

Abstract
Adolescence is a critical period for brain maturation characterized by the reorganization of interacting neural networks. In particular the prefrontal cortex, a region involved in executive function, undergoes synaptic and neuronal pruning during this time in both humans and rats. Our laboratory has previously shown that rats lose neurons in the medial prefrontal cortex (mPFC) and there is an increase in white matter under the frontal cortex between adolescence and adulthood. Female rats lose more neurons during this period, and ovarian hormones may play a role as ovariectomy before adolescence prevents neuronal loss. However, little is known regarding the timing of neuroanatomical changes that occur between early adolescence and adulthood. In the present study, we quantified the number of neurons and glia in the male and female mPFC at multiple time points from preadolescence through adulthood (postnatal days 25, 35, 45, 60 and 90). Females, but not males, lost a significant number of neurons in the mPFC between days 35 and 45, coinciding with the onset of puberty. Counts of GABA immunoreactive cell bodies indicated that the neurons lost were not primarily GABAergic. These results suggest that in females, pubertal hormones may exert temporally specific changes in PFC anatomy. As expected, both males and females gained white matter under the prefrontal cortex throughout adolescence, though these gains in females were diminished after day 35, but not in males. The differences in cell loss in males and females may lead to differential vulnerability to external influences and dysfunctions of the prefrontal cortex that manifest in adolescence.
Keywords: prefrontal cortex, adolescence, pruning, stereology
Adolescence, often defined as the period between puberty and adulthood, is a time characterized by neuroanatomical changes that coincide with an increased vulnerability to a variety of clinical disorders including depression, anxiety and schizophrenia (Spear, 2000; Steinberg, 2005; Paus et al., 2008). The prefrontal cortex (PFC), a region primarily involved in executive function and emotional regulation, continues to develop across much of the lifespan with adolescence as a critical period of development. In the human PFC, there is significant synaptic pruning during the adolescent period. Between early adolescence and adulthood, there is a decrease in dendritic spine density on PFC pyramidal neurons (Petanjek et al., 2011) and a decrease in synaptic density (Huttenlocher & Dabholkar, 1997). This period also coincides with an overall decrease in PFC volume (Gogtay et al., 2004; Lenroot & Giedd, 2006). Importantly, there is evidence for differences in the timing and trajectory of PFC development for males and females (Lenroot & Giedd, 2006).
Because of the difficulty in quantification and parcellation of the human PFC and given the homology between the rat and primate frontal cortex (Uylings et al., 2003), rat models for PFC development have been reliably used. Similar to primates, the rodent PFC continues to develop into adulthood, as both its volume (Van Eden & Uylings, 1985a) and laminar specificity (Van Eden & Uylings, 1985b) peak during the periadolescent period and then decrease until at least 90 days of age. Synaptic pruning and dendritic organization have been documented in the adolescent rodent mPFC as well. Specifically, between adolescence and adulthood, both male and female rats lose dendritic spines while only female rats lose a significant number of dendrites (Koss et al., 2014). In addition, while the density of PFC neurons projecting to the amygdala decreases between adolescence and adulthood (Cressman et al., 2010), there is an increase in the density of fibers innervating the mPFC from the amygdala (Cunningham et al., 2002).
Evidence from our laboratory indicates that changes in synaptic density and volume could be related to a loss of total neurons in the region. In the rat mPFC, the number of neurons decreases between adolescence and adulthood with neuronal loss being considerably greater in females (Markham et al., 2007). There is further evidence that in females, the actions of hormones secreted after puberty lead to this decrease in mPFC neuron number, as ovariectomy before puberty prevents this loss when neuron number is assessed in adulthood (Koss et al., 2015). This is similar to findings in the primary visual cortex where ovarian hormones after puberty were found to play a role in neuronal pruning (Nuñez et al., 2002). It is currently unclear whether these neuronal losses in the mPFC involve GABAergic interneurons or glutaminergic pyramidal cells, or a combination of both. However, one study showed that ovariectomized adult females have a greater density of parvalbumin positive GABA neurons in the mPFC than intact females (Cholanian et al., 2014), suggesting the neuronal losses in intact females may detectably include GABA cells.
Counter to the pruning of many cellular components of the gray matter is the increase in white matter between the juvenile and adult period. Myelination is known to continue well into adulthood in the corpus callosum of rats (Nuñez et al., 2000). Electron microscopic analysis of the posterior (splenium) corpus callosum shows that there is an increase in the number of axons that are myelinated between the juvenile period and adulthood, even while axons are pruned (Kim & Juraska, 1997). Furthermore the presence of ovarian hormones from puberty on leads to a lower number of myelinated axons in this region in early adulthood (Yates & Juraska, 2008). Like the posterior corpus callosum, the volume of the white matter under the prefrontal cortex continues to increase between adolescence and adulthood in both male and female rats. However, a sex difference emerges at P90 with males having a larger white matter volume than females (Markham et al., 2007).
