

Abstract
Background
Fucoidan is a high-molecular polysaccharide whose main constituent is sulfated fucose. We specifically focused on the anti-proliferation activity of fucoidan and examined the underlying mechanism in MKN45 gastric cancer cells.
Methods
MKN45 cell proliferation was analyzed by BrdU assay and fucoidan cytotoxicity was examined by LDH and clonogenic assays. The Agilent Human microarray kit was used to identify upregulated and downregulated genes in response to fucoidan, and western blot analyses evaluated cell cycle proteins.
Results
Fucoidan impeded the MKN45 cell cycle by approximately 50%, and inhibited cell proliferation.LDH assays showed no immediate cytotoxic effects of fucoidan at 24 h exposure, however longer time courses revealed cell growth inhibition at 4 days in a dose-dependent manner. Microarray analysis identified MAP3K5, or ASK1 (apoptosis signal-regulating kinase),which was upregulated by 1.38-fold upon fucoidan treatment.Fucoidan increased ASK1 protein levels, while reducing phosphorylated ASK1 levels. Reduction of ASK1 by siRNA decreased proliferation of MKN45 cells.
Conclusion
Our findings show that fucoidan may suppress cellular proliferation and DNA synthesis in MKN45 cells by suppressing the ASK1-p38 signaling pathway through reduction of phosphorylated ASK1 levels.
Keywords: anti-proliferation activity;, ASK1;, cell cycle;, fucoidan;, MKN45
Fucoidan is a high-molecular polysaccharide only found in brown algae and its main constituent is sulfated fu-cose. The saccharide chain also includes galactose, mannose, xylose and uronic acid. Over the past decades, fucoidan has been extensively studied, owing to its numerous interesting biological activities, such as antioxidant,1 immunomodulatory,2 antiviral,3 antithrombotic4 and anticoagulant effects.5 In particular, we have focused on the anti-cancer activities of fucoidan.
Multiple studies have demonstrated antitumor activity for many polysaccharides. Fucoidans from Eisenia bicyclis and Laminaria japonica are effective against sarcoma 180. 6, 7 Fucoidan from L. japonica can inhibit hepatoma QGY7703 cell growth in logarithmic phase in vitro, thus restraining the growth of tumors.8 Fucoidans from L. saccharina, L. digitata, Fucus serratus, F. distichus and F. vesiculosus strongly blocked MDA-MB-231 breast carcinoma cell adhesion to platelets, an effect that might have critical implications in tumor metastasis.9
Among its various anti-cancer effects, we have focused on the antiproliferation activities of fucoidan. We hypothesized that fucoidan exerts its anti-cancer effects through alterations of gene expression. Thus, in this study, we performed mRNA microarray analysis to examine the effects of fucoidan on cellular gene expression. We selected MKN45 gastric cancer cells for analysis, as this common cancer cell line is relatively easy to culture and is often used for experimental analyses.10– 12
MATERIALS AND METHODS
Reagents
The human gastric adenocarcinoma cell line, MKN45 was obtained from the Riken BioResource Center (National Bio-Resource Project of the Ministry of Education, Culture, Sports, Science and Technology, Tsukuba, Japan) in November 20, 2012. Cells were maintained in RPMI 1640 supplemented with 10% fetal bovine serum (FBS). MKN45 gastric adenocarcinoma cells were obtained from a metastatic liver tumor in a 62-year-old Japanese female.
Dulbecco’s modified Eagle medium (DMEM), trypsin-EDTA, and the lactate dehydrogenase (LDH) assay kit were obtained from Wako (Tokyo, Japan). FBS was from Biowest (Nuaillé, France). Rabbit polyclonal anti-ASK1 (H-300), anti-ERK1 (K-23), mouse monoclonal anti-phospho-ERK (E-4), and ASK1 siRNA were purchased from Santa Cruz Biotech (Santa Cruz, CA). Rabbit polyclonal anti-phospho-ASK1 (Thr845), rabbit monoclonal anti-p38 MAPK (D13E1) and antiphospho-p38 MAPK (Thr180/Tyr182) (D3F9) were from Cell Signaling Technology (Danvers, MA). Horseradish peroxidase-conjugated anti-rabbit immunoglobulin (IgG), anti-mouse IgG, and the ECL plus detection kit were from GE Healthcare (Buckinghamshire, United Kingdom). Alexa Fluor 488-conjugated anti-rabbit IgG was from Invitrogen (Carlsbad, CA).
