

Abstract
AIM: Hydrogen sulfide (H2S) is a prominent gaseous constituent of the gastrointestinal (GI) tract with known cytotoxic properties. Endogenous concentrations of H2S are reported to range between 0.2-3.4 mmol/L in the GI tract of mice and humans. Considering such high levels we speculate that, at non-toxic concentrations, H2S may interact with chemical agents and alter the response of colonic epithelium cells to such compounds. The GI tract is a major site for the absorption of phytochemical constituents such as isothiocyanates, flavonoids, and carotenoids, with each group having a role in the prevention of human diseases such as colon cancer. The chemopreventative properties of the phytochemical agent β-phenyethyl isothiocyanate (PEITC) are well recognized. However, little is currently known about the physiological or biochemical factors present in the GI tract that may influence the biological properties of ITCs. The current study was undertaken to determine the effects of H2S on PEITC mediated apoptosis in colon cancer cells.
METHODS: Induction of apoptosis by PEITC in human colon cancer HCT116 cells was assessed using classic apoptotic markers namely SubG1 population analysis, caspase-3 like activity and nuclear fragmentation and condensation coupled with the MTT [3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrasodium bromide] viability assay and LDH leakage.
RESULTS: PEITC significantly induced apoptosis in HCT116 cells as assessed by SubG1 population formation, nuclear condensation, LDH leakage and caspase-3 activity after 24 h, these data being significant from control groups (P < 0.01). In contrast, co-treatment of cells with physiological concentrations of H2S (0.1-1 mmol/L) prevented PEITC mediated apoptosis as assessed using the parameters described.
CONCLUSION: PEITC effectively induced cell death in the human adenocarcinoma cell line HCT116 in vitro through classic apoptotic mechanisms. However, in the presence of H2S, apoptosis was abolished. These data suggest that H2S may play a significant role in the response of colonic epithelial cells to beneficial as well as toxic agents present within the GI tract.
Keywords: Apoptosis, Colon cancer, Hydrogen sulfide, β-phenylethyl isothiocyanate
INTRODUCTION
Cruciferous vegetables such as broccoli, watercress and Bru-ssels sprouts are an important food source of phytochemicals known as glucosinolates (GSLs). Following tissue disruption GSLs can be converted by plant and/or enteric gut bacterial myrosinase to the organosulfur compounds isothiocyanates (ITCs)[1,2]. Exposure to ITCs can reduce carcinogen induced tumor formation in a diverse range of organs such as lung, mammary gland, esophagus, liver and intestine[3,4]. Supporting these findings are recent epidemiological studies showing that consumption of cruciferous vegetables, the primary source of ITCs in the human diet, can reduce cancer incidence in humans[5,6]. Due to the promising chemopreventative properties of crucifers and ITCs, numerous reports have attempted to determine the mechanism(s) of action of ITC. ITCs can inhibit phase I enzymes[7] and induce phase II detoxification enzymes[8-12]. Moreover, the induction of apoptosis in target tissues and cancer cells has also been demonstrated. Previous investigations have shown that β-phenylethyl isothiocyanate (PEITC), benzyl isothiocyanate (BITC), allyl isothiocyanate (AITC) and sulforaphane (MSB) can induce apoptosis in both in vitro and in vivo situations[13-19].
Considering the beneficial effects of ITC exposure in the diet, surprisingly few studies have placed much emphasis on endogenous or exogenous factors that may alter the beneficial properties of these phytochemical agents. To date, storage and cooking regimes, plant and bacterial myrosinase activity and altered conversion of GSLs to non-bioactive nitriles have been shown to influence the response of host tissues and cells to ITCs[20-24]. As ITCs are predominately absorbed along the gastrointestinal tract (GI) it is conceivable that the colonic environment may also contribute to the host’s response to ITCs.
Hydrogen sulfide (H2S) is a well known pungent gas and a prominent constituent present in the GI lumen. Exogenous sources show high toxicity towards the nervous, cardiovascular and respiratory systems in humans[25]. The toxic effect of H2S exposure are associated with its ability to bind reversibly to the heme site of cytochrome c aa3 site of cytochrome c oxidase thus inhibiting oxidative phosphorylation, although other mechanism(s) may also be involved. In addition, endogenous sites of H2S production also exist in mammals[26]. Catabolism of the amino acids L-cysteine and homocysteine by cystathionine-β-synthetase (CBS) generates appreciable levels of H2S in the brain. CBS is highly expressed in the hippocampus and cerebellum and CBS knockout mice have no detectable amounts of H2S in brain tissues. In contrast, H2S is also formed from the same precursors by cystathionine-β-lyase (CSE) which is the predominant enzyme in the vasculature. In addition, mammalian cells may also be exposed to high levels of H2S through the actions of enteric sulfate reducing bacteria in the GI tract[27]. Recent, studies have shown that H2S concentrations in the large intestine of mice ranges between 0.2-1 mmol/L and are comparable to the concentration of H2S in human feces (0.3-3.4 mmol/L)[28,29]. Considering the high in vivo levels of H2S it is plausible that such concentrations may directly affect the GI environment. Indeed, several clinical trials have shown a correlation between H2S levels and the development of colorectal cancer and ulcerative colitis, such studies highlighting a potential deleterious effect of H2S exposure[30]. However, it is not currently known whether H2S is the causative factor or a consequence of such diseases. In a more recent study Deplancke and Gaskins[31] demonstrated that H2S promotes cell cycle entry, MAPkinase activation, and cell proliferation in non-transformed intestinal IEC-18 cell in vitro. This data suggests a link between H2S exposure and cell cycle dysregulation which perhaps contributes to colon cancer and ulcerative colitis development. However, considering the potential toxic effects of H2S the GI tract has developed mechanism(s) to reduce the adverse effects of H2S such as increased rhodanese expression and activity, detoxification via oxidation to thiosulfate and bicarbonate secretion[32-34]. Therefore, H2S production in health subjects may not lead to disease formation due to adequate protective mechanisms. Recent studies have shown that H2S prevents glutamate induced oxidative stress in human neurons by stimulating glutathione biosynthesis[35]. H2S has also been shown to inhibit peroxynitrite, hydrogen peroxide and superoxide mediated cellular damage and death in neurons and myocytes by acting as an antioxidant[36,37]. Likewise, organisms like Saccharomyces cerevisiae and deep sea hydrothermal vent Archea use H2S for the detoxification of methylmercury and heavy metal ion (Cu, Zn and Co) induced toxicity[38,39], thus physiological concentrations in the human GI tract may protect colon cancer cells from undergoing apoptosis.
