

Abstract
Coptis chinensis rhizomes (CR) are one important ingredient of traditional Chinese herbal formulas such as San-Huang-Xie-Xin-Tang which is used for treatment of cardiovascular and neurodegenerative diseases. Recent studies suggest that the extract of CR might be a potential therapeutic agent for amelioration of neurological disorders associated with oxidative stress. In the present study we aimed at revealing the main active compound(s) of the CR extract and at investigating the mechanism of action. Four main alkaloids of the CR extract (berberine, coptisine, jatrorrhizine, and palmatine) were selected for this study. Results showed that out of those alkaloids only pretreatment with coptisine significantly attenuated tert-butylhydroperoxide induced reduction of cell viability, increased rate of apoptosis, and declined mitochondrial membrane potential. Elisa assay and quantitative real-time PCR analyses revealed that thioredoxin-interacting protein (TXNIP) gene expression was downregulated by coptisine, which could explain the neuroprotective effect, hypothetically, by strengthening the thioredoxin defense system against oxidative stress and attenuation of apoptosis signal-regulating kinase (Ask1) mediated apoptotic signaling. A comparison between coptisine and CR extract identified coptisine as the main single component responsible for the neuroprotective effect. Based on the results the CR extract and coptisine are promising candidate agents for prevention or improvement of diabetic neuropathy and neurodegenerative disorders.
1. Introduction
Coptis chinensis (Franch) rhizomes (CR), commonly known as Coptidis rhizoma or “Huang Lian,” has been used in Traditional Chinese Medicine (TCM) since ancient times and was recommended by famous physicians in TCM history like Li Shizhen, Tang Shenwei, and Tao Hongjing for inflammatory diseases [1].
CR is a main ingredient of multiple historical prescriptions, for example, San-Huang-xie-xin-tang (SHXT), which originated in the Ming dynasty. SHXT is composed of Coptis chinensis (Franch) (Coptidis rhizoma), Rheum officinale Baill (Rhei rhizoma), and Scutellaria baicalensis Georgi (Scutellariae radix) [2]. Recent studies showed that SHXT has neuroprotective properties due to its anti-inflammatory and antioxidative effects [3–5]. It has been shown that one of the main ingredients of SHXT, the dried root CR also known as Chinese goldthread (huang-lian in Chinese), is effective for the treatment of neurodegenerative disease associated with oxidative stress [4, 6, 7]. Some of its single compounds showed neuroprotective [8], neuroregenerative [9], anti-apoptotic [10], and antioxidative [6] effects which strongly point out that CR is one main component in decoctions used for the treatment of oxidative stress associated with neurodegenerative disease.
Reactive oxygen species (ROS) have a significant impact on the development of neurodegenerative disease like Alzheimer or Parkinson's disease [11–15]. Cells including neurons are usually well-protected from ROS-induced cytotoxicity by the endogenous antioxidant system. However, if the oxidative stress exceeds the antioxidative capacity of this system it can lead to deoxyribonucleic acid (DNA) demethylation, histone acetylation, oxidative protein and lipid modification, increase of intracellular calcium ions (Ca2+), depolarization of the mitochondrial membrane potential (MMP), and release of cytochrome C into the cytosol and, as a consequence, to apoptosis or necrosis [15–19]. For this reason, therapeutic strategies targeting ROS-induced cytotoxicity are needed and could have a major impact on the treatment of neurodegenerative diseases, which are associated with oxidative stress.
We recently showed that 2 h and 24 h pretreatment of SH-SY5Y neuroblastoma cells with the watery extract of CR (CRE) significantly attenuated tert-butylhydroperoxide- (t-BOOH-) induced cytotoxicity [20]. Further statistical analysis revealed that 24 h pretreatment with CRE was more effective than 2 h pretreatment.
The present study aimed at revealing the main active compound of CRE, which is responsible for the cytoprotective effect of the extract, and at comparing the effectiveness of the single compounds with the whole extract.
2. Materials and Methods
2.1. Drugs and Reagents
tert-Butylhydroperoxide (t-BOOH), thiazolyl blue tetrazolium bromide (MTT), and dimethyl sulfoxide (DMSO) were purchased from Sigma (Taufkirchen, Germany). 2′,7′-Dichlorodihydrofluorescein diacetate (H2DCFH-DA), Mitotracker Red CMX Ros, and Hoechst 33342 were obtained from Life Technologies (Darmstadt, Germany). Berberine (Ber), coptisine (Cop), jatrorrhizine (Jat), and palmatine (Pal) were ordered from Cfm Oskar Tropitzsch (Markdredwitz, Germany). All other reagents were purchased from Roth (Karlsruhe, Germany).
2.2. Herbal Preparation
Coptis chinensis (Franch) was obtained from China Medica (Ch. B. 930034; 83684 Tegernsee, Germany), as dried rhizome. Identity and purity were confirmed according to the Pharmacopoeia of the People's Republic of China [21]. Sebastian Kneipp research laboratory for residue analysis and organic trace analysis (Bad Wörishofen, Germany) certified that heavy metal, pesticide, and microbiological contamination were below the guideline of the Pharmacopoeia Europaea [22] and Regulation (EC) Number 396/2005 of the European Commission.
Coptis chinensis extract was prepared as described previously [20]. 10 g of grounded rhizome was boiled in 100 mL distilled deionized water (DDW) for 30 minutes and the extract was centrifuged afterwards. Supernatant was collected and the residue was extracted a second time with 100 mL DDW. Combined supernatants were dried with a rotary-vacuum evaporator (60°C, 200 mbar; Rotavapor-R, Büchi) and a vacuum concentrator (Bachofer). Dried extracts were stored at −20°C until use.
2.3. HPLC-Analysis
High-performance liquid chromatography (HPLC) analysis was performed according to the method described previously [20]. Briefly, chromatographic separation was conducted on an Alltima C18 (250 mm × 4.6 mm × 5 μm, S/N: 213100139, temperature: 25°C) column with 0.1% trifluoroacetic acid (A) and acetonitrile (B) as mobile phase and at a flow rate of 1 mL/min. Berberine (Ber), coptisine (Cop), jatrorrhizine (Jat), and palmatine (Pal) were used as reference standard compounds.
