

Abstract
Salmonella species are zoonotic pathogens and leading causes of food borne illnesses in humans and livestock1. Understanding the mechanisms underlying Salmonella-host interactions are important to elucidate the molecular pathogenesis of Salmonella infection. The Gentamicin protection assay to phenotype Salmonella association, invasion and replication in phagocytic cells was adapted to allow high-throughput screening to define the roles of deletion mutants of Salmonella enterica serotype Typhimurium in host interactions using RAW 264.7 murine macrophages. Under this protocol, the variance in measurements is significantly reduced compared to the standard protocol, because wild-type and multiple mutant strains can be tested in the same culture dish and at the same time. The use of multichannel pipettes increases the throughput and enhances precision. Furthermore, concerns related to using less host cells per well in 96-well culture dish were addressed. Here, the protocol of the modified in vitro Salmonella invasion assay using phagocytic cells was successfully employed to phenotype 38 individual Salmonella deletion mutants for association, invasion and intracellular replication. The in vitro phenotypes are presented, some of which were subsequently confirmed to have in vivo phenotypes in an animal model. Thus, the modified, standardized assay to phenotype Salmonella association, invasion and replication in macrophages with high-throughput capacity could be utilized more broadly to study bacterial-host interactions.
Keywords: Infectious Diseases, Issue 90, Salmonella enterica Typhimurium, association, invasion, replication, phenotype, intracellular pathogens, macrophages
Introduction
Nontyphoidal Salmonella are important causes of enteric diseases in all vertebrates. Salmonellosis in humans is among the top bacterial food-borne diseases1. Characterization of the molecular mechanisms that underpin the interactions of Salmonella with their animal hosts is mainly achieved through the study of Salmonella enterica serotype Typhimurium (STM) in tissue culture and animal models of infection. Gaining insights in STM-host interactions will help us understand how Salmonella survive and grow inside host cells. The first challenge in studying these interactions is to identify as many participating factors as possible from both host and pathogen, but these endeavors are largely obstructed by the significant difficulties of dealing with two independent complex biological systems simultaneously, i.e., host and Salmonella,under physiological conditions. Additionally, the large repertoire of Salmonella and host genes potentially encoding factors involved in host interactions require high-throughput biological platform to tackle this challenge.
A modified, standardized assay to phenotype Salmonella association, invasion and replication in macrophages with high-throughput capacity was developed to examine a large set of genes likely engaging in Salmonella-host interactions. The Gentamicin protection assay was developed in 19732, but was first thoroughly described by Elsinghorst in 19943,4. It has now become a standard tool for studying many intracellular bacterial pathogens ex vivo, including Salmonella5,6. Internalized bacteria avoid being killed by some antibiotics, like Gentamicin, that cannot penetrate eukaryotic cells3. By taking advantage of this phenomenon, the Gentamicin protection assay measures the survival and growth of intracellular bacterial pathogens. Three events during the infection, i.e., association with eukaryotic cells, invasion and replication, can be evaluated for intracellular bacterial pathogens based on the time interval between infection, Gentamicin treatment, and further incubation (Figure 1). Eukaryotic cell lines provide a physiological environment that is less complex than relevant animal models for host-pathogen interaction studies.
The Gentamicin protection assay is an appropriate platform to study STM-host interactions, but the standard assay in a 24-well culture dish has low-throughput capacity. Computational analysis of in vivo datasets identified 149 Salmonella gene products that are predicted to interact with approximately 300 host gene products (unpublished data). The standard Gentamicin protection assay does not have the capacity to phenotype this number of mutants efficiently.
In addition, the Gentamicin protection assay can theoretically detect the invasion of even a single bacterium. Because of this inherent sensitivity, the raw data are susceptible to technical variances when repeated at different times. The internal controls and relative data presentation after normalization are essential for meaningful interpretation of the results. Given these considerations, a modified, standardized Gentamicin protection assay was developed to enhance testing capacity and increase precision.
The following protocol is detailed and illustrated to perform the modified Gentamicin protection assay using 96-well culture dishes and the murine macrophage RAW264.7 cell line. Compared to the standard protocol in 24-well culture dishes, the modified protocol has the following advantages: 1) Using 96-well culture dishes allows up to 10 different mutant strains to be phenotyped including internal positive and negative controls with sufficient statistical power; 2) The variance of results is significantly reduced, because the mutant strains are tested in the same culture dish and at the same time; 3) The use of multichannel pipettes increases throughput while reducing operator fatigue. Lastly, comparing to 24-well culture dishes, concerns of less host cells per well in 96-well culture dish were addressed through protocol optimization and standardization.