Preparation of fucoidan
High molecular weight fucoidan was obtained from Marine Products Kimuraya (Sakaiminato, Japan) as a sulfated polysaccharide, extracted from the brown seaweed Cladosiphon okamuranus (Okinawa Mozuku) as previously described.13, 14 The mean molecular weight was 4 × 105 and the active ingredient contained 14.2% sulfate (SO42−). The material has been previously used in studies on biological activity and with respect to safety of human use.15– 17
Cell culture, cell growth and cell death assay
Cells were cultured in DMEM with 10% FBS. For fucoidan treatment, culture medium was replaced with fresh medium mixed with indicated concentrations of fucoidan, and cells were cultured until the assay. Cell growth assay was performed by counting cell numbers using a hemocytometer. Cytotoxic effects were evaluated by the LDH assay following the manufacturer’s protocol. Briefly, extracellular LDH activity in the medium was evaluated by an enzymatic reaction that results in formazan products, which were measured with a spectrophotometer (Tecan Sunrise Rainbow, Tecan Japan, Tokyo) at 600 nm
siRNA transfection
Transfection of siRNA was performed with Lipofectamine RNAiMAX (Invitrogen) following the manufacturer’s protocol. Briefly, ASK1 siRNA (sc-20748; Santa Cruz Biotech) was mixed with Lipofectamine RNAiMAX reagent in serum-free DMEM. Suspended cells were added to the mixture and the cell mixture was plated and cultured for 24 h before experimental analysis.
Microarray expression analysis
Total RNA was collected from MKN45 cells using the RNeasy mini kit (Qiagen, Hilden, Germany). Microarray expression analyses were performed using the Agilent SurePrint G3 Human GE v2 8×60K Microarray (Design ID: 039494) (Agilent Technologies, Palo Alto, CA).
BrdU assay
BrdU assay was performed using the BrdU Cell Proliferation assay kit (Merck, Marmstadt, Germany) following the manufacturer’s protocol. Briefly, cells in a 96-well plate were cultured in medium with BrdU for 12 h. BrdU incorporation was detected with anti-BrdU. Signals were measured by spectrophotometer analysis at 450/540 nm.
Western blot
Cells were lysed by sonication in 10 mM Tris-HCl (pH 7.4), 150 mM NaCl, 1 mM EDTA, 1 mM EGTA, and a protease inhibitor cocktail (Roche Diagnostics, Mannheim, Germany) at 4 °C. Protein concentrations were determined with a protein assay rapid kit (Wako,Tokyo). Cell lysates (10 µg of protein) were separated by SDS-PAGE and transferred to a PVDF membrane. The membrane was incubated with primary antibodies at 1:1000 dilution with TBS-Tween 20 buffer, followed by incubation with HRP-conjugated secondary antibody at 1:5000 dilution. The membranes were processed using the ECL detection kit and images were obtained using a LAS-4000 image analyzer (Fujifilm, Tokyo).
Immunofluorescence staining
Cells were plated on coverslips and fixed with 3.7% formaldehyde in PBS for 30 min, followed by permeabilization in 0.1% Triton X-100 in PBS for 15 min. Cells were then incubated with anti-phospho-p38 antibody (1:200 in 0.1% BSA/PBS) for 1 h, followed by incubation with Alexa Fluor 488-conjugated anti-rabbit IgG for 1 h. The coverslips were mounted with mounting medium containing DAPI (ProLong Gold Antifade Mounting with DAPI; Invitrogen), and fluorescence images were obtained with confocal microscopy (Leica TCS-SP2, Wetzler, Germany).
RESULTS
Fucoidan impedes the cell cycle
We first examined the effects of fucoidan on proliferation of MKN45 cells using the BrdU incorporation assay. Low doses of fucoidan had a minimal impact on MKN45 cell proliferation, while high doses of fucoidan, over 5 mg/mL, dramatically decreased BrdU incorporation. The rate of MKN45 cell proliferation inhibition by fucoidan was approximately 50% (Fig. 1A).
Fig. 1.
