

Abstract
While there are currently many well-established topical hemostatic agents for field administration, there are still limited tools to staunch bleeding at less accessible injury sites. Current clinical methods of restoring hemostasis after large volume blood loss include platelet and clotting factor transfusion, which have respective drawbacks of short shelf-life and risk of viral transmission. Therefore, synthetic hemostatic agents that can be delivered intravenously and encourage stable clot formation after localizing to sites of vascular injury are particularly appealing. In the past three decades, platelet substitutes have been prepared using drug delivery vehicles such as liposomes and PLGA nanoparticles that have been modified to mimic platelet properties. Additionally, structural considerations such as particle size, shape, and flexibility have been addressed in a number of reports. Since platelets are the first responders after vascular injury, platelet substitutes represent an important class of intravenous hemostats under development. More recently, materials affecting fibrin formation have been introduced to induce faster or more stable blood clot formation through fibrin crosslinking. Fibrin represents a major structural component in the final blood clot, and a fibrin-based hemostatic mechanism acting downstream of initial platelet plug formation may be a safer alternative to platelets to avoid undesired thrombotic activity. This review explores intravenous hemostats under development and strategies to optimize their clotting activity.
INTRODUCTION
A blood clot is a biopolymer-colloid composite that prevents bleeding from damaged vasculature. The colloid component consists of activated platelets bound to subendothelial proteins that are exposed after vascular injury (i.e. collagen, von Willebrand factor) and aggregated to form a platelet plug at the site of injury (Figure 1). The biopolymer component is subsequently formed by the coagulation cascade. Locally-activated thrombin enzyme cleaves circulating fibrinogen to form fibrin monomers. Fibrin monomers then self-polymerize into fibers to form a viscoelastic biopolymer network interspersed through the platelet plug. For small injuries, this composite is sufficient for maintaining hemostasis, the prevention of blood loss from damaged blood vessels. However, for more severe bleeding from traumatic injury, surgery, or bleeding disorders, hemostatic agents that augment the natural clotting process or physically seal the wound itself are needed to staunch bleeding. Hemostats for field administration after traumatic injury are particularly important as hemorrhage is responsible for 33–56% of prehospital deaths1 and 90% of preventable military battlefield casualties2, and immediate intervention is believed to be key in reducing early hemorrhage-related mortality and morbidity.
Figure 1.

Schematic of the clotting process showing initial platelet plug formation during primary hemostasis (A) followed by fibrin formation during secondary hemostasis (B). During primary hemostasis, circulating platelets adhere to the subendothelial matrix through binding of the platelet receptors GPIbα and GPIa-IIa/GPVI to von Willebrand Factor (vWF) and collagen, respectively. Upon platelet activation, GPIIb-IIIa goes through a conformational change which enables it to bind fibrinogen. Platelet aggregation is induced by multiple platelets binding the same fibrinogen molecule. During secondary hemostasis, activated thrombin enzyme cleaves the N-termini of Aα (red) and Bβ (green) polypeptide chains, revealing knob A and knob B peptide domains, respectively. Knobs A and B interact with holes a and b in the D nodules of other fibrin monomers to form half-staggered, double-stranded protofibrils. Non-specific interaction between αC domains causes lateral aggregation of protofibrils to form fibrin fibers.
There are many well-established clinically-used topical hemostatic agents. These include gel sealants derived from a mixture of biological materials (i.e. human fibrinogen and thrombin, bovine collagen, and human platelets) and absorbent hemostatic dressings manufactured with fibrin, chitin/chitosan, or mineral zeolites used to increase concentration of clotting factors, platelets, and erythrocytes at the site of injury.3 Topical hemostats are limited to treating visible and accessible injuries. Transfusion of blood products (i.e. fresh frozen plasma, platelets) and recombinant clotting factors is used to restore clotting function in those who are coagulopathic after large volume blood loss and are the only current intravenous hemostats in use.4 However, biological agents have many limitations such as risk of immunogenicity and viral transmission, restrictive storage conditions, short shelf-life, and involved manufacturing processes. In contrast, synthetic polymers and polymeric nanoparticles have tunable physical and chemical properties and, in general, have more straightforward manufacturing processes and longer shelf-lives compared to biological products. Therefore, current efforts are underway to develop synthetic hemostats that can be administered systemically for quick resolution of bleeding at inaccessible injury sites.
Blood clot formation is heavily mediated by specific protein-protein interactions. These interactions are responsible for platelet adhesion at injury sites, platelet aggregation, and fibrin polymerization. Strategies for inducing hemostasis have therefore predominantly focused on leveraging these interactions to accelerate and stabilize the assembly of non-covalent bonds between clot components. To do so, artificial platelets and fibrin-modulating polymers and particles have been designed with multivalent display of binding motifs to promote clot aggregation at the injured blood vessel. Furthermore, in the case of synthetic platelet substitutes, the shape and flexibility of particles on which these binding motifs are displayed have proven important in optimizing particle rolling velocities and contact area with the tissue surface, two contributing factors for platelet adhesion.5 The design of intravenous hemostats with site-specific activity has relied upon knowledge and strategies developed by the systemic drug delivery field. This review discusses the materials and engineering design considerations used in synthetic intravenous hemostats under pre-clinical development.
ARTIFICIAL PLATELETS
Primary hemostasis
Platelets are anucleate cell fragments 1–3 μm in diameter formed from the cytoplasm of megakaryocytes.6,7 During primary hemostasis, a platelet plug is formed at the injury site (Figure 1A). As the first responders after injury, platelets instigate clot formation by first marginating to the damaged endothelium, binding to proteins in the exposed subendothelial matrix, and finally aggregating after activation. The biconvex discoid shape of platelets facilitates margination towards vessel walls under blood flow.8 Upon reaching an injured wall, platelets are captured by shear-induced binding of the platelet surface glycoprotein GPIbα to von Willebrand factor (vWF) immobilized on the exposed subendothelial matrix. Reversible tethering by vWF allows platelets to spread and roll along the tissue surface for subsequent shear-independent binding of GPIa-IIa and GPVI to collagen, which stabilizes platelet adhesion.5,9,10 Platelets are activated by collagen-binding and agonists such as adenosine diphosphate (ADP) and thrombin. During activation, platelets take on a stellate shape and the surface integrin GPIIb-IIIa undergoes a conformational change which enables it to bind to any of three peptide domains in fibrinogen (i.e. RGD motifs: RGDF, RGDS; H12 sequence: HHLGGAKQAGDV).11–13 In this “sticky” state, platelets are thus able to aggregate by binding the same fibrinogen molecule, which has a total of 6 possible platelet-binding domains due to its symmetrical structure. Additional ADP is secreted by activated platelets for further local platelet activation. The following sections discuss the design and optimization of platelet substitutes. For further reading solely on synthetic platelets, an excellent prior review is available from the Sen Gupta lab.14
Fibrinogen and RGD peptide-coated microparticles
There are approximately 80,000 GPIIb-IIIa receptors per platelet15, and up to 40,000 fibrinogen can reportedly bind to activated platelets16–18. The first platelet substitutes were designed to promote platelet aggregation through surface display of fibrinogen or RGD-containing peptide (Table 1).19–22 In their constructs, Agam et al. covalently coupled fibrinogen to the surface of human platelets19 and erythrocytes20 by fixation with formaldehyde and showed enhanced platelet aggregation when incubated with activated platelets in vitro as well as restoration of normal bleeding times (i.e. time to clot) when delivered to thrombocytopenic rats (i.e. rats with low platelet counts). Untreated thrombocytopenic rats had bleeding times of 18 ± 1.5 min, whereas thrombocytopenic rats injected with fibrinogen-bearing erythrocytes had bleeding times of 4.5 ± 1.0 min, closer to the 2.5 ± 0.1 min bleeding time observed in non-thrombocytopenic rats.20 In later work, Levi et al. proposed the use of Synthocytes™, 3.5–4.5 μm-diameter human albumin microcapsules adsorbed with fibrinogen, for treatment of severe thrombocytopenia.22,23 In contrast to control rabbits with normal ear bleeding times of 1.7 ± 0.4 min, thrombocytopenic rabbit models had prolonged bleeding times of 21.7 ± 4.4 min. Intravenous injection of Synthocytes™ resulted in corrected bleeding times of 5.2 ± 1.7 min. While fibrinogen-functionalized materials bridge activated platelets, aggregate formation is also likely due to thrombin-induced fibrin formation. Fibrin polymerization between surfaces of modified platelets and erythrocytes would entrap platelets resulting in aggregate formation.
Table 1.