2.4. Cell Culture
Human neuroblastoma SH-SY5Y cells were cultivated in RPMI 1640 medium containing 10% fetal calf serum, 100 U/mL penicillin, and 100 μg/mL streptomycin. Cells were grown in a humid atmosphere of 5% CO2 and 95% air at 37°C. All cell culture reagents were obtained from Sigma (Taufkirchen, Germany).
2.5. Cell Viability Measurements
Cell viability was determined by the MTT assay. Ber, Cop, Jat, and Pal stock solutions were diluted in medium to their final concentration and sterilely filtrated, and different concentrations were added at the start of the incubation time for 24 h. Afterwards, cells were incubated for 2 h with 100 μM t-BOOH. Medium containing t-BOOH was removed, cells were washed with Dulbecco's phosphate-buffered saline (DPBS), and 1 mM MTT solution was added for 2 h. Subsequently, 100 μL 2-Propanol was added and the plate was agitated for 1 h at 450 revolutions per minute (rpm) at room temperature (RT). Absorption was measured 3 times at 570 nm.
2.6. Quantification of ROS
Reactive oxygen species in the cells were measured with the 2′,7′-dichlorodihydrofluorescein diacetate probe (H2DCFH-DA). SH-SY5Y cells were seeded into a 96-well microplate (4∗104 cells/well) and incubated with 100 μg/mL CRE or 20 μM Cop for 24 hours. Afterwards, fresh medium containing 20 μM H2DCFH-DA was added for 30 min at 37°C in the dark. Subsequently, cells were washed with DPBS and medium containing 100 μM t-BOOH was added. Fluorescence was measured every 10 minutes for 120 minutes (excitation: 485 nm; emission: 528 nm).
2.7. Quantification of Apoptotic Nuclei and Mitochondrial Membrane Potential
Apoptotic nuclei and mitochondrial membrane potential were measured using Mitotracker Red CMX Ros and Hoechst 33342. Cells were stained first with 25 nM Mitotracker Red CMX Ros for 1 hour, followed by 20 minutes fixation with 4% paraformaldehyde (PFA) and 10 minutes Hoechst 33342 (4 μM) staining. Images were captured with a Leica microscope and an Axiovision camera. To determine MMP, the intensity sum of the Mitotracker Red CMX Ros fluorescence was measured for each cell. Apoptosis was detected by analyzing the morphology of the Hoechst 33342 stained nuclei. Experiments were repeated 3 times and at least 1600 cells were analyzed for each group.
2.8. RT-PCR
Total RNA was isolated with the RNeasy MINI Kit (Quiagen; Hilden; Germany) according to the manufacturer's instructions. Reverse transcription was carried out with the high capacity RNA-to-cDNA Kit (Applied Biosystems). For semiquantitative analysis, LightCycler 480 SYBR Green 1 Master (Roche) and the following human specific primers were used: TXNIP 5′-GATCACCGATTGGAGAGCCC-3′ and 5′-TGCAGGGATCCACCTCAGTA-3′; GAPDH 5′-GCATCTTCTTTTGCGTCGCC-3′ and 5′-CCCAATACGACCAAATCCGTTG-3′. Obtained data were analyzed as previously described [23].
2.9. Human TXNIP Elisa Assay
Cells were seeded in T-25 flasks (2.5 × 106 cells/flask) and treated for 24 hours with CRE or Cop. Total proteins were isolated with mammalian cell lysis reagent (CellLytic M) according to the manufacturer's instructions and stored at −80°C until use.
Protein concentration was determined with Roti-Quant following the protocol provided by the manufacturer. Human TXNIP protein concentration was measured with the CircuLex Human TXNIP ELISA Kit (MBL international cooperation, Biozol, Germany) following the instructions of the provided protocol. Experiments were repeated three times and each sample was measured in duplicates.
2.10. Statistics
Data are presented as means ± standard error of the mean (SEM) of n experiments. Statistical significance between groups was determined with OriginLab pro 8.5 by ANOVA, followed by the Bonferroni Post Hoc Test. P < 0.05 was considered as statistically significant.
3. Results
3.1. HPLC Analysis
Watery extraction of CR yields 21.5% by weight of the dried herb. The CRE was composed of 554.9 ± 14.6 mg/g Ber, 60.6 ± 0.5 mg/g Cop, and 51.9 ± 0.2 mg/g Pal. Representative high-performance liquid chromatogram of the extract and of the mixed standard compounds is shown in Figure 1.
Figure 1.
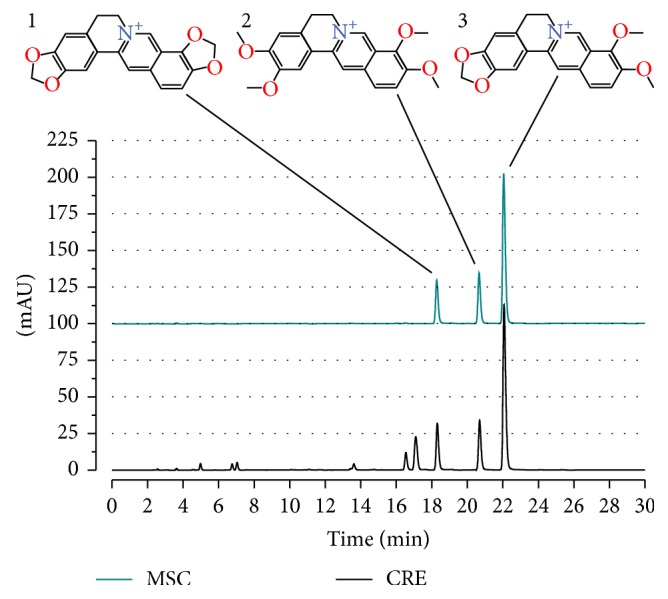
High-performance liquid chromatogram of the watery extract from Coptis chinensis (Franch). Blue line represents the HPLC chromatogram of mixed standard compounds (MSC); 1: coptisine, 2: palmatine, 3: berberine. Note that coptisine, berberine, and palmatine are the main components of the CRE. ACD/ChemSketch (http://www.acdlabs.com/resources/freeware/chemsketch/) was used to design chemical structures.