A critical question that remains unanswered is the trajectory of these neuroanatomical changes, specifically whether they occur gradually throughout the adolescent period or within a well-defined temporal window that may coincide with puberty. If puberty is a key component for these changes, there should be differences between males and females due to the difference in pubertal timing and the different hormones involved. Here we quantified neuron and glial cell number, along with the volume of the white matter under the prefrontal cortex, in male and female rats. In addition to counts of all neurons in the mPFC, we stereologically counted the number of GABA immunoreactive interneurons. Most previous studies have compared two ages: juvenile/early adolescence and adult. Here we examine five ages from the juvenile period to puberty through adulthood in order to delineate when these anatomical changes occur.
EXPERIMENTAL PROCEDURES
Subjects
Subjects were the offspring of Long-Evans hooded rats obtained from Harlan Laboratories (Indianapolis, IN) and bred in the vivarium in the Psychology Department at the University of Illinois. All animals were weaned on postnatal day (P) 24 and housed with same-sex littermates in pairs or triplets until sacrifice. Tissue from both male and female rats was collected at P25, P35, P45, P60 and P90 (n= 10–11 per group) for a total of 105 animals. P25 is during the preadolescent period, P35 approximates the period of pubertal onset in females, P45 approximates puberty onset in males, and P60 and 90 represent the transition from late adolescence to early adulthood. For each animal that was not sacrificed before pubertal onset, the day the animal reached puberty was recorded. For females, vaginal opening was used as a marker for puberty, since this coincides with surges of both luteinizing hormone and estrogen secretion (Castellano et al., 2011). For males, preputial separation was used for marking puberty, as this coincides with the major surge in androgens (Korenbrot et al., 1977). Each age group was comprised of animals from a minimum of five litters, and at each age, no more than two animals of the same sex came from the same litter. All animals were kept on a 12:12-h light-dark cycle with ad libitum access to food and water. All procedures were approved by the University of Illinois Institutional Care and Use Committee, and adhere to the National Institute of Health guidelines on the ethical use of animals.
Histology
Rats were given a lethal dose of sodium pentobarbital, and were perfused intracardially with 0.1M phosphate buffered saline (PBS) (pH 7.4) followed by 4% paraformaldehyde fixative in PBS. Brains were removed and post-fixed for an additional 24 hours and were then cryoprotected in a PBS solution containing 30% sucrose for three days. Once a brain had sunk in sucrose, it was sliced coronally into 40μm sections with a freezing microtome. Every fifth section containing PFC was temporarily placed into 0.1M PBS and then mounted on gelatin-coated slides. Once dried, sections were stained with Methylene Blue/Azure II. Staining procedures were identical to those described in Markham et al. (2007).
Volume and Cell Number Estimation
Parcellation of the mPFC and adjacent white matter was conducted as previously described by our laboratory (Markham et al., 2007) using the StereoInvestigator software program (MicroBrightField). Briefly, the ventral mPFC (prelimbic (PL) and infralimbic (IL) subregions) was identified based on the cytoarchitectonic criteria delineated in Van Eden & Uylings (1985a, 1985b) (Figure 1A). The boundary of the PL and the anterior cingulate was determined by the broadening of layer V cells and increase in density of layer III cells in the anterior cingulate region, along with a thin “empty band” that is visibly less dense. The ventral boundary of the IL is visible via a loss of laminar organization between cell layers. Within the ventral mPFC layers II/III and V/VI were parcellated for analysis. Layer I was excluded from the analysis due to its lack of neuronal cell bodies.
Figure 1.

(A) Coronal section showing parcellated borders of mPFC including layers 2/3 and 5/6, and adjacent white matter. PL= prelimbic, IL = infralimbic. (B) Photograph within the PL mPFC displaying stereological counting frame with “inclusion” (white) and “exclusion” (black) lines. A characteristic example of a neuron (N) and glia (G) is provided.
Parcellation was conducted by an experimenter blind to the age and sex of the animals. Frontal white matter volume and mPFC volume was calculated from every mounted section between the most anterior mounted section containing the genu of the corpus callosum and the caudal end of the IL mPFC, when the corpus callosum joins both hemispheres at the midline. Using StereoInvestigator, the experimenter defined the area of the frontal white matter, made easily distinguishable by its color and lack of cell bodies, and the area of the ventral mPFC (as described above). Additionally, the post shrinkage section thickness was measured by calculating the difference in focal depth of the top and bottom of visible tissue, made possible by a motorized stage controller (Prior Scientific) which measures z-axis movement in the software. Average section thickness for each animal was estimated from more than 50 sites. Total volume of the frontal white matter and mPFC was calculated by multiplying the average thickness by the area.