In summary, the modified, standardized assay to in vitro phenotype Salmonella or other bacterial association, invasion and replication in phagocytic cells increases precision and achieves high-throughput capacity while reducing operator fatigue.
Protocol
1. Murine Macrophage RAW264.7 Cell Culture
Grow low passage number murine macrophage cells, RAW264.7 (The ATCC® Number, TIB-71) in a T-75 cell culture flask vented filter cap in Dulbecco's Modified Eagle Medium (DMEM) supplemented with 10% fetal bovine serum (FBS), 0.5% NaHCO3, and 1% 100x Nonessential amino acids (NEAA) at 37°C, in a 5% CO2 incubator.
Once cells reach a 60-80% confluence in the flask, use a cell scraper to harvest cells, and count the cells in a hemacytometer and calculate the cell concentration.
Resuspend the cells and dilute the cell concentration into 2.5 x 105/ml in fresh DMEM cell culture medium. Use multichannel pipettes to plate 200 μl of DMEM cell culture medium containing 5 x 104 cells in each well of 96-well cell culture plates.
Plate four wells of each Salmonella strain. Set up three separate plates for one set of phagocytic cells invasion assay, and mark them with “association”, “invasion” and “replication”, respectively.
Place the plates in the 5% CO2 incubator at 37 °C ON to allow the macrophage cells to attach to the bottom of the wells.
2. Preparation of Salmonella Wild-type and Mutants
On the same day of preparing the macrophage RAW264.7 cells, pick single colonies of wild-type (WT) Salmonella enterica serotype Typhimurium 14028s, DinvA mutant, DphoP mutant, and the desired test mutant strains, and culture them in Luria-Bertani (LB) broth supplemented with antibiotics as appropriate. Use each set of the phagocytic cells invasion assays to test up to 10 mutant strains (DinvA mutant and DphoP mutant are defective in invasion and intracellular replication, respectively).
Grow the bacteria for 14 hr at 37 °C with shaking at 220 rpm in 5 ml each of LB broth in loosely capped 14 ml polypropylene in a round bottom tube. LB broth contains appropriate antibiotics, in our case, most of mutant strains are resistant to Kanamycin, 50 µg/ml
The next day, subculture each of the ON cultures of Salmonella strains in plain 5 ml of LB broths at a ratio of 1:50 for an additional 4 hr with shaking at 220 rpm.
Read OD600 values of each bacterial culture on a spectrophotometer, and each read should range from 0.6-1.2 to optimize Salmonella pathogenicity island-1 type III secretion system (SPI-1 TTSS) gene expression for invasion.
Use the formula of 1 OD600 = 7.5 x 108 CFU/ml to estimate the bacterial concentration, then dilute bacteria to a concentration of 5 x 106 CFU/ml in fresh DMEM cell culture medium.
3. Invasion Assay in 96-well Culture Plate
Remove three 96-well cell culture plates from the CO2 incubator and wash each well once with 200 μl of 1x PBS buffer.
After washing and complete removal of PBS, add 200 μl bacteria in DMEM medium (see above) into each well. This results in 1 x 106 Salmonella cells in each well and multiplicity of infection (MOI) of 20:1.
Centrifuge the plates at 1,000 x g in a sealed carrier for 10 min, then replace plates (time zero) in a 37 °C, 5% CO2 incubator for 30 min. For each strain, set up at least four duplicate wells.
After 30 min, remove plates from the incubator and wash three times with 200 μl of 1x PBS to remove unassociated bacteria.
Following washing, save the plate labeled with “association” for further treatment. Add 200 μl of DMEM medium containing 100 μg/ml Gentamicin to each well of the plates marked with “invasion” and “replication” in order to kill the extracellular Salmonella. Return the plates to 37 °C, 5% CO2 incubator for an additional hr.
Save one well from each infected strain for recording macrophage cell count, treat the rest of wells on the “association” plate with 200 μl of 1x PBS containing 1% Triton X-100 for 10 min. The Triton solution will lyse macrophage cells and release associated Salmonella.
Harvest Salmonella from each well and place them into 1.5 ml Eppendorf tubes. Perform three 10 fold serial dilutions with 900µl 1x PBS on the harvested samples from each well, vortex is performed between each dilution.