3.2. Effect of CRE and CR Main Alkaloids on Cell Viability
To investigate the neuroprotective effect of CR main alkaloids against t-BOOH-induced toxicity, we first examined if those single components exhibited any cytotoxic effect in the concentration range between 0.1 and 40 μM. Results showed no significant cytotoxic effect of Ber, Cop, Jat, and Pal in comparison to the medium control (P < 0.01, Figure 2). Treatment of cells with 100 μM t-BOOH resulted in significant decrease of cell viability to 53.2 ± 1.7% (P < 0.01; versus medium control). Pretreatment of the cells with Ber, Jat, and Pal (0.1–40 μM) before t-BOOH-induced oxidative stress showed no significant protective effect compared to the t-BOOH control (Figures 2(a), 2(c), and 2(d)). However, pretreatment with 1–40 μM Cop for 24 hours resulted in a significant increase of cell viability from 53.2 ± 1.7% (t-BOOH control) up to 65.6 ± 2.6% (40 μM, Figure 2(b)).
Figure 2.

Effect of CR main alkaloids on cell viability in SH-SY5Y cells. Cells were pretreated for 24 h with (0–40 μM) of Ber (a), Cop (b), Jat (c), and Pal (d) before the cells were exposed for 2 h to either medium (medium control) or 100 μM t-BOOH (t-BOOH control). Results represent mean cell viability ± SEM of four independent experiments conducted in triplicates. Only treatment with Cop resulted in a significant increase of cell viability. ∗ P < 0.05, ∗∗ P < 0.01 versus medium or t-BOOH control.
Previously, we showed that 24-hour pretreatment with the watery extract of CR had a significant protective effect against t-BOOH-induced oxidative stress, whereas 100 μg/mL was most effective [20]. To investigate if Cop is responsible for the previously reported protective effect of CRE, we compared 24-hour pretreatment of CRE (100 μg/mL) with Cop (20 μM). Results revealed that CRE increased cell viability to 71.9 ± 2.3% and Cop to 64.6 ± 1.9% (Figure 3). Statistical analyses showed that CRE is significantly more effective than Cop (P < 0.05). However, 80% of the cell viability increase of CRE could be achieved by Cop (Figure 3).
Figure 3.
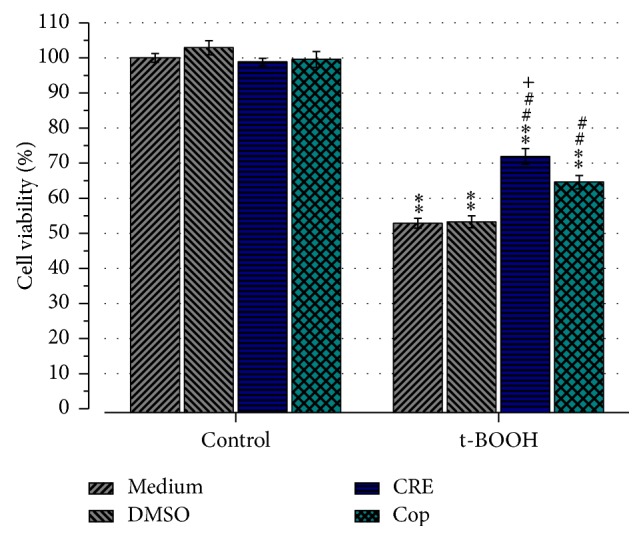
Effect of CRE and Cop on cell viability in SH-SY5Y cells. Cells were pretreated for 24 h with CRE (100 μg/mL) or Cop (20 μM). Note that treatment with Cop achieves 80% of the effect of CRE. ∗∗ P < 0.01 versus medium or DMSO control. ## P < 0.01 versus t-BOOH-medium or t-BOOH-DMSO. + P < 0.05 versus t-BOOH-Cop.
3.3. Effect of CRE and Cop on Intracellular Reactive Oxygen Production
To investigate the antioxidative effect of CRE and Cop, we analyzed the production of intracellular reactive oxygen species induced by t-BOOH treatment. The mean dichlorofluorescein (DCF) fluorescence of medium control cells was set to 100%. Results showed that both CRE and Cop had no effect on the baseline DCF fluorescence. Two-hour exposure to 100 μM t-BOOH produced a significant increase in DCF fluorescence to 252.1 ± 6.7% (P < 0.01; t-BOOH control). Furthermore, results revealed that only CRE significantly reduced mean DCF fluorescence to 227.7 ± 2.5% (P < 0.5 versus t-BOOH control, Figure 4).
Figure 4.

Effect of CRE (100 μg/mL) and Cop (20 μM) on DCF fluorescence was investigated in SH-SY5Y cells. Increase in DCF fluorescence indicates increased intracellular ROS. (a) shows the evolution of ROS generation; t-BOOH (upper 4 lines) or PBS (lower 4 lines) was added at the start of the measurements. Results are expressed as mean DCF fluorescence ± SEM normalized to the medium control of three independent experiments which were conducted in triplicate. (b) represents the mean sum of the DCF fluorescence signal normalized to the medium control, which was set to 100%. Note that only pretreatment with CRE significantly reduces mean DCF fluorescence. ∗∗ P < 0.01 versus control. + P < 0.05 versus Cop.
3.4. Effect of CRE and Cop on Apoptotic Rate
In order to investigate the effect of CRE and Cop on t-BOOH-induced apoptosis we analyzed the morphology of the nucleus. Results showed that the apoptotic rate in both medium and DMSO control groups was 5.3 ± 0.5% and 6.2 ± 0.6%, respectively. Two hours of t-BOOH treatment increased apoptotic rate in the medium and DMSO control group significantly to 46.3 ± 1.2% and 47.1 ± 1.5% (P < 0.01, Figure 5). Twenty-four hour pretreatment with CRE (100 μg/mL) or Cop (20 μM) reduced apoptosis significantly to 28.0 ± 1.3% and 36.5 ± 1.4%, respectively (P < 0.01). Statistical analysis revealed that CRE attenuates t-BOOH-induced apoptosis significantly more than Cop (P < 0.05).