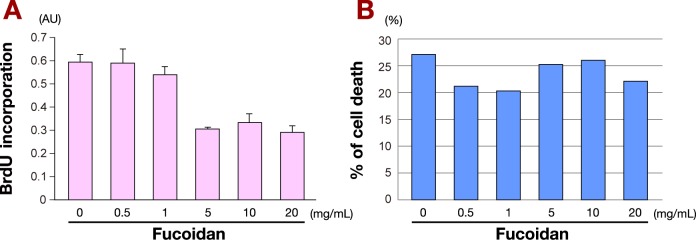
Effect of fucoidan on MKN45 cells.
A: Fucoidan impedes proliferation of MKN45 cells. The rate of MKN45 cell proliferation inhibition in the presence of increasing amounts of fucoidan was examined using BrdU incorporation assay. Over 5 mg/mL of fucoidan resulted in decreased BrdU incorporation.Inhibition rate was approximately 50%.
B: Cytotoxic effects of fucoidan in MKN45 cells. Cytotoxic effects of increasing amounts of fucoidan in MKN45 cells were examined by LDH assay. Cells were cultured with fucoidan as indicated for 24 h. Increasing fucoidan concentrations did not affect the rate ofcell death, even at high doses.
AU, arbitrary unit.
Cytotoxicity of fucoidan by LDH assay
The cytotoxic effects of treatment of MKN45 cells with increasing amounts of fucoidan for 24 h were examined by LDH assay (Fig. 1B). The results showed that 24 h fucoidan treatment did not induce cell death, even at high doses.
Clonogenic assay reveals fucoidan cytotoxic effects
To gain fuller insight into the potential cytotoxic effects of fucoidan, we performed clonogenic assays and extended the incubation time to 6 days (Fig. 2A). Starting at day 4, the numbers of cells were clearly reduced with larger doses of fucoidan. Colony formation assays showed similar results (Fig. 2B). We observed lower cell density of MNK45 cells cultured with high doses of fucoidan, and this reduction occurred in a dose-dependent manner.
Fig. 2.

Fucoidan inhibition of MKN45 cells.
A: Fucoidan inhibition of MKN45 cell growth. Clonogenic assays were performed by culturing MKN45 cells for 6 days in the presence of increasing amounts of fucoidan as indicated (F0-F20), and counting cells with a hemocytometer. Starting at day 4, the numbers of cells were clearly reduced with higher doses of fucoidan.
B: Fucoidan inhibits colony formation of MKN45 cells in a dose-dependent manner. Colony formation assay of MKN45 cells incubated with increasing amounts of fucoidan. Cell density was lower with increasing concentrations of fucoidan.
Microarray analyses of fucoidan-treated cells
To identify potential genes involved in mediating the cellular effects of fucoidan, total RNA was collected from MKN45 cells and microarray expression analyses were performed using the Agilent Human microarray kit. A total of 20278 sequences were examined at DNA Chip Research (Kanagawa, Japan), and 496 genes were found to be upregulated and 774 genes were downregulated by fucoidan. Table 1 lists the genes in MKN45 cells with significant changes in expression in response to fucoidan (P < 0.05).Among these genes, we observed that MAP3K5, also known as ASK1 (apoptosis signal-regulating kinase), was upregulated by 1.38-fold in response to fucoidan. Microarray analyses accession number:GSE58774.
Table 1. The differentially expressed G0 categories and gene lists in fucoidan treated MKN45 cells.
| Upregulated | Human insulin signaling (7) | SORBS1 |
| FLOT2 | ||
| PSCD3 | ||
| MAP4K1 | ||
| MAP3K5 | ||
| FOS | ||
| EGR1 | ||
| GPCRs, class a rhodopsin-like (5) | HTR6 | |
| OR3A3 | ||
| P2RY4 | ||
| ADRA2A | ||
| OR5V1 | ||
| T-cell receptor signaling pathway (5) | MAP4K1 | |
| RASGRP2 | ||
| FOS | ||
| CABIN1 | ||
| CARD11 | ||
| Monoamine GPCRs (2) | HTR6 | |
| ADRA2A | ||
| MAPK signaling pathway (5) | HSPA8 | |
| MAP4K1 | ||
| MAP3K5 | ||
| FOS | ||
| MAPT | ||
| Downregulated | Irinotecan pathway (Homo sapiens) (2) | CES1 |
| ABCC2 | ||
| Blood clotting cascade (2) | ERPINE1 | |
| F10 | ||
| Fatty acid omega oxidation (2) | CYP1A2 | |
| CYP1A1 | ||
| Focal adhesion (9) | TNC | |
| EGFR | ||
| CAV1 (A_24_P12626) | ||
| CAV3 | ||
| CAV1 (A_23_P134454) | ||
| RAPGEF1 | ||
| COMP | ||
| MAPK8 | ||
| Complement activation, classical pathway (3) | CFH | |
| SERPINE1 | ||
| F10 |
Genes that showed significant (P < 0.05) changes in upregulated or downregulated expression in MKN45 cells in the presence of fucoidan.