Quantification of neurons and glia in the mPFC was performed identically to previous studies from our laboratory (Nuñez et al., 2002; Markham et al., 2007). To obtain cell density, unbiased stereological counts of neurons and glial cells were performed using the StereoInvestigator optical disector. Counting frames (45μm × 45 μm) were randomly distributed across each section by the software. Each frame had ‘inclusion’ and ‘exclusion’ lines that determine which cells to include and exclude in the density calculation (Figure 1B). To prevent overcounting larger cells, a cell was only counted if the bottom of the cell was included in the z-axis within the counting frame (i.e. if it appeared in focus within the counting depth). As the experimenter traversed through the z-axis of a given counting frame (12 μm depth), individual neurons and glial cells, distinguishable by morphological characteristics (Figure 1B), were recorded with the use of distinct markers. Software presets were created to ensure that a minimum of 400 neurons and 150 glial cells were counted within each layer (II/III, V/VI) of the ventral mPFC. To obtain cell density, we divided the number of cells counted by the volume of the counting frame. Then, density was multiplied by the volume of the mPFC to obtain estimates for the total number of neurons and of glia.
GABA Immunocytochemistry and Analysis
Alternate brain sections from animals described above were processed for GABA immunocytochemistry. Based on the analysis of mPFC neuron number, only P25 and P90 male subjects and P35 and P45 female subjects were used. Free-floating sections were rinsed in 0.05M tris-buffered saline (TBS), then blocked in TBS containing 20% normal goat serum (NGS), 1% hydrogen peroxide and 1% bovine serum albumin (BSA) for 30 min. Sections were then incubated for 48 hours in a GABA primary antisera (rabbit polyclonal; Sigma; 1:3000) diluted in TTG: TBS containing 0.3% Triton-X-100 and 2% NGS). Following primary antibody incubation, sections were rinsed 3 times in TTG (5 min each) and placed in a solution containing 5μg/ml goat anti-rabbit biotinylated secondary antibody (Vector Labs) in TTG for 90 min at room temperature, followed by incubation in Avidin-Biotin Complex in TBS (Vectastain Elite Kit, Vector Labs) for 1 hour. Sections were then rinsed three times in TBS and incubated in diaminobenzadine (Sigma Fast Tabs) for 2 minutes. After 5 rinses in TBS, sections were mounted on gelatin-coated slides and coverslipped with Permount.
This procedure labeled GABA immunoreactive (ir) cell bodies in the mPFC (Figure 5A) and stereological counts of GABAir cells were conducted as described above. For each animal GABA cell densities were obtained from two sections containing the mPFC, and StereoInvestogator presets were such that a minimum of 250 cells were counted for every subject. GABA cell densities were then multiplied by mPFC volume to estimate the total number of GABA positive cells in the region. The ratio of GABA cells to total mPFC neurons was also calculated.
Figure 5.
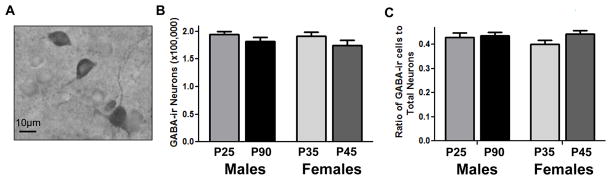
Total number of GABA positive cells in the male and female mPFC at select ages. The number of GABA immunoreactive cells (A) did not significantly change in female rats between P35 and P45 or in males between P25 and P90 (B). There was a non-significant trend (p<.076) for an increase in the ratio of GABA neurons to total mPFC neurons in females between P35 and P45 (C).
Statistical Analysis
Based on previous work from our laboratory, we hypothesized a different trajectory of neuroanatomical changes in males and females. As a result, males and females were analyzed separately in these experiments. For neuron number, glial cell number and the volume of the white matter a one way ANOVA (age) was run for males and females separately, with litter as a cofactor. For each of these measures, four a priori comparisons were examined for post hoc analysis using the Fisher’s least significant difference (LSD) test: P25 vs. P35, P35 vs. P45, P45 vs. P60 and P60 vs. P90. For analysis of GABA cell counts, separate t-tests were conducted for males (P25 vs. P90) and females (P35 vs. P45).
RESULTS
Age of Puberty
For animals that were not sacrificed prior to pubertal onset, males reached puberty at an average age of 44.9 days with a range of 42–48. The average age for the onset of puberty in females was 34.9 with a range of 32–38.