Figure 5.
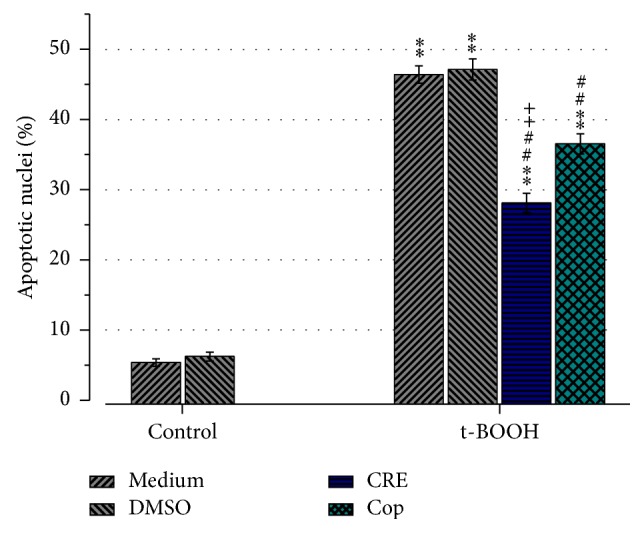
Effect of CRE (100 μg/mL) or Cop (20 μM) on t-BOOH-induced apoptosis. Results represent the percentage of apoptotic nuclei ± SEM from the total number of counted nuclei. 3 independent experiments were conducted and 1600 cells were analyzed at least in each group. Results showed that both COP and CRE significantly attenuate t-BOOH-induced apoptosis. ∗∗ P < 0.01 in comparison to the control (no treatment); ## P < 0.01 versus t-BOOH-medium or t-BOOH-DMSO and ++ P < 0.01 versus t-BOOH-Cop.
3.5. Effect of CRE and Cop on Mitochondrial Membrane Potential
Since it was previously reported that oxidative stress induced by t-BOOH leads to opening of the mitochondrial permeability transition (MPT) pore and loss of mitochondrial membrane potential [18, 19, 24], we investigated if CRE or Cop could attenuate the loss of mitochondrial membrane potential (MMP) in SH-SY5Y cells. Results showed that 2-hour exposure of SH-SY5Y cells with 100 μM t-BOOH significantly reduced the fluorescence signal to 44.2 ± 2.1% (t-BOOH-medium control) and 49.2 ± 1.5% (t-BOOH-DMSO control) in comparison to the medium control (P > 0.01), which was set to 100% (Figure 6). Pretreatment of the cells for 24 hours with 100 μg/mL CRE or 20 μM Cop significantly increased the MMP compared to the t-BOOH medium control to 78.2 ± 4.3% and 67.9 ± 1.7%, respectively (P < 0.01). Comparison between the CRE and Cop group showed that CRE was significantly more effective than Cop (P > 0.05).
Figure 6.
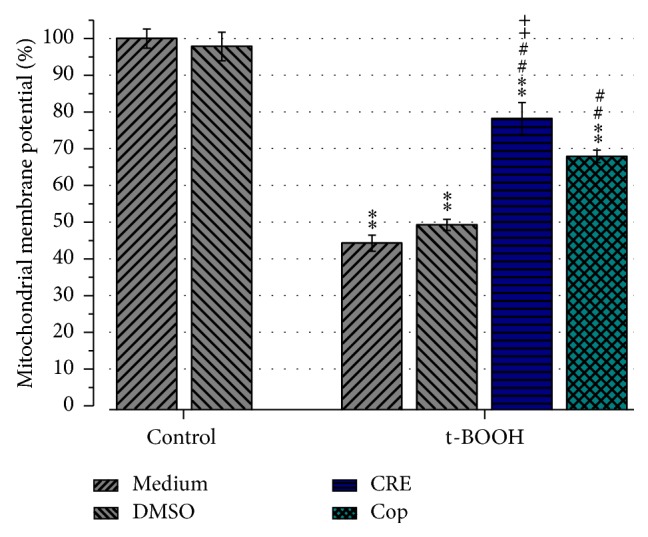
Effect of 24 h pretreatment with CRE (100 μg/mL) and Cop (20 μM) on mitochondrial membrane potential. Pretreated cells were challenged with oxidative stress induced by 2 h t-BOOH treatment. The mean MMP ± SEM was normalized to the medium control which was set to 100%. Experiments were repeated twice and for each group the MMP of 1600 cells were measured at least. Pre-treatment of the cells with CRE and Cop significantly increased MMP compared to the t-BOOH-medium control. ∗∗ P < 0.01 versus control; ## P < 0.01 versus t-BOOH-medium or t-BOOH-DMSO and ++ P < 0.01 versus t-BOOH-Cop.
3.6. Effect of CRE and Cop on TXNIP Expression
QRT-PCR results confirmed that treatment of the cells with CRE or Cop leads to a significant downregulation of TXNIP compared with the control group to 49.5 ± 3.8% and 71.9 ± 3.9%, respectively (P < 0.01; Figure 7). A comparison between CRE and Cop groups revealed that the downregulation of TXNIP was significantly pronounced in the CRE group (P < 0.05).
Figure 7.