Summary of intravenous hemostats and their mechanism of action.
| Mechanism of Action | Class of particle or polymer | Targeting ligand(s) | References |
|---|---|---|---|
| Platelet substitutes | |||
|
| |||
| Binds to subendothelial matrix | Liposome | rGPIα-IIα | Nishiya et al. (2001)31 |
| rGPIbα | Takeoka et al. (2002)5 | ||
| vWF binding peptide (VBP): TRYLRIHPQSQVHQI Collagen binding peptide (CBP): [GPO]7* |
Ravikumar et al. (2012)37 | ||
| Binds to platelets | Platelet | Fixation of fibrinogen on surface using formaldehyde | Agam et al. (1983)19 |
| Erythrocyte | Fixation of fibrinogen on surface using formaldehyde | Agam et al. (1992)20 | |
| Conjugation of Ac-CGGRGDF-NH2 via a heterobifunctional linker (thromboerythrocyte) | Coller et al. (1992)21 | ||
| Albumin microparticles | Surface adsorption of fibrinogen (Synthocytes™) | Levi et al. (1999)22 | |
| rGPIα-IIα | Teramura et al. (2003)93 | ||
| Surface display of H12 peptide HHLGGAKQAGDV | Okamura et al. (2005)33–35 | ||
| Liposome | Surface display of H12 peptide HHLGGAKQAGDV | Okamura et al. (2005)32 | |
| GSSSGRGDSPA and P-selectin binding peptide DAEWVDVS conjugated to liposomal membrane lipids | Modery et al. (2011)36 | ||
| PLL-PLGA nanoparticles | PEG arms terminated with GRGDS conjugated to NP surface | Bertram et al. (2009)41 | |
| Binds to subendothelial matrix and platelets | Liposome | Extraction and incorporation of platelet membrane proteins including GPIbα, GPIIb-IIIα, and GPIV/III into the liposomal membrane (plateletsome) | Rybak et al. (1993)29 |
| Surface display of VBP, CBP, and fibrinogen mimetic peptides (i.e. cyclo-CNPRGDY(OEt)RC or GRGDS) | Ravikumar et al. (2012)38 Modery-Pawlowski et al. (2013)39 Anselmo et al. (2014)45 |
||
| Binds to and activates platelets | Lipsomes | H12 peptide-coated liposomes encapsulating ADP | Okamura et al. (2009)40 |
|
| |||
| Fibrin modulators | |||
|
| |||
| Crosslinks fibrin | Ultra-low crosslinked p(NIPAm)-AAc microgels | Fibrin-binding sdFvs conjugated to the surface of ~1 μm-diameter microgel particles (platelet-like particles) | Brown et al. (2014)91 |
| Poly(HEMA) | Fibrin-binding peptides (Ac- Y(DGl)C(HPr)YGLCYIQGK-Am*†) grafted onto a linear poly(HEMA) backbone (polySTAT) | Chan et al. (2015)88 | |
O and HPr = hydroxyproline; DGl = D-glutamic acid; Ac = acetylation; Am = amidation.
Cyclized via C3 and C8 residues.
To avoid risk of infectious contaminants in purified fibrinogen, Coller et al. opted to functionalize erythrocytes with fibrinogen-mimetic peptides containing an RGD sequence (Ac-CGGRGDF-NH2) synthesized using solid phase peptide synthesis.21 This product, called thromboerythrocytes, represents one of the earliest uses of peptides in platelet engineering. Since RGD peptides are three orders of magnitude smaller than fibrinogen molecules, a significantly greater number of ligands were available for binding to GPIIb-IIIa (~0.5–1.5 x 106 peptides per erythrocyte) compared to fibrinogen-modified erythrocytes (~58 fibrinogen per erythrocyte)20. Interaction between surface-bound peptides and GPIIb-IIIa was confirmed after inhibitors of fibrinogen-platelet binding (i.e. free RGD peptide, monoclonal antibody against GPIIb-IIIa) successfully knocked down platelet aggregation.
While these hemostatic agents were effective at inducing platelet aggregation, translation of these materials into clinical use is limited by possible immunogenic responses against the biologically-derived components as well as difficulties in scale-up. Therefore, more recent iterations of platelet substitutes have focused on surface modification of synthetic particle platforms such as liposomes and polymer-based nanoparticles.
Liposome-based platelet substitutes
Since the beginning of lipid vesicle research in the 1960s, liposomes have remained a key technology in the drug delivery field due to their ability to carry both hydrophobic and hydrophilic cargo, straightforward surface functionalization through lipid modification, and tunable residence time in circulation.24 Platelet-like liposomes were initially synthesized to study the function of glycoproteins in the platelet membrane and were done so by reconstituting isolated platelet membrane proteins in liposomal membranes through reverse-phase sonication or evaporation and subsequent extrusion.25–27 Plateletsomes, liposomes containing at least 15 different platelet membrane proteins including GPIb, GPIIb-IIIa, and GPVI/III, were first evaluated in thrombocytopenic rats and shown to reduce tail bleeding times by 42% after intravenous injection.28,29 Similar liposome-based platelet substitutes were also synthesized using recombinant glycoproteins, rGPIbα and rGPIIα-IIIα.30,31 Following this work, platelet substitutes were functionalized with peptide ligands rather than whole proteins due to increased knowledge of peptide binding domains, the relative ease of peptide synthesis, the monodispersity of peptide products, and the controlled manner in which they can be conjugated to surfaces. Common peptide ligands employed include vWF-binding peptide (VBP: TRYLRIHPQSQVHQI) and collagen-binding peptide (CBP: [GPO]7) to mimic platelet adhesion and fibrinogen-mimetic peptides (H12 peptide and RGD peptides) and peptide binding the P-selectin surface marker for activated platelets (DAEWVDVS) to induce platelet aggregation.32–39 More recent synthetic platelets have been engineered to induce platelet plug formation through multiple mechanisms. Heteromultivalent liposomes with dual adhesive and platelet-aggregating abilities were synthesized by conjugation of VBP, CBP, and cyclic RGD peptides to DSPE-PEG2000-COOH via their N-terminus by standard carbodiimide chemistry and subsequent incorporation into liposomes at no more than 5 mol %.38,39 In another example of liposome-based platelets with dual mechanism, Okamura et al. used H12-targeted liposomes to deliver ADP for platelet activation.40 Mechanistic studies showed that aggregation of dye-loaded H12-liposomes with activated platelets caused release of the dye, which suggested that increased platelet aggregation in the presence of H12-(ADP)-liposome was due to an aggregation-dependent release of the platelet agonist.
Polymeric nanoparticles
In one of the first reported application of synthetic polymer-based platelet substitutes, the Lavik group engineered a hemostatic nanoparticle consisting of a poly(lactic-co-glycolic acid)-poly-L-lysine (PLGA-PLL) block copolymer core (170 nm diameter) conjugated to a corona of PEG arms terminated with RGD peptides.41,42 Two PEG linkers, PEG 1500 and PEG 4600, were used to display RGD peptides, and the peptides RGD, RGDS, and GRGDS were evaluated to determine if addition of flanking residues would increase platelet aggregation. The combination of PEG 4600 linker with GRGDS peptide led to the greatest aggregation in vitro. When administered in a rat model of femoral artery injury at a concentration of 20 mg/mL in a 0.5-mL vehicle solution, 4600-GRGDS nanoparticles also showed the greatest hemostatic effect by halving bleeding time. Additionally, intravenous injections of nanoparticles after a rat liver trauma model reduced blood loss after injury and significantly increased 1-hr survival to 80% compared to 40% and 47% survival of animals injected with GRGDS-absent nanoparticles and saline controls, respectively.43
Design considerations
Particle size, shape, and flexibility
Platelets are biconvex discoids rather than spheres, and it is this discoid form which facilitates platelet margination to vessel walls.8,44 Studies by Takeoka et al. suggests that after initial attachment to surfaces, deformation of flexible membranes by hemodynamic shear flow increases the contact area between particle and tissue surfaces, providing more opportunity for reversible binding to vWF (Figure 2A).5 vWF tethering of particles slow rolling velocities5 (Figure 2B), and longer contact times at tissue surfaces then allow adhesion-stabilizing collagen-binding to occur. However, platelet-like particles have been predominantly synthesized using spherical particles with minimal optimization of particle membrane flexibility.38–41 In a recent report, a layer-by-layer (LbL) method was used to synthesize platelet-like nanoparticles (PLNs) that more closely resemble the natural platelet shape (Figure 3A) in order to investigate the effect of particle size, shape, and flexibility on vessel adhesion and hemostasis.45 Spherical polystyrene (PS) template particles were coated with alternating layers of polycationic poly(allylamine) hydrochloride (PAH) and polyanionic bovine serum albumin (BSA). After crosslinking the coating, the sacrificial PS templates were removed by dissolution, resulting in the collapse of the 4 bilayers into a flexible discoid shape. In vitro adhesion studies using microfluidic channels revealed first that adherence was greatest for 200 nm spherical particles compared to 1- and 2-μm spherical particles and second that adherence was greatest for flexible PLNs compared to rigid discs and spheres (PLN > disc > sphere, Figure 3B).45 These results indicate that discoid shape and particle flexibility contribute to particle margination and adhesion. However, the results of the particle size study conflict with previous studies reported by Charoenphol et al. in which binding efficiency of sialyl Lewisa (sLea)-coated polystyrene spheres in endothelialized microfluidic chambers increased with increasing particle size and wall shear rate for 500 nm–5 μm sized particles (Figure 3C).46 Computational modeling by Müller et al. also support greater margination of micron-sized particles than nano-sized particles to vessel walls.47 This discrepancy is likely due to the fact that the former conducted flow assays with particles suspended in saline while the work by Charoenphol et al. and Müller et al. took into consideration red blood cells (RBCs). Particle margination toward the vessel wall is largely dependent on red blood cells, which occupy the center of the blood vessel during hemodynamic flow due to lift force. The area near vessel walls are therefore an RBC-free zone which is available for particles excluded from the vessel center to occupy.48 Despite conflicting in vitro reports on size effects, greater hemostatic effect was observed in vivo for smaller-sized PLNs. In mouse tail transections, bleeding stopped more rapidly after intravenous injection of 200-nm peptide-functionalized PLNs than intravenous injections of their micron-sized or rigid counterparts (Figure 3D). The reduced efficacy of larger, micron-sized PLNs is likely due to shorter circulation time as larger particles are more easily sequestered by scavenger cells (i.e. macrophages) in the reticuloendothelial (RES) system.49–51
Figure 2.