In view of the large quantities of H2S in the GI tract we have systematically evaluated whether H2S can alter the biological response of human colon cancer HCT116 cells to the chemopreventative agent PEITC. Such data may provide a novel insight as to how H2S, at non-toxic concentrations, may act indirectly on colonic cells apoptosis.
MATERIALS AND METHODS
Cells and chemicals
Human colon cancer HCT116, HT29, SW480 and LS174T cell lines were purchased from the American Culture Collection (Rockville, MD). HCA-7 cells were kindly supplied by Susan. C. Kirkland, Royal Postgraduate Medical School, UK. β-Phenylethyl isothiocyanate, DMSO, ATP, SDS, penicillin and streptomycin were purchased from Sigma-Aldrich (St. Louis, MO, USA); MEM, trypsin and FBS from GIBCO BRL (Gaithersburg, MD, USA).
Cell culture
HCT116, HT29, HCA-7 and LS174T cells were cultured in complete DMEM (containing 100 mL/L FBS, 100 000 U/L penicillin, 100 mg/L streptomycin, pH 7.4) in 75-cm culture flasks at 37°C in 50 mL/L CO2. Similarly, SW480 cells were grown in RPMI 1640 medium under the above cell culture conditions.
Cell culture treatment
NaSH was used to generate readily controlled amounts of H2S as it dissociates to Na+and HS- in solution, then HS-associates with H+ to produce H2S. We herein use the term H2S to reflect the sum of the species H2S, HS-or S2-present at physiological pH. PEITC was dissolved in DMSO to a final concentration of 100 mmol/L and adjusted to the required concentration by dilution in cell culture medium. All reagents were prepared fresh for each individual experiment.
Cell viability testing
Cell viability was determined using the MTT [3-(4, 5-dimethylthiazol-2-yl)-2, 5-diphenyl tetrasodium bromide] viability assay as previously described[40]. Activity of lactate dehydrogenase (LDH) in the medium was measured using an Abbott VP Biochemical Analyser with the test kit (Abbott Laboratories, Irving, TX, USA). The total LDH activity was determined by ultrasonication and assessed by expressing as percentage LDH leakage (LDH in medium/total LDH activity × 100).
Detection of nuclear morphological changes and apoptosis
To assess apoptotic cell death morphologic changes in chromatin structure were detected by DAPI staining. Briefly, attached cells were harvested washed with PBS, and re-suspended in PBS prior to fixing with 700 mL/L ethanol for 15 min at room temperature. Cells were centrifuged, re-suspended in PBS and stained with 2 g/L DAPI in the dark for 5 min. Cell suspensions were mounted on glass slides and subjected to fluorescence microscopic examination using a Nikon photomicroscope (Thornwood, NY, USA). Apoptotic cells were identified by their morphology and by the condensation and fragmentation of their nuclei. DNA fragmentation in HCT116 cells was measured using propodium iodide staining followed by flow cytometry analysis as described[41]. From each treatment 10 000 cells were analyzed using flow cytometry (Coulter Epics Elite ESP, Miami, FL, USA). Data was analyzed using WinMDI 2.7 software (Scripps Institute, La Jolla, CA, USA).
Measurement of caspase activity
Caspase-3 activity was determined spectrofluorometrically using the synthetic substrate Ac-DEVD-AFC respectively (BIOMOL Research Labs, Plymouth, PA, USA) as previously described[42,43]. Cell homogenates were incubated with synthetic substrate 0.2 mmol/L at 37°C/2 h. The fluorescence intensity of liberated AFC was monitored using a Tecan spectrofluor plus plate reader (USA) using an excitation wavelength of 400 nm and an emission wavelength of 505 nm.
Statistical analysis
All data are represented by three separate experiments. All experimental data consists of mean±SD unless otherwise stated and were analyzed by one-way ANOVA.