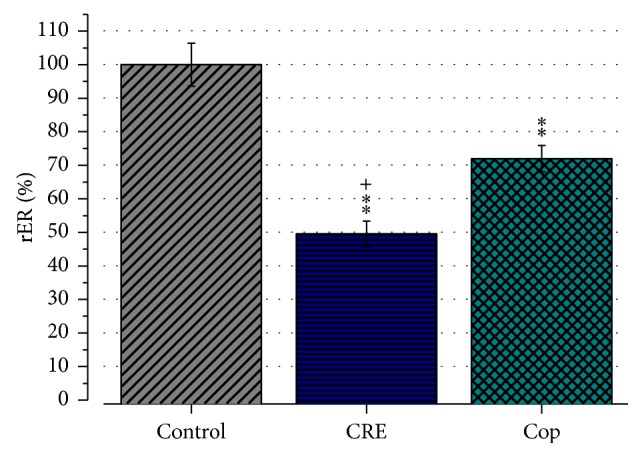
Effect of CRE (100 μg/mL) and Cop (20 μM) on TXNIP gene expression. Results represent mean relative gene expression ratio (rER) ± SEM. Experiments were performed in triplicates and repeated twice. Data were normalized to the nontreatment control which was set to 100%. Note that treatment of the cells with CRE or Cop leads to a significant downregulation of TXNIP. ∗∗ indicates P < 0.01 versus control. + P < 0.05 versus Cop.
Furthermore, results of the TXNIP protein expression revealed that the TXNIP concentration in the control, CRE, and Cop group was 1189.2 ± 86.6 pg/mg total protein, 689.4 ± 53.2 pg/mg total protein, and 958.7 ± 70.4 pg/mg total protein, respectively (Figure 8). Statistical analysis showed that both CRE and Cop reduced the TXNIP protein concentration significantly when compared to the control (P < 0.01 and P < 0.05) and that CRE was significantly more effective than Cop (P < 0.01).
Figure 8.
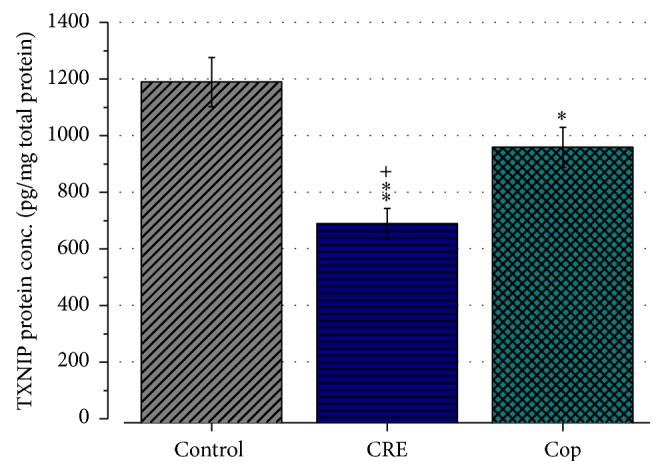
TXNIP protein concentration of 1 mg extracted total protein. Results represent mean TXNIP concentration ± SEM of three independent experiments preformed in duplicates. TXNIP protein concentration was significantly reduced by 24 h CRE (100 μg/mL) and Cop (20 μM) treatment. ∗ P < 0.05, ∗∗ P < 0.01 in comparison to the control; and + P < 0.05 versus Cop.
4. Discussion
In recent years many studies focused on cellular injuries related to oxidative stress and natural antioxidants with neuroprotective potential. A plurality of the investigated natural herbal extracts or single compounds exert their protective effect via removal of surplus ROS or by the prevention of ROS generation. However, it turned out to be that many natural antioxidants, even if they show some beneficial effect in cell culture experiments, have no or just limited effect in humans [25]. Therefore, other natural pharmaceuticals which act via a different mechanism to protect the cells from oxidative damage are needed.
In this study we investigated the neuroprotective effect against t-BOOH-induced oxidative damage of the main compounds of CR, a very important herb in Chinese medicine, which is used in many different herbal prescriptions.
Our results confirmed that 2-hour treatment of SH-SY5Y cells with t-BOOH resulted in a significant cell viability reduction with a strong dose-dependency. There was no attenuation of the cell viability loss detectable by pretreatment with different concentrations of Ber, Jat, and Pal. However, pretreatment with Cop resulted in a dose dependent reduction of t-BOOH-induced cytotoxicity by improvement of the mitochondrial membrane potential and attenuation of apoptosis. A comparison with the cytoprotective effect of CRE revealed that pretreatment with Cop in a comparable concentration, as in the CRE, achieved approximately 80% of the effect of the CR watery extracts. This suggests that Cop alone is not the only compound responsible for the neuroprotective effect; however, it is the main active compound. It has been shown previously that the extract of Coptis chinensis includes the following compounds: magnoflorine, groenlandicine, berberastine, demethyleneberberine, lycoranine B, jatrorrhizine/columbamine, epiberberine, coptisine, thalifendine/berberrubine, palmatine, berberine and dihydrochelerythrine [26–29]. In this study, we used only 4 alkaloids of the extract, namely, berberine, coptisine, jatrorrhizine, and palmatine. Therefore, we hypothesize that the cytoprotective effect of CR against t-BOOH-induced oxidative stress could be imitated by the combination of two or more single compounds.
It was previously reported by Drahota et al. that treatment with t-BOOH leads to a reduction of the MMP and increases apoptosis in vitro [30]. Therefore, we tested if Cop pretreatment attenuates the reduction of the MMP as well as the increased apoptotic rate and compared the results with the effect of the CRE. Our results revealed that pretreatment of SH-SY5Y cells with Cop or CRE improved the MMP compared to the t-BOOH control and that the CRE was significantly more effective than Cop. The same finding also applies to the reduction of t-BOOH-induced apoptosis by Cop and CRE. Furthermore, these findings strengthen the hypothesis that the cytoprotective effect of the CRE could only be explained by a combination of different individual compounds. Nonetheless, Cop seems to be the main active component which protects the cells against t-BOOH-induced oxidative stress.
Based upon these results, it is suggested that the cytoprotective effect is at least partly achieved by improved mitochondrial function and reduction of apoptosis.
To verify if the neuroprotective effect of CRE and COP against t-BOOH-induced oxidative stress could be explained furthermore by the antioxidative properties of the extract we investigated the antioxidative effect of CRE and Cop with the cell-based H2DCFH-DA assays. The results showed that only the CRE showed a slight antioxidative effect. However, this effect was too small to explain the present neuroprotective effect of the CRE. The results suggest that the amount of antioxidative compounds in the extract or the concentration of Cop was too low to have a significant influence on the ROS generation induced by the rather high dose of t-BOOH.