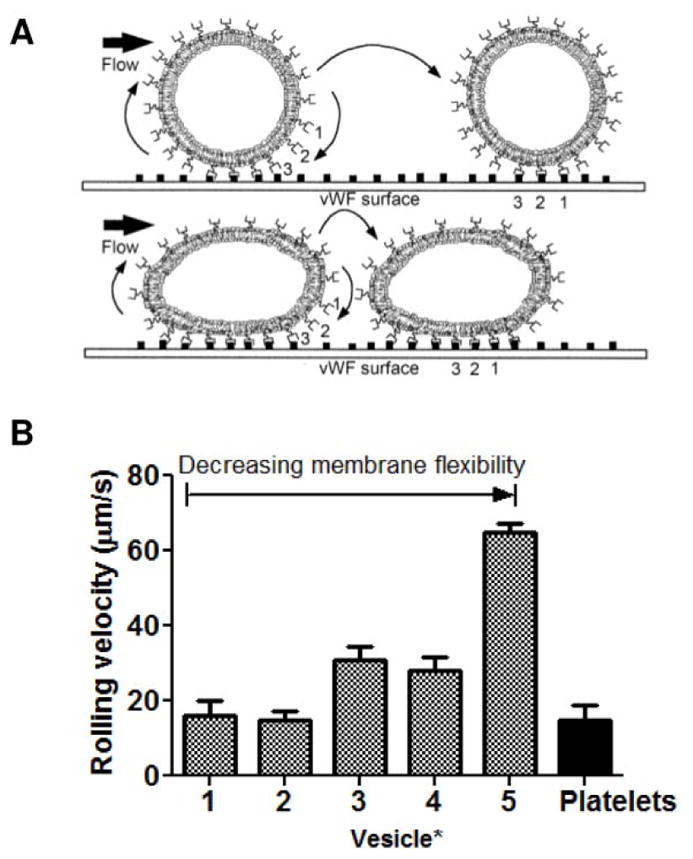
Effects of particle rigidity on rolling velocity of particles along an adhesive surface [reproduced with permission from 5]. (A) Illustration of expected rolling mechanism of rigid vWF-targeted particles (top) versus flexible vWF-targeted particles (bottom) along a vWF-coated surface. (B) Rolling velocities of liposomes with a range of membrane flexibilities. *1, EYL/DPPE = 10/1; 2, POPC/DPPE = 10/1; 3, EYL/cholesterol/DPPE = 5/5/1; 4, POPC/cholesterol/DPPE = 5/5/1; 5, DPPC/cholesterol/DPPE = 5/5/1 (by mol).
Figure 3.

Data showing effects of particle size, shape, and flexibility on adhesion to surfaces under flow and in vivo hemostatic function. (A) Synthesis of platelet-like nanoparticle (PLN) without targeting peptides. SEM imaging confirmed collapse of the flexible PAH-BSA shell after dissolution of the polystyrene core. Scale bar = 200 nm [reproduced with permission from 45]. (B) Quantification of adhesion to anti-OVA antibody-coated microfluidic chambers of OVA-covered spheres, rigid discs, and PLNS at fixed particles size (200 nm). *Denotes statistical difference (P < 0.05) from all other groups [reproduced with permission from 45]. (C) Quantification of adhesion to endothelialized microfluidic chambers of sLea spherical particles of various sizes under a range of shear rates [reproduced with permission from 46]. (D) Bleeding times after intravenous injection of 15 mg/kg PLN formulations followed by tail transections in Balb/c mice. *Denotes statistical difference (P < 0.05) from saline and PLNs without peptides [reproduced with permission from 45].
Pharmacokinetics and biodistribution are critical to the efficacy of systemically-administered hemostatic materials. After intravenous injection, hemostatic materials should circulate in the blood long enough to accumulate and act at the site of vascular injury and be excreted from the body with reasonable half-lives so as not to cause long-term risk of thrombosis. Patients sustaining traumatic injury are usually transported within an hour to a Level 1 trauma center.52 Within that hour of time, exsanguination is responsible for greater than one-third of deaths1 and is therefore a reasonable time in which intravenous hemostats should be circulating and active. Particle circulation time is dependent on a number of factors including particle size, shape, and flexibility. Nanoparticles can be rapidly removed from circulation by macrophages lining the sinusoids in the liver and in the red pulp of the spleen. This phenomenon is readily observed in biodistribution studies for platelet substitutes such as PLGA-PLL-PEG-RGD nanoparticles where nearly 70% of particles are taken up by macrophages in the liver 5 minutes after injection41 and also PLNs which demonstrate significant accumulation in the lungs, liver, and spleen 1 hour after injection. Phagocytosis of particles can be inhibited, however, by altering particle shape.51,53 Uptake is highly dependent on local particle curvature at the point of macrophage attachment, and particles with low curvature surfaces are more difficult to phagocytose than those with high curvature. For example, particles with high aspect ratios (AR), such as worm-like particles (AR>20), only have two points at either end of the worm with high curvature for phagocytosis compared to spherical particles which have equal curvature all around for easy uptake.53 Therefore, in addition to better margination under flow, elongated particles are more difficult to sequester and therefore have longer circulation times in vivo54,55. Flexible filamentous micelles have been shown to circulate for longer periods of time compared to rigid, crosslinked micelles of the same geometry.55 Therefore, membrane flexibility also appears to inhibit macrophage uptake. As previously mentioned, smaller particles are desirable to minimize sequestration by the RES system.51 Aside from better circulation time, smaller particles can prevent potential cardiopulmonary complications due to unintended blockage in the lungs and vessels.56,57 Even at submicron size (~200 nm), PLNs and PLGA-PLL-PEG-NP produced cardiopulmonary complications (evidenced by elevated heart rate and gasping) after intravenous injection of high doses.41,45
Ligand specificity
For particle-induced platelet aggregation, it is especially important that targeting ligands be specific to activated platelets to minimize risk of thrombosis. Binding to receptors on circulating quiescent platelets can cause off-target platelet aggregation and possibly even systemic platelet activation. A caveat of using RGD peptides is the lack of specificity to the GPIIb-IIIa integrin.14 The RGD motif is conserved across many extracellular matrix proteins mediating cell adhesion such as fibrinectin, victronectin, collagen, and laminin58 and is, therefore, recognized by multiple integrin receptors. Of the 5 platelet integrins, GPIIb-IIIa11,12, αvβ359, and α5β1 receptors60 have RGD-dependent binding mechanisms to their respective ligands, thus, increasing the likelihood of undesired interaction between RGD-functionalized materials and inactive platelets. However, modifications to RGD peptides, such as cyclization to restrict conformational flexibility and optimization of flanking residues, have been shown to increase specificity for certain integrin receptors.61–63 For example, liposomes functionalized with the cyclic RGD peptide, cyclo-CNPRGDY(OEt)RC, induced significant aggregation when mixed with ADP-activated platelets while inducing minimal aggregation when mixed with quiescent platelets (Figure 4A).38 Addition of flanking residues to RGD peptides have been shown to produce a more active binding conformation and induce greater activated platelet aggregation in vitro (GRGDS > RGDS > RGD) without binding and/or activing quiescent platelets (Figure 4B).41 When injected intravenously, PLGA-PLL-PEG-RGD nanoparticles (Figure 4C) with flanked RGD peptides reduced bleeding time more so than nanoparticles with non-flanked RGD peptides (Figure 4D-E), demonstrating that this increased peptide bioactivity translates to improved hemostatic function in vivo.41 As discussed in the next section, particle specificity for activated platelets can be further modulated by optimizing the linker length used to couple peptides to particle surfaces.64
Figure 4.

