

Local and global dynamics in folded RNAs occur over broad timescales spanning picoseconds to minutes.1 Slow motions likely play predominant roles in governing RNA folding and ribonucleoprotein assembly reactions. However, slow local motions are extremely difficult to detect, especially for large RNAs with complex structures.
The local environment and degree of flexibility can be evaluated at nucleotide resolution for RNAs of any size using selective 2′-hydroxyl acylation analyzed by primer extension (SHAPE) chemistry.2 RNA nucleotides exist in equilibrium between constrained (closed) and flexible (open) states. The 2′-OH group in flexible nucleotides preferentially adopts an open, reactive, conformation that facilitates reaction with electrophilic reagents to form a 2′-O-adduct (Figure 1). SHAPE experiments work well using electrophiles based on the isatoic anhydride (IA) scaffold.2a,3 Positions that form 2′-O-adducts are detected by primer extension.2
Figure 1.

Mechanistic framework for RNA SHAPE chemistry.
IA derivatives both react with the RNA 2′-OH group and also undergo concurrent degradation by hydrolysis (Figure 1). 2′-OH reactivity is thus conveniently monitored by allowing a reaction to proceed until the reagent has been consumed, either by hydrolysis or reaction with RNA. At this end point, the fraction adduct at any nucleotide (f) is
| (1) |
Where
| (2) |
and the rate of hydrolysis has been shown to be proportional to the rate of adduct formation,2b,3 kadduct/khydrolysis = β. These relationships lead to two limits. In limit 1, kopen + kclose ≫ kadduct[reagent]
| (3) |
For limit 2, kopen + kclose ≪ kadduct[reagent]
| (4) |
It should therefore be possible to monitor local nucleotide dynamics in RNA under conditions where limit 2 applies by varying the reactivity (or khydrolysis) of the hydroxyl-selective electrophile. IA has a hydrolysis half-life (t1/2) of 430 s at 37 °C. Electron-withdrawing substituents at the cyclic amine (R1) or in the benzene ring (R2) enhance reagent reactivity. Compared to IA, N-methyl isatoic anhydride (NMIA), 4-nitroisatioc anhydride (4NIA), and 1-methyl 7-nitroisatoic anhydride (1M7)3 have progressively shorter hydrolysis half-lives (table, Figure 1).
To investigate if distinct local nucleotide dynamics can be captured by varying the SHAPE electrophile, we focused on an important variation in RNA structure: the C2′-endo conformation. Although C2′-endo nucleotides are relatively rare, they are highly over-represented in important RNA tertiary interactions and in catalytic active sites.4 Local structure at tandem G•A mismatches depends on the local sequence context.5 Guanosine nucleotides in G•A pairs adopt the C2′-endo conformation in the sequences (UGAA)25a and (GGAU)2,5b the C3′-endo conformation typical of standard A-form helix geometry in (CGAG)2,5c and a mixture of C2′-endo/C3′-endo conformations in (UGAG)2.5b We constructed a simple hairpin RNA (termed the C2′-endo RNA) containing each of these sequences. Because the G•A mismatch-containing sequences are palindromic, there are two equivalent examples of each G•A pair in the RNA, including four total C2′-endo nucleotides (in red, Figure 2A).
Figure 2.
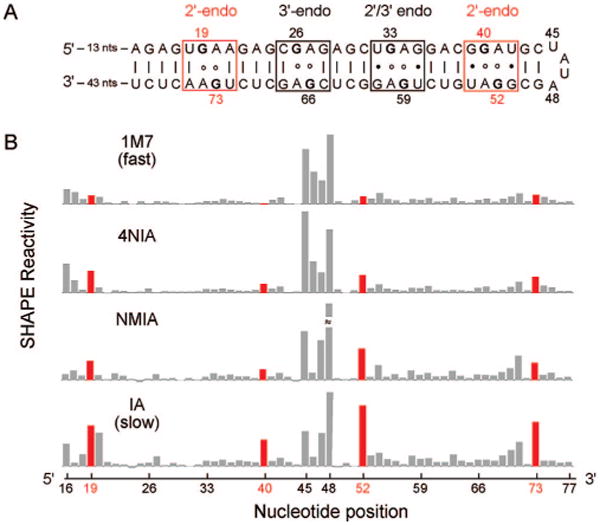
(A) The C2′-endo RNA construct contains tandem G•A base pairs that adopt either C2′- or C3′-endo ribose conformations. (B). Absolute SHAPE reactivities as a function of reagent electrophilicity.