We previously reported that genome-wide transcriptome analysis on Human Genome U219 microarrays (Affymetrix, SantaClara, USA) of SH-SY5Y cells revealed that 24 h treatment with CRE significantly regulated only the expression of two genes: mitochondrially encoded NADH dehydrogenase 1 (MTND1) and TXNIP [20]. In this work we hypothesized that downregulation of TXNIP, a 50-kDa protein which belongs to the α-arrestin protein family, could be at least partly responsible for the neuroprotective effect of CRE on t-BOOH-induced oxidative damage. It is well-known that TXNIP is an inhibitor of thioredoxin (Trx), which is essential for intracellular responses to ROS [31–33]. In case of oxidative stress TXNIP binds to Trx, which prevents Trx in playing its major role as antioxidant, in DNA synthesis and repair, in cellular signaling and transcription control, and in inhibition of apoptotic pathways [31–36].
Therefore, downregulation of TXNIP by CRE or Cop could reduce the amount of inactivated Trx, resulting in an attenuation of apoptotic signaling in the cell [37, 38]. This suggests that downregulation of TXNIP could protect SH-SY5Y cells from apoptosis, improve DNA repair as well as cellular signaling, and strengthen oxidative defense system during high oxidative stress. Our results revealed that both CRE and Cop decreased TXNIP, mRNA, and protein concentration. The reduction of TXNIP, at least for Cop, seems to be too low to explain the present neuroprotective effect of Cop on t-BOOH-induced oxidative stress. However, the reduction of TXNIP expression should have an impact on cellular ROS level. Yoshihara et al. reported that downregulation of TXNIP attenuates the inhibition of the Trx redox cycle by TXNIP and this results in a reduction of cellular oxidative stress [39]. Since our results showed only a small reduction of ROS by CRE and no effect of Cop, one might speculate that the present downregulation of TXNIP was not sufficient enough to have a significant influence on the ROS level generated by the relatively high t-BOOH concentration used in this study. Gao et al. showed that inhibition of TXNIP expression by activation of AMP-activated protein kinase protects podocytes from oxidative stress induced injury, without any influence of the cellular ROS level [38]. They hypothesized that a reduction of TXNIP expression leads to an attenuation of the ASK1-P58 signaling pathway which protects the cells from oxidative stress induced injuries. Nevertheless, further studies should clarify if downregulation of TXNIP by CRE and Cop also leads to a attenuation of the ASK1-P58 signaling pathway in SH-SY5Y cells and whether this mechanism is involved in the neuroprotective effect of CRE and Cop against oxidative stress induced cytotoxicity.
5. Conclusion
In conclusion, we showed that pretreatment of SH-SY5Y neuroblastoma cells with Cop attenuated t-BOOH-induced decrease in cell viability, reduction of the MMP, and increase of apoptosis. Further analysis showed that the protective effect of CRE against t-BOOH-induced cytotoxicity was more pronounced than the effect of Cop. Nonetheless, we could achieve approximately 80% of the effectiveness of CRE by treatment with Cop. This leads us to the conclusion that Cop is the main active single component of CRE for the prevention of t-BOOH-induced oxidative stress cell injury and that the protective effect might be based on an improvement of the mitochondrial function as well as on the attenuation of proapoptotic pathways, hypothetically, the ASK1-P58 signaling pathway.
This research work provided evidence that CRE and Cop might have potential therapeutic value for the treatment of neurodegenerative diseases associated with oxidative stress, like Alzheimer's disease or Parkinson's disease.
Acknowledgments
The study was supported by the Innovationsstiftung Hamburg and the HanseMerkur Versicherungsgruppe. The authors would like to thank PD Dr. M. Jendrach (Experimental Neurology, Department of Neurology, Goethe University Hospital, Frankfurt am Main, Germany) for the SH-SY5Y cells and Christine Knies (Institute of Anatomy and Experimental Morphology, University Medical Center Hamburg-Eppendorf, Hamburg, Germany) for her technological support. The authors also acknowledge Professor Dr. C. Lohr (Division of Animal Physiology, University of Hamburg; Hamburg, Germany) for his support in form and content.
Abbreviations
- Ask1:
Apoptosis signal-regulating kinase
- Ber:
Berberine
- Ca2+:
Calcium ion
- Cop:
Coptisine
- CR:
Coptis chinensis rhizomes
- CRE:
Coptis chinensis rhizomes watery extract
- DCF:
Dichlorofluorescein
- DDW:
Distilled deionized water
- DMSO:
Dimethyl sulfoxide
- DNA:
Deoxyribonucleic acid
- DPBS:
Dulbecco's phosphate-buffered saline
- H2DCFH-DA:
2′,7′-Dichlorodihydrofluorescein diacetate
- HPLC:
High-performance liquid chromatography
- Jat:
Jatrorrhizine
- MMP:
Mitochondrial membrane potential
- MSC:
Mixed standard compounds
- MTT:
Thiazolyl blue tetrazolium bromide
- Pal:
Palmatine
- PFA:
Paraformaldehyde
- qRT-PCR:
Quantitative real-time-PCR
- ROS:
Reactive oxygen species
- rpm:
Revolutions per minute
- RT:
Room temperature
- SEM:
Standard Error of the Mean
- SHXT:
San-Huang-xie-xin-tang
- t-BOOH:
tert-Butylhydroperoxide
- TCM:
Traditional Chinese Medicine
- Trx:
Thioredoxin.
Conflict of Interests
The authors declare that there is no conflict of interests regarding the publication of this paper.