Data showing effects of ligand specificity and linker length on platelet aggregation and in vivo hemostatic function. Peptide modifications such as cyclization increase binding specificity of RGD moieties for activated platelets and specificity is confirmed in experiments such as that shown in (A), where inactive and ADP-activated fluorescently-labeled platelets are incubated with unmodified and cRGD-modified liposomes to confirm significant particle-induced aggregation in active platelets and minimal aggregation with inactive platelets [reproduced with permission from 38]. (B) In vitro comparison of platelet aggregation produced by incubation of fluorescently-labeled platelets with RGD versus flanked RGD peptides attached to 1500 or 4600 Da PEG. *P < 0.05 for comparison to PEG 4600 alone [reproduced with permission from 41]. (C) Schematic of PLGA-PLL-PEG-RGD nanoparticles [reproduced with permission from 41]. Bleeding times of rats intravenously injected with 20 mg/mL suspension of PLGA-PLL-PEG-RGD nanoparticles synthesized with (D) PEG 1500 and (E) PEG 4600 followed by injury to the femoral artery [reproduced with permission from 41]. (D) *P < 0.01, ***P < 0.001 for comparison to PEG 1500. (E) * P < 0.05, ***P < 0.001 for comparison to saline and #P < 0.05 for comparison to rFVIIa.
Linker length
Thromboerythrocytes development revealed that interaction between RGD peptide ligands and platelets is highly dependent on linker length 21,64. The length of the glycine linker or n in the peptide (G)n-RGDF was shown to determine the extent of interaction between peptide-functionalized surfaces and platelets.64 When conjugated with RGD peptides with linker lengths of 1, 3, and 9, polyacrylonitrile beads demonstrated little interaction with platelets, highly selective aggregation with active platelets, and strong interaction with both active and inactive platelets, respectively. A 3-residue linker (CGG) was therefore used in the preparation of the thromboerythrocytes by Coller et al.21 The positive correlation between linker length and platelet interaction is likely due to closer proximity of RGD to receptors on platelet surfaces, increased conformational flexibility for binding, increased potential for assuming a secondary structure, or a combination of these factors.64 Consistent with these observations, longer PEG (4600 Da) used in the construction of PLGA-PLL-PEG-RGD nanoparticles produced greater platelet aggregation in vitro than shorter PEG (1500 Da) (Figure 4B) and reduced bleeding time to a greater degree in animal models of femoral artery injury (Figure 4D-E). For most liposomal formulations, peptides (VBP, CBP, and cyclic RGD) were conjugated to DSPE-PEG2000.36–38
Ligand or receptor density
Ligand density has been shown to be an important factor when engineering targeted nanoparticles for application in drug delivery and imaging65–70 and is, likewise, critical when engineering hemostatic particles mimicking platelet adhesion or platelet aggregation as demonstrated in multiple studies9,39,71,72. When flowed through a perfusion chamber, liposomes of varying rGPIbα surface densities (0, 5.27 x 103, 1.00 x 104 molecules per liposome) and fixed rGPIα-IIα density (2 x 103 molecules per liposome) showed rGPIbα density-dependent adhesion of collagen-immobilized surfaces in the presence of soluble vWF.9 At high shear rates (1200 and 2400 s−1), surface adhesion by functionalized liposomes increased with increasing rGPIbα. Interestingly, liposomes with the higher rGPIbα density had increasing adhesion with increasing shear whereas liposomes with no rGPIbα or the lower rGPIbα density demonstrated decreasing surface coverage with increasing shear. Similarly, experiments with varying rGPIα-IIα (0, 0.96 x 103, 2.17 x 103 molecules per liposome) and fixed rGPIbα (1.00 x 104 molecules per liposome) showed rGPIα-IIα density-dependent adhesion of collagen surfaces by functionalized liposomes. Liposomes with rGPIbα alone demonstrated transient adherence but no stable adhesion. These studies suggest that higher ligand density for targeting vWF and collagen is ideal for initial particle tethering and subsequent adhesion stabilization, especially under high shear rates.
For PLGA-PLL-PEG nanoparticle optimization, increasing GRGDS peptide content on nanoparticles by100-fold decreased the required in vitro and in vivo dose for achieving hemostasis by 10-fold and 8-fold, respectively.43,71 However, dose-dependent adverse effects were observed with the high density GRGDS formulation (GRGDS-NP100) compared to the lower-density formation (GRGDS-NP1). For example, while GRGDS-NP100 at 5 mg/kg doses could produce similar levels of improved 1-hr survival and reduced blood loss in rat liver trauma models as GRGDS-NP1 at 40 mg/kg, increasing the dose of GRGDS-NP100 to 20 mg/kg or 40 mg/kg drastically reduced percentage of animals surviving to 1 hour (Figure 5A-B). Adverse effects at high GRGDS-NP100 dosages can be attributed to saturation of GPIIb-IIIa receptors on activated platelets, which would inhibit platelet-platelet interaction, thus, preventing platelet aggregation and inducing anticoagulation (Figure 5C). This same phenomena was observed by Coller et al., who reported the loss of thromboerythrocyte aggregation with platelets in wells coated with high concentrations of fibrinogen.72 These studies are important in highlighting that increased hemostatic potency from high ligand density is offset by safety concerns of bleeding out due to overdosing.
Figure 5.

Data showing the effects of GRGDS ligand density on in vivo hemostatic function. Comparison between 1-hour survival (top) and blood loss (bottom) of lethal liver injury rat models after intravenous injection of (A) 40 mg/kg GRGDS-NP1 and (B) 5, 20, and 40 mg/kg GRGDS-NP100 demonstrates an 8-fold reduction in effective particle dosage (i.e. increased potency) when ligand density is increased 100-fold [reproduced with permission from 43,71]. However, adverse effects are observed at 20 and 40 mg/kg GRGDS-NP100 most likely due to saturation of GPIIb-IIIa receptors on platelets which would prevent platelet aggregation (C). (B) *P < 0.05 compared to saline.
Ligand synergism
vWF- and collagen-binding work synergistically for platelet adhesion, and are, therefore, generally coupled on heteromultivalent platelet substitutes (Figure 6A).9,38,39,45 When compared to liposomes with VBP or CBP alone, heteromultivalent liposomes with both peptides had significantly greater adhesion to vWF- and collagen-coated plates at all tested shear rates (5, 30, 55 dynes/cm2).39 The optimal ratio of vWF- to collagen-targeting ligand was determined using liposomes with varying ratios of DSPE-PEG-VBP and DSPE-PEG-CBP (80:20, 60:40, 50:50, 40:60, 20:80) at a fixed 5 mol % DSPE-PEG-peptide composition (Figure 6B-D).39 At low shear rate, liposomes with a greater proportion of CBP had the greatest adhesion at 30 min and retention at 45 min (Figure 6B) while those with a greater proportion of VBP had the greatest adhesion and retention at high shear rate (Figure 6D). At medium shear rate, near 1:1 ratio of CBP to VBP yielded the greatest adhesion and retention (Figure 6C). These studies demonstrate that collagen-binding is the dominant mechanism for adhesion under low shear while vWF-binding is the dominant mechanism under high shear. Furthermore, the ratio of targeting ligands can be optimized to maximize the adhesiveness of particle platforms.
Figure 6.