RESULTS
Evaluation of PEITC mediated reduction of colon cancer cell viability
Accumulating evidence has shown that cancer cells treated with PEITC undergo apoptosis by inducing mitochondrial dysfunction, sequential caspase activation, and MAPKinase mediated apoptotic signaling[14,16]. To evaluate the cytotoxic effects of PEITC on the viability of colon cancer cells, we preferentially screened several commonly available cell lines namely colorectal carcinoma HCT116, and colorectal adenocarcinoma cell lines HT29, SW480, HCA-7 and LS174T cells, following 24h treatments with increased concentrations of PEITC. Cell viability was determined by MTT assay. All cells lines were responsive to PEITC treatment with viability decreasing in a concentration dependent manner (Figure 1).
Figure 1.
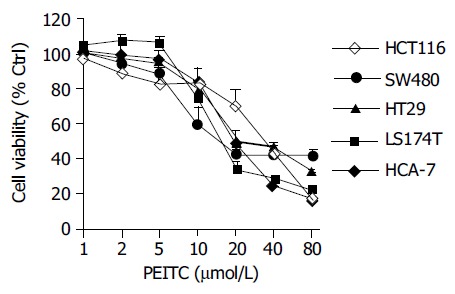
(A) Concentration dependant loss of cell viability induced by PEITC in colon cancer cells as determined at 24 h using the MTT viability assay. Data are representative of three or more separate experiments (mean±SD).
Induction of apoptosis by PEITC
We next selected two cell lines for further study these being the HCT116 and HT29 cell lines. Several key parameters that are known to be involved in PEITC induced apoptosis in HCT116 cells and HT29 cells were evaluated. Previous investigations have shown both HCT116 and HT29 cells are responsive to PEITC treatment, which we needed to reconfirm under our laboratory conditions[14,44]. As represented in Figure 2, PEITC mediated a time and concentration dependant induction of caspase-3 like activity in HCT116 and HT29 cells. Caspase activity was determined by measurement of AFC fluorescence intensity derived from the catalytic cleavage of the substrate Ac-DEVD-AFC by endogeneous caspase. The fluorescence intensity of treated over control cells, was used to determine the fold increase in caspase activity. Supporting a role of caspases in PEITC induced apoptosis was the inhibitory effects of the pharmacological caspase inhibitors Z-VAD-FMK (pan-caspase inhibitor), Ac-LEHD-CHO (caspase 9 inhibitor), and Ac-DEVD-CHO (caspase 3 inhibitor) on PEITC induced apoptosis. In both cell types tested all inhibitors reduced DNA fragmentation as determined by measurement of SubG1 populations by propodium iodide staining and LDH leakage (Figure 2).
Figure 2.
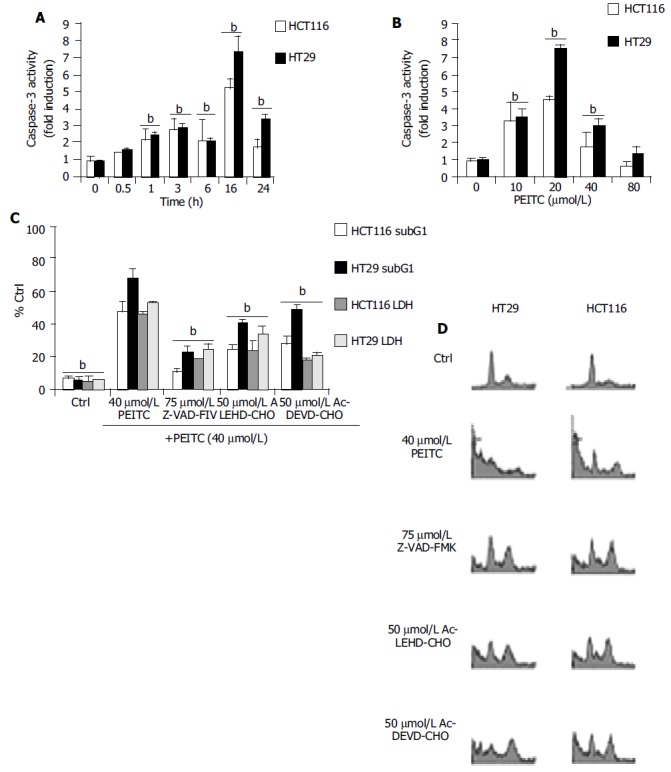
A: Time dependant Induction of caspase -3 activities in HCT116 and HT29 cells treated with 40 µmol/L PEITC as determined at 0.5, 1, 3, 6, 16 and 24 h. B: Concentration dependant activation of caspase-3 as determined at 24 h after incubation with PEITC (10-80 µmol/L). C: Z-VAD-FMK, Ac-LEHD-CHO and Ac-DEVD-CHO inhibition on PEITC mediated apoptosis as determined at 24 h. bP < 0.01, significant difference compared to control groups. Cells were treated with individual caspase inhibitors for 1 h prior to the addition of 40 µmol/L PEITC. SubG1 populations were determined by propodium iodide staining followed by flow cytometry analysis as described in the materials and methods. bP < 0.01, comparing PEITC (alone) with control groups. D: Representative flow cytometry analysis of HCT116 and HT29 cells was used to determine the extent of DNA fragmentation in cells treated with PEITC (40 µmol/L) in the presence of caspase inhibitors. Data were determined by PI staining at 24 h. Figure shows representative example of at least 3 experiments. In each analysis, 10 000 events were recorded. bP < 0.01, PEITC (alone) vs control and caspase inhibitor treated groups.