References
- 1.Li S. Bencao Gangmu—Compendium of Materia Medica. Vol. 2. Beijing, China: Foreign Language Press; 1999. [Google Scholar]
- 2.Dong S. K. J. Ming and Qing Chinese Masterpieces Series: Miraculous Recipe. Beijing, China: Traditional Chinese Medicine Press; 2000. First Published 1449, (Chinese) [Google Scholar]
- 3.Liou S.-F., Hsu J.-H., Liang J.-C., et al. San-Huang-Xie-Xin-Tang protects cardiomyocytes against hypoxia/reoxygenation injury via inhibition of oxidative stress-induced apoptosis. Journal of Natural Medicines. 2012;66(2):311–320. doi: 10.1007/s11418-011-0592-0. [DOI] [PubMed] [Google Scholar]
- 4.Lo Y. C., Shih Y. T., Tseng Y. T., Hsu H. T. Neuroprotective effects of san-huang-xie-xin-tang in the MPP+/MPTP models of Parkinson's disease in vitro and in vivo. Evidence-Based Complementary and Alternative Medicine. 2012;2012:10. doi: 10.1155/2012/501032.501032 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Shih Y.-T., Chen I.-J., Wu Y.-C., Lo Y.-C. San-Huang-Xie-Xin-Tang protects against activated microglia- and 6-OHDA-induced toxicity in neuronal SH-SY5Y cells. Evidence-Based Complementary and Alternative Medicine. 2011;2011:11. doi: 10.1093/ecam/nep025.429384 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Gong L.-L., Fang L.-H., Wang S.-B., et al. Coptisine exert cardioprotective effect through anti-oxidative and inhibition of RhoA/Rho kinase pathway on isoproterenol-induced myocardial infarction in rats. Atherosclerosis. 2012;222(1):50–58. doi: 10.1016/j.atherosclerosis.2012.01.046. [DOI] [PubMed] [Google Scholar]
- 7.Jung H. A., Min B.-S., Yokozawa T., Lee J.-H., Kim Y. S., Choi J. S. Anti-Alzheimer and antioxidant activities of coptidis rhizoma alkaloids. Biological and Pharmaceutical Bulletin. 2009;32(8):1433–1438. doi: 10.1248/bpb.32.1433. [DOI] [PubMed] [Google Scholar]
- 8.Luo T., Zhang H., Zhang W.-W., et al. Neuroprotective effect of Jatrorrhizine on hydrogen peroxide-induced cell injury and its potential mechanisms in PC12 cells. Neuroscience Letters. 2011;498(3):227–231. doi: 10.1016/j.neulet.2011.05.017. [DOI] [PubMed] [Google Scholar]
- 9.Han A. M., Heo H., Kwon Y. K. Berberine promotes axonal regeneration in injured nerves of the peripheral nervous system. Journal of Medicinal Food. 2012;15(4):413–417. doi: 10.1089/jmf.2011.2029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Miura N., Yamamoto M., Ueki T., Kitani T., Fukuda K., Komatsu Y. Inhibition of thymocyte apoptosis by berberine. Biochemical Pharmacology. 1997;53(9):1315–1322. doi: 10.1016/s0006-2952(97)87955-5. [DOI] [PubMed] [Google Scholar]
- 11.Finkel T., Holbrook N. J. Oxidants, oxidative stress and the biology of ageing. Nature. 2000;408(6809):239–247. doi: 10.1038/35041687. [DOI] [PubMed] [Google Scholar]
- 12.Perfeito R., Cunha-Oliveira T., Rego A. C. Reprint of: revisiting oxidative stress and mitochondrial dysfunction in the pathogenesis of Parkinson disease—resemblance to the effect of amphetamine drugs of abuse. Free Radical Biology and Medicine. 2013;62:186–201. doi: 10.1016/j.freeradbiomed.2013.05.042. [DOI] [PubMed] [Google Scholar]
- 13.Perry G., Cash A. D., Smith M. A. Alzheimer disease and oxidative stress. Journal of Biomedicine and Biotechnology. 2002;2(3):120–123. doi: 10.1155/s1110724302203010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Smith M. A., Perry G., Richey P. L., et al. Oxidative damage in Alzheimer's. Nature. 1996;382(6587):120–121. doi: 10.1038/382120b0. [DOI] [PubMed] [Google Scholar]
- 15.Vincent A. M., Russell J. W., Low P., Feldman E. L. Oxidative stress in the pathogenesis of diabetic neuropathy. Endocrine Reviews. 2004;25(4):612–628. doi: 10.1210/er.2003-0019. [DOI] [PubMed] [Google Scholar]
- 16.Amoroso S., Gioielli A., Cataldi M., di Renzo G., Annunziato L. In the neuronal cell line SH-SY5Y, oxidative stress-induced free radical overproduction causes cell death without any participation of intracellular Ca2+ increase. Biochimica et Biophysica Acta—Molecular Cell Research. 1999;1452(2):151–160. doi: 10.1016/s0167-4889(99)00110-x. [DOI] [PubMed] [Google Scholar]
- 17.Martín C., Martínez R., Navarro R., Ruiz-Sanz J. I., Lacort M., Ruiz-Larrea M. B. tert-butyl hydroperoxide-induced lipid signaling in hepatocytes: involvement of glutathione and free radicals. Biochemical Pharmacology. 2001;62(6):705–712. doi: 10.1016/s0006-2952(01)00704-3. [DOI] [PubMed] [Google Scholar]
- 18.Nieminen A.-L., Saylor A. K., Tesfai S. A., Herman B., Lemasters J. J. Contribution of the mitochondrial permeability transition to lethal injury after exposure of hepatocytes to t-butylhydroperoxide. Biochemical Journal. 1995;307(part 1):99–106. doi: 10.1042/bj3070099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Zhao K., Luo G., Giannelli S., Szeto H. H. Mitochondria-targeted peptide prevents mitochondrial depolarization and apoptosis induced by tert-butyl hydroperoxide in neuronal cell lines. Biochemical Pharmacology. 2005;70(12):1796–1806. doi: 10.1016/j.bcp.2005.08.022. [DOI] [PubMed] [Google Scholar]