Data showing the effects of ligand synergism on platelet adhesion. (A) Synergistic ligands can be incorporated into heteromultivalent liposomes using a mixture of DSPE-PEG-peptide conjugates during liposome synthesis [reproduced with permission from 38]. Liposomes with fixed 5 mol% DSPE-PEG-peptide was synthesized with varying ratios DSPE-PEG-CBP to DSPE-PEG-VBP to determine the optimal ratio for maximizing liposome adhesion to the subendothelial matrix. Liposomes were flowed through a parallel plate fluidic chamber over glass slides coated with 1:1 ratio of vWF to collagen under a range of shear stresses. Adhesion at (B) 5 dynes/cm2, (C) 30 dynes/cm2, and (D) 55 dynes/cm2 was quantified by measuring fluorescence from fluorescently-labeled liposomes [reproduced with permission from 39].
FIBRIN-CROSSLINKING AGENTS
Secondary hemostasis
During secondary hemostasis, the fibrin matrix is formed by the coagulation cascade (Figure 1B). Fibrinogen is the 340 kDa glycoprotein precursor of fibrin and, under normal physiologic conditions, circulates in the blood at an average concentration of 3.0 mg/mL.73 Fibrinogen is made of three pairs of polypeptide chains– Aα, Bβ, and γ.74 It has a symmetric elongated structure made up of three nodules, a central E nodule containing the N-termini of all six chains from which the chains extend as two sets of three-chained coiled coils into two distal D nodules containing the C-termini of the Bβ and γ chains (βC and γC). The Aα chains exit the D nodules and travel back to the E nodule where their C-termini (αC) interact with each other and the E nodule. After tissue damage, thrombin is activated locally as a result of the coagulation cascade and cleaves fibrinopeptides A and B (FPA and FPB) from the N-termini of Aα and Bβ chains, respectively, exposing A and B knob peptide domains. Knobs A and B, in what are now fibrin monomers, interact with holes a and b in γC and βC, respectively, resulting in the formation of half-staggered, double-stranded fibrin protofibrils. Non-specific interactions of αC cause protofibrils to aggregate laterally, bundle into fibers, and branch to form a three-dimensional insoluble hydrogel scaffold for platelets, blood cells, and other clot components. The transglutaminase Factor XIIIa (FXIIIa) then stabilizes fibrin by creating intra- and inter-fiber crosslinks through amide bond formation between lysine and glutamic acid residues.
Polymers for modulating formation of the fibrin matrix
Difficulty in clotting, or coagulopathy, can result from the absence, deficiency, or dysfunction of clotting factor(s) which participate in the coagulation cascade ultimately leading to thrombin activation. A shortage of functional clotting factors can be due to genetic defect as in hemophilia, the result of large volume blood loss as in trauma-induced coagulopathy (TIC), or the result of anticoagulant use (e.g. warfarin). When thrombin activation is inhibited, fibrin formation is impeded. Fibrin matrices that form at low thrombin levels are more porous and made up of loose, thick fibrin fibers as opposed to denser networks of thin fibers.75–77 The former, in addition to producing mechanically weaker gels for maintaining hemostasis, is also more susceptible to enzymatic degradation (i.e. fibrinolysis).78 Therefore, affecting fibrin formation using materials that mimic clotting factors is another strategy for inducing strong clot formation for hemostasis.
Early successful attempts at developing a substitute for the thrombin-modified fibrinogen E domain using synthetic polymer constructs were reported by Lorand et al.79 Using the synthetic peptide mimic of knob A (GPRP)80, Lorand et al. synthesized a double-headed ligand bis(Gly-Pro-Arg-Pro-amido)polyethylene glycol and showed that it could replace the thrombin-modified E nodule for non-covalent fibrinogen-fibrinogen or D-D crosslinking via interaction with two hole a’s in neighboring γ-chains (Figure 7A).79 Presence of free knob A peptide mimic in a mixture of thrombin and fibrinogen inhibits clot formation due to competition with fibrin monomers for hole a.80 However, as demonstrated by Lorand et al., a bivalent construct terminated with knob A peptides is able to crosslink fibrin. Furthermore, this behavior was biphasic with “productive” crosslink-forming behavior at the lower polymer concentration range and “non-productive” crosslink-absent behavior at the higher polymer concentration range (Figure 7B). This biphasic behavior was attributed to inter-polymer competition for available hole a’s (two per fibrinogen molecule) making it difficult for the unbound end of a bound polymer to find a second hole a for crosslinking. Additionally, these studies showed that 900 Da PEGs were large enough to span the distance of two hole a’s (a minimum of 32 Å)81, an important engineering design specification for future fibrin-crosslinking constructs. Lorand et al. were the first to demonstrate in vitro that fibrin polymerization in the absence of thrombin could be driven by synthetic polymers. However, due to their ability to crosslink fibrinogen, knob A-terminated polymers should be limited to local administration rather than systemic administration.
Figure 7.
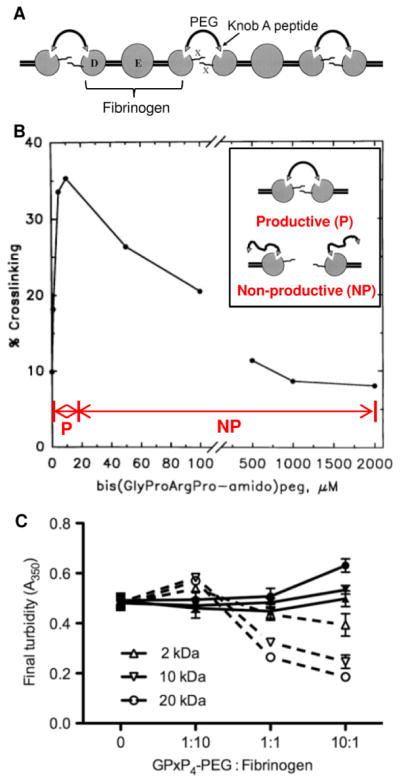
Knob A-peptide terminated polymers for fibrin(ogen) crosslinking in vitro. (A) A bivalent 900 Da PEG terminated on either end with knob A peptide (GPRP), bis(GlyProArgPro-amido)peg, was synthesized to crosslink fibrinogen D nodules in the absence of thrombin. (B) Bis(GlyProArgPro-amido)peg demonstrated biphasic behavior where lower “productive” polymer concentrations formed D-D crosslinks and higher “non-productive” polymer concentrations showed less efficient crosslinking due to competition of polymers for the finite number of hole a’s. (A-B) Reproduced with permission from 79. Copyright © 1998 National Academy of Sciences, U.S.A. (C) Biphasic behavior was also observed in turbidity measurements for fibrin formed with 4-arm PEG-knob A conjugates of various molecular weights. Filled in symbols and solid lines represent scrambled conjugate controls and empty symbols and dotted lines represent knob A conjugates [reproduced with permission from 82].
In subsequent work, Soon et al. conjugated cysteine-terminated knob A peptide mimics (GPRPAAC) to two-arm and four-arm maleimide-functionalized PEGs (2–20 kDa). The conjugates were evaluated in vitro with thrombin and were used to modulate fibrin structure for tissue engineering applications rather than drive thrombin-independent fibrin polymerization.82 Biphasic behavior was again observed in these studies. At higher molar ratios (1:1 and 10:1) of GPRP4-PEG to fibrinogen, final clot turbidities were significantly reduced which is likely due to inhibited fibrin formation evidenced by prolonged clotting times and the reduction of clottable protein from >90% to 70%. This behavior is, again, likely due to competition between conjugates as well as competition with naturally-occurring knob:hole interactions. At the lower 1:10 molar ratio of GPRP4-PEG to fibrinogen, final clot turbidities were slightly increased indicating larger fiber diameters and higher fiber density (Figure 7C). The behavior seen at the 1:10 molar ratio suggests that lower concentrations of polymers might have resulted in the same “productive” behavior observed in the work by Lorand et al. However, further investigation was completed only at the 1:1 molar ratio which continued to show “non-productive” behavior as evidenced by the reduced elastic modulus (i.e. stiffness) of fibrin. Later work from this group reported the use of PEGylated knob peptides as anticoagulants.83
Due to the accessibility of hole a’s in both fibrin and fibrinogen, knob A-terminated polymers can cause thrombosis (off-target clot formation) if administered systemically and are, therefore, not suitable as intravenous hemostats. However, knob A peptides can be readily substituted with other peptides or proteins that specifically bind fibrin and not fibrinogen to create hemostatic polymers that only induce clotting at sites of injury. Recently, our lab reported the use of fibrin-binding peptides identified via phage display by Kolodziej et al84–87 to create linear polymer hemostats (PolySTAT) for crosslinking fibrin (Figure 8A).88 PolySTAT is composed primarily of a poly(hydroxyethyl)methacrylate [p(HEMA)] backbone (80%) with pendant cyclic fibrin-binding peptides conjugated to N-hydroxysuccinimidyl ester methacrylate (NHSMA) comonomers. Polymers with average molecular weight of 45 kDa and ~ 16 peptides were synthesized by reversible addition-fragmentation chain-transfer (RAFT) polymerization. Confocal images of fluorescently-labeled fibrin formed with fluorescent PolySTAT showed integration of the polymer into fibrin fibers, and SEM images showed altered fibrin networks which were denser and less porous than controls (Figure 8B). Elastic moduli of PolySTAT-crosslinked fibrin was increased 2–3 fold, and with the addition of plasmin into the purified system, the PolySTAT-integrated fibrin network showed significantly reduced fibrinolysis compared to scrambled controls. Increased mechanical strength as well as resistance to enzymatic breakdown is particularly important to resolve bleeding in conditions such as TIC, where clot strength is reduced and hyperfibrinolytic activity may be observed.89,90 Intravenous injection of PolySTAT at a dose of 15 mg/kg in rat models of femoral artery injury and fluid resuscitation resulted in significantly greater survival rates compared to scrambled controls (Figure 8C) as well as reduced blood loss (Figure 8D) and fluid resuscitation requirements to maintain blood pressure above 60 mm Hg.
