations because the values of spatial autocorrelation were low (Mantel test p > 0.05, n = 89)44,45. A Mantel test measures the correlation between two matrices typically containing measures of distance. The Mantel test was performed using the package ‘ade4’ for R package.
Occurrences of the most frequent bird species first using the new food source was analyzed in terms of differences among habitats (urban or rural) and difference in sex ratio of the great tit using new feeders with a chi-square test46,47, and the Bonferroni adjustment method was applied, multiplying the p-values with the number of comparisons. Differences in average time taken for birds to use a novel source of food (expressed in minutes) between male and female great tits were tested with the Mann-Whitney U-test.
Finally, a generalized additive mixed model (GAMM) with the package ‘mgcv’7,48 was used to study differences in latency when the first bird started to use the novel food source. GAMMs are especially designed for inference of relationships of clustered and correlated data by adding random effects to the additive predictor, which account for that correlation. In this study, GAMM was used to take into account any potential effect on the studied behaviour, related to differences among cities where data were collected.
The variable “minutes” was used as response variable. The covariates entered in the full model were bird species richness, date, temperature, environment (urban or rural), number of dogs, cats and human walkers, number of feeders and number of corvids. In this case, we specified as random effects the grouping structure of the data defined by the “city identity” where the observations were collected. A Poisson distribution error was assumed for the response variable.
The frequency of technical innovations was estimated as the sum of novel feeding techniques, novel techniques in an anthropogenic context, novel parasitic behaviour, novel commensal behaviour, novel mutualistic behaviour, novel prototool behaviour, novel tool behaviour and novel caching behaviour36.
Additional Information
How to cite this article: Tryjanowski, P. et al. Who started first? Bird species visiting novel birdfeeders. Sci. Rep. 5, 11858; doi: 10.1038/srep11858 (2015).
Acknowledgments
We thank E. Mróz, J.J. Nowakowski and P. Czechowski for help in the field.
Footnotes
Author Contributions idea – P.T., A.P.M.; data analysis: P.T., F.M., P.S., A.P.M.; data collection: P.T., A.G., P.I., C.M., P.Z., D.W.; writing the manuscript: all authors.
References
- Newton I. Population limitation in birds (Academic Press, London, 1998). [Google Scholar]
- Robb G. N., McDonald R. A., Chamberlain D. E. & Bearhop S. Food for thought: Supplementary feeding as a driver of ecological change in avian populations. Front. Ecol. Environ. 6, 476–484 (2008). [Google Scholar]
- Houston A. I. & McNamara J. M. A theoretical investigation of the fat reserves and mortality levels of small birds in winter. Ornis Scand. 24, 205–219 (1993). [Google Scholar]
- Bednekoff P. A. & Krebs J. R. Great tit fat reserves, effects of changing and unpredictable feeding day length. Funct. Ecol. 9, 457–462 (1995). [Google Scholar]
- Pravosudov V. V. & Grubb T. C. Jr. Energy management in passerine birds during the nonbreeding season: A review. Curr. Ornithol. 14, 189–234 (1997). [Google Scholar]
- Atkinson P. W., Fuller R. A. & Vickery J. A. Large-scale patterns of summer and winter bird distribution in relation to farmland type in England and Wales. Ecography 25, 466–480 (2002). [Google Scholar]
- Gosling S. D. From mice to men, what can we learn about personality from animal research. Psychol. Bull. 127, 45–86 (2000). [DOI] [PubMed] [Google Scholar]
- Ducatez S., Clavel J. & Lefebvre L. Ecological generalism and behavioral innovation in birds. Technical intelligence or the simple incorporation of new foods? J. Anim. Ecol. 84, 79–89 (2015). [DOI] [PubMed] [Google Scholar]
- Møller A. P. Successful city dwellers, a comparative study of the ecological characteristics of urban birds in the Western Palearctic. Oecologia 159, 849–858 (2009). [DOI] [PubMed] [Google Scholar]
- Sol D., Griffin A. S., Bartomeus I. & Boyce H. Exploring or Avoiding Novel Food Resources? The Novelty Conflict in an Invasive Bird. PLoS ONE 6, e195352011 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Galbraith J. A. et al. Risks and drivers of wild bird feeding in urban areas of New Zealand. Biol. Conserv. 180, 64–74 (2014). [Google Scholar]
- Luniak M. Synurbization – adaptation of animal wildlife to urban development. Pp. 50–55. In Shaw W. W., Harris L. K. & Vandruff L. (Eds.), Proceedings of the 4th International Urban Wildlife Symposium. University of Arizona, Tucson, Arizona, U.S.A. (2004). [Google Scholar]
- Ditchkoff S. S., Saalfeld S. T. & Gibson C. J. Animal behavior in urban ecosystems: Modifications due to human-induced stress. Urban Ecosyst. 9, 5–12 (2006). [Google Scholar]
- Fuller R. A., Irvine, K. N., Davies, Z. G., Armsworth, P. R. & Gaston K. J. Interactions between people and birds in urban landscapes. Stud. Avian Biol. 45, 249–266 (2012). [Google Scholar]
- Maciusik B., Lenda M. & Skórka P. Corridors, local food resources and climatic conditions affect the utilization of the urban environment by the Black-headed Gull Larus ridibundus in winter. Ecol. Res. 25, 263–272 (2010). [Google Scholar]
- Tvardikova K. & Fuchs R. Tits recognize the potential dangers of predators and harmless birds in feeder experiments. J. Ethol. 30, 157–165 (2012). [Google Scholar]