Fucoidan downregulates cell cycle promoting proteins
From the results of the BrdU incorporation assay and mRNA microarray analysis, we hypothesized that fucoidan affected the expressions of cell cyclerelated genes. Western blot analysis revealed that treatment of MNK45 cells with over 5 mg/mL of fucoidan resulted in decreased levels of phosphorylated ASK1 and phosphorylated p38 MAPK. In contrast, the expression of ASK1 was slightly increased in a dose-dependent manner. The expressions of p38 MAPK, ERK, and phosphorylated ERK were unchanged (Fig. 3A).
Fig. 3.
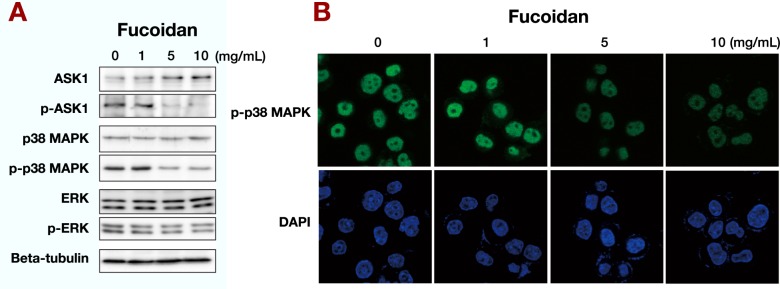
Effects of fucoidan on phosphorylation.
A: Fucoidan downregulates cell-cycle promoting proteins. Western blot analysis revealed that over 5 mg/mL of fucoidan deceased the levels of phosphorylated ASK1 and phosphorylated p38 MAPK. However, the expression of ASK1 was slightly increased in a dosedependent manner. The expressions of p38 MAPK, ERK, and phosphorylated ERK were not changed.
B: Fucoidan reduces phosphorylated p38 in the nucleus. MKN45 cells were incubated with fucoidan as indicated and analyzed by immunofluorescence for phosphorylated p38. Treatment with fucoidan at 5 and 10 mg/mL clearly reduced the levels of phosphorylated p38 protein in the nucleus of MKN45 cells, which was consistent with the western blot results.
We also performed fluorescence immunostaining of MNK45 cells with phosphorylated p38 antibody and found that 5 and 10 mg/mL fucoidan treatment clearly reduced phosphorylated p38 protein levels in the nucleus (Fig. 3B), which was consistent with the western blot results. These results suggested the possibility that fucoidan inhibited proliferation of MKN45 cells by suppressing the ASK1-p38 signaling pathway.
ASK1 knockdown reduces proliferation of MKN45 cells
To examine the effect of ASK1 suppression on MKN45 cell proliferation, ASK1 siRNA was transfected into MKN45 cells. Western blot analyses confirmed that ASK1 siRNA reduced ASK1 protein levels compared with control cells (Fig. 4A). BrdU assay revealed that genetic inhibition of ASK1 repressed MKN45 cell proliferation (Fig. 4B).
Fig. 4.