Figure 8.

In vitro characterization of PolySTAT-modified fibrin and in vivo evaluation of hemostatic efficacy. (A) PolySTAT is a synthetic poly(HEMA) polymer with multivalent display of fibrin-binding peptides (shown in red). (B) PolySTAT-modified fibrin has a denser mesh structure compared to control fibrin (SEM scalebar = 10 μm). Intravenous injection of PolySTAT in a rat femoral injury model significantly (C) increases survival rate and (D) reduces hemorrhage rate [reproduced with permission from 88].
Fibrin-binding microgel particles
In another recent publication, ultra-low crosslinked (ULC) poly(N-isopropylacrylamide-co-acrylic acid) (pNIPAm-AAc) microgel particles with surface-conjugated single domain variable fragments (sdFv’s) with affinity for fibrin, termed platelet-like particles (PLPs), were reported for their application as intravenous hemostats in injured rat models (Figure 9A).91 PLPs were engineered with low core crosslinking densities (<0.5%) to create particle bodies with high deformability to mimic platelet deformation upon fibrin formation. Despite their given name, PLPs are technically unresponsive until secondary hemostasis and were included in this section because of their enhancement of fibrin formation rather than initial platelet plug formation. PLPs added to platelet-poor plasma (PPP) promoted fibrin formation in an endothelialized microfluidic device to nearly comparable fibrin levels observed with platelet-rich plasma (PRP) and, similar to PolySTATs, produced denser fibrin networks (Figure 9B). Bleeding times were halved in rat models of femoral vein injury. In addition to high deformability, PLPs were likened to platelets because of clot collapse that was observed 24–48 h after clot formation (Figure 9C). A caveat, as the authors also noted, is that clot retraction due to actomyosin contraction in native platelets attached to the fibrin matrix occurs significantly more rapidly (within minutes to an hour after clot formation)92.
Figure 9.
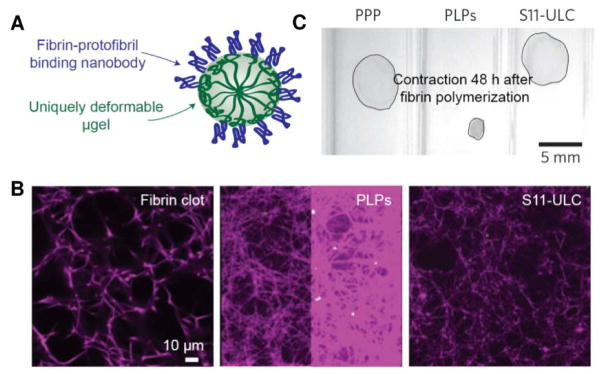
In vitro characterization of PLP-modified fibrin. (A) PLPs have multivalent display of fibrin-binding nanobodies. (B) PLP increases the density of the fibrin matrix in comparison to non-binding microgel controls (S11-ULC). (C) PLP integration in clots results in contraction 48 hours after fibrin polymerization [reproduced with permission from 91].
CONCLUSIONS
Current methods of resolving bleeding after traumatic injury include using topical hemostats which are limited to treating accessible wounds and by transfusion of blood products or recombinant clotting factors to restore hemostatic function. Biologically-derived products have many limitations such as short shelf life, involved manufacturing processes, and risk of viral transmission. Therefore, there has been growing interest in developing synthetic hemostatic agents for systemic administration to resolve bleeding in less accessible injuries (e.g. internal bleeding in the trunk). Numerous platelet substitutes have been developed using nanoparticles commonly used in drug delivery such as liposomes and PLGA nanoparticles and have been evaluated on their ability to mimic platelet margination and adhesion to vessel walls and to induce platelet aggregation. Investigation of these platelet-like constructs have demonstrated the importance of physical parameters (i.e. particle size, shape, and flexibility) in platelet mimicry as well as methodical approaches to combining and displaying binding motifs for optimal hemostatic activity specific to the site of vascular injury. With more recent materials, we have seen emergent properties mimicking platelet contraction and deformation. Dynamic particles capable of mimicking platelet contraction at the necessary timescale could potentially stiffen clots to the same degree as native platelet contraction, and thus induce hemostasis more rapidly. Recent materials have also started engaging fibrin to stabilize clot structure and provide an orthogonal approach to hemostasis. Future work on intravenous hemostats will likely focus on the amalgamation of both optimized physical particle properties and binding motif arrangement into one platelet construct. Furthermore, future hemostatic agents may be engineered to engage both primary and secondary hemostasis mechanisms for a stronger hemostatic response.
Acknowledgments
This work is supported by NIH NIBIB 1R21EB018637 and the Bioengineering Cardiovascular Training Grant NIH 2T32EB001650-06A2 (LWC).
Footnotes
SUPPORTING INFORMATION
No supporting information is available.
References
- 1.Kauvar DS, Lefering R, Wade CE. Impact of hemorrhage on trauma outcome: an overview of epidemiology, clinical presentations, and therapeutic considerations. J Trauma. 2006;60:S3–11. doi: 10.1097/01.ta.0000199961.02677.19. [DOI] [PubMed] [Google Scholar]
- 2.Eastridge BJ, Mabry RL, Seguin P, Cantrell J, Tops T, Uribe P, Mallett O, Zubko T, Oetjen-Gerdes L, Rasmussen TE, Butler FK, Kotwal RS, Kotwal RS, Holcomb JB, Wade C, Champion H, Lawnick M, Moores L, Blackbourne LH. Death on the battlefield (2001–2011): implications for the future of combat casualty care. J Trauma Acute Care Surg. 2012;73:S431–7. doi: 10.1097/TA.0b013e3182755dcc. [DOI] [PubMed] [Google Scholar]
- 3.Achneck HE, Sileshi B, Jamiolkowski RM, Albala DM, Shapiro ML, Lawson JH. A comprehensive review of topical hemostatic agents: efficacy and recommendations for use. Ann Surg. 2010;251:217–228. doi: 10.1097/SLA.0b013e3181c3bcca. [DOI] [PubMed] [Google Scholar]
- 4.Spahn DR, Bouillon B, Cerny V, Coats TJ, Duranteau J, Fernandez-Mondejar E, Filipescu D, Hunt BJ, Komadina R, Nardi G, Neugebauer E, Ozier Y, Riddez L, Schultz A, Vincent JL, Rossaint R. Management of bleeding and coagulopathy following major trauma: an updated European guideline. Crit Care. 2013;17:R76. doi: 10.1186/cc12685. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Takeoka S, Teramura Y, Okamura Y, Tsuchida E, Handa M, Ikeda Y. Rolling properties of rGPIbalpha-conjugated phospholipid vesicles with different membrane flexibilities on vWf surface under flow conditions. Biochem Biophys Res Commun. 2002;296:765–770. doi: 10.1016/s0006-291x(02)00934-8. [DOI] [PubMed] [Google Scholar]
- 6.Pease DC. An electron microscope study of red bone marrow. Blood. 1956;11:501–526. [PubMed] [Google Scholar]
- 7.Machlus KR, Italiano JE., Jr The incredible journey: From megakaryocyte development to platelet formation. J Cell Biol. 2013;201:785–796. doi: 10.1083/jcb.201304054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Gentile F, Chiappini C, Fine D, Bhavane RC, Peluccio MS, Cheng MMC, Liu X, Ferrari M, Decuzzi P. The effect of shape on the margination dynamics of non-neutrally buoyant particles in two-dimensional shear flows. J Biomech. 2008;41:2312–2318. doi: 10.1016/j.jbiomech.2008.03.021. [DOI] [PubMed] [Google Scholar]
- 9.Nishiya T, Kainoh M, Murata M, Handa M, Ikeda Y. Reconstitution of adhesive properties of human platelets in liposomes carrying both recombinant glycoproteins Ia/IIa and Ib alpha under flow conditions: specific synergy of receptor-ligand interactions. Blood. 2002;100:136–142. doi: 10.1182/blood.v100.1.136. [DOI] [PubMed] [Google Scholar]