Inhibition of PEITC mediated apoptosis by low molecular weight sulfur compounds
Because PEITC is absorbed along the GI tract we next evaluated whether hydrogen sulfide and other low molecular weight sulfur compounds could protect HCT116 cells from PEITC mediated apoptosis. Incubation of HCT116 cells with up to 1 mmol/L concentrations of hydrogen sulfide (H2S) and its metabolic by-products sulfate (SO4), sulfite (SO3) and thiosulfate (S2O32-) had no appreciable effects on cell viability as determined using the MTT assay and LDH leakage at 24 h (Figure 3). Co-treatment of individual sulfur compounds (SO3, SO4, S2O32-) at 1 mmol/L concentrations failed to prevent the apoptotic effects of PEITC (40 µmol/L). In contrast, H2S co-treatment but not pre-treatment inhibited PEITC induced apoptosis in a concentration dependant manner that was comparable to the low molecular weight thiol N-acetylcysteine (Figure 3). Associated with the inhibitory effects of both H2S and NAC was a significant reduction in caspase activity (Figure 3D).
Figure 3.
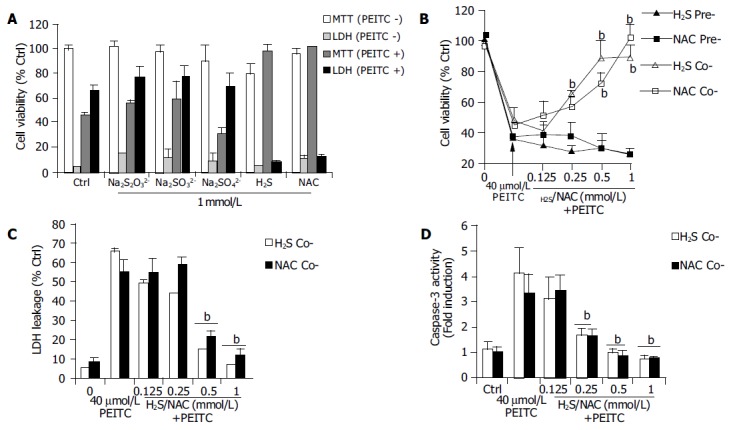
A: Effects of low molecular weight sulfur compounds on PEITC mediate loss of cell viability; B and C: Inhibition of PEITC mediated loss of cell viability in HCT116 cells following either co-treatment or pre-treatment (24 h) with H2S or NAC (125 µmol-1 mmol/L) as determined by MTT or LDH leakage. Data are expressed as mean±SD, (n = 8); D: Co-treatment with either H2S or NAC (125 µmol-1 mmol/L) inhibits PEITC induced caspase-3 activity as determined at 16 h and Data are representative of 3 or more separate experiments conducted on separate occasions. mean±SD, bP < 0.01 vs PEITC treated groups.
H2S and NAC protect against BSO mediated sensitization of HCT116 cells to PEITC induced apoptosis
Intracellular depletion of glutathione (GSH) can be achieved using buthionine-S-sulfoximine (BSO), an inhibitor of the rate limiting enzyme required for GSH biosynthesis. BSO treatment sensitizes cells to apoptotic agents by reducing intracellular GSH levels. The sensitizing effects of combined treatment of PEITC and BSO in HCT116 cells were next investigated in the presence of H2S and NAC. BSO at the concentration used had no appreciable effects on cell viability. Pre-treatment with 0.5 mmol/L BSO/24 h increased the apoptotic response of HCT116 cells to PEITC (40 µmol/L) as determined by the MTT assay and LDH leakage (Figure 4A). Indeed, BSO pre-treatment decreased cell viability by 83% and increased LDH leakage to 84.9% (P < 0.01) in PEITC treated cells. Cellular protection was afforded by co-treatment with either H2S or NAC (1 mmol/L). Moreover, evaluation of morphological and nuclear condensation also demonstrated that both H2S and NAC prevent BSO mediated sensitization of HCT116 cells to PEITC.
Figure 4.