Plate the third dilution, 10-3, on either LB (WT) or LB Kan (mutants) plates. Label the plates with appropriate Salmonella strain name, dilution number, time point and date.
After vortexing the third dilution tube, dispense 10 µl sample on the surface of the agar and repeat four times for a total of 5 drops. Be sure to space the five 10 μl drops evenly on the Petri dish plates7.
After the drops soak into the agar, turn the plates over and incubate overnight at 37 °C, 5% CO2 incubator.
Treat wells saved for counting of macrophages with 150 µl 1xPBS containing 0.25% Trypsin-EDTA for 10 min.
After trypsinization, place macrophages into 1.5 ml Eppendorf tubes and treat them with 50 µl of FBS to neutralize Trypsin/EDTA solution, stain the cells with 0.4% Trypan blue and score them through using hemacytometer.
When the one hr incubation has elapsed, remove the “invasion” and “replication” plates from the CO2 incubator, and wash each well as “Step 3.4”.
Save the “invasion” plate for further treatment as “Step 3.6-3.12”.
Add 200 µl of DMEM medium containing 10 μg/ml Gentamicin to the “replication” plate to maintain clearance of extracellular Salmonella in the medium. Return the plate to 37 °C, 5% CO2 incubator for an additional 22.5 hr.
The next day, remove the “replication” plate from 37 °C, 5% CO2 incubator, and wash each well as “Step 3.4” and treat them as “Step 3.6-3.12”.
Lastly, remove the agar plates after incubation overnight in 37 °C incubator and score the number of bacterial colonies.
A good colony distribution should be between 10 and 100 colonies, each spot approximately contains 2 to 20 colonies, it would be hard to score if there were more than 20 colonies on one spot. If the count is too low or too high, replate the sample with 10-fold higher or lower dilutions as needed.
4. Data Analysis
Calculate the CFU (colony formation units per ml) of each plate based on the plating volume and dilution factor.
Ascertain the number of Salmonella per macrophage through the recorded macrophage cell count from each strain.
Calculate the geometric means of CFU per macrophage from at least three independent experiments for cell association, invasion or replication, respectively, and normalize further to the WT for the relative value.
Analyze the data for association, invasion, and replication by Student’s t-test respectively, by comparing each strain to WT, and determine the statistical significance by the p-value.
Representative Results
See representative results (Figure 2) after the data are plotted based on the modified phagocytic cell invasion assay. The data include five different strains, WT, ΔinvA, ΔphoP, mutant A, and mutant B. ΔinvA, known to be defective for invasion, and ΔphoP known to be defective for replication8, are used as positive controls to assess the experimental validity. Indeed, in the modified invasion assay, a ΔinvA mutant is internalized poorly by RAW264.7 cells and a ΔphoP mutant had reduced replication after 24 hr. Mutants A and B are representative strains assayed using this protocol. Mutant A shows significant reduction of replication (Figure 2), similar to a ΔphoP mutant; interestingly, mutant B shows a significant increase in replication (Figure 2). The data clearly indicate both of those mutants have altered phenotypes for invasion and survival in macrophages.
In summary, numerous Salmonella deletion mutants were individually phenotyped for involvement in the bacterial association, invasion, and replication in RAW264.7 macrophages using the modified invasion assay. Twenty of 38 tested mutants so far displayed variable but significant defects in bacterial infection of macrophages (Table 1), thus the modified assay to in vitro phenotype Salmonella or other bacterial association, invasion and replication in phagocytic cells increases precision and high-throughput capacity while reducing operator fatigue.
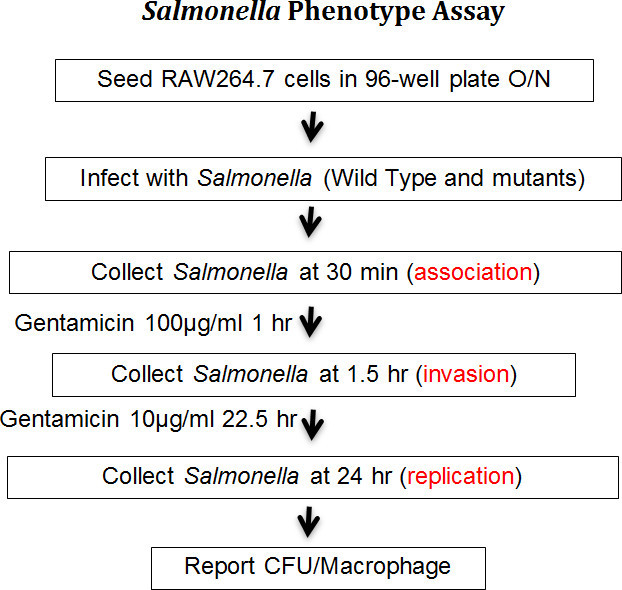 Figure 1. Working scheme of high-throughput assay. Gentamicin protection assay is modified, standardized to phenotype numerous Salmonella mutants simultaneously in 96-well plates. The key steps are performed as above to examine association, invasion and replication of Salmonella in RAW264.7 macrophage cells.