- 10.Chen H, Locke D, Liu Y, Liu C, Kahn ML. The platelet receptor GPVI mediates both adhesion and signaling responses to collagen in a receptor density-dependent fashion. J Biol Chem. 2002;277:3011–3019. doi: 10.1074/jbc.M109714200. [DOI] [PubMed] [Google Scholar]
- 11.Pytela R, Pierschbacher MD, Ginsberg MH, Plow EF, Ruoslahti E. Platelet membrane glycoprotein IIb/IIIa: member of a family of Arg-Gly-Asp--specific adhesion receptors. Science (80-) 1986;231:1559–1562. doi: 10.1126/science.2420006. [DOI] [PubMed] [Google Scholar]
- 12.Plow EF, Ginsberg MH. Cellular adhesion: GPIIb-IIIa as a prototypic adhesion receptor. Prog Hemost Thromb. 1989;9:117–156. [PubMed] [Google Scholar]
- 13.Salsmann A, Schaffner-Reckinger E, Kabile F, Plancon S, Kieffer N. A new functional role of the fibrinogen RGD motif as the molecular switch that selectively triggers integrin alphaIIbbeta3-dependent RhoA activation during cell spreading. J Biol Chem. 2005;280:33610–33619. doi: 10.1074/jbc.M500146200. [DOI] [PubMed] [Google Scholar]
- 14.Modery-Pawlowski CL, Tian LL, Pan V, McCrae KR, Mitragotri S, Sen Gupta A. Approaches to synthetic platelet analogs. Biomaterials. 2013;34:526–541. doi: 10.1016/j.biomaterials.2012.09.074. [DOI] [PubMed] [Google Scholar]
- 15.Wagner CL, Mascelli MA, Neblock DS, Weisman HF, Coller BS, Jordan RE. Analysis of GPIIb/IIIa receptor number by quantification of 7E3 binding to human platelets. Blood. 1996;88:907–914. [PubMed] [Google Scholar]
- 16.Bennett JS, Vilaire G. Exposure of platelet fibrinogen receptors by ADP and epinephrine. J Clin Invest. 1979;64:1393–1401. doi: 10.1172/JCI109597. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Marguerie GA, Edgington TS, Plow EF. Interaction of fibrinogen with its platelet receptor as part of a multistep reaction in ADP-induced platelet aggregation. J Biol Chem. 1980;255:154–161. [PubMed] [Google Scholar]
- 18.Niiya K, Hodson E, Bader R, Byers-Ward V, Koziol JA, Plow EF, Ruggeri ZM. Increased surface expression of the membrane glycoprotein IIb/IIIa complex induced by platelet activation. Relationship to the binding of fibrinogen and platelet aggregation. Blood. 1987;70:475–483. [PubMed] [Google Scholar]
- 19.Agam G, Livne A. Passive participation of fixed platelets in aggregation facilitated by covalently bound fibrinogen. Blood. 1983;61:186–191. [PubMed] [Google Scholar]
- 20.Agam G, Livne AA. Erythrocytes with covalently bound fibrinogen as a cellular replacement for the treatment of thrombocytopenia. Eur J Clin Invest. 1992;22:105–112. doi: 10.1111/j.1365-2362.1992.tb01943.x. [DOI] [PubMed] [Google Scholar]
- 21.Coller BS, Springer KT, Beer JH, Mohandas N, Scudder LE, Norton KJ, West SM. Thromboerythrocytes. In vitro studies of a potential autologous, semi-artificial alternative to platelet transfusions. J Clin Invest. 1992;89:546–555. doi: 10.1172/JCI115619. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Levi M, Friederich PW, Middleton S, de Groot PG, Wu YP, Harris R, Biemond BJ, Heijnen HF, Levin J, ten Cate JW. Fibrinogen-coated albumin microcapsules reduce bleeding in severely thrombocytopenic rabbits. Nat Med. 1999;5:107–111. doi: 10.1038/4795. [DOI] [PubMed] [Google Scholar]
- 23.Davies AR, Judge HM, May JA, Glenn JR, Heptinstall S. Interactions of platelets with Synthocytes, a novel platelet substitute. Platelets. 2002;13:197–205. doi: 10.1080/0953371027000. [DOI] [PubMed] [Google Scholar]
- 24.Allen TM, Cullis PR. Liposomal drug delivery systems: from concept to clinical applications. Adv Drug Deliv Rev. 2013;65:36–48. doi: 10.1016/j.addr.2012.09.037. [DOI] [PubMed] [Google Scholar]
- 25.Sie P, Gillois M, Boneu B, Chap H, Bierme R, Douste-Blazy L. Reconstitution of liposomes bearing platelet receptors for human von Willebrand factor. Biochem Biophys Res Commun. 1980;97:133–138. doi: 10.1016/s0006-291x(80)80145-8. [DOI] [PubMed] [Google Scholar]
- 26.Baldassare JJ, Kahn RA, Knipp MA, Newman PJ. Reconstruction of platelet proteins into phospholipid vesicles. Functional proteoliposomes. J Clin Invest. 1985;75:35–39. doi: 10.1172/JCI111693. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Parise LV, Phillips DR. Platelet membrane glycoprotein IIb-IIIa complex incorporated into phospholipid vesicles. Preparation and morphology. J Biol Chem. 1985;260:1750–1756. [PubMed] [Google Scholar]
- 28.Rybak ME. A platelet substitute-the plateletsome to be used in transfusion therapy. US Army Medical Research and Development Command; Fort Detrick, Frederick, Maryland: 1989. pp. 1–41. [Google Scholar]
- 29.Rybak ME, Renzulli LA. A liposome based platelet substitute, the plateletsome, with hemostatic efficacy. Biomater Artif Cells Immobil Biotechnol. 1993;21:101–118. doi: 10.3109/10731199309117350. [DOI] [PubMed] [Google Scholar]
- 30.Kitaguchi T, Murata M, Iijima K, Kamide K, Imagawa T, Ikeda Y. Characterization of liposomes carrying von Willebrand factor-binding domain of platelet glycoprotein Ibalpha: a potential substitute for platelet transfusion. Biochem Biophys Res Commun. 1999;261:784–789. doi: 10.1006/bbrc.1999.1088. [DOI] [PubMed] [Google Scholar]
- 31.Nishiya T, Kainoh M, Murata M, Handa M, Ikeda Y. Platelet interactions with liposomes carrying recombinant platelet membrane glycoproteins or fibrinogen: approach to platelet substitutes. Artif Cells Blood Substit Immob Biotechnol. 2001;29:453–464. doi: 10.1081/bio-100108550. [DOI] [PubMed] [Google Scholar]
- 32.Okamura Y, Maekawa I, Teramura Y, Maruyama H, Handa M, Ikeda Y, Takeoka S. Hemostatic effects of phospholipid vesicles carrying fibrinogen gamma chain dodecapeptide in vitro and in vivo. Bioconjug Chem. 2005;16:1589–1596. doi: 10.1021/bc050178g. [DOI] [PubMed] [Google Scholar]
- 33.Okamura Y, Takeoka S, Teramura Y, Maruyama H, Tsuchida E, Handa M, Ikeda Y. Hemostatic effects of fibrinogen gamma-chain dodecapeptide-conjugated polymerized albumin particles in vitro and in vivo. Transfusion. 2005;45:1221–1228. doi: 10.1111/j.1537-2995.2005.00173.x. [DOI] [PubMed] [Google Scholar]
- 34.Okamura Y, Fujie T, Maruyama H, Handa M, Ikeda Y, Takeoka S. Prolonged hemostatic ability of polyethylene glycol-modified polymerized albumin particles carrying fibrinogen gamma-chain dodecapeptide. Transfusion. 2007;47:1254–1262. doi: 10.1111/j.1537-2995.2007.01265.x. [DOI] [PubMed] [Google Scholar]
- 35.Okamura Y, Fujie T, Nogawa M, Maruyama H, Handa M, Ikeda Y, Takeoka S. Haemostatic effects of polymerized albumin particles carrying fibrinogen gamma-chain dodecapeptide as platelet substitutes in severely thrombocytopenic rabbits. Transfus Med. 2008;18:158–166. doi: 10.1111/j.1365-3148.2008.00860.x. [DOI] [PubMed] [Google Scholar]
- 36.Modery CL, Ravikumar M, Wong TL, Dzuricky MJ, Durongkaveroj N, Sen Gupta A. Heteromultivalent liposomal nanoconstructs for enhanced targeting and shear-stable binding to active platelets for site-selective vascular drug delivery. Biomaterials. 2011;32:9504–9514. doi: 10.1016/j.biomaterials.2011.08.067. [DOI] [PubMed] [Google Scholar]
