

Abstract
When attending for impending visual stimuli, cognitive systems prepare to identify relevant information while ignoring irrelevant, potentially distracting input. Recent work (Marini et al., 2013) showed that a supramodal distracter-filtering mechanism is invoked in blocked designs involving expectation of possible distracter stimuli, although this entails a cost (distraction-filtering cost) on speeded performance when distracters are expected but not presented. Here we used an arrow-flanker task to study whether an analogous cost, potentially reflecting the recruitment of a specific distraction-filtering mechanism, occurs dynamically when potential distraction is cued trial-to-trial (cued distracter-expectation cost). In order to promote the maximal utilization of cue information by participants, in some experimental conditions the cue also signaled the possibility of earning a monetary reward for fast and accurate performance. This design also allowed us to investigate the interplay between anticipation for distracters and anticipation of reward, which is known to engender attentional preparation. Only in reward contexts did participants show a cued distracter-expectation cost, which was larger with higher reward prospect and when anticipation for both distracters and reward were manipulated trial-to-trial. Thus, these results indicate that reward prospect interacts with the distracter expectation during trial-by-trial preparatory processes for potential distraction. These findings highlight how reward guides cue-driven attentional preparation.
Keywords: distraction-filtering cost, cued distracter-expectation cost, reward prospect, attentional preparation, flanker task, cueing paradigm
1. Introduction
The guidance of appropriate and efficient human behavior relies on both the selection of relevant information and the inhibition of irrelevant information from the environment. Attentional control mechanisms guide these selection and inhibition processes. Both the selection of relevant information and the suppression of irrelevant, distracting stimuli are exposed to moment-to-moment changes in the environment. Attentional mechanisms, such as proactive regulation of current goals and priorities, are invoked to help withstand these potentially distracting changes, while momentary lapses in such proactive attentional mechanisms typically are promptly recovered from by reactive enhancements in attentional control (e.g., Weissman, Roberts, Visscher, & Woldorff, 2006; Walsh, Buonocore, Carter, & Mangun, 2011; Braver, Paxton, Locke, & Barch, 2009). The capability of proactively preparing for, and reactively adjusting to, the multifaceted inputs that arise from our changing environment is a key feature of attentional control. The neurocognitive study of attention has revealed physiological and psychological processes that intervene with both proactive and reactive dynamics for the prevention of distraction and for the suppression of distracting elements during visual tasks (see Geng, 2014, for a review). The proactive suppression of distracters might occur when the observer has some prior information about the characteristics of impending distracter stimuli, such as the spatial location where distracters might be presented, or their color. In such circumstances, control mechanisms may be able to pre-emptively orient attention toward relevant locations or features and away from those of distracter stimuli.
Cueing paradigms are particularly suitable for the study of attentional preparation, including for the possibility of distracter stimuli. In a previous study on sensory-related modulation of distracter suppression, a visual cue was shown prior to a visual target for predicting both the presence (or absence) and the spatial location of a potential visual distracter (Ruff & Driver, 2006). Other studies have used cues for predicting the probability of distracting elements being presented in the visual stimulus display (Awh, Matsukura, & Serences, 2003; Serences, Yantis, Culberson, & Awh, 2004). Moreover, cueing paradigms allow making inferences about preparatory processes while disentangling cue-driven processing from responses evoked by probe stimuli (e.g., van den Berg, Krebs, Lorist, & Woldorff, 2014; Schevernels, Krebs, Santens, Woldorff, & Boehler, 2014).
Although cueing paradigms afford some useful advantages, a different experimental paradigm has been also used for creating an expectation for potential distraction. In fact, a recent multisensory attention study (Marini, Chelazzi, & Maravita, 2013) used a novel approach, namely the “Distraction context manipulation paradigm,” for studying mechanisms for dealing with possible impending distractions. In this paradigm, rather than using a predictive cue on each trial, the expectation for distraction was created at the block level by using two different types of experimental blocks. One type of block had an expectation of distraction because both distracter-absent and distracter-present trials were intermixed (the Mixed block), while the other type of block included only distracter-absent trials (the Pure block) and therefore engendered no expectation for distracters. The critical comparison in the Distraction Context Manipulation paradigm was between distracter-absent trials in the Mixed block (i.e., the block with occasional distracters) and distracter-absent trials in the Pure block (i.e., the completely distracter-free block). This comparison revealed that responses to distracter-absent trials in the Mixed blocks incurred a cost in response times (RT) compared to the physically identical distracter-absent trials in the Pure block. This cost, termed the distraction-filtering cost, was inversely correlated to the behavioral cost on RTs (incongruent versus congruent trials) caused by distracter stimuli in distracter-present trials, and likely reflects the proactive engagement of a distraction-filtering mechanism that is invoked in potentially distracting contexts in order to limit the negative impact of distraction.
In the present study, we aimed at further exploring this distraction-filtering mechanism by investigating whether a similar distraction-filtering cost (hereinafter, the cued distracter-expectation cost) can also be invoked in more dynamic situations by using a cued variant of the arrow flanker task (Eriksen & Eriksen, 1974; see also Experiment 6 in Marini et al., 2013) in which we modulated the expectation for distraction on a trial-by-trial basis. On every trial, the stimulus display was preceded by the presentation of an informative cue that predicted either the absence (with 100% validity) or the possible presence (with 75% validity) of distracting stimuli in the upcoming stimulus display. Because of such experimental design, 25% of stimuli following a distraction-predicting cue were actually distracter-absent trials, although preparatory processes for dealing with the possibility of distracting stimuli would have been invoked on these trials. In a similar vein to the aforementioned blocked-paradigm study (Marini et al., 2013), in this cueing version of the paradigm the critical comparison for evidencing the behavioral cost of the distraction-filtering mechanism was contrasting the distracter-absent trials preceded by a cue predicting no distracters versus the distracter-absent trials preceded by a cue predicting distracters. In the latter situation, akin to distracter-absent trials in the Mixed block of Marini et al.'s study, preparatory mechanisms for dealing with expected distraction could manifest as a cost on performance when the anticipated distraction does not occur (“cued distracter-expectation cost”).
It is worth noting that the dynamic invoking of a distraction-filtering mechanism on a trial-by-trial basis is probably much more cognitively demanding than its strategic and sustained activation throughout an experimental block. Therefore, it is also possible that the relatively costly distraction-filtering mechanism would not be dynamically invoked unless an extra incentive is provided for promoting the optimization of attentional preparation. Such an incentive might be provided by incorporating the possibility of a monetary reward (Della Libera & Chelazzi, 2006; Seifert, Naumann, Hewig, Hagemann & Bartussek, 2006; Hubner & Schlosser, 2010).
Attention and reward are intimately related through different underlying behavioral and neural mechanisms (see Chelazzi, Perlato, Santandrea & Della Libera, 2013, for a review). The level of interplay between reward and attention that closely relates to the present study derives from the mobilization of cognitive and attentional resources in the effort of maximizing the outcome of the current task (e.g., Locke & Braver, 2008). The incentive role of reward and its effects on behavior can be achieved through modulations of attentional and cognitive control. These modulations include enhancing attentional preparation under conditions of reward prospect (Van den Berg et al., 2014), enhanced recruitment of cognitive resources (Krebs, Boehler, Roberts, Song, & Woldorff, 2012; Vassena, Silvetti, Boehler, Achten, Fias et al., 2014), reduction of conflict (Padmala & Pessoa, 2011; Krebs, Boehler, & Woldorff, 2010), stopping-induced stimulus devaluation (Wessel, O'Doherty, Berkebile, Linderman & Aron, 2014), reactive response inhibition (Boehler, Schevernels, Hopf, Stoppel, & Krebs, 2014), and optimized memory encoding (Marini, Marzi, & Viggiano, 2011). Moreover, the Eriksen flanker task in association with monetary rewards has been already used in previous studies of selective attention (Seifert et al. 2006; Hubner & Schlosser, 2010), cognitive control (Braem, Hickey, Duthoo & Notebaert, 2014) and conflict adaptation (van Steenbergen, Band & Hommel, 2009; Braem, Vertguts, Roggeman & Notebaert, 2012), suggesting the usefulness of this paradigm for the goals of the present study.