- 20.Friedemann T., Otto B., Klätschke K., et al. Coptis chinensis Franch. exhibits neuroprotective properties against oxidative stress in human neuroblastoma cells. Journal of Ethnopharmacology. 2014;155(1):607–615. doi: 10.1016/j.jep.2014.06.004. [DOI] [PubMed] [Google Scholar]
- 21.Pharmacopeia C. C. Pharmacopoeia of the People's Republic of China 2010. China Medical Science Press; 2010. [Google Scholar]
- 22.European Pharmacopoeia Commission and Council of Europe European Directorate for the Quality of Medicines (EDQM) European Pharmacopoeia. 8th. Deutscher Apotheker; 2014. [Google Scholar]
- 23.Schefe J. H., Lehmann K. E., Buschmann I. R., Unger T., Funke-Kaiser H. Quantitative real-time RT-PCR data analysis: current concepts and the novel ‘gene expression's CT difference’ formula. Journal of Molecular Medicine. 2006;84(11):901–910. doi: 10.1007/s00109-006-0097-6. [DOI] [PubMed] [Google Scholar]
- 24.Nieminen A. L., Byrne A. M., Herman B., Lemasters J. J. Mitochondrial permeability transition in hepatocytes induced by t- BuOOh: NAD(P)H and reactive oxygen species. American Journal of Physiology—Cell Physiology. 1997;272(4, part 1):C1286–C1294. doi: 10.1152/ajpcell.1997.272.4.C1286. [DOI] [PubMed] [Google Scholar]
- 25.Halliwell B. Free radicals and antioxidants—quo vadis? Trends in Pharmacological Sciences. 2011;32(3):125–130. doi: 10.1016/j.tips.2010.12.002. [DOI] [PubMed] [Google Scholar]
- 26.Chuang W.-C., Young D.-S., Liu L. K., Sheu S.-J. Liquid chromatographic-electrospray mass spectrometric analysis of Coptidis Rhizoma. Journal of Chromatography A. 1996;755(1):19–26. doi: 10.1016/s0021-9673(96)00591-2. [DOI] [Google Scholar]
- 27.Ding P.-L., Chen L.-Q., Lu Y., Li Y.-G. Determination of protoberberine alkaloids in Rhizoma Coptidis by ERETIC 1H NMR method. Journal of Pharmaceutical and Biomedical Analysis. 2012;60:44–50. doi: 10.1016/j.jpba.2011.10.030. [DOI] [PubMed] [Google Scholar]
- 28.Jiang X., Huang L.-F., Wu L.-B., Wang Z.-H., Chen S.-L. UPLC-QTOF/MS analysis of alkaloids in traditional processed Coptis chinensis franch. Evidence-Based Complementary and Alternative Medicine. 2012;2012:6. doi: 10.1155/2012/942384.942384 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Wang D., Liu Z., Guo M., Liu S. Structural elucidation and identification of alkaloids in Rhizoma Coptidis by electrospray ionization tandem mass spectrometry. Journal of Mass Spectrometry. 2004;39(11):1356–1365. doi: 10.1002/jms.727. [DOI] [PubMed] [Google Scholar]
- 30.Drahota Z., Křiváková P., Červinková Z., et al. Tert-butyl hydroperoxide selectively inhibits mitochondrial respiratory-chain enzymes in isolated rat hepatocytes. Physiological Research. 2005;54(1):67–72. doi: 10.33549/physiolres.930578. [DOI] [PubMed] [Google Scholar]
- 31.Lu J., Holmgren A. Thioredoxin system in cell death progression. Antioxidants & Redox Signaling. 2012;17(12):1738–1747. doi: 10.1089/ars.2012.4650. [DOI] [PubMed] [Google Scholar]
- 32.Ma Q. Transcriptional responses to oxidative stress: pathological and toxicological implications. Pharmacology & Therapeutics. 2010;125(3):376–393. doi: 10.1016/j.pharmthera.2009.11.004. [DOI] [PubMed] [Google Scholar]
- 33.Nordberg J., Arnér E. S. J. Reactive oxygen species, antioxidants, and the mammalian thioredoxin system. Free Radical Biology and Medicine. 2001;31(11):1287–1312. doi: 10.1016/S0891-5849(01)00724-9. [DOI] [PubMed] [Google Scholar]
- 34.Lu J., Chew E.-H., Holmgren A. Targeting thioredoxin reductase is a basis for cancer therapy by arsenic trioxide. Proceedings of the National Academy of Sciences of the United States of America. 2007;104(30):12288–12293. doi: 10.1073/pnas.0701549104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Saitoh M., Nishitoh H., Fujii M., et al. Mammalian thioredoxin is a direct inhibitor of apoptosis signal-regulating kinase (ASK) 1. The EMBO Journal. 1998;17(9):2596–2606. doi: 10.1093/emboj/17.9.2596. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Saxena G., Chen J., Shalev A. Intracellular shuttling and mitochondrial function of thioredoxin-interacting protein. The Journal of Biological Chemistry. 2010;285(6):3997–4005. doi: 10.1074/jbc.m109.034421. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Al-Gayyar M. M. H., Abdelsaid M. A., Matragoon S., Pillai B. A., El-Remessy A. B. Thioredoxin interacting protein is a novel mediator of retinal inflammation and neurotoxicity. British Journal of Pharmacology. 2011;164(1):170–180. doi: 10.1111/j.1476-5381.2011.01336.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Gao K., Chi Y., Sun W., Takeda M., Yao J. 5′-AMP-activated protein kinase attenuates adriamycin-induced oxidative podocyte injury through thioredoxin-mediated suppression of the apoptosis signal-regulating kinase 1-P38 signaling pathway. Molecular Pharmacology. 2014;85(3):460–471. doi: 10.1124/mol.113.089458. [DOI] [PubMed] [Google Scholar]
- 39.Yoshihara E., Chen Z., Matsuo Y., Masutani H., Yodoi J. Thiol redox transitions by thioredoxin and thioredoxin-binding protein-2 in cell signaling. Methods in enzymology. 2010;474:67–82. doi: 10.1016/S0076-6879(10)74005-2. [DOI] [PubMed] [Google Scholar]