Figure 1. Working scheme of high-throughput assay. Gentamicin protection assay is modified, standardized to phenotype numerous Salmonella mutants simultaneously in 96-well plates. The key steps are performed as above to examine association, invasion and replication of Salmonella in RAW264.7 macrophage cells.
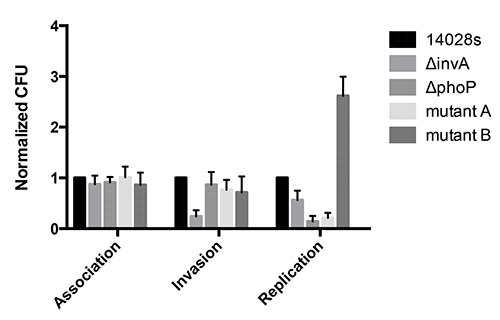 Figure 2. Representative results. The high-throughput assays are performed for Salmonella strains — WT, ΔinvA, ΔphoP, mutant A, and mutant B — in RAW264.7 macrophage cells. For each strain, the geometric means of CFU per macrophage are obtained from three independent experiments for cell association, invasion and replication, respectively, which are further normalized to the WT and graphic representation is shown. Error bars denote standard error, and statistical significance of each strain referenced to WT was determined by Student’s t-test. *p<0.05. MOI=20.
Figure 2. Representative results. The high-throughput assays are performed for Salmonella strains — WT, ΔinvA, ΔphoP, mutant A, and mutant B — in RAW264.7 macrophage cells. For each strain, the geometric means of CFU per macrophage are obtained from three independent experiments for cell association, invasion and replication, respectively, which are further normalized to the WT and graphic representation is shown. Error bars denote standard error, and statistical significance of each strain referenced to WT was determined by Student’s t-test. *p<0.05. MOI=20.
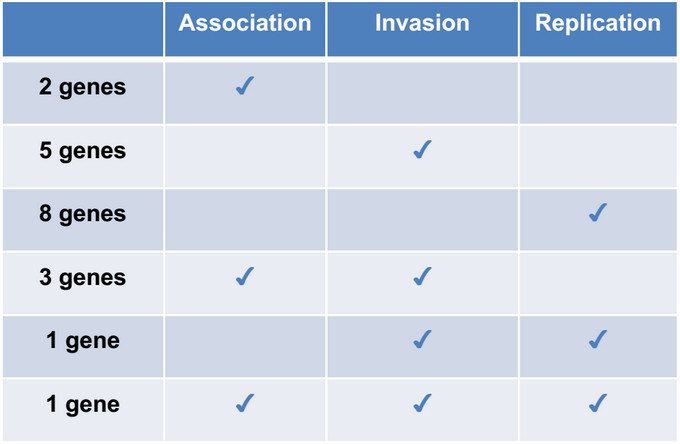 Table 1. Phenotypes of candidate genes. Candidate Salmonella mutants are phenotyped by the modified, standardized assay for association, invasion and replication in RAW264.7 macrophage cells. 20 mutants have significant defects in association, invasion and/or replication.
Table 1. Phenotypes of candidate genes. Candidate Salmonella mutants are phenotyped by the modified, standardized assay for association, invasion and replication in RAW264.7 macrophage cells. 20 mutants have significant defects in association, invasion and/or replication.
Discussion
The Gentamicin protection assay is widely used to study the invasion and replication of intracellular bacterial pathogens inside host cell, and it is especially an important biological tool for studying pathogens, like Salmonella, whose invasion is the prerequisite step for establishing infection1. The standard Gentamicin protection assay in Salmonella research community is implemented in 24-well culture dish5. Though the use of 48 or even 96-well plates were discussed before for high-throughput capacity9, no actual protocol has been demonstrated in detail and successfully employed to test a collection of bacterial mutants. Here, a modified and standardized protocol was developed to use 96-well culture dishes to phenotype Salmonella mutants. Notably, this protocol was only tested for Salmonella and phagocytic cells, thus modifications and optimization would obviously be necessary with other bacteria or host cells.