Given that reward prospect can optimize cognitive control and preparatory attention through several different mechanisms, we hypothesized that it might also help optimize the dynamic implementation of a cognitive set for distraction-filtering. Accordingly, we predicted that in our study the prospect of a monetary incentive for optimal (i.e., quick and accurate) performance might better promote the recruitment of the distraction-filtering mechanism in a dynamic trial-by-trial modulation of distracter expectation. Thus, when the cue predicted that a distracter was likely to be present, we expected to find more of a performance cost on trials when no distracter actually occurred and the cue also indicated reward prospect compared to when it indicated no (or low) reward prospect. This prediction is supported by recent work showing that reward-prospect cues modulated brain activity associated with attentional preparation for discriminating a Stroop color-word target (Van den Berg et al., 2014). In Van den Berg et al.'s study, differences in individual electrophysiological responses to reward-prospect cues were associated with differences in interference in the Stroop task, indicating that participants were able to utilize cue information for implementing efficient attentional control. Similarly, in the present study we predicted that participants would utilize cue information for invoking a distraction-filtering mechanism in a dynamic fashion when the cue was paired with the prospect of reward.
2. Methods
2.1 Participants
The study included two experiments involving a total of thirty-three participants, all with normal or corrected-to-normal vision and none with any known neurological condition. All participants gave written informed consent in accordance with protocols approved by the Duke Medical Center Institutional Review Board. Participants were compensated for their time by being paid 15 US dollars/hour, plus any reward they earned as a function of task performance (see Experimental Design).
Separate pools of subjects participated in the two experiments. Sixteen subjects participated in Experiment 1 (mean age 22.1, range 18-35, 8 females, 16 right-handed), and seventeen in Experiment 2 (mean age 22.7, range 18-35, 8 females, 17 right-handed). One participant in Experiment 2 was excluded from analysis because of very low response accuracy (i.e. overall accuracy 3 standard deviations below the group mean).
2.2 Experimental design
The two experiments differed in how Reward and No-Reward conditions were implemented in the experimental paradigm. In Experiment 1 all the trials carried reward prospect, but they were divided between low-reward and high-reward trials that were randomly intermingled. Experiment 2 included a reward context in which Reward trials and No-Reward trials were randomly intermixed within the same experimental block; in addition, in separate blocks participants also performed in a no-reward context that included only No-Reward trials.
For the entire duration of each experiment participants sat comfortably in a dimly illuminated room at a distance of 57 cm from the central point of a 24″ computer screen (Asus VG248QE, 1920 × 1080 pixels, refresh rate 120Hz). The experimental paradigm was programmed in Matlab (Mathworks Inc.) with Psychtoolbox 3.0 (Kleiner et al., 2007). Participants' responses were collected through button presses on a gamepad (Logitech Precision G-UG15). Participants received written task instructions and performed 20 practice trials prior to the beginning of the experiment.
Stimuli without distracters consisted of a single centrally presented arrow that was pointing either up or down. Stimuli with distracters consisted of a horizontal array of five flanking arrows that were pointing either up or down (Eriksen & Eriksen, 1974). The arrows were presented in black on a medium-gray background immediately below a central fixation cross which stayed visible throughout the experiment. Each arrow subtended a visual angle of 0.75 by 0.5 degrees, and the center-to-center distance between adjacent arrows was 0.75 degrees. The central arrow was always the target, whether presented alone or with flanking arrows; for the trials with flanking stimuli, the four flanking arrows (i.e., two on each side of the target) were the distracters. On congruent trials, target and distracters pointed in the same direction (either up or down), while on incongruent trials target and distracters pointed in opposite directions (either target up and distracter down or vice-versa). Participants were instructed to respond as quickly and as accurately as possible by pressing the gamepad button corresponding to the direction of the central target arrow while ignoring the direction of any distracters if present.
In both experiments, prior to the visual presentation of each stimulus, the participants were presented with a visual cue indicating whether a distracter might be presented or would definitely not be presented in the upcoming stimulus display. Participants were informed as to the meaning of the different cues at the beginning of the experiment. A pause screen, which was shown between blocks, reiterated cue meanings and, in Experiment 2, reported the type of the upcoming block (“possible reward” or “no reward”). The cue was a letter, whose identity indicated the potential for distraction and whose color indicated the presence or absence of reward possibility. An “X” indicated that no distracter was going to be presented while a “D” (for “distracter”) indicated that a distracter (either congruent or incongruent) would likely appear. The “X” cue had 100% validity, meaning that after an “X” distracter-absent stimuli were always presented. The “D” cue had 75% validity and thus the cue was followed by a distracter-absent stimulus display in 25% of these trials. The remaining trials in this group were divided between those with incongruent distracters (50%) and those with congruent distracters (25%). The cue letter was colored (blue/yellow, randomly assigned across participants), thereby providing information about possible reward in the current trial. Specifically, in Experiment 1 the cue color indicated the amount of potential reward (low/high); in Experiment 2 the cue color indicated whether the trial was associated with a potential reward or not (reward/no-reward). The stimulus display appeared 1500 ms after cue onset and stayed onscreen for 200 ms. After stimulus offset, participants had 1000 ms to make their response. A feedback stimulus, which was displayed for 200 ms, followed each reward-prospect trial with no delay and reported the earned reward. The experimental design for both experiments is schematically represented in Figure 1.
Figure 1. Experimental paradigm.
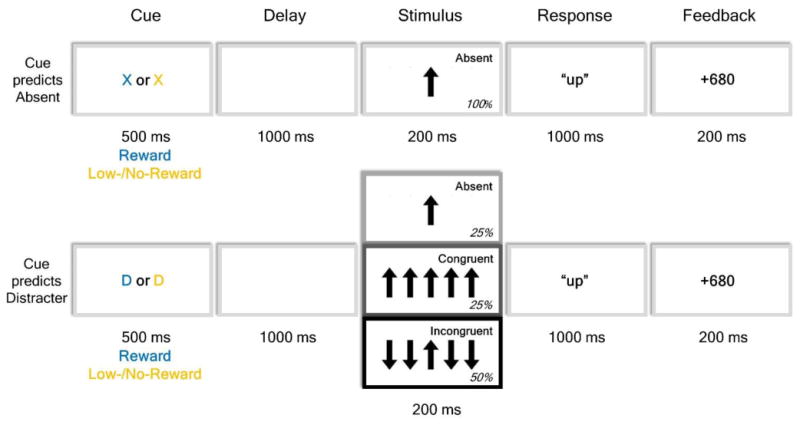
Schematic representation of the experimental paradigm. Each trial started with a letter cue whose identity predicted the possibility of distraction and whose color conveyed whether there was a prospect of reward. After a delay, an arrow flanker array was presented and participants indicated the direction of the central arrow. At the end of reward-prospect trials, a feedback stimulus showed the earned reward. In Experiment 1, the cue colors changed from trial to trial and indicated whether there was prospect of a low-value versus a high-value reward. In Experiment 2, the cue-colors also changed trial-wise in reward-context blocks, but indicated whether there was reward prospect or no reward prospect on that trial, and in addition no-reward context blocks were also included.