Neuron Number
Because our analysis revealed no differences in neuron counts between cerebral hemispheres, or between cortical layers (2/3, 5/6) in males or females at any age, data presented are pooled from left and right hemispheres from layers 2–6. Females lost a significant number of neurons after P35 (Figure 2A). A one-way ANOVA revealed a significant decrease in the number of neurons with age (F1,50=2.987, p=0.029). Post hoc analysis revealed that for females, there was a significant difference in neuron number between P35 and P45 (p=0.043). There were no differences between P25 and P35 (p=0.971), P45 and P60 (p=0.829) or P60 and P90 (p=0.798). Based on the timing of neuronal losses in females, we additionally ran a separate t-test for all female subjects at P35 with the independent variable being whether or not they reached puberty by that day. For females within the P35 group, there was no significant difference between those who had reached puberty and those who had not (p=0.13) (Figure 2B) although the direction of the means may suggest that the onset of puberty was a precipitating factor in the drop. In males, although mean neuron number appeared to decrease incrementally, there was only a weak trend for age (F4,53=3.143, p=0.083) (Figure 3).
Figure 2.
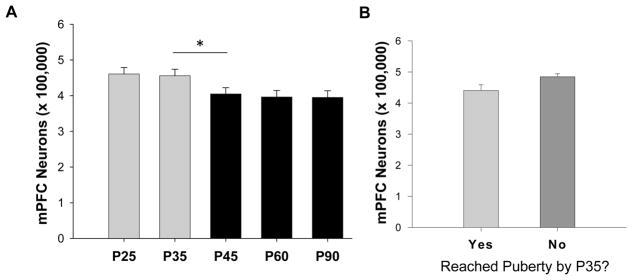
Total number of neurons in the female mPFC across adolescence. (A) Females lost a significant number of neurons between P35 and P45, coinciding with the onset of puberty (*p<0.05). There were no changes in neuron number after P45. (B) The P35 female group was divided into those who had reached puberty by P35 (n=6) and those who had not (n= 4). There was no statistical differences between these two subgroups, yet the direction of the means supports pubertal onset as a potential trigger for neuronal lose in females.
Figure 3.

Total number of neurons in the male mPFC across adolescence. There were no significant changes in neuron number in males across adolescence. There was a modest trend (p=0.083) for a decrease across the ages
Glia Number
Again, no differences were seen between hemispheres or cortical layers, so data was averaged for these factors for each animal. There was no effect of age in the total number of glial cells in either males (F4,53=0.501, p=0.735) or females (F4,50=1.101, p=0.368) (Figure 4).
Figure 4.
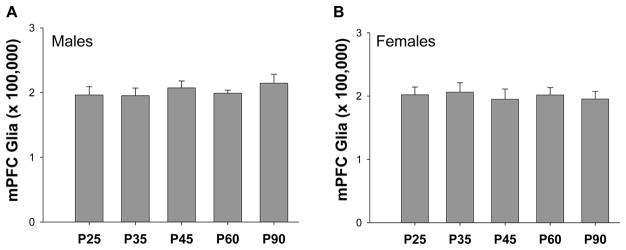
Total number of glial cells in the male and female mPFC across adolescence. There were no differences in glial cell number between any ages examined in male (A) or in females (B) rats.
GABA Neuron Number
In females, there was not a significant difference in the number of GABAir neurons between P35 and P45 (t1,19=1.411, p=0.176), nor was there any difference in males between P25 and P90 (t1,19=1.257, p=0.225) (Figure 5B). There was a non-significant trend towards an increase in the ratio of GABA neurons to total neurons in females between P35 and P45 (t1,19=1.880, p=0.076), while this trend was not found in males between P25 and P90 (t1,19=0.297, p=0.770) (Figure 5C).
Volume of White Matter under Frontal Cortex
The ANOVA for males showed a significant increase due to age (F4,53=64.729, p=<0.001) (Figure 6A). Post hoc analysis revealed that every age was significantly different from the next: P25 was different from P35 (p=0.001), P35 from P45 (p<0.001), P45 from P60 (p=0.005) and P90 from P60 (p=0.032). There was also a significant increase in females (F4,50=21.568, p=<0.001) (Figure 6B). Post hoc tests revealed a significant difference between P25 and P35 (p=0.001) and between P45 and P60 (p<0.001). Unlike in males, there was no significant difference between P35 and P45 (p=0.145), or between P60 and P90 (p=0.220).
Figure 6.

Changes in the volume of the white matter under the frontal cortex in males and females across adolescence. (A) In males, white matter under the frontal cortex increased progressively with age. Each age group was statistically significant from the other (p<0.05). (B) There was a significant increase in frontal white matter in females across adolescence. There was an increase from P25 to P35, and from P45 to P60 (p<0.05), but no differences between P35 and P45, or P60 and P90.
DISCUSSION
In the present study, there was a decrease in the number of neurons in the female mPFC between P35 and P45, which coincides with the onset of puberty. Furthermore, it appears that these neuronal losses were not predominantly GABAergic interneurons. Male rats showed only a weak trend for neuronal loss across ages. Additionally, while both males and females gained white matter volume under the prefrontal cortex, this increase was somewhat blunted in female rats after puberty. There was no detectable change in total glial cell number in males or females across the periadolescent period, though this could be related to the continuing proliferation of glial cells after early development through adolescence and beyond (Lee et al., 2000).