When the C2′-endo RNA was subjected to SHAPE analysis using the fastest reagent, 1M7, flexible nucleotides in the apical loop (nts 45–48) are reactive, while positions constrained by base pairing are unreactive, regardless of the sugar pucker (top panel, Figure 2B). When an otherwise identical experiment was performed with IA, which reacts 30-fold more slowly, nucleotides in the apical loop were again reactive, while most of the base paired nucleotides were unreactive, similar to their reactivities with 1M7. In strong contrast, the four G nucleotides that adopt the C2′-endo conformation were highly reactive, even more so than some nucleotides in the flexible loop (Figure 2B, bottom panel; red bars at positions 19, 40, 52, 73). For the two reagents with intermediate reactivities, 4NIA and NMIA, the C2′-endo positions are moderately reactive (Figure 2B). Nucleotides constrained in these C2′-endo conformations are thus unreactive toward fast-reacting electro-philes but are highly reactive toward the slower reagents.
Limit 1 (eq 3) predicts that adduct formation is a function of the equilibrium constant for formation of the open state [kopen/(kopen + kclose)] and is independent of the reagent hydrolysis rate. Most nucleotides thus far analyzed by SHAPE, including in tRNA2,6 and in an RNase P RNA,3 are characterized by limit 1.
In contrast, the observation of a strong dependence of adduct formation on khydrolysis suggests that limit 2 applies to the C2′-endo nucleotides in Figure 2. Limit 2 also implies (1) that the extent of reaction at C2′-endo nucleotides will be independent of reagent concentration and (2) that kobs reports kopen (compare eqs 3 and 4). We analyzed the concentration dependence for reaction at positions 52 and 73 using isatoic anhydride and found, as predicted by limit 2, that adduct formation is independent of reagent concentration under conditions where reaction of the unconstrained model nucleotide, pAp-ethyl, showed a clear concentration dependence (Figure S1).
We therefore estimated kopen at the C2′-endo nucleotides at positions 19/73 and 40/52 by fitting the extent of 2′-O-adduct formation as a function of khydrolysis to eq 4 (Figure 3). In both cases, kopen is 4 × 10−5 s−1. In contrast, reactivities for both pAp-ethyl and flexible loop nucleotides are independent of khydrolysis (Figure S2).
Figure 3.

Determination of kopen for C2′-endo positions 19/73 and 40/52. Lines represent a fit to eq 4, with the addition of a plateau term; rate constants are 4 (±2) × 10−5 s−1. Error bars indicate standard deviations from independent measurements.
Critically, some C2′-endo nucleotides thus experience extraordinarily slow local dynamics to form conformations reactive toward isatoic anhydride-based electrophiles.
We next explored whether the differential reactivity between 1M7 and IA can be used to identify nucleotides that undergo slow conformational dynamics in an RNA with a complex structure, the specificity domain of the Bacillus subtilis ribonuclease P enzyme (RNase P).7 After excluding nucleotides where the electron density was not well-defined or that participate in crystal contacts, we identified 10 C2′-endo nucleotides in the RNase P RNA (in color, Figure 4A).
Figure 4.
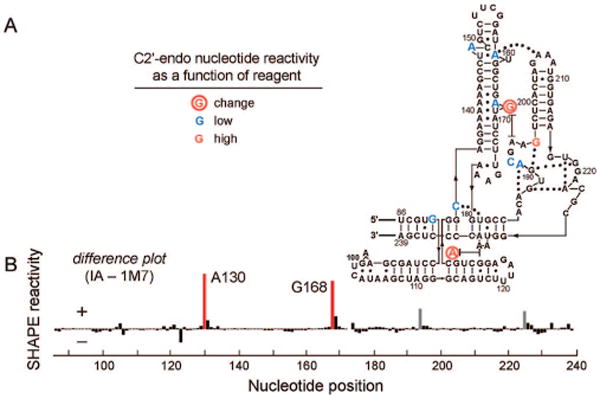
SHAPE reactivities at C2′-endo nucleotides in the specificity domain of RNase P. (A) Secondary structure showing SHAPE reactivities at the 10 well-defined C2′-endo nucleotides. (B) Differential SHAPE reactivities for slow (IA) versus fast (1M7) reagents.