There are several advantages to the modified Gentamicin protection assay to phenotype Salmonella association, invasion and replication in phagocytic cells. First, the 96-well culture dish format allows high-throughput testing and analysis, up to 10 different strains can be tested simultaneously with sufficient repetition statistical analysis and internal controls. Second, because of the high sensitivity of the assay, the raw data of the Gentamicin protection assay may be susceptible to technical variances when repeated at different times. Under this modified protocol, several strains are phenotyped under the same conditions throughout the process, reducing variance. Lastly, the use of multichannel pipettes increases efficiency and reduces operator fatigue. The new protocol facilitated the rapid phenotyping numerous Salmonella mutants while generating large amounts of quantitative, reproducible data.
One concern in doing invasion assays in 96-well culture dishes is that the wells contain fewer eukaryotic cells, and because of this quantitation could be compromised. To address this concern, a large series of assays were performed to optimize reproducibility. First, the RAW264.7 macrophages used in this protocol had fewer than 20 passages, and the cells were seeded into 96-well culture dish overnight before the assay. Second, the MOI was reduced to 20:1 from the regularly used 50:1 or 100:1. A MOI 20:1 ensures that 99% of macrophages will be exposed to infection by 10 to 30 bacteria calculated by Poisson distribution according to the stochastic point. It is thought that, not restricted to biological factors, cellular damage could be caused from overwhelming bacteria physical contacts10. Additionally, Salmonella invasion could induce apoptosis of macrophages, which is positively associated with high MOI11. Using a MOI of 20:1, cellular damage was found to be minimal, presumably due to limited bacteria-macrophage physical contact. Third, the incubation time interval was optimized for the three time points: association for 30 min with no Gentamicin; invasion for additional 1 hr (total 1.5 hr post-infection) with 100 μg/ml Gentamicin; replication for additional 22.5 hr (total 24 hr after infection) with 10 μg/ml Gentamicin. In our test, 5 μg/ml Gentamicin is sufficient to kill all the Salmonella in cell-free DMEM with 10% fetal bovine serum (unpublished data), thus, 100 μg/ml Gentamicin would be expected to ensure the rapid killing and 10 g/ml Gentamicin would maintain the clearance of extracellular bacteria in the cell culture medium for overnight incubation. In vivo, it takes about 15 min to detect tissue associated Salmonella with calf intestine epithelial cells12,13; in vitro, 30 min incubation is reported to be adequate for the wild-type to invade14. For the standard invasion assay, the work load of the first two time points is quite intense due to washing, serial dilution and plating. The precise time interval for each time point was achieved by utilizing the multichannel pipettes, because the device significantly increases the operator’s efficiency and precision. Furthermore, the sample plating technique was altered after serial dilution, instead of plating 100 μl and manually spreading, 5 separate drops of 10 μl were plated for each repeat well on plates which greatly reduces the plating time and increases the accuracy, because the final count per well is scored from 5 plated samples. Collectively, these optimization and standardization procedures increase the reproducibility of the assay, different operators can repeatedly obtain consistent results at different times.
This protocol employs phagocytic cells instead of epithelial cells for this assay in a smaller 96-well format. In studying the pathogenesis of salmonellosis, the most widely used epithelial cell line is Caco-2, derived from heterogeneous human epithelial colorectal adenocarcinoma. Using the regular 24-well culture dish invasion test with Caco-2 cells, the actual number of recovered bacteria at each time point, particularly the 22.5 hr time point for phenotyping replication, is much lower than when phagocytic cells, such as J774 or RAW264.7 macrophages, are used (unpublished data). Thus, this makes it more difficult to perform phenotyping using a 96-well culture dish when each well has fewer host cells. It is known that macrophages cells could actively engulf bacteria, which could lead to more bacteria being internalized, and it also cannot be ruled out that Salmonella apparently replicate more slowly in Caco-2 cells. The ability of Salmonella to survive and replicate inside macrophage cells is related to key roles in the pathogenesis of salmonellosis, i.e., triggering massive host inflammation responses and facilitating systemic infection1. We mainly employed the modified and standardized Gentamicin protection assay to phenotype a large set of computation pipeline selected genes for their potential engagements in host interactions. It turned out nearly 50% of the Salmonella mutants were phenotyped as genes involved in intracellular replication in phagocytic cells.