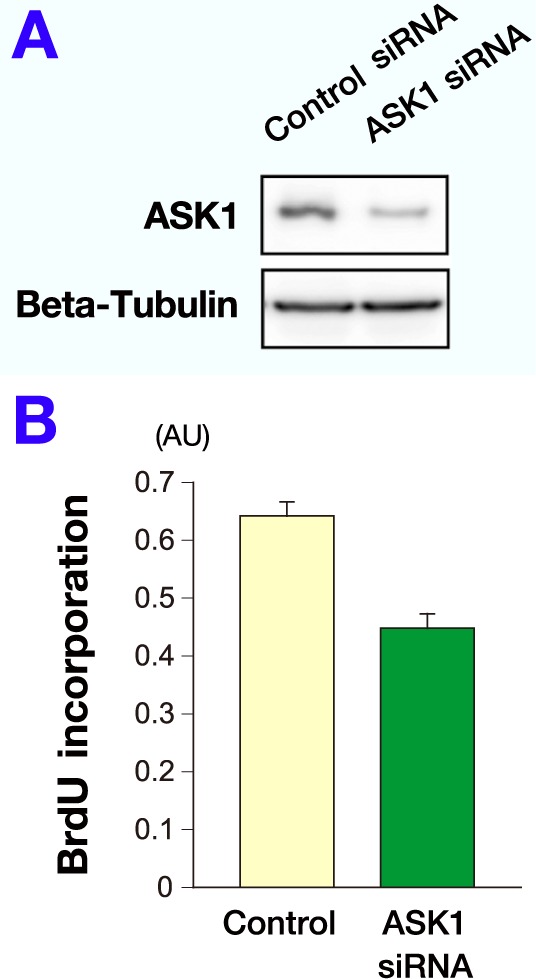
ASK1 knockdown decreases proliferation of MKN45 cells.
A: Western blot analysis confirmed that transfection with ASK1 siRNA reduced the expression level of ASK1 protein compared with control cells. Beta-tubulin was used as an internal control.
B: To examine the effect of ASK1 suppression on proliferation of MKN45 cells, ASK1 siRNA was transfected into MKN45 cells and Brdu assay was performed. Genetic inhibition of ASK1 repressed the proliferative activity of MKN45 cells.
AU, arbitrary unit.
DISCUSSION
Recent studies have shown that marine bioactive compounds can either modulate signaling pathways leading to cell cycle regulation or directly alter cell cycle regulatory molecules in cancer therapy. The anti-proliferative activity of fucoidan may be a result of its interference in cell cycle progression.
Previous studies demonstrated that fucoidan caused a cell cycle arrest at the G0/G1 phase in hepatocarcinoma cell lines18 and suppressed cell proliferation and arrested the cell cycle in hepatocellular carcinoma cell lines.19 Our BrdU results showed that cell proliferation was suppressed at a dose of fucoidan of over 5 mg/mL in 24 h, which impeded DNA synthesis and cell proliferation during S phase in MKN45 cells. This result demonstrated that fucoidan induced cell cycle arrest in MKN45 cells.
We explored the cytotoxic effects of fucoidan in MKN45 cells by LDH assay. After 24 h of incubation, fucoidan-treated MKN45 cells did not exhibit any elevation in LDH release compared with the control group, even at high doses (Fig. 1B). This result was consistent with previous studies.18, 19 However, other studies demonstrated cytotoxicity of fucoidan in different cancer cells. 19, 20
These discrepancies in cytotoxicity among fucoidans are likely due to variations in fucoidan polymer structure and function, as fucoidans originate from various species, anatomical regions, and under different growing conditions of brown seaweeds. Fucoidans are also subject to specific extraction and purification procedures. Furthermore, fucoidans may also exhibit celltype-specific effects, with different functions in different cancer cell lines. Additionally, even though cell proliferation was repressed by fucoidan in previous reports, the repression was not evident at 24 h by MTT assay.19 We also did not observe cytotoxic effects of fucoidan at 24 h. We speculated that a longer time course may better reveal fucoidan effects under long exposure conditions, and subsequently extended the incubation time to 6 days. Under longer treatment conditions, we observed both a marked decrease in cell number and reduction of cell density in a dose-dependent manner.
We performed mRNA microarray analysis to identify potential gene targets of fucoidan in mediating its cell cycle effects. MAP3K5 or ASK1 that was identified as a MAPK kinase kinase, was upregulated 1.38-fold in response to fucoidan. ASK1 is involved in regulating the cell cycle and is phosphorylated at the beginning of S phase.21 This suggested that phosphorylation might facilitate ASK1’s function in the spindle-kinetochore interaction, which presumably occurs during the S phase after the duplication of kinetochores. This also impliesthat decreasing phosphorylated ASK1 would impair the cell cycle during S phase.