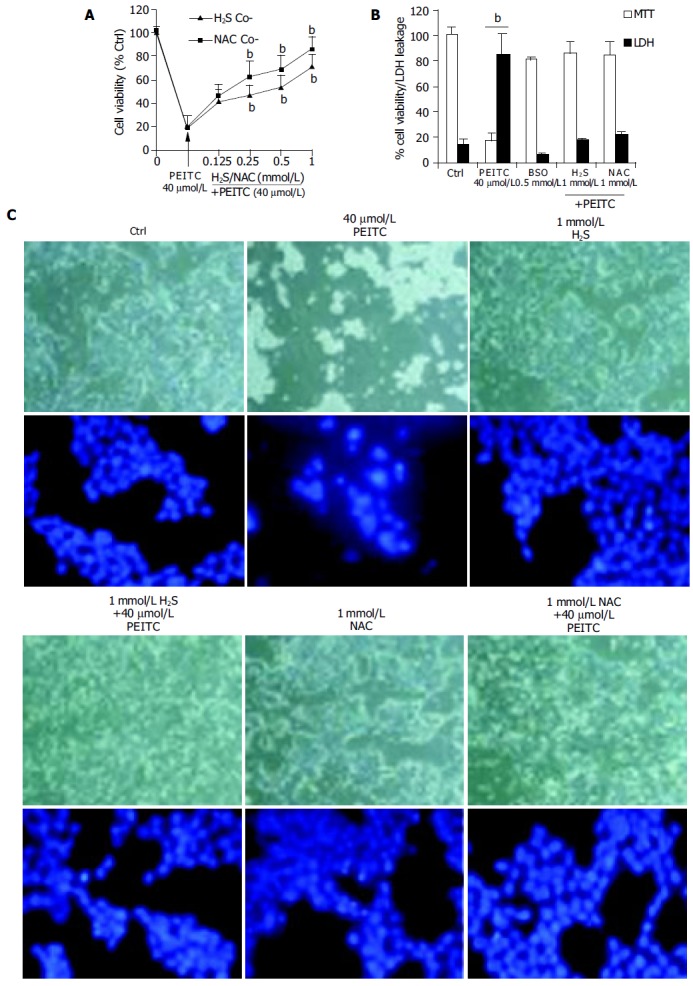
Inhibition of PEITC mediated loss in cell viability in GSH depleted cells following pre-treatment with 0.5 mmol/L BSO for 24 h. Both H2S and NAC inhibited PEITC induced loss of cell viability in concentration dependant manner as determined using the MTT assay (A) and (B) LDH leakage. (C) Respective
DISCUSSION
In the present investigation we sought to examine whether exogenous sources of the reductant H2S could alter the response of human colon cancer cells to β-phenylethyl isothiocyanate induced apoptosis. PEITC is known to possess potent chemopreventative properties against carcinogen induced colon cancer in rodent models, perhaps mediated largely by the induction of carcinogen detoxification pathways, anti-inflammatory affects, and apoptosis[8,11,18,19,45]. It is interesting to note, that previous investigations have shown that colon cancer cells are more sensitive to ITC mediated cytotoxicity than there detransformed counterparts. This property has been proposed to protect against the development of colorectal cancer by inhibiting the growth of transformed cell clones within the gastrointestinal mucosa[46,47]. However, factors that may limit these beneficial effects have been largely unexplored.
The biological properties of ITCs, including PEITC, are predominantly mediated by the electophilic central carbon atom of the isothiocyanate groups (R-N = C = S)[48]. The NCS residue readily reacts with nucleophilic centers of deprotonated thiols such as those present on glutathione (GSH) and N-acetylcysteine (NAC) leading to the formation of N-substituted mono-dithiocarbamate conjugates. Such interactions alter the intracellular redox state and activate phase II detoxification enzymes or apoptotic signaling pathways.
Considering that sulfur metabolizing bacteria generate appreciable levels of the thiol H2S in the GI tract, via metabolism of inorganic and organic sulfur compounds, we investigated whether H2S could act in a similar manner to GSH or NAC in the detoxification of PEITC. Early work conducted by Zuman and Zahradnik[49] demonstrated that organic ITCs can reversibly react with the hydrosulfide ion of H2S forming N-mono-substituted dithiocarbomates, such findings supporting our hypothesis. However, whether these interactions could change the biological properties of PEITC, as well as other ITCs were not explored. H2S concentrations in the GI tract range between 0.2-1 mmol/L in mice and up to 3.4 mmol/L in human feces, therefore chemical interaction between H2S and PEITC are highly plausible. On the basis of the chemical nature of H2S and its similar biological characteristics to that of both GSH and NAC e.g. antioxidant properties and a deprotonated thiol group in aqueous solution, perhaps H2S at physiological concentrations may impair the biological properties of electrohilic chemop-reventative agents against colon cancer cells.
In our study, PEITC reduced colon cancer cell viability in a concentration manner in all cell lines studied (Figure 1). In addition, we also confirmed that the loss of cell viability in HT29 and HCT116 cells was due to the induction of apoptosis as assessed by nuclear fragmentation and conde-nsation, LDH leakage, caspase-3 like activation and an increase in the SubG1 population (Figure 2). We next addressed using the HCT116 cell line whether H2S could impair PEITC induced apoptosis. In each assay examined H2S significantly reduced the apoptotic response of HCT116 cells to PEITC at concentrations well within the physiological reported levels (up to 3.4 mmol/L). Indeed, H2S was almost as effective on a molar basis at inhibiting PEITC induced apoptosis as the antioxidant thiol N-acetylcysteine, a known inhibitor of PEITC intracellular up-take[12]. We assume that the mechanism of inhibition by H2S is similar to NAC, although this requires further study.
Understanding the mechanisms by which PEITC mediates apoptosis and factors that may influence its in vivo efficacy is of considerable importance as PEITC mediated photographs and DAPI stained cells showing cell morphological changes and nuclear condensation. Data are representative of 3 or more separate experiments conducted on separate occasions. mean±SD, bP < 0.01 vs PEITC treated to control groups. apoptosis is thought to contribute to its chemopreventative effects. In particular, extensive induction in apoptosis by PEITC has been observed in cell culture studies and rodent models of colon cancer. Similarly, an inverse correlation between ITC exposure and a reduction in colon cancer prevalence has been demonstrated in some epidemiological studies. Whether elevated levels of H2S observed in colon cancer and ulcerative colitis impair chemopreventative or chemotherapeutic drug induced anti-inflammatory and apoptotic effects of has yet to be demonstrated.