Disclosures
We have nothing to disclose.
Acknowledgments
This project was supported partly by a grant for National Institutes of Health NIAID (for A.J.B. and L.G.A., R01 AI076246). The Salmonella mutant collection was partly supported by National Institutes of Health grants (for M.M., U01 A152237-05, R01 AI07397-01, R01 AI039557-11 and R01 AI075093-01), partly by National Institutes of Health grants (for H.A.P, R21 AI083964-01, 1R0 1AI083646-01, 1R56AI077645, R01 AI075093). We thank Steffen Prowollik for replica plating and confirming the mutants in the collection.
References
- Haraga A, Ohlson MB, Miller SI. Salmonellae interplay with host cells. Nat Rev Microbiol. 2008;6:53–66. doi: 10.1038/nrmicro1788. [DOI] [PubMed] [Google Scholar]
- Mandell GL. Interaction of intraleukocytic bacteria and antibiotics. The Journal of clinical investigation. 1973;52:1673–1679. doi: 10.1172/JCI107348. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Elsinghorst EA. Measurement of invasion by gentamicin resistance. Methods Enzymol. 1994;236:405–420. doi: 10.1016/0076-6879(94)36030-8. [DOI] [PubMed] [Google Scholar]
- Elsinghorst EA, Weitz JA. Epithelial cell invasion and adherence directed by the enterotoxigenic Escherichia coli tib locus is associated with a 104-kilodalton outer membrane protein. Infect Immun. 1994;62:3463–3471. doi: 10.1128/iai.62.8.3463-3471.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Behlau I, Miller SI. A PhoP-repressed gene promotes Salmonella typhimurium invasion of epithelial cells. J Bacteriol. 1993;175:4475–4484. doi: 10.1128/jb.175.14.4475-4484.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hueck CJ, et al. Salmonella typhimurium secreted invasion determinants are homologous to Shigella Ipa proteins. Mol Microbiol. 1995;18:479–490. doi: 10.1111/j.1365-2958.1995.mmi_18030479.x. [DOI] [PubMed] [Google Scholar]
- Steele-Mortimer O. Infection of epithelial cells with Salmonella enterica. Methods Mol Biol. 2008;431:201–211. doi: 10.1007/978-1-60327-032-8_16. [DOI] [PubMed] [Google Scholar]
- Foster JW, Spector MP. How Salmonella survive against the odds. Annu Rev Microbiol. 1995;49:145–174. doi: 10.1146/annurev.mi.49.100195.001045. [DOI] [PubMed] [Google Scholar]
- Edwards AM, Massey RC. Invasion of human cells by a bacterial pathogen. Journal of visualized experiments JoVE. 2011;10 doi: 10.3791/2693. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Molinari G, et al. The role played by the group A streptococcal negative regulator Nra on bacterial interactions with epithelial cells. Mol Microbiol. 2001;40:99–114. doi: 10.1046/j.1365-2958.2001.02373.x. [DOI] [PubMed] [Google Scholar]
- Van der Velden AW, Lindgren SW, Worley MJ, Heffron F. Salmonella pathogenicity island 1-independent induction of apoptosis in infected macrophages by Salmonella enterica serotype typhimurium. Infect Immun. 2000;68:5702–5709. doi: 10.1128/iai.68.10.5702-5709.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Santos RL, Zhang S, Tsolis RM, Baumler AJ, Adams LG. Morphologic and molecular characterization of Salmonella typhimurium infection in neonatal calves. Vet Pathol. 2002;39:200–215. doi: 10.1354/vp.39-2-200. [DOI] [PubMed] [Google Scholar]
- Lawhon SD, et al. Role of SPI-1 secreted effectors in acute bovine response to Salmonella enterica Serovar Typhimurium a systems biology analysis approach. PLoS One. 2011;6:e26869. doi: 10.1371/journal.pone.0026869. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Drecktrah D, Knodler LA, Galbraith K, Steele-Mortimer O. The Salmonella SPI1 effector SopB stimulates nitric oxide production long after invasion. Cell Microbiol. 2005;7:105–113. doi: 10.1111/j.1462-5822.2004.00436.x. [DOI] [PubMed] [Google Scholar]