In the MAPK pathway, p38 MAPK functions as a downstream kinase of ASK1. p38 MAPK was reported to promote cancer cell growth and survival.22 In a study investigating the effect of fucoidan on lung carcinoma, fucoidan treatment resulted in decreased phospho-p38 levels,23 which demonstrated anti-cancer effect due to fucoidan.
Another MAPK, ERKs are known to promote cell survival, differentiation, cell proliferation 24 and cell death.25 A previous study showed fucoidan clearly decreased the phosphorylation of ERK in HS-Sultan cells.26
In evaluating the results of the mRNA microarray analysis, we therefore focused on genes involved in cell cycle regulation to examine potential mediators of fucoidan. Based on the BrdU incorporation results, we hypothesized that the protein levels of ASK1 and its related proteins would be altered by fucoidan, and we performed western blot analysis of ASK1, p38 MAPK, and ERK steady state and phosphorylated forms to determine the effects of fucoidan.
Treatment with over 5 mg/mL fucoidan effectively deceased phosphorylated ASK1 and phosphorylated p38 MAPK levels, while ERK expression remained unchanged. The western blot results furthered the BrdU incorporation findings, suggesting that fucoidan caused reduction of phosphorylated ASK1 expression, arresting the cell cycle in S phase and eventually impairing DNA synthesis. This results in damaging proliferation of cancer cells.
Consistent with the microarray analysis, fucoidan increased ASK1 protein levels. Increased ASK1 protein could possibly result from a reduction of phosphorylated ASK1 by fucoidan. As a consequence of reduced phosphorylated ASK1, the mRNA levels were compensatory amplified, resulting in increased expression of ASK1.
A previous study demonstrated elevation of ASK1 in human gastric cancer and showed that overexpression of ASK1 promoted gastric cancer cell proliferation.27 Another report found that ASK1 inhibition prevents gastric cancer cell growth in vitro and in vivo. 28 This mechanism involved cyclin D, a known regulator of the G1 to S phase transition.29 However, this mechanism seemed specific to gastric cancer cells, as ASK1 knockdown in other cell lines did not inhibit proliferation. 28 The previous study supports the results of the current study,demonstrating that genetic inhibition of ASK1 repressed MKN45 cell proliferation.
To elucidate the mechanisms underlying the anticancer activities of fucoidan, determining its binding partners and targets would help clarify its activities. One study reported that fucoidan acts as a polysaccharide ligand of P-selectin, with nanomolar affinity.30 P-selectin is an adhesion molecule expressed at the surface of endothelial cells and platelets on activation. P-selectin promotes platelet, endothelium, and leukocyte interactions, and was reported to cause cardiovascular events.31
Fucoidan exhibited the highest affinity for Pselectin32and repressed its function in platelet aggregation.31 This implied that fucoidan targets the surface of endothelial cells, with potential consequences on downstream signal transduction pathways. Our findings show that fucoidan induces alterations of gene expression, which may possibly involve its function at the cell surface.
In conclusion, our study indicated that fucoidan may suppress cellular proliferation and DNA synthesis in MKN45 cells by suppressing the ASK1-p38 signaling pathway through a reduction in phosphorylated ASK1 levels. Our findings strongly suggest fucoidan as a potential therapeutic agent for cancer therapies. Further investigations would provide new approaches to the development of effective chemotherapy regimens.
Acknowledgments
Acknowledgments: This work was supported by Marine Products Kimuraya Co., Ltd. (Sakaiminato, Japan).
The authors declare no conflict of interest.