- Dhondt A. A. & Eyckerman R. Competition between the great tit and the blue tit outside the breeding season in field experiments. Ecology 61, 1291–1296 (1980). [Google Scholar]
- Ekman J. Tree use and predator vulnerability of wintering passerines. Ornis Scand. 17, 261–267 (1986). [Google Scholar]
- Carrascall M. & Moreno E. Proximal costs and benefits of heterospecific social foraging in the Great Tit, Parus major. Can. J. Zool. 70, 1947–1952 (1992). [Google Scholar]
- Suhonen J. Predation risk influences the use of foraging sites by tits. Ecology 74, 1197–1203 (1993). [Google Scholar]
- Kullberg C. Spatial niche dynamics under predation risk in the willow tit Parus montanus. J. Avian Biol. 29, 235–240 (1998). [Google Scholar]
- Lima S. L. Nonlethal effects in the ecology of predator-prey interaction. What are the ecological effects of anti-predator decision-making? BioScience 48, 25–34 (1998). [Google Scholar]
- Ekner A. & Tryjanowski P. Do small hole nesting passerines detect cues left by a predator? A test on winter roosting sites. Acta Ornithol. 43, 107–111 (2008). [Google Scholar]
- Seress G., Bókony V., Heszberger J. & Liker A. Response to predation risk in urban and rural house sparrows. Ethology 117, 896–907 (2011). [Google Scholar]
- Jokimäki J., Suhonen J., Jokimäki-Kaisanlahti M. L. & Carbó-Ramírez P. Effects of urbanization on breeding birds in European towns, Impacts of species traits. Urban Ecosyst. 10.1007/s11252-014-0423-7 (2014). [DOI] [Google Scholar]
- Tratalos J., Fuller R. A., Warren P. H., Davies R. G. & Gaston K. J. Urban form, biodiversity potential and ecosystem services. Landscape Urban Plan. 83, 308–317 (2007). [Google Scholar]
- Horn D. J. & Johansen S. M. A. comparison of birdfeeding practices in the United States and Canada. Wildlife Soc. Bull. 37, 293–300 (2013). [Google Scholar]
- Horn D. J., Fairbairn S. E. & Hollis R. J. Factors influencing the occurrence of birds that use feeders in Iowa. J. Iowa Acad. Sci. 109, 8–18 (2002). [Google Scholar]
- Amrhein V. Wild bird feeding (probably) affects avian urban ecology. In: Gil D. & Brumm H. Avian Urban Ecology, Behavioural and Physiological Adaptations (Oxford University Press, 2014). [Google Scholar]
- Lima S. L. Predators and the breeding bird: behavioral and reproductive flexibility under the risk of predation. Biol. Rev. 84, 485–513 (2009). [DOI] [PubMed] [Google Scholar]
- Overington S. E., Bogert N. J., Morand-Ferron J. & Lefebvre L. Technical innovation drive the relationship between innovativeness and residual brain size in birds. Anim. Behav. 78, 1001–1010 (2009). [Google Scholar]
- Chamberlain D. E. et al. Annual and seasonal trends in the use of garden feeders by birds in winter. Ibis 147, 563–575 (2005). [Google Scholar]
- Dingemanse N. J. et al. Variation in personality and behavioural plasticity across four populations of the great tit Parus major. J. Anim. Ecol. 81, 116–126 (2012). [DOI] [PubMed] [Google Scholar]
- Suhonen J. Predation risk influences the use of foraging sites by tits. Ecology 74, 1197–1203 (1993). [Google Scholar]
- Farine D. R. & Lang S. D. The early bird gets the worm, foraging strategies of wild songbirds lead to the early discovery of food sources. Biol. Lett. 9, 10.1098/rsbl.2013.0578 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Regelmann K. & Curio E. Why do great tit (Parus major) males defend their brood more than females do? Anim. Behav. 34, 1206–1214 (1986). [Google Scholar]
- Mettke-Hofmann C., Winkler H. & Leisler B. The significance of ecological factors for exploration and neophobia in parrots. Ethology 108, 249–272 (2002). [Google Scholar]
- Baker P. J. et al. Cats about town: is predation by free‐ranging pet cats Felis catus likely to affect urban bird populations? Ibis 150, 86–99 (2008). [Google Scholar]
- Loss S. R., Will T. & Marra P. P. The impact of free-ranging domestic cats on wildlife of the United States. Nature Commun. 4, 1396. 10.1038/ncomms2380 (2013). [DOI] [PubMed] [Google Scholar]
- Maklakov A. A. et al. Brains and the city, big-brained passerine birds succeed in urban environments. Biol. Lett. 7, 730. 10.1098/rsbl.2011.0341 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Møller A. P. Urban areas as refuges from predators and flight distance of prey. Behav. Ecol. 23, 1030–1035 (2012). [Google Scholar]
- Wilson W. E. Jr. The effects of supplemental feeding on wintering Black-capped Chickadees (Poecile atricapilla) in central Maine, Population and individual responses. Wilson Bull. 113, 65–72 (2001). [Google Scholar]
- McNamara J. R., Houston A. & Lima S. Foraging routines of small birds in winter, A theoretical investigation. J. Avian Biol. 25, 287–302 (1994). [Google Scholar]
- Tryjanowski P. et al. Winter bird assemblages in rural and urban habitats, a national survey. PLoS One, resubmitted (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Manly B. F. J. Randomization, Bootstrap and Monte Carlo Methods in Biology. Third Edition (Chapman and Hall/CRC, Boca Raton, 2006). [Google Scholar]
- Sokal R. R. & Rohlf F. J. Biometry, the principles and practice of statistics in biological research (W.H. Freeman, New York, NY, 1995). [Google Scholar]
- Wood S. N. Generalized Additive Models. An Introduction with R (London, UK, Chapman and Hall/CRC, Boca Raton, 2006).
- Wood S. N. Stable and efficient multiple smoothing parameter estimation for generalized additive models. J. Am. Stat. Assoc. 99, 673–686 (2004). [Google Scholar]