It has been shown that females lose neurons from the prelimbic and infralimbic mPFC between adolescence (P35) and adulthood (P90) (Markham et al., 2007). Here, we examine the trajectory of the neuronal losses took place between P35 and P45. Ovariectomy before puberty at P20 prevented this neuronal loss in females compared to intact females by adulthood (Koss et al., 2015), suggesting that ovarian hormones may play a role in the pruning of neurons. In the present study, neuronal losses coincided with pubertal onset provides more support for a role of ovarian hormones at puberty in the development of the PFC in adolescent females. This is similar to the visual cortex where the presence of ovarian hormones between puberty and adolescence has been shown to reduce neuron number (Nuñez et al., 2002). Estrogen actions have been shown to induce apoptosis during early development (Arai et al., 1996; Forger, 2009) so it is conceivable that they could do so during adolescence as well. There is currently a paucity of information regarding the complete ontogeny of estrogen receptor expression in the mPFC during adolescence, but there is evidence that in rats, ERα is present in the female mPFC at least until P25 (Wilson et al., 2011), and that ERβ is strongly expressed in the mPFC of young adult females (Shughrue et al., 1997). Future studies should examine the ontogeny of estrogen receptor localization during the pubertal period.
What currently remains unknown is whether these changes in neuron number can only occur during adolescence (i.e., a critical/sensitive period) or whether neurons in the mPFC can be affected by ovarian hormones after adolescence. There probably are limits to the effects witnessed in adolescence, given that ovariectomy and hormone replacement has no effect on mPFC number in middle aged (12–13 months) female rats (Chisholm et al., 2012). The results of the present study suggest that, at least in terms of neuron number, there is a brief period of rapid change coinciding withpuberty in females. Further evidence that the female mPFC is highly plastic during the pubertal period comes from Nemati and Kolb (2012) where female rats with induced damage to the PFC during early adolescence (P35), compared to late adolescence (P55), displayed greater dendritic reorganization and more prominent behavioral resilience in adulthood. Additionally, MRI analysis of human peripubertal adolescents indicates that the increase of estrogen at puberty is correlated with decreases in gray matter in females while no changes were found in the peripubescent males (Peper et al., 2009).
The present study also examined whether GABAergic interneurons were detectably lost. Previous work has suggested that there is an increase in the density of GABAergic interneurons in the female mPFC following ovariectomy (Cholanian et al., 2014), and a decrease in GABA cell density in males between adolescence and adulthood (Leussis et al., 2012). However, both of these studies only examined cell density and were restricted to parvalbumin labeled cells, a distinct subtype of GABA neurons. Stereological counts of all GABA positive cells in in the current study suggests that the total number of interneurons in males do not detectably change between preadolescence and adulthood, and that the neurons lost in the adolescent female mPFC were not predominantly GABAergic. This is further indicated by the nonsignificant trend for the ratio of GABAergic to the total number of neurons to increase between P35 and 45 when the total number of neurons decreases in females indicating that proportionately more non-GABAergic neurons are pruned than GABAergic. Given the extensive diversity of cortical pyramidal neurons, immunocytochemical analysis of glutamatergic pyramidal cells remains impractical (reviewed in Lodato et al., 2014). Thus, we cannot currently exclude the possibility that a small number of GABAergic cells were pruned.
White matter under the frontal cortex increased in both males and females throughout the periadolescent period. In males, this increase was progressive and continued at every age examined from P25 through P90. In females, there was no increase in white matter volume between P35 and P45, corresponding with puberty. Ovariectomy prior to puberty resulted in increased volume of white matter under the mPFC in adult females (Koss et al., 2014), extending the finding of Yates and Juraska (2008) that pubertal ovarian hormones slow the progression of myelination during adolescence in the posterior corpus callosum. Furthermore, in humans, while testosterone is positively correlated with white matter development, estradiol levels are negatively correlated with the size of frontal white matter in adolescent girls (Herting et al., 2012). We hypothesize that increases in myelination are the underlying cellular basis for the increases in the volume of the white matter in both sexes, and the hormones of puberty slow this process in females. All of this speculation awaits further studies of the cellular changes occurring in the anterior white matter at this time.
In general, the present results support the hypothesis that in females, pubertal onset coincides with a critical developmental period for the mPFC. Given this hypothesis, it is possible that puberty could play a role in improved performance on cognitive tasks in females, particularly those that depend on the mPFC. The transition from adolescence to early adulthood coincides with increased capacity for a variety of cognitive tasks, particularly those that involve cognitive flexibility (Koss et al., 2011; Andrzejewski et al., 2011; Naneix et al., 2012). Though there is currently no direct link between neuronal pruning in the mPFC and improved cognition, it is possible that this neuroanatomical change, in conjunction with other changes occurring during the pubertal period, could play an important role in increased capacity for executive function. Future studies should examine the possible effects of pubertal timing on the development of cognitive behavior.