REFERENCES
- 1. Wang J, Zhang Q, Zhang Z, Li Z. Antioxidant activity of sulfated polysaccharide fractions extracted from Laminaria japonica. Int J Biol Macromol. 2008; 42: 127-32. . [DOI] [PubMed] [Google Scholar]
- 2.Lekseyenko TV, Zhanayeva SY, Venediktova AA, Zvyagintseva TN, Kuznetsova TA, Besedonova NN. Antitumor and antimetastatic activity of fucoidan, a sulfated polysaccharide isolated from the Okhotsk Sea Fucus evanescens brown alga. Bull Exp Biol Med. 2007; 143: 730-2. . [DOI] [PubMed] [Google Scholar]
- 3.Maruyama H, Tamauchi H, Iizuka M, Nakano T. The role of NK cells in antitumor activity of dietary fucoidan from Undaria pinnatifida sporophylls (Mekabu). Planta Med. 2006; 72: 1415-7. . [DOI] [PubMed] [Google Scholar]
- 4. Simon M, Grandage VL, Linch DC, Khwaja A. Constitutive activation of the Wnt/beta-catenin signalling pathway in acute myeloid leukaemia. Oncogene. 2005; 24: 2410-20. . [DOI] [PubMed] [Google Scholar]
- 5. Ishii H, Iwatsuki M, Ieta K, Ohta D, Haraguchi N, Mimori K. Cancer stem cells and chemoradiation resistance. Cancer Sci. 2008; 99: 1871-7. . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Usui T, Asari K, Mizuno T. Isolation of highly purified fucoidan from Eisenia bicyclis and its anticoagulant and antitumor activities. Agric Biol Chem. 1980; 44: 1965-6. 10.1080/00021369.1980.10864249. [DOI] [Google Scholar]
- 7. Song JQ, Xu YT, Zhang HK. Immunomodulation action of sulfate polysaccharide of Laminaria japonica on peritoneal macrophages of mice. Chin J Immunol. 2000; 16: 70. [Google Scholar]
- 8. Shi ZY, Guo YZ, Wang Z. Pharmacological activity of fucoidan from Laminaria japonica. J Shanghai Fisheries Univ [Internet]. 2000. [cited 2013 June 30]; (3): 268-71. Available from: http://caod.oriprobe.com/articles/2721133/Pharmacological_activity_of_fucoidan_from_Laomaroa_japonica.htm. [Google Scholar]
- 9. Cumashi A, Ushakova NA, Preobrazhenskaya ME, D’Incecco A, Piccoli A, Totani L. A comparative study of the antiinflammatory, anticoagulant, antiangiogenic, and antiadhesive activities of nine different fucoidans from brown seaweeds. Glycobiology. 2007; 17: 541-52. . [DOI] [PubMed] [Google Scholar]
- 10. Cheng-Qian Y, Xin-Jing W, Wei-Xinbing , Zhuang-Lei G, Hong-Peng Z, Songde X. Lovastatin inhibited the growth of gastric cancer cells. Hepatogastroenterology. 2014; 61: 1- 4. . [PubMed] [Google Scholar]
- 11. Terai S, Fushida S, Tsukada T, Kinoshita J, Oyama K, Okamoto K. Bone marrow derived “fibrocytes” contribute to tumor proliferation and fibrosis in gastric cancer. Gastric Cancer. Epub 2014 May 4. . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Zheng Y, Wang DD, Wang W, Pan K, Huang CY, Li YF. Reduced expression of uroplakin 1A is associated with the poor prognosis of gastric adenocarcinoma patients. PLoS One. 2014; 9: e93073. . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Kawamoto H, Miki Y, Kimura T, Tanaka K, Nakagawa T, Kawamukai M. Effects of fucoidan from Mozuku on human stomach cell lines. Food Sci Technol Re. 2006; 12: 218-22. 10.3136/fstr.12.218. [DOI] [Google Scholar]
- 14. Morimoto M, Takatori M, Hayashi T, Mori D, Takashima O, Yoshida S. Depolymerization of sulfated polysaccharides under hydrothermal conditions. Carbohydr Res. 2014; 384: 56- 60. . [DOI] [PubMed] [Google Scholar]
