

Abstract
Ixodes persulcatus, Ixodes pavlovskyi, and Dermacentor reticulatus ticks inhabiting Western Siberia are responsible for the transmission of a number of etiological agents that cause human and animal tick-borne diseases. Because these ticks are abundant in the suburbs of large cities, agricultural areas, and popular tourist sites and frequently attack people and livestock, data regarding the microbiomes of these organisms are required. Using metagenomic 16S profiling, we evaluate bacterial communities associated with I. persulcatus, I. pavlovskyi, and D. reticulatus ticks collected from the Novosibirsk region of Russia. A total of 1214 ticks were used for this study. DNA extracted from the ticks was pooled according to tick species and sex. Sequencing of the V3-V5 domains of 16S rRNA genes was performed using the Illumina Miseq platform. The following bacterial genera were prevalent in the examined communities: Acinetobacter (all three tick species), Rickettsia (I. persulcatus and D. reticulatus) and Francisella (D. reticulatus). B. burgdorferi sensu lato and B. miyamotoi sequences were detected in I. persulcatus and I. pavlovskyi but not in D. reticulatus ticks. The pooled samples of all tick species studied contained bacteria from the Anaplasmataceae family, although their occurrence was low. DNA from A. phagocytophilum and Candidatus Neoehrlichia mikurensis was first observed in I. pavlovskyi ticks. Significant inter-species differences in the number of bacterial taxa as well as intra-species diversity related to tick sex were observed. The bacterial communities associated with the I. pavlovskyi ticks displayed a higher biodiversity compared with those of the I. persulcatus and D. reticulatus ticks. Bacterial community structure was also diverse across the studied tick species, as shown by permutational analysis of variance using the Bray-Curtis dissimilarity metric (p = 0.002). Between-sex variation was confirmed by PERMANOVA testing in I. persulcatus (p = 0.042) and I. pavlovskyi (p = 0.042) ticks. Our study indicated that 16S metagenomic profiling could be used for rapid assessment of the occurrence of medically important bacteria in tick populations inhabiting different natural biotopes and therefore the epidemic danger of studied foci.
Introduction
Ticks belonging to the genera Ixodes and Dermacentor (the family Ixodidae) are capable of transmitting a number of bacteria and viruses to humans, including causative agents of Lyme disease, rickettsioses, ehrlichioses, relapsing fever borreliosis, tularemia, Q fever, and tick-borne encephalitis [1,2,3]. In Western Siberia, I. persulcatus ticks are the main vector for tick-borne encephalitis and Kemerovo viruses [4,5,6] and many bacterial agents including Borrelia burgdorferi sensu lato, B. miyamotoi, “Candidatus Rickettsia tarasevichiae”, Anaplasma phagocytophilum, Ehrlichia muris, and Bartonella spp. [7,8,9,10,11,12,13,14]. I. pavlovskyi ticks inhabit the southern part of Western Siberia (Altai, Kuznetsk Alatau, and Salair mountains and foothill areas) and are responsible for transmission of infectious agents of Lyme disease and a tick-borne encephalitis virus [15,16]. Since the beginning of this century, I. pavlovskyi ticks have been abundant in the green belts of the Novosibirsk and Tomsk cities, which are located more northward in the Western Siberian Plain, and currently, this tick species is more prevalent compared with the I. persulcatus ticks and accounts for 82 and 90% of the collected samples, respectively [17,18,19]. A third Ixodidae tick species, Dermacentor reticulatus, is abundant in Western Siberia and is involved in the transmission of etiological agents of human and animal tick-borne diseases, such as Omsk hemorrhagic fever virus, B. burgdorferi s. l., Rickettsia raoultii, Bartonella quintana, and B. canis [11,14,20,21].
Recently, new microorganisms associated with human infections have been detected in different Ixodidae tick populations in the Asian region of Russia [6,9,12,22]; however, it is likely that a number of infectious agents are still unknown. Currently, 16S profiling is among the most powerful tools for examining microbial communities. This method uses polymerase chain reaction (PCR) with primers specific to the conserved regions of 16S rRNA genes to obtain a set of the corresponding DNA fragments. High-throughput sequencing of the amplicons is performed using second-generation sequencing platforms [23]. Analysis of the 16S rRNA gene sequences allows for phylotyping of the prokaryotes, including the organisms unable to be cultivated [24], which is especially important when analyzing symbiotic communities. This approach has been successfully used to examine the I. ricinus microbiome and allowed for the detection of various infectious agents within this species, including Borrelia species, Rickettsia spp., A. phagocytophilum, Ehrlichia spp., and “Candidatus Neoehrlichia mikurensis” [25,26,27]. Methods including 16S profiling were also applied for examination of microbial diversity in Amblyomma americanum, the most commonly encountered tick species in the southeastern-central USA [28,29], and it was found that some members of the bacterial community within A. americanum change during the tick life cycle. Furthermore, microbiomes from the salivary glands of Ixodes ovatus, I. persulcatus and Haemaphysalis flava ticks inhabiting Japan were examined [30]. This study indicated that bacterial communities in tick microbiomes were different between tick species [30].
I. persulcatus, I. pavlovskyi, and D. reticulatus are abundant in the green belts of large Siberian cities, agricultural areas, and popular tourist sites, and little is currently known about the bacterial communities associated with these ticks which are able to transmit a number of infectious agents and frequently attack people and home animals, so the goal of this study was to assess the taxonomic structure of I. persulcatus, I. pavlovskyi, and D. reticulatus microbiomes using high-throughput sequencing.
Methods
Tick collection and sample preparation
Adult ticks were collected by flagging from three sampling sites in the Novosibirsk region of Russia (Table 1). I. persulcatus ticks were flagged in site 1 (forest-steppe, Toguchin district); I. pavlovskyi and D. reticulatus ticks were flagged in sites 2 and 3, respectively (forest suburbs of Novosibirsk, Novosibirsk district). Coordinates of the sampling sites are presented in Table 1. Each tick species dominated in the selected site. No specific permission was required for our activities in locations involved in the study. Our field activities did not involve endangered or protected species. Ticks were sampled from 24 April to 30 May 2012 during the peak activity period for these Ixodidae ticks. Live ticks were taxonomically characterized by microscopic analysis using standard taxonomic keys. The relative abundance was assessed as the number of ticks collected per 1 km. Before study, ticks were stored at -70°C. A total of 1214 ticks were used for this study (I. persulcatus, n = 414; I. pavlovskyi, n = 436; D. reticulatus, n = 364).