In summary, our data reports for the first time that physiological concentrations of H2S can inhibit the apoptotic effects of the chemopreventative agent PEITC. It is possible based on our data that changes in H2S levels along the GI tract may influence the extent of PEITC mediated apoptosis in vivo. The potential interaction of H2S with other dietary constituents as well as chemotherapeutic drugs requires further study.
Footnotes
Science Editor Li WZ Language Editor Elsevier HK
References
- 1.Fahey JW, Zalcmann AT, Talalay P. The chemical diversity and distribution of glucosinolates and isothiocyanates among plants. Phytochemistry. 2001;56:5–51. doi: 10.1016/s0031-9422(00)00316-2. [DOI] [PubMed] [Google Scholar]
- 2.Rask L, Andréasson E, Ekbom B, Eriksson S, Pontoppidan B, Meijer J. Myrosinase: gene family evolution and herbivore defense in Brassicaceae. Plant Mol Biol. 2000;42:93–113. [PubMed] [Google Scholar]
- 3.Zhang Y, Talalay P. Anticarcinogenic activities of organic isothiocyanates: chemistry and mechanisms. Cancer Res. 1994;54:1976s–1981s. [PubMed] [Google Scholar]
- 4.Verhoeven DT, Verhagen H, Goldbohm RA, van den Brandt PA, van Poppel G. A review of mechanisms underlying anticarcinogenicity by brassica vegetables. Chem Biol Interact. 1997;103:79–129. doi: 10.1016/s0009-2797(96)03745-3. [DOI] [PubMed] [Google Scholar]
- 5.London SJ, Yuan JM, Chung FL, Gao YT, Coetzee GA, Ross RK, Yu MC. Isothiocyanates, glutathione S-transferase M1 and T1 polymorphisms, and lung-cancer risk: a prospective study of men in Shanghai, China. Lancet. 2000;356:724–729. doi: 10.1016/S0140-6736(00)02631-3. [DOI] [PubMed] [Google Scholar]
- 6.Zhao B, Seow A, Lee EJ, Poh WT, Teh M, Eng P, Wang YT, Tan WC, Yu MC, Lee HP. Dietary isothiocyanates, glutathione S-transferase -M1, -T1 polymorphisms and lung cancer risk among Chinese women in Singapore. Cancer Epidemiol Biomarkers Prev. 2001;10:1063–1067. [PubMed] [Google Scholar]
- 7.Conaway CC, Jiao D, Chung FL. Inhibition of rat liver cytochrome P450 isozymes by isothiocyanates and their conjugates: a structure-activity relationship study. Carcinogenesis. 1996;17:2423–2427. doi: 10.1093/carcin/17.11.2423. [DOI] [PubMed] [Google Scholar]
- 8.Rose P, Faulkner K, Williamson G, Mithen R. 7-Methylsulfinylheptyl and 8-methylsulfinyloctyl isothiocyanates from watercress are potent inducers of phase II enzymes. Carcinogenesis. 2000;21:1983–1988. doi: 10.1093/carcin/21.11.1983. [DOI] [PubMed] [Google Scholar]
- 9.Mithen R, Faulkner K, Magrath R, Rose P, Williamson G, Marquez J. Development of isothiocyanate-enriched broccoli, and its enhanced ability to induce phase 2 detoxification enzymes in mammalian cells. Theor Appl Genet. 2003;106:727–734. doi: 10.1007/s00122-002-1123-x. [DOI] [PubMed] [Google Scholar]
- 10.Zhang DD, Hannink M. Distinct cysteine residues in Keap1 are required for Keap1-dependent ubiquitination of Nrf2 and for stabilization of Nrf2 by chemopreventive agents and oxidative stress. Mol Cell Biol. 2003;23:8137–8151. doi: 10.1128/MCB.23.22.8137-8151.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Zhang Y, Talalay P. Mechanism of differential potencies of isothiocyanates as inducers of anticarcinogenic Phase 2 enzymes. Cancer Res. 1998;58:4632–4639. [PubMed] [Google Scholar]
- 12.Zhang Y. Molecular mechanism of rapid cellular accumulation of anticarcinogenic isothiocyanates. Carcinogenesis. 2001;22:425–431. doi: 10.1093/carcin/22.3.425. [DOI] [PubMed] [Google Scholar]
- 13.Gamet-Payrastre L, Li P, Lumeau S, Cassar G, Dupont MA, Chevolleau S, Gasc N, Tulliez J, Tercé F. Sulforaphane, a naturally occurring isothiocyanate, induces cell cycle arrest and apoptosis in HT29 human colon cancer cells. Cancer Res. 2000;60:1426–1433. [PubMed] [Google Scholar]
- 14.Hu R, Kim BR, Chen C, Hebbar V, Kong AN. The roles of JNK and apoptotic signaling pathways in PEITC-mediated responses in human HT-29 colon adenocarcinoma cells. Carcinogenesis. 2003;24:1361–1367. doi: 10.1093/carcin/bgg092. [DOI] [PubMed] [Google Scholar]