Participants earned reward points only for correct responses, and the actual number of points was calculated with the following exponentially decreasing function1:
The parameters m, a, ε and k were fixed and assumed the following values: m=2954, a=2.2, n=5, ε=46, k=1 (Experiment 1, high-reward condition; Experiment 2) or k=4 (Experiment 1, low-reward condition) and the independent variable x expressed reaction time in seconds (range 0.2 – 1.2). This method provided an incentive to maximize both response speed and accuracy. At the end of each block, points were converted into real money according to a fixed conversion rate (180000 points equated to 1 US dollar), and participants were informed about the amount they had earned so far. At the end of the experiment participants were paid, in addition to their hourly payment, the additional reward-based amount they had earned. The average reward-based addition across participants was 5.70 US dollars. The duration of Experiment 1 was about 50 minutes and the duration of Experiment 2 was about 75 minutes, with breaks given in between the experimental blocks.
2.3 Analysis
Trials were filtered to eliminate those with response time (RT) outliers, which were defined as responses faster than 200 ms (anticipatory responses) or exceeding the mean RT value for that subject and experimental condition plus three standard deviations (delayed responses) (Ratcliff, 1993). Accuracy and RT values were used to calculate the inverse efficiency (IE) score (Townsend & Ashby, 1983). Inverse efficiency is defined as the ratio between the mean RT and the proportion of correct responses, thereby allowing adjusting for speed-accuracy trade-offs. In tasks where both speed and accuracy index performance, inverse efficiency has been widely used in the domain of distraction filtering (Marini et al., 2013) as well as in studies of visual attention (e.g., Austen & Enns, 2003) and crossmodal perception (e.g., Heed, Habetz, Sebanz, & Knoblich, 2010; Marini, Tagliabue, Sposito, Hernandez-Arieta et al., 2014). IE values are actually identical to RTs when accuracy is at 100% and increase proportionally with increases in the error rate (e.g., a mean RT of 600 ms combined with an accuracy of 90% would give an IE = 600/.90 = 667 msa, where here we use the subscript “a” to show that the unit form of IE is an adjusted ms value). Because RT and accuracy showed fully compatible patterns, in order to achieve maximal statistical power we focused on the IE results. Nonetheless, we also conducted full statistical analyses on the RTs. The RT analyses yielded the same statistical results as the IE analyses, with only one exception, which we report in a footnote. Comprehensive RT, accuracy, and IE values are reported in Table 1A for the distracter-absent trials (from which the cued distracter-expectation cost is calculated) and in Table 1B for the distracter-present trials (from which the Flanker incongruency effect is calculated). Statistical comparisons were conducted by means of planned comparisons with paired-sample t-tests and by means of repeated-measure analyses of variance (ANOVA). When significant effects and interactions emerged in the ANOVAs, they were further explored with pairwise comparisons (t-tests). The family-wise error rate was controlled by applying the Holm-Bonferroni method (Holm, 1979). Analyses were performed with Statistica for Windows release 6.0 (StatSoft Italia SRL) and with the R software package (R Development Core Team, 2013).
Table 1A. Overall results for distracter-absent trials.
|
Inverse efficiency (msa) mean (SD) |
Reaction time (ms)
mean (SD) |
Accuracy (%) mean (SD) |
||||
|---|---|---|---|---|---|---|
|
|
||||||
| Cue predicts no distracter | Cue predicts distracter | Cue predicts no distracter | Cue predicts distracter | Cue predicts no distracter | Cue predicts distracter | |
| Experiment 1 | ||||||
|
|
||||||
| Low Reward | 386.4 (52) | 393.5 (55) | 369.6 (49) | 377.6 (50) | 95.7 (3) | 96 (2.6) |
| High Reward | 380.7 (38) | 398.2 (52) | 363.6 (38) | 375.5 (44) | 95.5 (3) | 94.3 (4.5) |
| Experiment 2 | ||||||
|
|
||||||
| No-Rew Context | 474.1 (68) | 478.6 (76) | 438.1 (63) | 443.9 (64) | 92.6 (6.3) | 93.2 (6.8) |
| Rew-Context No-Reward | 446.5 (45) | 456.7 (58) | 418.7 (45) | 423.7 (52) | 93.8 (4.8) | 93.1 (7) |
| Rew-Context Reward | 405.6 (43) | 424 (42) | 386.4 (43) | 397.6 (45) | 95.3 (3.2) | 93.8 (4.8) |
| Table 1B. Overall results for distracter-present trials. | ||||||
|---|---|---|---|---|---|---|
|
Inverse efficiency (msa) mean (SD) |
Reaction time (ms)
mean (SD) |
Accuracy (%) mean (SD) |
||||
|
| ||||||
| Congruent distracter | Incongruent distracter | Congruent distracter | Incongruent distracter | Congruent distracter | Incongruent distracter | |
| Experiment 1 | ||||||
|
|
||||||
| Low Reward | 391.3 (58) | 527.4 (56) | 381.3 (52) | 433.9 (51) | 97.4 (2.6) | 82.3 (10) |
| High Reward | 387 (47) | 543.2 (61) | 376.2 (48) | 430 (53) | 97.2 (3.1) | 79.2 (10.1) |
| Experiment 2 | ||||||
|
|
||||||
| No-Rew Context | 467.3 (65) | 591.5 (57) | 445.7 (60) | 498.8 (59) | 95.5 (4.6) | 84.6 (9.5) |
| Rew-Context No-Reward | 452.3 (51) | 579.5 (64) | 428.8 (53) | 478.7 (49) | 94.9 (6) | 83.3 (10.1) |
| Rew-Context Reward | 414 (42) | 554.9 (68) | 396.1 (42) | 447.8 (47) | 95.7 (3.7) | 81.3 (8.3) |
3. Results
3.3 Experiment 1 (low vs. high reward)
In Experiment 1, we investigated whether a “cued distracter-expectation cost” (i.e., slower responses in distracter-absent trials following a cue predicting probable distraction compared to a cue predicting no distracters), potentially related to the implementation of the distraction-filtering mechanism, was observed with a double cue indicating both the possibility of distracters and the prospect of reward in the next trial. A recent study showed that reward influences cognitive control by enhancing contextual features that lead to a rewarding outcome (Braem et al., 2014). Accordingly, we hypothesized that cueing the prospect of a different amount of reward on every trial (low versus high) might favor the utilization of distracter-predictive cue information and thereby provide an incentive to implement a distraction-filtering mechanism, especially on the high-reward trials.
The first ANOVA was focused on the comparison of distracter-absent trials under different cue and reward conditions. This ANOVA had a 2-by-2 within-subjects design factoring Reward (low, high) and Cued distracter-expectation (cue predicting distracter-absent, cue predicting distracter-present). This analysis revealed a main effect of Cued distracter-expectation [F(1,15)=5.83, p<.05], with slower performance in distracter-absent trials that were preceded by a cue predicting the likely occurrence of a distracter (mean IE ± SD: 396 ± 53 msa) compared to distracter-absent trials for which the cue predicted there would not be any distracters (mean IE ± SD: 384 ± 45 msa) (Figure 2, left graph). This cued distracter-expectation cost is fully compatible with the distraction-filtering cost found in previous work using blocked manipulation of possible distracters (Marini et al., 2013). On the other hand, there was no significant main effect of Reward level [F(1,15)=.02, p=.87]. A significant interaction between Reward and Cued distracter-expectation was observed, however ([F(1,15)=6.56, p<.05], Figure 2, left graph), due to the cued distracter-expectation cost being significantly larger2 in the high-reward condition (mean IE cost: 18 msa) [t(15)=2.78, p<.01, corrected-α=.025] than in the low-reward condition (mean IE cost: 7 msa [t(15)=1.56, p=.06, corrected-α=.05]).