Table 1. Library description.
| Species | Ixodes persulcatus | Ixodes pavlovskyi | Dermacentor reticulatus | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Flagging site | Site 1 | Site 2 | Site 3 | |||||||||
| Site description | forest-steppe | forest suburbs | forest suburbs | |||||||||
| Site coordinates | N55°01'07'', E84°06'41'' | N54°49'21'', E83°06'41'' | N54°53'15'', E83°08'45" | |||||||||
| Sex | male | female | male | female | male | female | ||||||
| Library name | PersM.1 | PersM.2 | PersF.3 | PersF.4 | PavlM.5 | PavlM.6 | PavlF.7 | Pavl.F8 | DermM.9 | DermM.10 | DermF.11 | DermF.12 |
| Number of ticks in library | 120 | 120 | 87 | 87 | 120 | 120 | 98 | 98 | 89 | 88 | 94 | 93 |
| Number of reads in library | 5168 | 5806 | 14043 | 11884 | 12796 | 12999 | 19161 | 12466 | 5322 | 9372 | 12639 | 10992 |
DNA extraction and sequencing
Before DNA extraction, ticks were washed in distilled water, 70% ethanol and distilled water as described previously [25]. Then ticks were grouped according to their species and sex; each group consisted of eight I. persulcatus or I. pavlovskyi males, or up to five I. persulcatus, I. pavlovskyi females, or up to five D. reticulatus ticks. Ticks within each group were frozen (30 min/ – 80°C), macerated with a sterile micropestle, and total DNA was extracted using the Proba NK kit (DNA-technology, Russia) according to the manufacturer’s instruction. This kit provides DNA extraction from 100 mkg specimen. Aliquots of DNA from each group were subsequently combined resulted in 12 pooled samples (persM.1, persM.2, persF.3, persF.4, pavlM.5, pavlM.6, pavlF.7, pavlF.8, dermM.9, dermM.10, dermF.11, and dermM.12) providing two biological replicates for each ‘one species-one sex’ combination. Each pooled sample included 88–120 males or 93–98 females (Table 1).
The presence of bacterial DNA was assessed by PCR using primers that specifically amplified the hypervariable regions V3-V5 of 16S rRNA genes [24]. 10 ng of DNA was used as a template in PCR. To prevent contamination, we performed DNA isolation, PCR master mix assembly, and amplification in separate rooms. Aerosol-free pipette tips were used at each stage. We included negative control reactions with bi-distilled water in each experiment.
Fusion primers comprised of the fragment-specific 3’ end, barcode, and adapter sequence for the Illumina MiSeq platform (Illumina) were used for amplification. The length of the amplified fragment was approximately 430 bp. Sequencing by paired-end reads with a read length of 250 nucleotides was conducted on the Illumina MiSeq platform at the Genomics Core Facility of the Siberian Branch of the Russian Academy of Sciences.
Data analysis
Paired-end reads from Illumina Miseq platform were assembled using Pandaseq v2.4. Reads containing more than six ambiguous bases were eliminated from the dataset [31]. The pooled samples were taxonomically characterized using the QIIME v. 1.7.0 software package [32]. Analysis included read clustering by the UCLUST software package with a minimum similarity coefficient of 97% [33]. Representative sequences for each operational taxonomic unit (OTU) were selected. Taxonomic classification for each representative was performed using the RDP classifier with 80% confidence [34]. Taxonomic trees were constructed using the MEGAN software package.
Hierarchical clustering, between-class analysis and diversity analysis were performed using the R programming language. Hierarchical clustering of pooled samples was performed using hclust and visualized as a heatmap. Diversity analysis was completed using the “vegan” package [35]. Between-class analysis was performed using the package “ade4” with euclidean and UniFRAC distances [36]. The Permutational Multivariate Analysis of Variance (PERMANOVA) function from package “vegan” was used to assess the effect of gender and species on taxonomic profiles. The permuted p-value was obtained by the Bray-Curtis dissimilarity measure for 10 000 permutations.
Results
Bacterial diversity estimation
The 16S rRNA gene fragments used for phylotyping contained V3, V4 and V5 hypervariable regions. These regions allow for effective genus-level classification of bacteria [24,37]. A total of 12 pooled samples were sequenced, resulting in twelve libraries. The number of reads per library was 6152 to 23 477 (Table 1). To avoid biodiversity inflation caused by sequencing errors and PCR chimeras, all OTUs with single observation count were removed from data. In total, 25767 OTUs were singletons while 2802 OTUs were presented in two or more copies. DNA sequences from 726 OTUs were detected in the I. persulcatus ticks (520 in male ticks and 390 in female ticks); 2306 OTUs in the I. pavlovskyi ticks (1709 in males and 1316 in females); and 654 OTUs in the D. reticulatus ticks (474 in males and 437 in females). Extrapolated richness of the bacterial communities from the examined groups was assessed using chao and bootstrap estimators (Table 2). Because of the different data volumes for each library, the samples were subsampled according to the read number in the smallest library. Shannon index and beta diversity index were calculated on rarefied data, as well. This estimation proved that the bacterial communities detected in I. pavlovskyi ticks were the most diverse, containing 2058 ± 232 OTUs and Shannon index of 3.86 ± 0.73. The D. reticulatus and I. persulcatus microbiomes were less diverse, comprised of 637 ± 65 OTUs (H = 2.01 ± 0.23) for D. reticulatus, and 773 ± 98 OTUs (H = 2.06 ± 0.27) for I. persulcatus ticks.