A potential contribution to the occurrence of neuronal loss in females, but not males, during the peripubertal period might involve the environmental stimulation of social behavior. There is some precedent for this, in that neuronal survival is influenced by maternal stimulation during early postnatal development (Moore et al., 1992). In adolescence, males tend to exhibit greater levels of play behavior, an effect that is at least partially mediated by perinatal hormonal influences (reviewed in Auger & Oleson, 2009). There is evidence that play behavior during the juvenile and adolescent period affects neural reward circuits, and involves the neocortex (reviewed in Vanderschuren et al., 1997). It is possible that this type of behavior is a form of environmental enrichment, perhaps preventing neuronal loss in males. The role of the environment in protecting against or promoting neuronal loss during adolescence should be explored.
The present study indicates that male puberty is not associated with changes in mPFC neuron number. This does not necessarily mean that other facets of development are not occurring in males at the same time. For example, in males, the density of both D1 and D2 dopamine receptors increases between preadolescence and early adulthood (Andersen et al., 2000). Additionally, TH fibers increase between adolescence and adulthood (Naneix et al., 2012). Along with the trajectory of dopaminergic innervation in the adolescent mPFC, other synaptic changes that may be occurring during this time should be explored. Research on the timing of changes in the adolescent mPFC may implicate potential mechanisms that lead to vulnerability and dysfunction of the mPFC during adolescence that vary in timing and degree in males and females.
Highlights.
Female rats lose neurons in the medial prefrontal cortex between postnatal days 35 and 45.
Males do not lose a significant number of neurons from preadolescence to adulthood.
There were no changes in glial cell number in males or females from preadolescence to adulthood.
Males and females gained white matter under the frontal cortex during this time.
Acknowledgments
We thank Taehyeon Kim for her assistance and the Beckman Institute Imaging Technology Group for use of the microscopy suite. This work was supported by NIH MH099625 to JM Juraska.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Andersen SL, Thompson AT, Rutstein M, Hostetter JC, Teicher MH. Dopamine receptor pruning in prefrontal cortex during the periadolescent period in rats. Synapse. 2000;37:167–169. doi: 10.1002/1098-2396(200008)37:2<167::AID-SYN11>3.0.CO;2-B. [DOI] [PubMed] [Google Scholar]
- Andrzejewski ME, Schochet TL, Feit EC, Harris R, McKee BL, Kelley AE. A comparison of adult and adolescent rat behavior in operant learning, extinction and behavioral inhibition paradigms. Behavioral Neuroscience. 2011;125 (1):93–105. doi: 10.1037/a0022038. [DOI] [PubMed] [Google Scholar]
- Arai Y, Sekine Y, Murakami S. Estrogen and apoptosis in the developing sexually dimorphic preoptic area in female rats. Neuroscience Research. 1996;25 (4):403–407. doi: 10.1016/0168-0102(96)01070-x. [DOI] [PubMed] [Google Scholar]
- Auger AP, Olesen KM. Brain sex differences and the organisation of juvenile social play behavior. Journal of Neuroendocrinology. 2009;21:519–525. doi: 10.1111/j.1365-2826.2009.01871.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Castellano JM, Bentsen AH, Sanchez-Garrido MA, Ruiz-Pino F, Romero M, Garcia-Galiano D, Aguilar E, Pinilla L, Dieguez C, Mikkelsen JD, Tena-Sempere M. Early metabolic programming of puberty onset: Impact of changes in postnatal feeding and rearing conditions on the timing of puberty and the development of the hypothalamic kisspeptin system. Endocrinology. 2011;152 (9):3396–3408. doi: 10.1210/en.2010-1415. [DOI] [PubMed] [Google Scholar]
- Chisholm NC, Packard AR, Koss WA, Juraska JM. The effects of long-term treatment with estradiol and medroxyprogesterone acetate on tyrosine hydroxylase fibers and neuron number in the medial prefrontal cortex of aged female rats. Endocrinology. 2012;153 (10):4874–4882. doi: 10.1210/en.2012-1412. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cholanian M, Lobzova A, Das Bhargab, Yelleswarapu C, Donaldson ST. Digital holographic microscopy discriminates sex differences in medial prefrontal cortex GABA neurons following amphetamine sensitization. Pharmacology, Biochemistry and Behavior. 2014;124:326–332. doi: 10.1016/j.pbb.2014.06.026. [DOI] [PubMed] [Google Scholar]