- 15.Rose P, Whiteman M, Huang SH, Halliwell B, Ong CN. beta-Phenylethyl isothiocyanate-mediated apoptosis in hepatoma HepG2 cells. Cell Mol Life Sci. 2003;60:1489–1503. doi: 10.1007/s00018-003-3150-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Rose P, Armstrong JS, Chua YL, Ong CN, Whiteman M. Beta-phenylethyl isothiocyanate mediated apoptosis; contribution of Bax and the mitochondrial death pathway. Int J Biochem Cell Biol. 2005;37:100–119. doi: 10.1016/j.biocel.2004.05.018. [DOI] [PubMed] [Google Scholar]
- 17.Xu K, Thornalley PJ. Studies on the mechanism of the inhibition of human leukaemia cell growth by dietary isothiocyanates and their cysteine adducts in vitro. Biochem Pharmacol. 2000;60:221–231. doi: 10.1016/s0006-2952(00)00319-1. [DOI] [PubMed] [Google Scholar]
- 18.Chung FL, Conaway CC, Rao CV, Reddy BS. Chemoprevention of colonic aberrant crypt foci in Fischer rats by sulforaphane and phenethyl isothiocyanate. Carcinogenesis. 2000;21:2287–2291. doi: 10.1093/carcin/21.12.2287. [DOI] [PubMed] [Google Scholar]
- 19.Smith TK, Mithen R, Johnson IT. Effects of Brassica vegetable juice on the induction of apoptosis and aberrant crypt foci in rat colonic mucosal crypts in vivo. Carcinogenesis. 2003;24:491–495. doi: 10.1093/carcin/24.3.491. [DOI] [PubMed] [Google Scholar]
- 20.Matusheski NV, Juvik JA, Jeffery EH. Heating decreases epithiospecifier protein activity and increases sulforaphane formation in broccoli. Phytochemistry. 2004;65:1273–1281. doi: 10.1016/j.phytochem.2004.04.013. [DOI] [PubMed] [Google Scholar]
- 21.Hwang ES, Jeffery EH. Effects of different processing methods on induction of quinone reductase by dietary broccoli in rats. J Med Food. 2004;7:95–99. doi: 10.1089/109662004322984770. [DOI] [PubMed] [Google Scholar]
- 22.Keck AS, Qiao Q, Jeffery EH. Food matrix effects on bioactivity of broccoli-derived sulforaphane in liver and colon of F344 rats. J Agric Food Chem. 2003;51:3320–3327. doi: 10.1021/jf026189a. [DOI] [PubMed] [Google Scholar]
- 23.Rouzaud G, Young SA, Duncan AJ. Hydrolysis of glucosinolates to isothiocyanates after ingestion of raw or microwaved cabbage by human volunteers. Cancer Epidemiol Biomarkers Prev. 2004;13:125–131. doi: 10.1158/1055-9965.epi-085-3. [DOI] [PubMed] [Google Scholar]
- 24.Rouzaud G, Rabot S, Ratcliffe B, Duncan AJ. Influence of plant and bacterial myrosinase activity on the metabolic fate of glucosinolates in gnotobiotic rats. Br J Nutr. 2003;90:395–404. doi: 10.1079/bjn2003900. [DOI] [PubMed] [Google Scholar]
- 25.U.S. Environmental Protection Agency EPA/635/R-03/005 [www document]. URL http: //www.epa.gov/iris [accessed 1 November 2003] [Google Scholar]
- 26.Kamoun P. Endogenous production of hydrogen sulfide in mammals. Amino Acids. 2004;26:243–254. doi: 10.1007/s00726-004-0072-x. [DOI] [PubMed] [Google Scholar]
- 27.Deplancke B, Finster K, Graham WV, Collier CT, Thurmond JE, Gaskins HR. Gastrointestinal and microbial responses to sulfate-supplemented drinking water in mice. Exp Biol Med (Maywood) 2003;228:424–433. doi: 10.1177/153537020322800413. [DOI] [PubMed] [Google Scholar]
- 28.Florin T, Neale G, Gibson GR, Christl SU, Cummings JH. Metabolism of dietary sulphate: absorption and excretion in humans. Gut. 1991;32:766–773. doi: 10.1136/gut.32.7.766. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Magee EA, Richardson CJ, Hughes R, Cummings JH. Contribution of dietary protein to sulfide production in the large intestine: an in vitro and a controlled feeding study in humans. Am J Clin Nutr. 2000;72:1488–1494. doi: 10.1093/ajcn/72.6.1488. [DOI] [PubMed] [Google Scholar]
- 30.Huycke MM, Gaskins HR. Commensal bacteria, redox stress, and colorectal cancer: mechanisms and models. Exp Biol Med (Maywood) 2004;229:586–597. doi: 10.1177/153537020422900702. [DOI] [PubMed] [Google Scholar]
- 31.Deplancke B, Gaskins HR. Hydrogen sulfide induces serum-independent cell cycle entry in nontransformed rat intestinal epithelial cells. FASEB J. 2003;17:1310–1312. doi: 10.1096/fj.02-0883fje. [DOI] [PubMed] [Google Scholar]
- 32.Picton R, Eggo MC, Merrill GA, Langman MJ, Singh S. Mucosal protection against sulphide: importance of the enzyme rhodanese. Gut. 2002;50:201–205. doi: 10.1136/gut.50.2.201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Furne J, Springfield J, Koenig T, DeMaster E, Levitt MD. Oxidation of hydrogen sulfide and methanethiol to thiosulfate by rat tissues: a specialized function of the colonic mucosa. Biochem Pharmacol. 2001;62:255–259. doi: 10.1016/s0006-2952(01)00657-8. [DOI] [PubMed] [Google Scholar]