Table 2. Diversity analysis.
| Library number | PersM.1 | PersM.2 | PersF.3 | PersF.4 | PavlM.5 | PavlM.6 | PavlF.7 | PavlF.8 | DermM.9 | DermM.10 | DermF.11 | DermF.12 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Tick species | Ixodes persulcatus | Ixodes pavlovskyi | Dermacentor reticulatus | |||||||||
| Tick sex | male | female | male | female | male | female | ||||||
| Per-group diversity statistics | ||||||||||||
| Number of OTUs per group | 520 | 390 | 1709 | 1316 | 474 | 437 | ||||||
| Number of OTUs after rarefaction | 511 | 252 | 1233 | 792 | 400 | 307 | ||||||
| Extrapolated richness (Chao) | 699 ± 28 | 367 ± 24 | 1721 ± 47 | 1182 ± 46 | 558 ± 27 | 404 ± 19 | ||||||
| Extrapolated richness (bootstrap) | 598 ± 88 | 297 ± 47 | 1447 ± 214 | 937 ± 146 | 470 ± 71 | 357 ± 51 | ||||||
| Shannon index (H) | 2.29 ± 0.06 | 1.83 ± 0.03 | 4.48 ± 0.05 | 3.23 ± 0.24 | 2.18 ± 0.18 | 1.83 ± 0.05 | ||||||
| Beta diversity | 0.52 | 0.56 | 0.53 | 0.57 | 0.53 | 0.49 | ||||||
| Per-species diversity statistics | ||||||||||||
| Tick species | Ixodes persulcatus | Ixodes pavlovskyi | Dermacentor reticulatus | |||||||||
| Number of OTUs per species | 726 | 2306 | 654 | |||||||||
| Number of OTUs after rarefaction | 638 | 1693 | 532 | |||||||||
| Extrapolated richness (Chao) | 973 ± 46 | 2749 ± 89 | 849 ± 50 | |||||||||
| Extrapolated richness (bootstrap) | 773 ± 98 | 2058 ± 232 | 637 ± 65 | |||||||||
| Shannon index (H) | 2.06 ± 0.27 | 3.86 ± 0.73 | 2.01 ± 0.23 | |||||||||
| Beta diversity | 1.56 | 1.59 | 1.28 | |||||||||
Sample clustering and community structure
To detect differences in microbiome structures among different tick species, hierarchical clustering and between-class analysis were used. The most frequently represented OTUs in the pooled samples were used in hierarchical cluster analysis based on a distance matrix (Fig 1). Between-class analysis based on the taxonomic composition demonstrated that the species-level differences in the examined samples are detectable within the first two variation components (Fig 2). The intra-species variation within samples from the I. persulcatus and D. reticulatus ticks were relatively small versus that of I. pavlovskyi. A strict separation of the studied tick groups according to the tick species criterion was observed. When using the PCoA analysis with weighted unifrac distance measures, the first two components account for 73.51% of the total variation and provide for a more strict separation of the groups (Fig 2). PERMANOVA analysis based on in-sample OTU amounts also indicates a strict separation in microbiome structure among these three tick species (F-stat = 5.323, df = 2, p = 0.02).
Fig 1. Heatmap based on 30 mostly reprepreted OTUs.

Heatmap was constructed using euclidean distance method for log-scaled in-sample taxon share values. Sample names on the X axis consist of tick species, sex and library number. OTU names on the Y axis contain OTU's lowest common ancestor name and unique integer OTU key.
Fig 2. Between-class analysis plots for taxon-based distances.

A) MDS plot based on PCA for 150 taxa with the largest share. Principal component analysis was performed using package “ade4” with data scaling and centering. B) MDS plot based on the UniFRAC distance. Calculation of UniFRAC distance was performed using QIIME package. First and second principal components describe 40.69% and 32.82% of total variance respectively.
Sex-specific differences in the studied microbiomes were detected. Using distance matrix with euclidean distances, it was shown that microbiome of D. reticulatus ticks was the most homogeneous across all pooled samples while the sex-specific differences within the I. pavlovskyi and I. persulcatus samples were more significant (Fig 1). Within-group variations across the studied samples were varied. The male I. persulcatus and female D. reticulatus groups displayed the least intragroup variation versus the male and female I. pavlovskyi ticks that had a considerably higher variation value. PERMANOVA analysis based on in-sample OTU amounts shows that sex-specific variations in microbiome structure were substantial in the I. persulcatus (F-stat = 11.841, df = 1, p = 0.042) and I. pavlovskyi samples (F-stat = 4.0852, df = 1, p = 0.042) and insignificant in the D. reticulatus samples (F-stat = 2.7301, df = 1, p = 0.333).
Taxonomic composition
Occurrence of the detected taxa is shown on the MEGAN taxonomic tree (S1 Table, S1 Fig). The bacterial DNA sequences of Alpha- and Gammaproteobacteria types were prevalent in all of the assayed pooled samples with 30.2% and 60.8% average occurrence, respectively. The most abundant families were Rickettsiaceae (20.5%, on average, varying from 1.5% in I. pavlovskyi to 21.1% in D. reticulatus and 41.1% in I. persulcatus), Francisellaceae (16.7–55% in D. reticulatus) and Moraxellaceae (from 9% to 76% in I. persulcatus).
DNA sequences belonging to the bacterial genera Acinetobacter (Gammaproteobacteria, Pseudomonadales, Moraxellaceae) and Rickettsia (Alphaproteobacteria, Rickettsiales, Rickettsiaceae) were most abundant in all samples; however, the abundances of these groups in the I. persulcatus, I. pavlovskyi, and D. reticulatus bacterial communities varied considerably. Acinetobacter bacteria were prevalent in the female I. pavlovskyi (65.4%) and male I. persulcatus (45.6%) ticks and ranged from 8.8 to 31.3% in the microbiomes of the remaining tick groups (Fig 3, S1 Table, S1 Fig). In addition to Acinetobacter spp., other members of the order Pseudomonadales and families Moraxellaceae (Enhydrobacter spp., Psychrobacter spp., etc.) and Pseudomonadaceae (Pseudomonas spp.) were also detected. The occurrence of bacteria belonging to the genus Rickettsia was higher in the I. persulcatus (males, ~41%, and females, ~35%) and D. reticulatus (males, 18.5%, and females, 23.7%) ticks (Fig 3, S1 Fig), but considerably lower in I. pavlovskyi ticks, amounting to 2.5 and 3.8% in males and females, respectively. In addition, DNA from the Rickettsiales bacterium “Montezuma” was highly abundant (~ 40%) in the microbiome of female I. persulcatus ticks, whereas its rate did not exceed 1% in male I. persulcatus ticks and other pooled samples (Fig 3, S1 Fig).