Figure 2. Results of the cued distracter-expectation cost analysis.
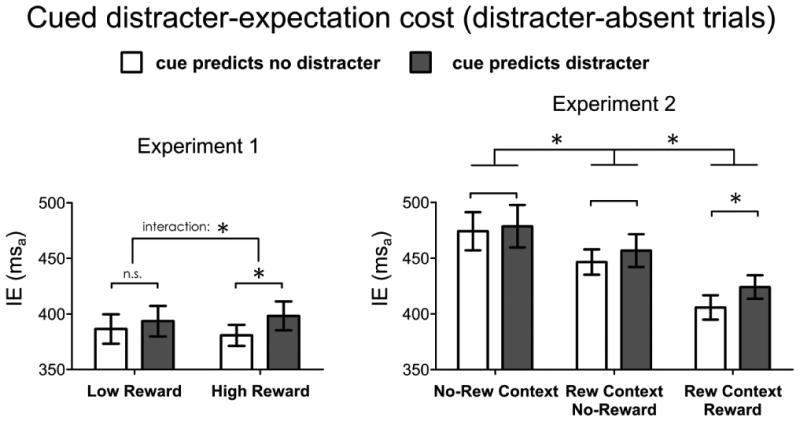
Average inverse efficiency values (IE) and standard errors of the mean in the two experiments, for the comparison of the behavioral responses on distracter-absent trials that were preceded by a cue predicting no distracter (white bars) or by a cue predicting potential distracter (gray bars). Results are showed separately for each one of the reward-related conditions within each experiment. A higher cued distracter-expectation cost, likely indicating the preparatory effort to implement distraction-filtering, was observed in the mostly rewarding conditions (high-reward condition in Experiment 1, reward trials in the reward context in Experiment 2). Additionally, reward-context in Experiment 2 improved performance on distracter-absent trials, relative to the no-reward context. This was observed in the reward context even in absence of reward in the current trial, although the improvement was larger when reward prospect was specifically indicated for the current trial.
For the distracter-present trials, we performed an ANOVA to examine the conflict-related effects in the different reward-prospect conditions by comparing IEs in congruent and incongruent trials as a function of reward level. This ANOVA had a 2-by-2 within-subjects design, with factors of Reward (low, high) and Distracter Type (congruent, incongruent). As expected, there was a main effect of conflict, with congruent-distracter stimuli eliciting faster responses (mean IE ± SD: 389 ± 52 msa) than did incongruent-distracter stimuli (mean IE ± SD: 535 ± 58 msa) [F(1,15)=73.7, p<.001] (Figure 3, left graph). We observed no significant main effect of Reward in this analysis [F(1,15)=3.95, p=.07], as well as no significant interaction between the Reward and Congruency factors [F(1,15)=3.92, p=.07]. Thus, as in the flanker study by Hubner & Schlosser (2010) and in the cued Stroop study by Van den Berg et al. (2014), we observed that cued reward prospect did not lead to any overall reduction of distracter interference across subjects (Van den Berg et al., 2014; but see Padmala & Pessoa, 2011).
Figure 3. Results of the flanker incongruency effect analysis.
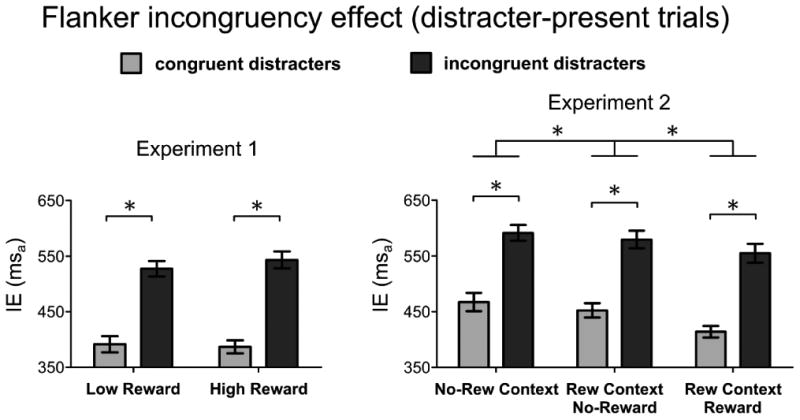
Average inverse efficiency values (IE) and standard errors of the mean in the two experiments, for the comparison of the behavioral responses on distracter-present trials that presented congruent distracter stimuli (light gray bars) versus incongruent distracter stimuli (dark gray bars). Results are showed separately for each one of the reward-related conditions within each experiment. In both experiments, congruent trials elicited better performance overall compared to incongruent trials. The magnitude of the incongruency effect was not modulated by the reward condition in either experiment. In Experiment 2, however, no-reward trials (congruent and incongruent together) of the reward-context had better performance compared to the same trials in the no-reward context, and reward trials (congruent and incongruent together) had better performance compared to the no-reward trials (congruent and incongruent together) of both the reward-context and the no-reward context.
Experiment 1 thus showed that reward-prospect, when cued on a trial-by-trial basis, effectively invokes attentional preparatory processes that seem similar to those observed at the block level (Marini et al., 2013), as indicated by the incursion of a cued distracter-expectation cost. Additionally, the magnitude of the cued distracter-expectation cost was directly related to the amount of reward, with this cost being larger when the monetary incentive was higher. Such a results suggests that the higher the reward, the stronger the attentional preparation.
3.4 Experiment 2 (reward-context blocks [with both reward and no-reward trials], and no-reward blocks)
In Experiment 2, we investigated: (i) whether distraction-informative cues delivered with no reward are sufficient to trigger the cued distracter-expectation cost observed in Experiment 1; and (ii) whether the cued distracter-expectation cost is observed when the presence of reward is at the blocked/context level, or alternatively occurs only when the possibility of reward is indicated in a given trial. Experiment 2 comprised both no-reward and reward context conditions, run in separate experimental blocks: in some blocks there was no reward nor reward prospect (no-reward-context blocks), and in the other blocks no-reward and reward trials were intermixed (reward-context blocks). Thus, in the no-reward context blocks, the cue signaled only the potential for distracters (present or absent) and therefore its color was unchanging trial-to-trial. In contrast, in the reward-context blocks there was a double cue on each trial (similar to Experiment 1) that signaled both the potential for distraction (present or absent, depending on cue letter) and the potential for reward (present or absent, depending on cue color).
For investigating preparatory effects under different conditions of reward, we conducted a repeated-measure ANOVA on distracter-absent trials with the factors of Reward (no-reward-context and the conditions in the reward context: no-reward, reward) and Cued distracter-expectation (predicting distracter-absent, predicting distracter-present). We identified significant main effects for both Reward [F(2,30)=15.02, p<.001] and Cued-distracter-expectation [F(1,15)=6.95, p<.05] (Figure 2, right graph). More specifically, in the distracter-absent condition participants were fastest on reward trials in the reward-context blocks (mean IE ± SD: 415 ± 42 msa), next fastest on no-reward trials in the reward-context blocks (mean IE ± SD: 451 ± 50 msa), and slowest on the no-reward trials in the no-reward-context blocks (mean IE ± SD: 476 ± 71 msa). This effect of reward context on no-reward trials constitutes a previously unidentified result that suggests that reward-related effects might entail a strategic component that favors no-reward trials occurring in a reward context over the same trials in a no-reward context.