- 15. Osaki T, Kitahara K, Okamoto Y, Imagawa T, Tsuka T, Miki Y. Effect of fucoidan extracted from mozuku on experimental cartilaginous tissue injury. Mar Drugs. 2012; 10: 2560-70. . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Ikeguchi M, Yamamoto M, Arai Y, Maeta Y, Ashida K, Katano K. Fucoidan reduces the toxicities of chemotherapy for patients with unresectable advanced or recurrent colorectal cancer. Oncol Lett. 2011; 2: 319-22. . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Abe S, Hiramatsu K, Ichikawa O, Kawamoto H, Kasagi T, Miki Y. Safety evaluation of excessive ingestion of mozuku fucoidan in human. J Food Sci. 2013; 78: T648-51. . [DOI] [PubMed] [Google Scholar]
- 18. Nagamine T, Hayakawa K, Kusakabe T, Takada H, Nakazato K, Hisanaga E. Inhibitory effect of fucoidan on Huh7 hepatoma cells through downregulation of CXCL12. Nutr Cancer. 2009; 61: 340-7. . [DOI] [PubMed] [Google Scholar]
- 19. Fukahori S, Yano H, Akiba J, Ogasawara S, Momosaki S, Sanada S. Fucoidan, a major component of brown seaweed, prohibits the growth of human cancer cell lines in vitro. Mol Med Rep. 2008; 1: 537-42. . [PubMed] [Google Scholar]
- 20. Yang C, Chung D, Shin I, Lee H, Kim J, Lee Y. Effects of molecular weight and hydrolysis conditions on anticancer activity of fucoidans from sporophyll of Undaria pinnatifida. Int J Biol Macromol. 2008; 43: 433-7. . [DOI] [PubMed] [Google Scholar]
- 21. Li Y, Elledge SJ. The DASH complex component ASK1 is a cell cycle-regulated Cdk substrate in Saccharomyces cerevisiae. Cell Cycle. 2003; 2: 143-8. . [PubMed] [Google Scholar]
- 22.Junttila MR, Li SP, Westermarck J. Phosphatase-mediated crosstalk between MAPK signaling pathways in the regulation of cell survival. FASEB J. 2008; 22: 954-65. . [DOI] [PubMed] [Google Scholar]
- 23. Boo HJ, Hyun JH, Kim SC, Kang JI, Kim MK, Kim SY. Fucoidan from Undaria pinnatifida induces apoptosis in A549 human lung carcinoma cells. Phytother Res. 2011; 25: 1082-6. . [DOI] [PubMed] [Google Scholar]
- 24. Seger R, Krebs EG. The MAPK signaling cascade. FASEB J. 2003; 9: 726-35. . [PubMed] [Google Scholar]
- 25. Cheung EC, Slack RS. Emerging role for ERK as a key regulator of neuronal apoptosis. Sci STKE. 2004; 251: 45. . [DOI] [PubMed] [Google Scholar]
- 26. Aisa Y, Miyakawa Y, Nakazato T, Shibata H, Saito K, Ikeda Y. Fucoidan induces apoptosis of human HS-sultan cells accompanied by activation of caspase-3 and downregulation of ERK pathways. Am J Hematol. 2005; 78: 7-14. . [DOI] [PubMed] [Google Scholar]
- 27. Hayakawa Y, Hirata Y, Nakagawa H, Sakamoto K, Hikiba Y, Kionshita H. Apoptosis signal-regulating kinase 1 and cyclin D1 compose a positive feedback loop contributing to tumor growth in gastric cancer. Proc Natl Acad Sci USA. 2011; 108: 780-5. . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Hayakawa Y, Hirata Y, Sakitani K, Nakagawa H, Nakata W, Kinoshita H. Apoptosis signal-regulating kinase-1 inhibitor as a potent therapeutic drug for the treatment of gastric cancer. Cancer Sci. 2012; 103: 2181-5. . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Sherr CJ. Cancer cell cycles. Science. 1996; 274: 1672-7. . [DOI] [PubMed] [Google Scholar]
- 30. Rouzet F, Bachelet-Violette L, Alsac JM, Suzuki M, Meulemans A, Louedec L. Radiolabeled fucoidan as a p-selectin targeting agent for in vivo imaging of plateletrich thrombus and endothelial activation. J Nucl Med. 2011; 52: 1433-40. . [DOI] [PubMed] [Google Scholar]
- 31. Yokoyama S, Ikeda H, Haramaki N, Yasukawa H, Murohara T, Imaizumi T. Platelet P-selectin plays an important role in arterial thrombogenesis by forming large stable platelet-leukocyte aggregates. J Am Coll Cardiol. 2005; 45: 1280-6. . [DOI] [PubMed] [Google Scholar]
- 32. Bachelet L, Bertholon I, Lavigne D, Vassy R, Jandrot-Perrus M, Chaubet F. Affinity of low molecular weight fucoidan for P-selectin triggers its binding to activated human platelets. Biochim Biophys Acta. 2009; 1790: 141-6. . [DOI] [PubMed] [Google Scholar]