Fig 3. Bar graph with the share of the 8 most represented taxa.

Share of Rickettsiales order and Enterobacteriaceae family cover only hits that hadn't be included in the lower levels of these taxa.
Bacteria from the Francisella genus (Gammaproteobacteria, Thiotrichales, Francisellaceae) were prevalent in the microbiomes of D. reticulatus ticks, representing approximately 50% of sequences (Fig 3, S1 Table), and were almost absent in the microbiomes of the Ixodes ticks. In addition to Francisella spp., Francisella-like endosymbionts [38] belonging to the family Francisellaceae were also detected in D. reticulatus ticks. The 16S rRNA gene fragments used in this study did not allow discrimination of pathogenic and non-pathogenic Francisella species.
DNA sequences of medically significant bacteria belonging to the genus Borrelia (Spirochaetes, Spirochaetales, Spirohaetaceae) were present in I. persulcatus and I. pavlovskyi ticks (S1 Table, S1 Fig). The selected 16S rRNA locus made it possible to assign borrelias to individual genetic groups. Both B. burgdorferi s. l. and B. miyamotoi were detected in the I. persulcatus and I. pavlovskyi ticks. No Borrelia spp. were found in the D. reticulatus ticks. Several pooled samples contained medically relevant bacteria from the family Anaplasmataceae (Alphaproteobacteria, Rickettsiales), although their occurrence was low (S1 Table, S1 Fig). DNA sequences from Ehrlichia spp. that were most similar to those of E. muris were detected in I. persulcatus and D. reticulatus ticks. I. persulcatus and I. pavlovskyi ticks contained A. phagocytophilum DNA; in addition, DNA sequences from Candidatus Neoehrlichia mikurensis were found in I. pavlovskyi male ticks.
Almost all pooled samples contained small amounts of DNA sequences from Pseudomonas spp., Chryseobacterium spp., Acidovorax spp., Janthinobacterium spp., Sphingobacterium spp., and other free-living saprophytic bacteria (Fig 3, S1 Table, S1 Fig). Presumably, these bacteria inhabit the tick chitin exoskeleton. Some animal and human symbiotic bacteria—Bacteroides, Prevotella, Escherichia, Enterobacter, Bifidobacterium, Yersinia, and others—were also detected in small amounts (S1 Table, S1 Fig).
In addition, DNA sequences from the bacteria belonging to the genus Ricketsiella recently classified as pathogens of arthropods were found in the D. reticulatus and I. pavlovskyi ticks [39]. These sequences were present in all D. reticulatus samples (0.01–0.9%) and males I. pavlovskyi samples (0.6–23.6%).
Discussion
The microbiomes associated with ticks contain symbiotic and parasitic species as well as infection agents transmitted by ticks to their vertebrate hosts, whose pathogenicity for ticks is unknown [40,41]. Here, we describe the biodiversity of the bacterial communities associated with the I. persulcatus, I. pavlovskyi, and D. reticulatus ticks that inhabit Western Siberia and frequently attack people and livestock in this region. Ticks were collected from three sampling sites, which are located within the Novosibirsk region and display similar climates typical for Northern forest-steppe area in Siberia. In these sampling sites, larval and nymphal stages of the studied ixodidae ticks are mainly fed on the northern birch mouse (Sicista betulina), the striped field mouse (Apodemus agrarius), the grey red-backed vole (Myodes rufocanus), the northern red-backed vole (M. rutilus), and the common shrew (Sorex araneus). These sampling sites were selected because each tick species was prevalent in the corresponding site and other ixodidae ticks were collected in solitary cases. The relative abundance of I. persulcatus, I. pavlovskyi, and D. reticulatus ticks in sampling sites 1, 2, and 3 in 2012 was 32.4, 31.8, and 41,9 individuals per km, respectively. Other ixodidae tick species were found in corresponding sampling sites in single cases.
More than 1200 ticks of three species were included in this study. To assess the bacterial communities associated with these tick species, 16S metagenomics profiling was performed on twelve pools–four pools for each species (two males and two females). Each pool contained 87–120 ticks. Such tick quantity increases result reliability for large-scale population studies. It should be noted that the prevalence of infectious agents in individual ticks inhabiting Novosibirsk region was studied earlier and it was found that the occurrence of certain infectious agents is low [9,12,14].
We detected both species- and sex-specific differences among the microbiomes of these three tick species. Different clustering algorithms demonstrated that the I. persulcatus, I. pavlovskyi, and D. reticulatus microbiomes differ in both their alpha-diversity levels and taxonomic structures. Taxonomic diversity of the symbiotic microbiomes in the studied ticks was relatively low when compared with that of higher animals and ecological systems. Notably, the symbiotic diversity of the I. pavlovskyi microbiomes exceeded that of the I. persulcatus and D. reticulatus ticks in a statistically significant manner. In addition to alpha-diversity, the examined microbiomes differed in their dominant species and abundances of various subdominants.
Each tick species was collected from a different sampling site associated with different microhabitats that could contribute to the variation in the studied microbiomes. However, these sampling sites are located in one geographic area with similar climate and reservoir host composition, so, we assume that at least a part of the differences among microbiomes of these tick species is species-specific. Earlier analyses of the microbial populations contained in the salivary glands of I. ovatus, I. persulcatus and H. flava ticks collected from the Shizuoka Prefecture of Japan indicated differential bacterial composition among these tick species [30]. However, whether the differences observed in that study and in our work are associated with both tick species and habitats requires further examination. In addition, this study revealed sex-specific differences in the bacterial communities present in the I. persulcatus and I. pavlovskyi ticks. Our data are consistent with results observed in analyses of the microbiome of A. americanum ticks [29].