For assessing the presence of a cued distracter-expectation cost in the different distracter-absent conditions, we conducted planned pairwise comparisons (corrected for the family-wise error rate with the Holm-Bonferroni method) between cue-predicts-absent-distracter and cue-predicts-present-distracter trials for each of the reward conditions (in the no-reward context condition, and in the no-reward and reward conditions occurring in the reward-context blocks). Results revealed that only in the reward condition was a significant cued distracter-expectation cost observed [t(15)=4.39, p<.001, corrected-α=.0167], whereas the tendency for a cued distracter-expectation cost was not significant in the no-reward-context blocks, nor in the no-reward condition in the reward-context blocks [t(15)=0.57, p=.29, corrected-α=.05, and t(15)=1.6, p=.07, corrected-α=.025, respectively] (Figure 2, right graph). Moreover, we conducted a repeated-measures ANOVA within the reward-context blocks, with the factors of Cued-Reward (no-reward, reward) and Cued distracter-expectation (predicting distracter-absent, predicting distracter-present). This analysis revealed a main effect of Reward [F(1,15)=15.8, p<0.01], with better performance on reward versus no-reward trials (mean IEs ± SD: 415 ± 42 msa and 451 ± 50, respectively), and a main effect of Cued distracter-expectation [F(1,15)=9.3, p<0.01], with slower responses after cues predicting distraction compared to cues predicting no-distraction (mean IEs ± SD: 426 ± 43 msa and 440 ± 46, respectively). The interaction between Reward and Cued distracter-expectation was not significant [F(1,15)=2.31, p=0.15].
Conflict-related interference effects were explored with a separate ANOVA on distracter-present trials, with the factors of Reward (no-reward context, and conditions in reward context: no-reward, reward) and Distracter Type (congruent, incongruent). As with the distracter-absent trials described above, performance on the distracter-present trials was fastest in the Reward condition in the reward-context blocks, intermediate in the No-Reward condition in the reward-context blocks, and slowest in the no-reward context blocks (average IEs ± SD: 484 ± 44 msa, 516 ± 46 msa, and 529 ± 53 msa, respectively) [F(2,30)=14.77, p<.001]. A significant main effect of conflict was also observed, with incongruent distracters eliciting slower responses than congruent distracters [F(1,15)=75.8, p<.001] (see Figure 3, right graph). No interactions of these factors emerged, however. Subsequent pairwise comparisons revealed that, similarly to Experiment 1, as well as several recent related studies (Hubner & Schlosser, 2010; van den Berg et al., 2014), the magnitude of the incongruency cost on behavioral performance was not reduced by the prospect of a monetary reward (all p-values > 0.05).
In order to test our prediction that the utilization of the reward cue promoted attentional preparation for potential distraction, we conducted an across-subject correlation analysis in the reward-context blocks between the reward effect (no-reward IEs minus reward trials IEs) and the cued distracter-expectation cost. We did find such a correlation in the distracter-absent trials of Experiment 2; the larger the reward effect, the more the cued distracter-expectation cost [r = 0.56, t(14) = 2.53, p = 0.024] (see Figure 4). This indicates that the degree of the engagement of a distraction-filtering mechanism depends on the degree of utilization of the cue. Participants who relied more on the cue information, as indicated by a larger reward effect, also exhibited a stronger attentional preparation, as indicated by a larger cued distracter-expectation cost.
Figure 4. Across-subject correlation analysis of cue utilization and attentional preparation.
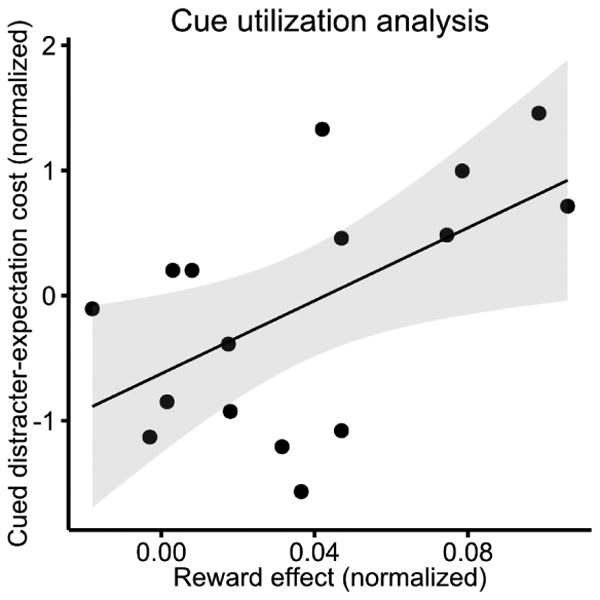
Across-subjects correlation between the effect of reward (positive numbers represent faster IEs for reward versus no-reward trials) and the cued distracter-expectation cost in the reward-context blocks of Experiment 2. The shaded grey area represents the 95% confidence interval of the linear fit. The relationship between the reward effect and cued distracter-expectation cost indicates that subjects who used the cue information more, reflected by showing a larger reward effect, also showed a greater preparation for distracter possibility, as indicated by a larger cued distracter-expectation cost.
Results of Experiment 2 thus confirm: (1) that monetary incentive speeds up performance in an arrow flanker conflict task; and (2) that the largest cued distracter-expectation cost is observed with simultaneous cueing of both potential distraction and reward-prospect, suggesting the possible recruitment of a specific distracter-filtering preparation mechanism. This experiment also revealed that reward has an advantageous effect on performance not only when there is monetary incentive at a single trial level, but also at the context-level, as indicated by the facilitation shown on no-reward trials occurring in a context in which they are intermixed with actual reward trials, compared to no-reward trials occurring when there is no reward context.
4. Discussion
Simultaneous cueing of distraction and reward elicits a cued distracter-expectation cost as part of attentional preparation
In this study, we explored preparatory mechanisms of attention that are engaged for dealing with potential upcoming distraction in a dynamic setting in which expectation for distraction could change on a trial-by-trial basis. Marini and colleagues (2013) previously showed that, in situations with occasional distracter stimuli, a distraction-filtering mechanism can be proactively invoked depending on contextual circumstances, such as the probability of distraction within an experimental block. The activation of such a distraction-filtering mechanism entailed a behavioral cost, however, which was termed a distraction-filtering cost. A critical aspect of that study was that expectation for potential distraction was manipulated at the block level, rather than on a trial-to-trial basis. Consequently, when the presence of distracters was highly likely, the distraction-filtering mechanism could be invoked in a tonic way and sustained throughout an experimental block.
In the current study, we aimed at investigating whether a similar distraction-filtering mechanism can be utilized in a more dynamic trial-to-trial setting, in which a cue indicates if the upcoming target is likely to have distracters or not. In order to create (or to not create) an expectation for distraction, two different types of cues were alternatively presented: either a cue predicting no distracters would occur or a cue predicting that distracters were likely. Crucially, the cue predicting no distraction was always valid, while the cue predicting distraction was valid in 75% of trials, so that it was followed on 25% of trials by a distracter-absent stimulus display. Thus, these imparted an expectation for distraction, although in 25% of trials the displayed target stimuli were physically identical to those in trials where cue predicted no distraction, allowing for a comparison of identical distracter-absent trials in different distracter-potential contexts. If the relatively costly preparatory processes for dealing with upcoming distraction can be dynamically invoked after cues predicting distraction (but not after cues predicting no distraction), then a cued distracter-expectation cost should be observed in this comparison -- i.e., a performance drop on distracter-absent trials when they were preceded by cues predicting distraction, compared to when they were preceded by cues predicting no distraction.