- Cressman VL, Balaban J, Steinfeld S, Shemyakin A, Graham P, Parisot N, Moore H. Prefrontal cortical inputs to the basal amygdala undergo pruning during late adolescence in the rat. Journal of Comparative Neurology. 2010;518 (14):2693–2709. doi: 10.1002/cne.22359. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cunningham MG, Bhattacharyya S, Benes FM. Amygdalo-cortical sprouting continues into early adulthood: implications for the development of normal and abnormal function during adolescence. Journal of Comparative Neurology. 2002;453 (2):116–130. doi: 10.1002/cne.10376. [DOI] [PubMed] [Google Scholar]
- Forger NG. Control of cell number in the sexually dimorphic brain and spinal cord. Journal of Neuroendocrinology. 2009;21 (4):393–399. doi: 10.1111/j.1365-2826.2009.01825.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gogtay N, Giedd JN, Lusk L, Hayashi KM, Greenstein D, Vaituzis AC, Nugent TF, 3rd, Herman DH, Clasen LS, Toga AW, Rapoport JL, Thompson PM. Dynamic mapping of human cortical development during childhood through early adulthood. Proc Natl Acad Sci. 2004;101(21):8174–8179. doi: 10.1073/pnas.0402680101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Herting MM, Maxwell EC, Irvine C, Nagel BJ. The impact of sex, puberty, and hormones on white matter microstructure in adolescents. Cerebral Cortex. 2012;22 (9):1979–1992. doi: 10.1093/cercor/bhr246. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huttenlocher PR, Dabholkar AS. Regional differences in synaptogenesis in human cerebral cortex. Journal of Comparative Neurology. 1997;387:167–178. doi: 10.1002/(sici)1096-9861(19971020)387:2<167::aid-cne1>3.0.co;2-z. [DOI] [PubMed] [Google Scholar]
- Kim JHY, Juraska JM. Sex differences in the development of axon number in the splenium of the rat corpus callosum from postnatal day 15 through 60. Developmental Brain Research. 1997;102:77–85. doi: 10.1016/s0165-3806(97)00080-1. [DOI] [PubMed] [Google Scholar]
- Korenbrot CC, Huhtaniemi IT, Weiner RI. Preputial separation as an external sign of pubertal development in the male rat. Biology of Reproduction. 1977;17 (2):298–303. doi: 10.1095/biolreprod17.2.298. [DOI] [PubMed] [Google Scholar]
- Koss WA, Franklin AD, Juraska JM. Delayed alternation in adolescent and adult male and female rats. Developmental Psychobiology. 2011;53 (7):724–731. doi: 10.1002/dev.20543. [DOI] [PubMed] [Google Scholar]
- Koss WA, Belden CE, Hristov AD, Juraska JM. Dendritic remodeling in the adolescent medial prefrontal cortex and the basolateral amygdala of male and female rats. Synapse. 2014;68 (2):61–72. doi: 10.1002/syn.21716. [DOI] [PubMed] [Google Scholar]
- Koss WA, Lloyd MM, Sadowski RN, Wise LM, Juraska JM. Gonadectomy before puberty decreases the number of neurons and glia in the medial prefrontal cortex of female, but not male, rats. Developmental Psychobiology. 2015;57(3):305–12. doi: 10.1002/dev.21290. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee JC, Mayer-Proschel M, Rao MS. Gliogenesis in the central nervous system. Glia. 2000;30 (2):105–121. doi: 10.1002/(sici)1098-1136(200004)30:2<105::aid-glia1>3.0.co;2-h. [DOI] [PubMed] [Google Scholar]
- Lenroot RK, Giedd JN. Brain development in children and adolescents: Insights from anatomical magnetic resonance imaging. Neurosci Biobehav Rev. 2006;30 (6):718–729. doi: 10.1016/j.neubiorev.2006.06.001. [DOI] [PubMed] [Google Scholar]
- Lodato S, Shetty AS, Arlotta P. Cerebral cortex assembly: generating and reprogramming projection neuron diversity. Trends in Neurosciences. 2014;38 (2):117–125. doi: 10.1016/j.tins.2014.11.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leussis MP, Freund N, Brenhouse HC, Thompson BS, Andersen SL. Depressive-like behavior in adolescents after maternal separation: Sex differences, controllability and GABA. Developmental Neuroscience. 2012;34:210–217. doi: 10.1159/000339162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Markham JA, Morris JR, Juraska JM. Neuron number decreases in the rat ventral, but not dorsal, medial prefrontal cortex between adolescence and adulthood. Neuroscience. 2007;144:961–968. doi: 10.1016/j.neuroscience.2006.10.015. [DOI] [PubMed] [Google Scholar]