Three prevalent bacterial genera—Acinetobacter, Rickettsia, and Francisella—accounted for 68% of the microbiomes of the studied tick species. The Acinetobacter bacteria are free-living aerobic saprophytes with oxidative metabolism. Some Acinetobacter species are typical representatives of nosocomial infections. These bacteria have been found in arthropods, including ticks [42]. 16S profiling has allowed for the detection of Acinetobacter in I. ricinus, I. ovatus, I. persulcatus and H. flava ticks in small amounts [27,30], whereas we detected these bacteria in all of the assayed pooled samples in considerably larger amounts from 8.8 to 74% of the microbiome. Other free-living aerobic saprophytes, including various members of the family Pseudomonadaceae, were also detected in all pooled samples examined, though in smaller amounts.
The share of bacteria belonging to the genus Rickettsia in the I. persulcatus and D. reticulatus microbiomes was large versus that of the I. pavlovskyi microbiome, in which Rickettsia spp. were fewer. It is unclear whether this is determined by a considerable variation in the rate of rickettsia-infected ticks in particular sampling sites or by different tropism of Rickettsia spp. to these tick species, including two closely related ixodes ticks species. Earlier investigations demonstrated that the occurrence of Rickettsia spp. sequences can be high in both I. persulcatus and D. reticulatus [11,31,43]. The 16S rRNA gene fragments used for amplification are highly conserved within the Rickettsia, which hinders their species-level identification. Nevertheless, this allowed for detection of DNA indicating the presence of the Rickettsiales bacterium “Montezuma”, a recently discovered bacterium, which has not been affiliated with any of the Rickettsiales families and forms a separate cluster in this order [44, 45]. Before this study, the Rickettsiales bacterium “Montezuma” had been found only in I. persulcatus ticks, whereas we for the first time detected it in I. pavlovskyi ticks.
The genus Francisella is composed of gram-negative intracellular parasites, in particular, the tularemia (F. tularensis) and septicemia (F. philomiragia) agents. The bacteria of this genus account for approximately half of the D. reticulatus microbiomes (both male and female) versus 0.1% of the microbiomes from the examined Ixodes tick species. Earlier investigations showed the presence of Francisella-like endosymbionts in D. reticulatus ticks [38] and a high infection rate of D. variabilis ticks with F. tularensis, though, this high rate of infection was inconsistent with the rare transmission of this disease via tick bites [46]. Meanwhile, the presence of Francisella spp. bacteria in the I. persulcatus, I. ovatis microbiomes was not demonstrated [30]. So, we could hypothesize that Francisella spp is a common member of microbiomes associated with different species of the Dermacentor genus.
The bacterial pathogens belonging to the genera Borrelia, Ehrlichia, and Anaplasma are known to be detectable in Ixodes ticks [1,2]. We also identified DNA sequences of the bacteria belonging to these genera in the pooled samples of I. persulcatus and I. pavlovskyi ticks with both B. burgdorferi s. l. and B. miyamotoi found in these two tick species. Recently, B. miyamotoi was isolated from I. pavlovskyi ticks inhabiting Hokkaido, Japan [47] and here we first report the presence of B. miyamotoi in I. pavlovskyi ticks in Siberia. These results indicated that I. pavlovskyi along with I. persulcatus could be a common tick vector for this pathogenic bacterium. No borrelias were detected in D. reticulatus microbiome, which is in agreement with some studies of these ticks in other regions [48]. Sometimes, borrelia DNA could be found in solitary D. reticulatus ticks, but this vector is not competent to transmit Borrelia spp. [14,49]. In our study, D. reticulatus ticks were collected in a sampling site where this tick species was dominated, while I. persulcatus and I. pavlovskyi ticks were rare species. Therefore, the possibility for D. reticulatus ticks to feed on the same reservoir host as ixodes ticks and acquire Borrelia spp. was low. DNA sequences of A. phagocytophilum were revealed in both Ixodes tick species as well. Notably, this is the first report of A. phagocytophilum and Candidatus Neoehrlichia mikurensis in I. pavlovskyi ticks. In D. reticulatus pooled samples, DNA sequences from bacteria belonging to the genus Ehrlichia were detected, which is the first discovery of these bacteria in this tick species. Further examinations are necessary to determine Ehrlichia genetic variants associated with D. reticulatus ticks.
In summary, 16S metagenomic profiling of the bacterial communities associated with I. persulcatus, I. pavlovskyi, and D. reticulatus ticks collected in the Novosibirsk region of Russia indicated that only several bacterial taxa with high abundance form the major share of microbiomes in all studied tick groups. This approach confirmed the results of long-term examination of bacterial pathogen occurrence in I. persulcatus ticks in Novosibirsk region and revealed that the spectrum of infectious agents found in the less-studied I. pavlovskyi and D. reticulatus ticks was wider than previously thought. Thus, this approach could be used for rapid assessment of the occurrence of medically important bacteria in tick populations inhabiting different natural biotopes and could therefore determine the epidemic danger of particular foci of infections transmitted by ticks.