- 34.Roediger WE, Lawson MJ, Kwok V, Grant AK, Pannall PR. Colonic bicarbonate output as a test of disease activity in ulcerative colitis. J Clin Pathol. 1984;37:704–707. doi: 10.1136/jcp.37.6.704. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Kimura Y, Kimura H. Hydrogen sulfide protects neurons from oxidative stress. FASEB J. 2004;18:1165–1167. doi: 10.1096/fj.04-1815fje. [DOI] [PubMed] [Google Scholar]
- 36.Whiteman M, Armstrong JS, Chu SH, Jia-Ling S, Wong BS, Cheung NS, Halliwell B, Moore PK. The novel neuromodulator hydrogen sulfide: an endogenous peroxynitrite 'scavenger'? J Neurochem. 2004;90:765–768. doi: 10.1111/j.1471-4159.2004.02617.x. [DOI] [PubMed] [Google Scholar]
- 37.Geng B, Chang L, Pan C, Qi Y, Zhao J, Pang Y, Du J, Tang C. Endogenous hydrogen sulfide regulation of myocardial injury induced by isoproterenol. Biochem Biophys Res Commun. 2004;318:756–763. doi: 10.1016/j.bbrc.2004.04.094. [DOI] [PubMed] [Google Scholar]
- 38.Ono B, Ishii N, Fujino S, Aoyama I. Role of hydrosulfide ions (HS-) in methylmercury resistance in Saccharomyces cerevisiae. Appl Environ Microbiol. 1991;57:3183–3186. doi: 10.1128/aem.57.11.3183-3186.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Edgcomb VP, Molyneaux SJ, Saito MA, Lloyd K, Böer S, Wirsen CO, Atkins MS, Teske A. Sulfide ameliorates metal toxicity for deep-sea hydrothermal vent archaea. Appl Environ Microbiol. 2004;70:2551–2555. doi: 10.1128/AEM.70.4.2551-2555.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Hansen MB, Nielsen SE, Berg K. Re-examination and further development of a precise and rapid dye method for measuring cell growth/cell kill. J Immunol Methods. 1989;119:203–210. doi: 10.1016/0022-1759(89)90397-9. [DOI] [PubMed] [Google Scholar]
- 41.Nicoletti I, Migliorati G, Pagliacci MC, Grignani F, Riccardi C. A rapid and simple method for measuring thymocyte apoptosis by propidium iodide staining and flow cytometry. J Immunol Methods. 1991;139:271–279. doi: 10.1016/0022-1759(91)90198-o. [DOI] [PubMed] [Google Scholar]
- 42.Thornberry NA, Rano TA, Peterson EP, Rasper DM, Timkey T, Garcia-Calvo M, Houtzager VM, Nordstrom PA, Roy S, Vaillancourt JP, et al. A combinatorial approach defines specificities of members of the caspase family and granzyme B. Functional relationships established for key mediators of apoptosis. J Biol Chem. 1997;272:17907–17911. doi: 10.1074/jbc.272.29.17907. [DOI] [PubMed] [Google Scholar]
- 43.Garcia-Calvo M, Peterson EP, Leiting B, Ruel R, Nicholson DW, Thornberry NA. Inhibition of human caspases by peptide-based and macromolecular inhibitors. J Biol Chem. 1998;273:32608–32613. doi: 10.1074/jbc.273.49.32608. [DOI] [PubMed] [Google Scholar]
- 44.Powolny A, Takahashi K, Hopkins RG, Loo G. Induction of GADD gene expression by phenethylisothiocyanate in human colon adenocarcinoma cells. J Cell Biochem. 2003;90:1128–1139. doi: 10.1002/jcb.10733. [DOI] [PubMed] [Google Scholar]
- 45.Jeong WS, Kim IW, Hu R, Kong AN. Modulatory properties of various natural chemopreventive agents on the activation of NF-kappaB signaling pathway. Pharm Res. 2004;21:661–670. doi: 10.1023/b:pham.0000022413.43212.cf. [DOI] [PubMed] [Google Scholar]
- 46.Musk SR, Johnson IT. Allyl isothiocyanate is selectively toxic to transformed cells of the human colorectal tumour line HT29. Carcinogenesis. 1993;14:2079–2083. doi: 10.1093/carcin/14.10.2079. [DOI] [PubMed] [Google Scholar]
- 47.Musk SR, Stephenson P, Smith TK, Stening P, Fyfe D, Johnson IT. Selective toxicity of compounds naturally present in food toward the transformed phenotype of human colorectal cell line HT29. Nutr Cancer. 1995;24:289–298. doi: 10.1080/01635589509514418. [DOI] [PubMed] [Google Scholar]
- 48.Podhradský D, Drobnica L, Kristian P. Reactions of cysteine, its derivatives, glutathione coenzyme A, and dihydrolipoic acid with isothiocyanates. Experientia. 1979;35:154–155. doi: 10.1007/BF01920581. [DOI] [PubMed] [Google Scholar]
- 49.Zahradnik R, Zuman P. Kinetics and decomposition mecha-nism of dithiocarbamic acids solutions. Polarographic study. Z Phys Chem. 1957;208:135–140. [Google Scholar]