- Moore CL, Dou H, Juraska JM. Maternal stimulation affects the number of motor neurons in a sexually dimorphic nucleus of the lumbar spinal cord. Brain Research. 1992;572:52–56. doi: 10.1016/0006-8993(92)90449-j. [DOI] [PubMed] [Google Scholar]
- Naneix F, Marchland AR, Di Scala G, Pape JR, Coutureau E. Parallel maturation of goal-directed behavior and dopaminergic systems during adolescence. Journal of Neuroscience. 2012;32 (46):16223–16232. doi: 10.1523/JNEUROSCI.3080-12.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nemati F, Kolb B. Recovery from medial prefrontal cortex injury during adolescence: implications for age-dependent plasticity. Behavioural Brain Research. 2012;229:168–175. doi: 10.1016/j.bbr.2012.01.002. [DOI] [PubMed] [Google Scholar]
- Nuñez JL, Nelson J, Pych JC, Kim JHY, Juraska JM. Myelination in the splenium of the corpus callosum in adult male and female rats. Developmental Brain Research. 2000;120:87–90. doi: 10.1016/s0165-3806(99)00193-5. [DOI] [PubMed] [Google Scholar]
- Nuñez JL, Sodhi J, Juraska JM. Ovarian hormones after postnatal day 20 reduce neuron number in the rat primary visual cortex. Journal of Neurobiology. 2002;52 (4):312–321. doi: 10.1002/neu.10092. [DOI] [PubMed] [Google Scholar]
- Paus T, Keshavan M, Giedd JN. Why do many psychiatric disorders emerge during adolescence? Nature Reviews: Neuroscience. 2008;9:947–957. doi: 10.1038/nrn2513. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peper JS, Brouwer RM, Schnack HG, van Baal GC, van Leeuwen M, van den Berg SM, Delemarre-Van de Waal HA, Boomsma DI, Kahn RS, Hulshoff Pol HE. Sex steroids and brain structure in pubertal boys and girls. Psychoneuroendocrinology. 2009;34 (3):332–342. doi: 10.1016/j.psyneuen.2008.09.012. [DOI] [PubMed] [Google Scholar]
- Petanjek Z, Judas M, Simic G, Rasin MR, Uylings HB, Rakic P, Kostovic I. Extraordinary neoteny of synaptic spines in the human prefrontal cortex. Proc Natl Acad Sci. 2011;108 (32):13281–13286. doi: 10.1073/pnas.1105108108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shughrue PJ, Lane MV, Merchenthaler I. Comparative distribution of estrogen receptor-α and –β mRNA in the rat central nervous system. Journal of Comparative Neurology. 1997;388:507–525. doi: 10.1002/(sici)1096-9861(19971201)388:4<507::aid-cne1>3.0.co;2-6. [DOI] [PubMed] [Google Scholar]
- Spear LP. The adolescent brain and age-related behavioral manifestations. Neurosci Biobehav Rev. 2000;24:417–463. doi: 10.1016/s0149-7634(00)00014-2. [DOI] [PubMed] [Google Scholar]
- Steinberg L. Cognitive and affective development in adolescence. Trends Cogn Sci. 2005;9:69–74. doi: 10.1016/j.tics.2004.12.005. [DOI] [PubMed] [Google Scholar]
- Uylings HBM, Groenewegen HJ, Kolb B. Do rats have a prefrontal cortex? Behavioral Brain Research. 2003;146:3–17. doi: 10.1016/j.bbr.2003.09.028. [DOI] [PubMed] [Google Scholar]
- Vanderschuren LJMJ, Niesinki RJM, Van Ree JM. The neurobiology of social play behavior in rats. Neuroscience and Biobehavioral Reviews. 1997;21 (3):309–326. doi: 10.1016/s0149-7634(96)00020-6. [DOI] [PubMed] [Google Scholar]
- Van Eden CG, Uylings HB. Postnatal volumetric development of the prefrontal cortex in the rat. Journal of Comparative Neurology. 1985a;241 (3):268–274. doi: 10.1002/cne.902410303. [DOI] [PubMed] [Google Scholar]
- Van Eden CG, Uylings HB. Cytoarchitectonic development of the prefrontal cortex in the rat. Journal of Comparative Neurology. 1985b;241 (3):253–267. doi: 10.1002/cne.902410302. [DOI] [PubMed] [Google Scholar]
- Wilson ME, Westberry JM, Trout Al. Estrogen receptor-alpha gene expression in the cortex: sex differences during development and in adulthood. Hormones and Behavior. 2011;59:353–357. doi: 10.1016/j.yhbeh.2010.08.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yates MA, Juraska JM. Pubertal ovarian hormone exposure reduces the number of myelinated axons in the splenium of the rat corpus callosum. Experimental Neurology. 2008;209:284–287. doi: 10.1016/j.expneurol.2007.09.013. [DOI] [PMC free article] [PubMed] [Google Scholar]