Supporting Information
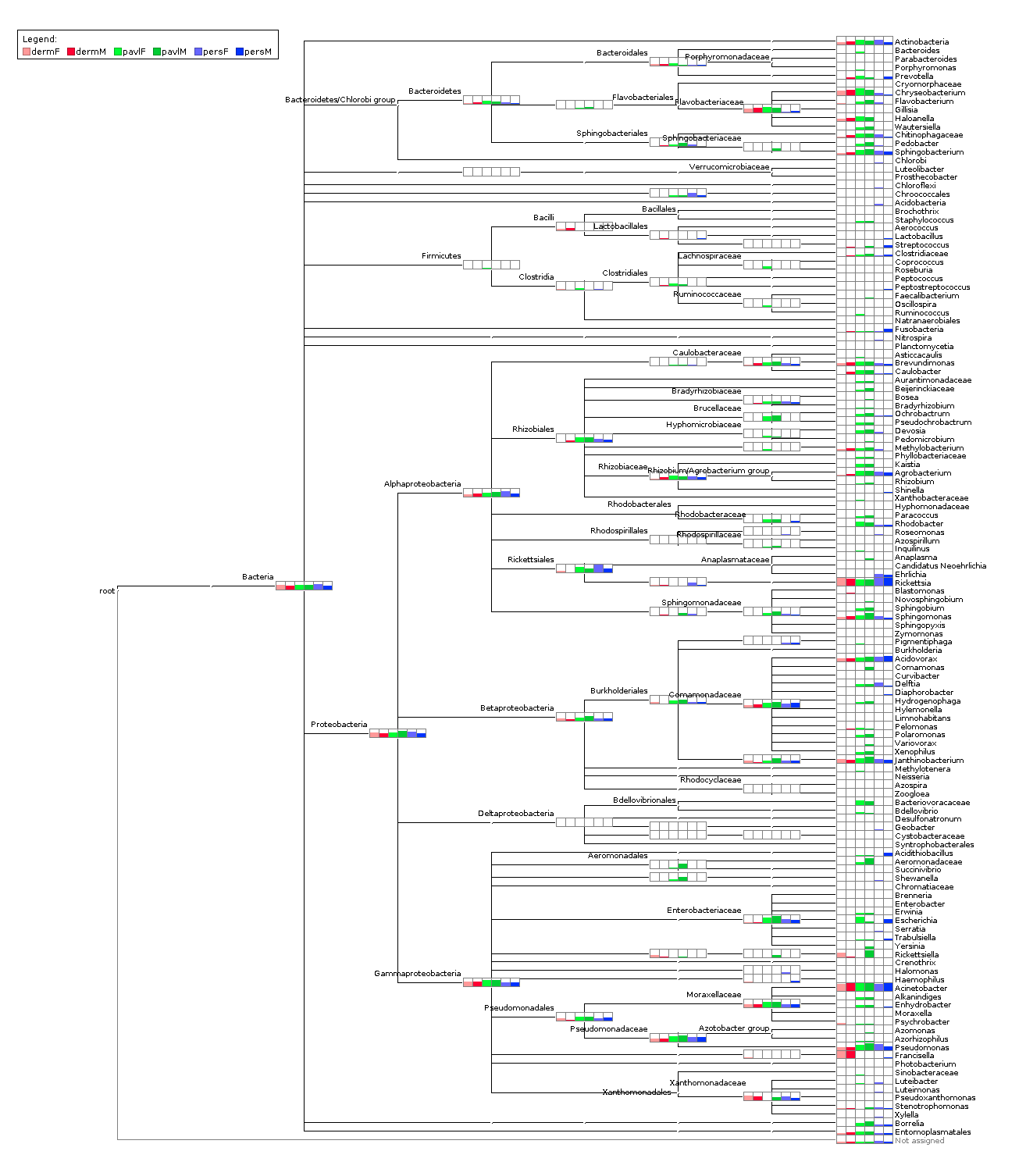
Samples in the tree were merged by tick species and gender. The height of the bars cohered to the log-scaled number of hits for each taxon.
(PNG)
{kind=link}
Each sheet in the file describes share of taxa for specific taxonomic level: genus, family, order, class. Taxonimic level is given on the sheet name.
(ZIP)
Data Availability
All sequence files are available from the NCBI database (accession number of Bioproject: PRJNA278801/SRP056314).
Funding Statement
This work was funded by the Russian Foundation for Basic Research (14-04-31549), http://www.rfbr.ru, and the Russian Ministry of Education and Science Projects (VI.55.1.1), http://mon.gov.ru. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Parola P, Raoult D (2001) Ticks and tickborne bacterial diseases in humans: an emerging infectious threat. Clin Infect Dis. 32: 897–928. [DOI] [PubMed] [Google Scholar]
- 2. Heyman P, Cochez C, Hofhuis A, van der Giessen J, Sprong H, Porter SR, et al. (2010) A clear and present danger: tick-borne diseases in Europe. Expert Rev Anti Infect Ther. 8: 33–50. 10.1586/eri.09.118 [DOI] [PubMed] [Google Scholar]
- 3. Dantas-Torres F, Chomel BB, Otranto D (2012) Ticks and tick-borne diseases: a One Health perspective. Trends Parasitol. 28: 437–446. 10.1016/j.pt.2012.07.003 [DOI] [PubMed] [Google Scholar]
- 4. Bakhvalova VN, Rar VA, Tkachev SE, Matveev VA, Matveev LE, Karavanov AS, et al. (2000) Tick-borne encephalitis virus strains of Western Siberia. Virus Res. 70:1–12. [DOI] [PubMed] [Google Scholar]
- 5. Mikryukova TP, Moskvitina NS, Kononova YV, Korobitsyn IG, Kartashov MY, Tyuten Kov OY, et al. (2014) Surveillance of tick-borne encephalitis virus in wild birds and ticks in Tomsk city and its suburbs (Western Siberia). Ticks Tick Borne Dis. 5: 145–151. 10.1016/j.ttbdis.2013.10.004 [DOI] [PubMed] [Google Scholar]
- 6. Tkachev S, Panov V, Dobler G, Tikunova N (2014) First detection of Kemerovo virus in Ixodes pavlovskyi and Ixodes persulcatus ticks collected in Novosibirsk region, Russia. Ticks Tick Borne Dis. 5: 494–496. 10.1016/j.ttbdis.2014.03.003 [DOI] [PubMed] [Google Scholar]
- 7. Beklemishev AB, Dobrotvorsky AK, Piterina AV, Ivanov ID, Nomokonova NY, Livanova NN (2003) Detection and typing of Borrelia burgdorferi sensu lato genospecies in Ixodes persulcatus ticks in West Siberia, Russia. FEMS Microbiol Lett. 227: 157–161. [DOI] [PubMed] [Google Scholar]
- 8. Fomenko NV, Livanova NN, Chernousova NY (2008) Diversity of Borrelia burgdorferi sensu lato in natural foci of Novosibirsk region. Int. J Med. Microbiol. 298(S1): 139–148. [Google Scholar]
- 9. Fomenko NV, Borgoiakov VIu, Panov VV (2011) Genetic features of Borrelia miyamotoi transmitted by Ixodes persulcatus . Mol Gen Mikrobiol Virusol. 2:12–17. [PubMed] [Google Scholar]
- 10. Shpynov S, Fournier PE, Rudakov N, Raoult D (2003) "Candidatus Rickettsia tarasevichiae" in Ixodes persulcatus ticks collected in Russia. Ann N Y Acad Sci. 990: 162–172. [DOI] [PubMed] [Google Scholar]
- 11. Shpynov S, Fournier PE, Rudakov N, Tarasevich I, Raoult D (2006) Detection of members of the genera Rickettsia, Anaplasma, and Ehrlichia in ticks collected in the Asiatic part of Russia. Ann N Y Acad Sci. 1078: 378–383. [DOI] [PubMed] [Google Scholar]
- 12. Rar VA, Livanova NN, Panov VV, Doroschenko EK, Pukhovskaya NM, Vysochina NP, et al. (2010) Genetic diversity of Anaplasma and Ehrlichia in the Asian part of Russia. Ticks Tick Borne Dis. 1: 57–65. 10.1016/j.ttbdis.2010.01.002 [DOI] [PubMed] [Google Scholar]
- 13. Morozova OV, Cabello FC, Dobrotvorsky AK (2004) Semi-nested PCR detection of Bartonella henselae in Ixodes persulcatus ticks from Western Siberia, Russia. Vector Borne Zoonotic Dis. 4: 306–309. [DOI] [PubMed] [Google Scholar]
- 14. Rar VA, Fomenko NV, Dobrotvorsky AK, Livanova NN, Rudakova SA, Fedorov EG, et al. (2005) Tickborne pathogen detection, Western Siberia, Russia. Emerg Infect Dis. 11: 1708–1715. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Korenberg EI, Nefedova VV, Romanenko VN, Gorelova NB (2010) The tick Ixodes pavlovskyi as a host of spirochetes pathogenic for humans and its possible role in the epizootiology and epidemiology of borrelioses. Vector Borne Zoonotic Dis. 10: 453–458. 10.1089/vbz.2009.0033 [DOI] [PubMed] [Google Scholar]
- 16. Chausov EV, Ternovoi VA, Protopopova EV, Kononova JV, Konovalova SN, Pershikova NL, et al. (2010) Variability of the tick-borne encephalitis virus genome in the 5' noncoding region derived from ticks Ixodes persulcatus and Ixodes pavlovskyi in Western Siberia. Vector Borne Zoonotic Dis. 10: 365–375. 10.1089/vbz.2009.0064 [DOI] [PubMed] [Google Scholar]