The results of Experiment 1 confirmed this prediction. A cued distracter-expectation cost was observed in distracter-absent trials when the cue predicted potential distracters compared to when it predicted there would be no distracters. The presence of the cued distracter-expectation cost likely reflects the dynamic and phasic engagement of a specific preparatory mechanism for filtering out potential distraction, presumably similar to the supramodal distraction-filtering mechanism characterized by Marini and colleagues (2013) where the distraction expectation was manipulated in a block-wise fashion. An additional result of Experiment 1 concerns the interdependence between the amount of reward and cued distracter-expectation cost. Specifically, Experiment 1 included a low-reward condition and a high-reward-condition, also indicated by the cue and occurring in randomized order (Della Libera & Chelazzi, 2006; Braem et al., 2014). We observed that the cued distracter-expectation cost, as reflected by the inverse efficiency measures and indicating the effort in attentional preparation specifically for the possibility of distracters, was significantly larger when the incentive was higher (i.e., in the high-reward condition compared to the low-reward condition). This difference derived from both an improvement on distracter-absent-cue trials and a slowing-down on distracter-present-cue trials. One of our predictions was that reward prospect optimizes attentional preparation and that this might include not only the implementation of a specific distraction-filtering mechanism but also a more efficient mobilization of cognitive resources for the task (e.g. Hubner & Schlosser, 2010). Accordingly, the observation of faster responses after distracter-absent cues in the high versus low reward conditions might be due to an increased efficiency in the mobilization of cognitive resources. This might in turn contribute to the instantiation of the relative cued distracter-expectation cost. Overall, the findings of Experiment 1 thus suggest that not only was the conjunctive cued manipulation of distraction prediction and reward amount effective in optimizing the dynamic preparation for distraction filtering, but also that such preparation directly depended on the value of the incentive.
Experiment 2 was designed to further explore the preparation for potential distraction by the use of another within-subject design in which trials with no reward prospect were delivered both in a separate block where no trials had reward prospect and as an additional manipulation within potentially rewarding blocks. In the no-reward context, the cue predicted solely the potential distraction, whereas in the reward context the cue was informative about both potential distraction and reward prospect. In this experiment, the interaction between cued reward prospect and cued distracter-expectation did not reach significance, which might suggest that the performance cost induced by a cue predicting a distracter-present trial relative to a cue predicting a distracter-absent trial is not strictly dependent on reward. However, further exploration of the results by means of planned pairwise comparisons showed that a significant cued distracter-expectation cost was found only in trials with reward-prospect occurring in the reward-context blocks. This result thus supports the view that when potential distraction is cued dynamically from trial-to-trial, a reliable cued distracter-expectation cost is incurred only when reward prospect is concurrently cued as well. Interestingly, a measure of the utilization of the cue, the reward effect, correlated across subjects with a measure of attentional preparation, the cued distracter-expectation cost. Thus, this finding confirms that the reward-related information embedded in the cue was effective in promoting attentional preparation for potential distraction.
The results of Experiments 1 and 2, taken together, suggest that in dynamic situations where there are environmental cues that provide additional information, two factors modulate the engagement of a specific distraction-filtering mechanism whose behavioral signature is the cued distracter-expectation cost. However, it is worth noting that in neither of the current experiments was the cued distracter-expectation cost inversely correlated to measures of distracter interference (i.e., incongruent vs. congruent distracters when they did occur), unlike the previous study by Marini and colleagues (2013). Accordingly, we cannot provide direct evidence here for the cued distracter-expectation cost being related to distraction filtering. We do not have a clear-cut explanation for the lack of interference reduction in the current study, and it would seem that this issue will need to be returned to in future work. In the current study, different mechanisms might have generated – or at least contributed to – the observed cued distracter-expectation cost. In this regard, we examine two possibilities. First, since the flanker interference effect does involve (although is not limited to) response-competition processes (e.g., Eriksen, 1995), one possibility is that participants adopted a more cautious response setting when the cue predicted distracters (compared to when the cue predicted no distracters). This might have been done in order to prevent the motor execution of an erroneous and selective response tendency (Cai, Oldenkamp & Aron, 2011) that could be triggered by the likely occurrence of incongruent flankers. If this were the case, however, the prolonged time-on-task in the cue-predicts-distracter trials (versus the cue-predicts-no-distracter trials) should have benefitted accuracy. We have addressed an analogous potential issue previously (Experiment 7 in Marini et al., 2013) by showing that the slowing-down of responses on distracter-absent trials embedded in a potentially distracting context was actually associated with a drop in accuracy. Similarly, here one might take note that in the high-reward condition of Experiment 1 and in the reward condition of Experiment 2, error rates were actually smaller in the cue-predicts-no-distracter condition compared to the cue-predicts-distracter condition (Table 1A). A second possibility for the observed cued distracter-expectation cost relates to the fact that participants were probably not expecting to see a distracter-absent target after cues predicting distraction. Deviation from expectation may trigger the involvement of control systems to make sure active schema, such as the task-set and the stimulus-response mapping, are still suitable (Norman & Shallice, 1986; Posner & DiGirolamo, 2000). In this regard, it should be noted that in our paradigm the presence (or absence) of distracters did not modify the required behavioral response and thus the task itself did not require reactive adjustments of any task-set or response-mapping control circuits. Accordingly, we have predicted and observed proactive modulations of control consisting in different preparation processes depending on the cue type. Therefore, although we cannot completely rule out the interpretation in terms of enhancement of control due to deviation from expectation, we think that it is unlikely to account for the observed cued distracter-expectation cost.
Whatever the mechanism(s) involved in generating the cued distracter-expectation cost, two factors seem to play an important role. One factor appears to be the actual prospect of a monetary reward, as indicated by the larger cued distracter-expectation cost in the high-reward condition (compared to the low-reward condition) of Experiment 1. The other factor is the pairing of the cue for reward prospect with trial-by-trial informative content concerning distraction potential, as demonstrated by the largest cued distracter-expectation cost observed in the reward trials of the reward block in Experiment 2. The presence of reward per se seems to facilitate a general preparation, while the pairing of reward and distraction-predicting information in the cue might promote the recruitment of a more specific distraction-filtering mechanism on a trial-by-trial basis. These aspects thus suggest that the presence of reward information in the cue may act by enhancing the relevance of the cue itself (Braem et al., 2014), thereby promoting the utilization of cue information for attentional preparation and distraction-filtering while pre-emptively maximizing the mobilization of cognitive resources for the task at hand.
Monetary reward prospect does not modulate the flanker incongruency effect
Some previous studies have shown that certain reward manipulations can reduce conflict-related incongruency effects (Padmala & Pessoa, 2011; Krebs et al., 2010; Krebs, Boehler, Appelbaum, & Woldorff, 2013). For instance, in a cued-reward Stroop-like study, Padmala and Pessoa (2011) found that when a relevant picture was overlaid with an irrelevant word, participants showed less interference from the irrelevant word in the reward condition compared to in the no-reward condition (as reflected by the amount of RT slowing when the irrelevant word was incongruent). Similarly, Krebs et al. (2010; 2013) found that in a word-color Stroop task (noncued) employing reward-associations for certain colors and not for others, participants showed less incongruency-related interference when a reward association was present for the relevant font-color dimension compared to when no such association was present. However, other studies have not consistently found a reward-mediated reduction in interference (Hubner & Schlosser, 2010; van den Berg, et al., 2014; Krebs, Boehler, Egner, & Woldorff, 2011). In line with the latter set of studies, in neither of the current experiments here did we find such reduction in interference from conflicting distracters under reward-prospect versus no-reward-prospect conditions.
There are several possible reasons why the conflict-related slowing was not reduced by the reward-prospect manipulation here. One possibility is that the paradigm utilized an arrow flanker task in which all the displayed visual stimuli shared the same perceptual and object-based characteristics (see also Discussion in van den Berg et al., 2014). More specifically, the target arrow, which was flanked by either congruent or incongruent arrows in distracter-present trials, shared its inherent features with those of those distracters (i.e., they were all arrows) and this might have limited the instantiation of a specific mechanism for enhancing the relevant dimension while suppressing the irrelevant one. This aspect would constitute a key difference relative to some of the previous studies, including the Padmala and Pessoa (2011) study where the features constituted by a relevant picture were overlaid with an irrelevant word (i.e., they were not embedded or integrated together in the same object), thus perhaps making it easier to selectively suppress or boost the processing of certain features. Moreover, similarly to the current study, Hubner and Schlosser (2010) did not observe a reduction of interference in a reward version of the flanker task. They concluded that the effect of reward is a mobilization of cognitive resources that enhances sensory coding unselectively, without modulating the scope of the attentional focus.