For the vast majority of RNase P nucleotides, including all positions with C3′-endo conformations, SHAPE reactivities were identical for both the fast (1M7) and slow (IA) reagents (Figure S3). These nucleotides reflect limit 1. The 10 well-defined C2′-endo nucleotides fell into three categories: (i) most C2′-endo nucleotides are highly constrained and, as expected,2a unreactive toward both reagents (blue nucleotides, Figure 4A); (ii) one nucleotide is not constrained and is reactive toward both electrophiles (red nucleotide, Figure 4A); and (iii) two C2′-endo positions show large changes in reactivity (nts A130 and G168, circled nucleotides and red bars, Figure 4). Two other nucleotides showed smaller changes in reactivity but were in regions of the structure where experimental electron density was poorly defined (gray columns, Figure 4B). A similar distribution of reactive and unreactive C2′-endo nucleotides occurs in the Tetrahymena P5-P4-P6 domain using NMIA.8
While the C2′-endo conformation by itself clearly does not govern SHAPE reactivity, a distinct class of C2′-endo nucleotides experiences extraordinarily slow local dynamics. These nucleotides in both the simple C2′-endo RNA (Figure 2B) and the RNase P RNA (Figure 4B) share key characteristics: (1) the ribose group has the C2′-endo conformation, and (2) the nucleotide conformation is partially constrained by base stacking and hydrogen bonding interactions (Figure S4). These C2′-endo dynamics are orders of magnitude slower than for other local RNA conformational changes such as base opening reactions9 and are also slower than folding processes that involve assembly of whole domains in large RNAs.1a
An instructive precedent for slow conformational changes at a single residue in a biopolymer is the cis–trans isomerization of prolyl residues in proteins. Proline cis-trans conformations interconvert on the order of 10−2−10−5 s−1,10 and are thus comparable to the rates measured here for local dynamics at some C2′-endo nucleotides in RNA. The cis-trans isomerization can function as a molecular switch in biology.10c
We postulate that slow conformational dynamics at C2′-endo nucleotides also have the potential to function as rate-determining molecular switches and will play important, but currently unexplored, roles in RNA folding, ligand recognition, and catalysis.
Supplementary Material
Supporting Figure 1. Concentration dependence for reaction at positions 52 and 73 in the C2′-endo RNA construct and for the (unconstrained) model nucleotide pAp-ethyl.
Supporting Figure 2. Absence of a dependence of fraction adduct formed as a function of khydrolysis for reaction of the (unconstrained) model nucleotide pAp-ethyl and for nucleotide 45 in the loop of the C2′-endo construct.
Supporting Figure 3. Absolute SHAPE reactivities and difference plot for RNase P using slow (IA) and fast (1M7) reagents. Red columns indicate nucleotide positions that show the greatest difference in reactivity between the two reagents. Reactivites at all other positions are identical within experimental error.
Supporting Figure 4. Base stacking and hydrogen bonding interactions at C2′-endo nucleotides that undergo slow local conformational changes. (A) C40 from the C2′-endo RNA construct12 and (B) A130 and (C) G168 in the RNase P RNA. Distances are in Å.
Acknowledgments
We are indebted to Doug Turner and Blanton Tolbert for insightful discussions. This work was supported by grants from the NSF (MCB-0416941) and NIH (AI068462) to K.M.W.
Footnotes
Supporting Information Available: Methods and four figures. This material is available free of charge via the Internet at http://pubs.acs.org.
References
- 1.See Figure 12 of: Buchmueller KL, Hill BT, Platz MS, Weeks KM. J Am Chem Soc. 2003;125:10850–10861. doi: 10.1021/ja035743+.Shajani Z, Varani G. Biopolymers. 2007;86:348–359. doi: 10.1002/bip.20650.