- 17. Romanenko VN (2005) The peculiarities of the biology of ticks inhabiting the environs of Tomsk City. Parazitologiia. 39: 365–370. [PubMed] [Google Scholar]
- 18. Livanova NN, Livanov SG, Panov VV (2011). Characteristics of the distribution of ticks Ixodes persulcatus and Ixodes pavlovskyi at the border between the forest and forest-steppe zones in the territory near Ob River. Parazitologiia. 45: 94–103. [PubMed] [Google Scholar]
- 19.Romanenko V, Leonovich S (2015) Long-term monitoring and population dynamics of ixodid ticks in Tomsk city (Western Siberia). Exp. Appl. Acarol. 10.1007/s10493-015-9879-2 [DOI] [PubMed]
- 20. Karan LS, Ciccozzi M, Yakimenko VV, Lo Presti A, Cella E, Zehender G, et al. (2014) The deduced evolution history of Omsk hemorrhagic fever virus. J Med Virol. 86: 1181–1187. 10.1002/jmv.23856 [DOI] [PubMed] [Google Scholar]
- 21. Mediannikov O, Matsumoto K, Samoylenko I, Drancourt M, Roux V, Rydkina E, et al. (2008) Rickettsia raoultii sp. nov., a spotted fever group rickettsia associated with Dermacentor ticks in Europe and Russia. Int J Syst Evol Microbiol. 58: 1635–1639. 10.1099/ijs.0.64952-0 [DOI] [PubMed] [Google Scholar]
- 22. Rar V, Epikhina T, Suntsova O, Kozlova I, Lisak O, Pukhovskaya NM, et al. (2014) Genetic variability of Babesia parasites in Haemaphysalis spp. and Ixodes persulcatus ticks in the Baikal region and Far East of Russia. Infect Genet Evol. 28: 270–275. 10.1016/j.meegid.2014.10.010 [DOI] [PubMed] [Google Scholar]
- 23. Hudson ME (2008) Sequencing breakthroughs for genomic ecology and evolutionary biology. Mol Ecol Resour. 8: 3–17. 10.1111/j.1471-8286.2007.02019.x [DOI] [PubMed] [Google Scholar]
- 24. Chakravorty S, Helb D, Burday M, Connell N, Alland D (2007) A detailed analysis of 16S ribosomal RNA gene segments for the diagnosis of pathogenic bacteria. J Microbiol Methods. 69: 330–339. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Carpi G, Cagnacci F, Wittekindt NE, Zhao F, Qi J, Tomsho LP, et al. (2011) Metagenomic profile of the bacterial communities associated with Ixodes ricinus ticks. PLoS One. 6:e25604 10.1371/journal.pone.0025604 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Nakao R, Abe T, Nijhof AM, Yamamoto S, Jongejan F, Ikemura T, et al. (2013) A novel approach, based on BLSOMs (Batch Learning Self-Organizing Maps), to the microbiome analysis of ticks. ISME J. 7: 1003–1015. 10.1038/ismej.2012.171 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Vayssier-Taussat M, Moutailler S, Michelet L, Devillers E, Bonnet S, Cheval J, et al. (2013) Next generation sequencing uncovers unexpected bacterial pathogens in ticks in western Europe. PLoS One. 8: e81439 10.1371/journal.pone.0081439 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Menchaca AC, Visi DK, Strey OF, Teel PD, Kalinowski K, Allen MS, et al. (2013) Preliminary assessment of microbiome changes following blood-feeding and survivorship in the Amblyomma americanum nymph-to-adult transition using semiconductor sequencing. PLoS One. 8: e67129 10.1371/journal.pone.0067129 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Williams-Newkirk AJ, Rowe LA, Mixson-Hayden TR, Dasch GA (2014) Characterization of the bacterial communities of life stages of free living lone star ticks (Amblyomma americanum). PLoS One. 9: e102130 10.1371/journal.pone.0102130 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Qiu Y, Nakao R, Ohnuma A, Kawamori F, Sugimoto C (2014) Microbial population analysis of the salivary glands of ticks; a possible strategy for the surveillance of bacterial pathogens. PLoS One. 9: e103961 10.1371/journal.pone.0103961 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Masella AP, Bartram AK, Truszkowski JM, Brown DG, Neufeld JD (2012) PANDAseq: paired-end assembler for illumina sequences. BMC Bioinformatics. 13: 31 10.1186/1471-2105-13-31 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Caporaso JG, Kuczynski J, Stombaugh J, Bittinger K, Bushman FD, Costello EK, et al. (2010) QIIME allows analysis of high-throughput community sequencing data. Nat Methods. 7: 335–336. 10.1038/nmeth.f.303 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Edgar RC (2010) Search and clustering orders of magnitude faster than BLAST. Bioinformatics. 26: 2460–2461. 10.1093/bioinformatics/btq461 [DOI] [PubMed] [Google Scholar]
- 34. Wang Q, Garrity GM, Tiedje JM, Cole JR (2007) Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl Environ Microbiol. 73: 5261–5267. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Oksanen J, Guillaume Blanchet F, Kindt R, Legendere P, Minchin PR, O’Hara RB, et al. (2013). vegan: Community Ecology Package. R package version 2.0–10.