In the present study, the target and distracters were spatially separated but their features were identical (again, both were arrows). Spatial attention modulated by reward prospect could perhaps not so selectively mitigate the processing of the conflicting stimuli or enhance that of the target stimulus because the conflicting and target stimuli shared the same features. This idea, together with previous results (Hubner & Schlosser, 2010), suggests that reward may be able to modulate attentional control processes for reducing conflict by drawing attention away from irrelevant features and/or from irrelevant stimuli and toward relevant ones, but only when the relevant and irrelevant features are more easily separable. The present results thus suggest that an incentive-driven prioritization of rewarded stimuli to reduce stimulus conflict does not seem to apply very selectively to the domain of spatial attention, or at least fine-grained selective spatial attention, when adjacent spatial locations in the stimulus display would need to be selectively enhanced or suppressed (such as in the flanker task). Thus, although previous studies have shown that reward-driven reduction of conflict can act at the feature-specific level under certain circumstances, the present evidence does not indicate a role for reward prospect in reducing interference from conflicting stimuli in the domain of fine-grained spatial attention.
Reward context improves performance in the absence of a current trial-specific reward
An additional result that emerged from Experiment 2 regards the behavioral performance at the context-level under different rewarding circumstances. In Experiment 2, in addition to reward-present trials, there were two types of context in which no-reward trials were delivered. No-reward-prospect trials were delivered both under conditions of no-reward context, i.e., in experimental blocks where all trials were of this type, and under conditions of reward context, i.e., in experimental blocks where these trials were intermixed with reward-present trials. We observed that performance in no-reward trials was significantly better in the reward context than in the no-reward context. This might be due to participants using an increased motivational effort throughout the blocks in which there was the possibility of earning monetary rewards. However, if the effect of reward consists uniquely in a generalized increase in motivational effort at the block-level in the reward context, then one should not expect differences between reward-present trials and no-reward trials in the reward context. Instead, we observed both facilitation at the block level in the reward context and an additional facilitation when reward-prospect was present at the trial level. One possibility is that some participants simply ignored (or forgot the meaning of) the reward-predicting cue in the reward context, while others employed that information. Coherently with this interpretation, the visual inspection of individual responses on distracter-absent trials showed that five participants did not improve their performance on no-reward trials of the reward (vs. no-reward) context and that four participants did not improve their performance on reward (vs. no-reward) trials of the reward context. An alternative possibility, however, is that reward prospect can act both at the trial level and at the context level. Participants might have pre-alerted their attentional systems tonically throughout the reward blocks and then they might have also reinforced or strengthened this engagement more phasically every time the cue predicted potential reward. Future research about the way reward improves behavioral performance in trials where it is not currently delivered (e.g., the present study; Zedelius et al., 2012) would be needed to further investigate the hypothesis of a dual-component (i.e., tonic and phasic) mechanism of reward.
Conclusions
The current work examined how distraction-filtering mechanisms interact with reward-related modulations of preparatory attention. Results revealed a multi-faceted interplay between these two processes. When no rewards were involved, the trial-to-trial dynamics of preparation for potential distraction appeared to rely mainly on a general resource mobilization, without specific utilization of a distraction-filtering mechanism. When there was a prospect of reward, however, the presence of a cued distracter-expectation cost suggests participants might have recruited a specific distraction-filtering mechanism, although the observed slowing down was not accompanied by a reduction of distracter interference and it thus might be due to other concurrent factors. The advantage of reward prospect on speeded RT-performance was observed both at the trial level, where it was strongest, and at the context-level, where it was still significantly present (i.e., on no-reward trials in a reward-context circumstance compared to the same trials in a no-reward context). These results, taken together, shed light on the complex dynamics by which reward shapes attentional preparation, and particularly into those mediated by prior information about potential distraction.
Acknowledgments
We thank Brittany Zulkiewicz, Zaynah Alam, Frank Lee, and Christine Tina Wei for their helpful assistance with data collection. We also thank two anonymous reviewers for suggesting several possible alternative interpretations of our results that we present in the Discussion. This work was supported by grants from the National Institutes of Health (R01-MH060415 and R01-NS051048) to M.G.W.
Footnotes
We are grateful to Prof. Mauro Marini for the mathematical formulation of the equation of this function.
In the RT data, this Reward by Cue-distraction interaction was not significant [F(1,15)=2.59, p=.12].
Contributor Information
Francesco Marini, Email: francesco.pd@gmail.com.
Berry van den Berg, Email: berryv.dberg@gmail.com.
Marty G. Woldorff, Email: woldorff@duke.edu.
References
- Austen EL, Enns JT. Change detection in an attended face depends on the expectation of the observer. Journal of Vision. 2003;3:64–74. doi: 10.1167/3.1.7. [DOI] [PubMed] [Google Scholar]
- Awh E, Matsukura M, Serences JT. Top-down control over biased competition during covert spatial orienting. Journal of Experimental Psychology: Human Perception and Performance. 2003;29:52–63. doi: 10.1037//0096-1523.29.1.52. [DOI] [PubMed] [Google Scholar]
- Boehler CN, Schevernels H, Hopf JM, Stoppel CM, Krebs RM. Reward prospect rapidly speeds up response inhibition via reactive control. Cognitive Affective and Behavioral Neurosciences. 2014 doi: 10.3758/s13415-014-0251-5. in press. [DOI] [PubMed] [Google Scholar]
- Braem S, Verguts T, Roggeman C, Notebaert W. Reward modulates adaptation to conflict. Cognition. 2012;125:324–332. doi: 10.1016/j.cognition.2012.07.015. [DOI] [PubMed] [Google Scholar]
- Braem S, Hickey C, Duthoo W, Notebaert W. Reward determines the context-sensitivity of cognitive control. Journal of Experimental Psychology: Human Perception and Performance. 2014;40(5):1769–1778. doi: 10.1037/a0037554. [DOI] [PubMed] [Google Scholar]
- Braver TS, Paxton JL, Locke HS, Barch DM. Flexible neural mechanisms of cognitive control within human prefrontal cortex. Proceedings of the National Academy of Sciences of the USA. 2009;106(18):7351–7356. doi: 10.1073/pnas.0808187106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cai W, Oldenkamp CL, Aron AR. A proactive mechanism for selective suppression of response tendencies. The Journal of Neuroscience. 2011;31(16):5965–5969. doi: 10.1523/JNEUROSCI.6292-10.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Della Libera C, Chelazzi L. Visual selective attention and the effects of monetary rewards. Psychological Science. 2006;17(3):222–227. doi: 10.1111/j.1467-9280.2006.01689.x. [DOI] [PubMed] [Google Scholar]