- 2.(a) Merino EJ, Wilkinson KA, Coughlan JL, Weeks KM. J Am Chem Soc. 2005;127:4223–4231. doi: 10.1021/ja043822v. [DOI] [PubMed] [Google Scholar]; (b) Wilkinson KA, Merino EJ, Weeks KM. J Am Chem Soc. 2005;127:4659–4667. doi: 10.1021/ja0436749. [DOI] [PubMed] [Google Scholar]; (c) Wilkinson KA, Merino EJ, Weeks KM. Nat Protoc. 2006;1:1610–1616. doi: 10.1038/nprot.2006.249. [DOI] [PubMed] [Google Scholar]
- 3.Mortimer SA, Weeks KM. J Am Chem Soc. 2007;129:4144–4145. doi: 10.1021/ja0704028. [DOI] [PubMed] [Google Scholar]
- 4.Richardson JS, et al. RNA. 2008;14:465–481. doi: 10.1261/rna.657708. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.(a) Heus HA, Wijimenga SS, Hoppe H, Hilbers CW. J Mol Biol. 1997;271:147–158. doi: 10.1006/jmbi.1997.1158. [DOI] [PubMed] [Google Scholar]; (b) Tolbert BS, Kennedy SD, Schroeder SJ, Krugh TR, Turner DH. Biochemistry. 2007;46:1511–1522. doi: 10.1021/bi061350m. [DOI] [PMC free article] [PubMed] [Google Scholar]; (c) Santa Lucia JJ, Turner DH. Biochemistry. 1993;32:12612–12623. doi: 10.1021/bi00210a009. [DOI] [PubMed] [Google Scholar]
- 6.Wang B, Wilkinson KA, Weeks KM. Biochemistry. 2008;47:512–524. doi: 10.1021/bi702372x. [DOI] [PubMed] [Google Scholar]
- 7.Krasilnikov AS, Yang X, Pan T, Mondragon A. Nature. 2003;421:760–764. doi: 10.1038/nature01386. [DOI] [PubMed] [Google Scholar]
- 8.Vicens Q, Gooding AR, Laederach A, Cech TR. RNA. 2007;13:536–548. doi: 10.1261/rna.400207. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.(a) Leroy JL, Bolo N, Figueroa N, Plateau P, Gueron M. J Biomol Struct Dyn. 1985;2:915–939. doi: 10.1080/07391102.1985.10507609. [DOI] [PubMed] [Google Scholar]; (b) Spies MA, Schowen RL. J Am Chem Soc. 2002;124:14049–14053. doi: 10.1021/ja012272n. [DOI] [PubMed] [Google Scholar]
- 10.(a) Grathwohl C, Wuthrich K. Biopolymers. 1981;20:2623–2633. [Google Scholar]; (b) Schoetz G, Trapp O, Schurig V. Electrophoresis. 2001;22:2409–2415. doi: 10.1002/1522-2683(200107)22:12<2409::AID-ELPS2409>3.0.CO;2-C. [DOI] [PubMed] [Google Scholar]; (c) Lu KP, Finn G, Lee TH, Nicholson LK. Nat Chem Biol. 2007;3:619–629. doi: 10.1038/nchembio.2007.35. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supporting Figure 1. Concentration dependence for reaction at positions 52 and 73 in the C2′-endo RNA construct and for the (unconstrained) model nucleotide pAp-ethyl.
Supporting Figure 2. Absence of a dependence of fraction adduct formed as a function of khydrolysis for reaction of the (unconstrained) model nucleotide pAp-ethyl and for nucleotide 45 in the loop of the C2′-endo construct.
Supporting Figure 3. Absolute SHAPE reactivities and difference plot for RNase P using slow (IA) and fast (1M7) reagents. Red columns indicate nucleotide positions that show the greatest difference in reactivity between the two reagents. Reactivites at all other positions are identical within experimental error.
Supporting Figure 4. Base stacking and hydrogen bonding interactions at C2′-endo nucleotides that undergo slow local conformational changes. (A) C40 from the C2′-endo RNA construct12 and (B) A130 and (C) G168 in the RNase P RNA. Distances are in Å.