- 36. Dray S, Dufour A (2007) The ade4 package: Implementing the duality diagram for ecologists. J Stat Software 22: 1–20. [Google Scholar]
- 37. Wang Y, Qian PY (2009) Conservative fragments in bacterial 16S rRNA genes and primer design for 16S ribosomal DNA amplicons in metagenomic studies. PLoS One 4: e7401 10.1371/journal.pone.0007401 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Michelet L, Bonnet S, Madani N, Moutailler S (2013) Discriminating Francisella tularensis and Francisella-like endosymbionts in Dermacentor reticulatus ticks: evaluation of current molecular techniques. Vet Microbiol. 163: 399–403. 10.1016/j.vetmic.2013.01.014 [DOI] [PubMed] [Google Scholar]
- 39. Cordaux R, Paces-Fessy M, Raimond M, Michel-Salzat A, Zimmer M, Bouchon D (2007) Molecular characterization and evolution of arthropod-pathogenic Rickettsiella bacteria. Appl Environ Microbiol. 73: 5045–5047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Beninati T, Lo N, Sacchi L, Genchi C, Noda H, Bandi C (2004) A novel alpha-proteobacterium resides in the mitochondria of ovarian cells of the ticks I. ricinus . Appl Environ Microbiol. 70: 2596–2602. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Rymaszewska A. (2007) Symbiotic bacteria in oocyte and ovarian cell mitochondria of the tick Ixodes ricinus: biology and phylogenetic position. Parasitol Res. 100: 917–920. [DOI] [PubMed] [Google Scholar]
- 42. Murrell A, Dobson SJ, Yang X, Lacey E, Barker SC (2003) A survey of bacterial diversity in ticks, lice and fleas from Australia. Parasitol Res. 89: 326–334. [DOI] [PubMed] [Google Scholar]
- 43. Alberdi MP, Nijhof AM, Jongejan F, Bell-Sakyi L (2012) Tick cell culture isolation and growth of Rickettsia raoultii from Dutch Dermacentor reticulatus ticks. Ticks Tick Borne Dis. 3(5–6): 349–354. 10.1016/j.ttbdis.2012.10.020 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Mediannikov OIu, Ivanov LI, Nishikawa M, Saito R, Sidel'nikov IuN, Zdanovskaia NI, et al. (2004) Microorganism "Montezuma" of the order Rickettsiales: the potential causative agent of tick-borne disease in the Far East of Russia. Zh Mikrobiol Epidemiol Immunobiol. 1: 7–13. [PubMed] [Google Scholar]
- 45. Eremeeva ME, Oliveira A, Moriarity J, Robinson JB, Tokarevich NK, Antyukova LP, et al. (2007) Detection and identification of bacterial agents in Ixodes persulcatus Schulze ticks from the north western region of Russia. Vector Borne Zoonotic Dis. 7: 426–436. [DOI] [PubMed] [Google Scholar]
- 46. Goethert HK, Telford SR 3rd (2010) Quantum of infection of Francisella tularensis tularensis in host-seeking Dermacentor variabilis . Ticks Tick Borne Dis. 1: 66–68. 10.1016/j.ttbdis.2010.01.001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Takano A, Toymane K, Konnai S, Ohashi K, Nakao M, Ito T, et al. (2014) Tick surveillance for relapsing fever spirochete Borrelia miyamotoi in Hokkaido, Japan. PLoS One 9: e104532 10.1371/journal.pone.0104532 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48. Richter D, Kohn C, Matuschka FR (2013) Absence of Borrelia spp., Candidatus Neoehrlichia mikurensis, and Anaplasma phagocytophilum in questing adult Dermacentor reticulatus ticks. Parasitol Res. 112: 107–111. 10.1007/s00436-012-3110-8 [DOI] [PubMed] [Google Scholar]
- 49. Lledó L, Gegúndez MI, Giménez-Pardo C, Álamo R, Fernández-Soto P, Nuncio MS, et al. (2014) A seventeen-year epidemiological surveillance study of Borrelia burgdorferi infections in two provinces of northern Spain. Int J Environ Res Public Health. 11: 1661–1672. 10.3390/ijerph110201661 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Samples in the tree were merged by tick species and gender. The height of the bars cohered to the log-scaled number of hits for each taxon.
(PNG)
Each sheet in the file describes share of taxa for specific taxonomic level: genus, family, order, class. Taxonimic level is given on the sheet name.
(ZIP)
Data Availability Statement
All sequence files are available from the NCBI database (accession number of Bioproject: PRJNA278801/SRP056314).