- Eriksen BA, Eriksen CW. Effects of noise letters upon identification of a target letter in a non-search task. Perception & Psychophysics. 1974;16:143–149. [Google Scholar]
- Eriksen CW. The flankers task and response competition: A useful tool for investigating a varietyof cognitive problems. Visual Cognition. 1995;2(2-3):101–118. [Google Scholar]
- Geng JJ. Attentional mechanisms of distractor suppression. Current Directions in PsychologicalScience. 2014;23(2):147–153. [Google Scholar]
- Heed T, Habets B, Sebanz N, Knoblich G. Others' actions reduce crossmodal integration in peripersonal space. Current Biology. 2010;20:1345–1349. doi: 10.1016/j.cub.2010.05.068. [DOI] [PubMed] [Google Scholar]
- Holm S. A simple sequentially rejective multiple test procedure. Scandinavian Journal of Statistics. 1979;6:65–70. [Google Scholar]
- Hubner R, Schlosser J. Monetary reward increases attentional effort in the flanker task. Psychonomic Bulletin & Review. 2010;17(6):821–826. doi: 10.3758/PBR.17.6.821. [DOI] [PubMed] [Google Scholar]
- Kleiner M, Brainard D, Pelli D, Ingling A, Murray R, Broussard C. What's new in Psychtoolbox-3. Perception. 2007;36(14):1–1. [Google Scholar]
- Krebs RM, Boehler CN, Appelbaum LG, Woldorff MG. Reward associations reduce behavioral interference by changing the temporal dynamics of conflict processing. PLoS ONE. 2013;8(1):e53894. doi: 10.1371/journal.pone.0053894. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Krebs RM, Boehler CN, Egner T, Woldorff MG. The neural undeprinnings of how reward associations can both guide and misguide attention. The Journal of Neuroscience. 2011;31(26):9752–9759. doi: 10.1523/JNEUROSCI.0732-11.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Krebs RM, Boehler CN, Roberts KC, Song AW, Woldorff MG. The involvement of the dopaminergic midbrain and cortico-striatal-thalamic circuits in the integration of reward prospect and attentional task demands. Cerebral Cortex. 2012;22:607–615. doi: 10.1093/cercor/bhr134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Krebs RM, Boehler CN, Woldorff MG. The influence of reward associations on conflict processing in the Stroop task. Cognition. 2010;117:341–347. doi: 10.1016/j.cognition.2010.08.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Locke HS, Braver TS. Motivational influences on cognitive control: behavior, brain activation, and individual differences. Cognitive Affective and Behavioral Neurosciences. 2008;8(1):99–112. doi: 10.3758/cabn.8.1.99. [DOI] [PubMed] [Google Scholar]
- Marini F, Chelazzi L, Maravita A. The costly filtering of potential distraction: evidence for a supramodal mechanism. Journal of Experimental Psychology: General. 2013;142(3):906–922. doi: 10.1037/a0029905. [DOI] [PubMed] [Google Scholar]
- Marini F, Marzi T, Viggiano MP. “Wanted!” The effects of reward on face recognition: electrophysiological correlates”. Cognitive Affective and Behavioral Neurosciences. 2011;11(4):627–643. doi: 10.3758/s13415-011-0057-7. [DOI] [PubMed] [Google Scholar]
- Marini F, Tagliabue CF, Sposito AV, Hernandez-Arieta A, Brugger P, Estévez N, Maravita A. Crossmodal representation of a functional robotic hand arises after extensive training in healthy participants. Neuropsychologia. 2014;53:178–186. doi: 10.1016/j.neuropsychologia.2013.11.017. [DOI] [PubMed] [Google Scholar]
- Norman DA, Shallice T. Attention to action. Springer; 1986. [Google Scholar]
- Padmala S, Pessoa L. Reward reduces conflict by enhancing attentional control and biasing visual cortical processing. Journal of Cognitive Neuroscience. 2011;23(11):3419–3432. doi: 10.1162/jocn_a_00011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Posner MI, DiGirolamo GJ. Conflict, target detection and cognitive control. In: Parasuraman R, editor. The attentive brain. MIT Press; 2000. [Google Scholar]
- R Development Core Team. R Foundation for Statistical Computing; Vienna, Austria: 2013. R: A language and environment for statistical computing. URL: http://www.R-project.org/ [Google Scholar]
- Ratcliff R. Methods for dealing with reaction times outliers. Psychological Bulletin. 1993;114:510–532. doi: 10.1037/0033-2909.114.3.510. [DOI] [PubMed] [Google Scholar]
- Ruff CC, Driver J. Attentional preparation for a lateralized visual distractor: Behavioral and fMRI evidence. Journal of Cognitive Neuroscience. 2006;18(4):522–538. doi: 10.1162/jocn.2006.18.4.522. [DOI] [PubMed] [Google Scholar]
- Schevernels H, Krebs RM, Santens P, Woldorff MG, Boehler CN. Task preparation processes related to reward prediction precede those related to task-difficulty expectation. Neuroimage. 2014;84:639–647. doi: 10.1016/j.neuroimage.2013.09.039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Seifert J, Naumann E, Hewing J, Hagemann D, Bartussek D. Motivated executive attention-incentives and the noise-compatibility effect. Biological Psychology. 2006;71:80–89. doi: 10.1016/j.biopsycho.2005.03.001. [DOI] [PubMed] [Google Scholar]
- Serences JT, Yantis S, Culberson A, Awh E. Preparatory activity in visual cortex indexes distractor suppression during covert spatial orienting. Journal of Neurophysiology. 2004;92:3538–3545. doi: 10.1152/jn.00435.2004. [DOI] [PubMed] [Google Scholar]
- Stanisor L, van der Togt C, Pennartz CMA, Roelfsema PR. A unified selection signal for attention and reward in primary visual cortex. Proceedings of the National Academy of Sciences of the United States of America. 2013;110(22):9136–9141. doi: 10.1073/pnas.1300117110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Townsend JT, Ashby FG. Stochastic modeling of elementary psychological processes. Cambridge: Cambridge University Press; 1983. [Google Scholar]
- Van den Berg B, Krebs RM, Lorist MM, Woldorff MG. Utilization of reward-prospect enhances preparatory attention and reduces stimulus conflict. Cognitive Affective and Behavioral Neurosciences. 2014;14(2):561–577. doi: 10.3758/s13415-014-0281-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Van Steenbergen H, Band GPH, Hommel B. Reward counteracts conflict adaptation: evidence for a role of affect in executive control. Psychological Science. 2014;20:1473–1477. doi: 10.1111/j.1467-9280.2009.02470.x. [DOI] [PubMed] [Google Scholar]
- Vassena E, Silvetti M, Boehler CN, Achten E, Fias W, Verguts T. Overlapping neural systems represent cognitive effort and reward anticipation. PLoS ONE. 2014;9(3):e91008. doi: 10.1371/journal.pone.0091008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Walsh BJ, Buonocore MH, Carter CS, Mangun GR. Integrating conflict detection and attentional control mechanisms. Journal of Cognitive Neuroscience. 2011;23(9):2211–2221. doi: 10.1162/jocn.2010.21595. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weissman DH, Roberts KC, Visscher KM, Woldorff MG. The neural bases of momentary lapses in attention. Nature Neuroscience. 2006;9(7):971–978. doi: 10.1038/nn1727. [DOI] [PubMed] [Google Scholar]
- Wessel JR, O'Doherty JP, Berkebile MM, Linderman DL, Aron AR. Stimulus devaluation induced by stopping action. Journal of Experimental Psychology: General. 2014;143(6):2316–2329. doi: 10.1037/xge0000022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Woldorff MG, Hazlett CJ, Fichtenholtz HM, Weissman DH, Dale AM, Song AW. Functional parcellation of attentional control regions of the brain. Journal of Cognitive Neuroscience. 2004;16(1):149–165. doi: 10.1162/089892904322755638. [DOI] [PubMed] [Google Scholar]
- Zedelius CM, Veling H, Bijleveld E, Aarts H. Promising high monetary rewards for future task performance increases intermediate task performance. PLoS ONE. 2012;7(8):e42547. doi: 10.1371/journal.pone.0042547. [DOI] [PMC free article] [PubMed] [Google Scholar]