- 37.Ravikumar M, Modery CL, Wong TL, Dzuricky M, Sen Gupta APD. Mimicking Adhesive Functionalities of Blood Platelets using Ligand-Decorated Liposomes. Bioconjug Chem. 2012;23:1266–1275. doi: 10.1021/bc300086d. [DOI] [PubMed] [Google Scholar]
- 38.Ravikumar M, Modery CL, Wong TL, Gupta AS. Peptide-decorated liposomes promote arrest and aggregation of activated platelets under flow on vascular injury relevant protein surfaces in vitro. Biomacromolecules. 2012;13:1495–1502. doi: 10.1021/bm300192t. [DOI] [PubMed] [Google Scholar]
- 39.Modery-Pawlowski CL, Tian LL, Ravikumar M, Wong TL, Sen Gupta A. In vitro and in vivo hemostatic capabilities of a functionally integrated platelet-mimetic liposomal nanoconstruct. Biomaterials. 2013;34:3031–3041. doi: 10.1016/j.biomaterials.2012.12.045. [DOI] [PubMed] [Google Scholar]
- 40.Okamura Y, Takeoka S, Eto K, Maekawa I, Fujie T, Maruyama H, Ikeda Y, Handa M. Development of fibrinogen gamma-chain peptide-coated, adenosine diphosphate-encapsulated liposomes as a synthetic platelet substitute. J Thromb Haemost. 2009;7:470–477. doi: 10.1111/j.1538-7836.2008.03269.x. [DOI] [PubMed] [Google Scholar]
- 41.Bertram JP, Williams CA, Robinson R, Segal SS, Flynn NT, Lavik EB. Intravenous hemostat: nanotechnology to halt bleeding. Sci Transl Med. 2009;1:11ra22. doi: 10.1126/scitranslmed.3000397. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Hans ML, AML Biodegradable nanoparticles for drug delivery and targeting. Curr Opin Solid State Mater Sci. 2002;6:319. [Google Scholar]
- 43.Shoffstall AJ, Atkins KT, Groynom RE, Varley ME, Everhart LM, Lashof-Sullivan MM, Martyn-Dow B, Butler RS, Ustin JS, Lavik EB. Intravenous hemostatic nanoparticles increase survival following blunt trauma injury. Biomacromolecules. 2012;13:3850–3857. doi: 10.1021/bm3013023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Doshi N, Prabhakarpandian B, Rea-Ramsey A, Pant K, Sundaram S, Mitragotri S. Flow and adhesion of drug carriers in blood vessels depend on their shape: A study using model synthetic microvascular networks. J Control Release. 2010;146:196–200. doi: 10.1016/j.jconrel.2010.04.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Anselmo AC, Modery-Pawlowski CL, Menegatti S, Kumar S, Vogus DR, Tian LL, Chen M, Squires TM, Sen Gupta A, Mitragotri S. Platelet-like Nanoparticles: Mimicking Shape, Flexibility, and Surface Biology of Platelets To Target Vascular Injuries. ACS Nano. 2014;8:11243–53. doi: 10.1021/nn503732m. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Charoenphol P, Huang RB, Eniola-Adefeso O. Potential role of size and hemodynamics in the efficacy of vascular-targeted spherical drug carriers. Biomaterials. 2010;31:1392–402. doi: 10.1016/j.biomaterials.2009.11.007. [DOI] [PubMed] [Google Scholar]
- 47.Muller K, Fedosov DA, Gompper G. Margination of micro- and nano-particles in blood flow and its effect on drug delivery. Sci Rep. 2014;4:4871. doi: 10.1038/srep04871. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Müller K, Fedosov Da, Gompper G. Margination of micro- and nano-particles in blood flow and its effect on drug delivery. Sci Rep. 2014;4:4871. doi: 10.1038/srep04871. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Decuzzi P, Pasqualini R, Arap W, Ferrari M. Intravascular delivery of particulate systems: does geometry really matter? Pharm Res. 2009;26:235–43. doi: 10.1007/s11095-008-9697-x. [DOI] [PubMed] [Google Scholar]
- 50.Jones SW, Roberts RA, Robbins GR, Perry JL, Kai MP, Chen K, Bo T, Napier ME, Ting JP, Desimone JM, Bear JE. Nanoparticle clearance is governed by Th1/Th2 immunity and strain background. J Clin Invest. 2013;123:3061–3073. doi: 10.1172/JCI66895. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Champion Ja, Mitragotri S. Role of target geometry in phagocytosis. Proc Natl Acad Sci U S A. 2006;103:4930–4. doi: 10.1073/pnas.0600997103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Swaroop M, Straus DC, Agubuzu O, Esposito TJ, Schermer CR, Crandall ML. Pre-hospital transport times and survival for Hypotensive patients with penetrating thoracic trauma. J Emerg Trauma Shock. 2013;6:16–20. doi: 10.4103/0974-2700.106320. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Champion Ja, Mitragotri S. Shape induced inhibition of phagocytosis of polymer particles. Pharm Res. 2009;26:244–9. doi: 10.1007/s11095-008-9626-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Arnida Janát-Amsbury MM, Ray a, Peterson CM, Ghandehari H. Geometry and surface characteristics of gold nanoparticles influence their biodistribution and uptake by macrophages. Eur J Pharm Biopharm. 2011;77:417–23. doi: 10.1016/j.ejpb.2010.11.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Geng Y, Dalhaimer P, Cai S, Tsai R, Tewari M, Minko T, Discher DE. Shape effects of filaments versus spherical particles in flow and drug delivery. Nat Nanotechnol. 2007;2:249–255. doi: 10.1038/nnano.2007.70. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Dobrovolskaia MA, Aggarwal P, Hall JB, McNeil SE. Preclinical studies to understand nanoparticle interaction with the immune system and its potential effects on nanoparticle biodistribution. Mol Pharm. 2008;5:487–495. doi: 10.1021/mp800032f. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.He C, Hu Y, Yin L, Tang C, Yin C. Effects of particle size and surface charge on cellular uptake and biodistribution of polymeric nanoparticles. Biomaterials. 2010;31:3657–66. doi: 10.1016/j.biomaterials.2010.01.065. [DOI] [PubMed] [Google Scholar]
- 58.Bennett JS, Berger BW, Billings PC. The structure and function of platelet integrins. J Thromb Haemost. 2009;7(Suppl 1):200–205. doi: 10.1111/j.1538-7836.2009.03378.x. [DOI] [PubMed] [Google Scholar]
- 59.Cheresh DA. Human endothelial cells synthesize and express an Arg-Gly-Asp-directed adhesion receptor involved in attachment to fibrinogen and von Willebrand factor. Proc Natl Acad Sci U S A. 1987;84:6471–6475. doi: 10.1073/pnas.84.18.6471. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Pierschbacher MD, Ruoslahti E. Cell attachment activity of fibronectin can be duplicated by small synthetic fragments of the molecule. Nature. 1984;309:30–33. doi: 10.1038/309030a0. [DOI] [PubMed] [Google Scholar]
- 61.Pierschbacher MD, Ruoslahti E. Influence of stereochemistry of the sequence Arg-Gly-Asp-Xaa on binding specificity in cell adhesion. J Biol Chem. 1987;262:17294–17298. [PubMed] [Google Scholar]
- 62.Cheng S, Craig WS, Mullen D, Tschopp JF, Dixon D, Pierschbacher MD. Design and synthesis of novel cyclic RGD-containing peptides as highly potent and selective integrin alpha IIb beta 3 antagonists. J Med Chem. 1994;37:1–8. doi: 10.1021/jm00027a001. [DOI] [PubMed] [Google Scholar]
- 63.Srinivasan R, Marchant RE, Gupta AS. In vitro and in vivo platelet targeting by cyclic RGD-modified liposomes. J Biomed Mater Res A. 2009;93:1004–1015. doi: 10.1002/jbm.a.32549. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Beer JH, Springer KT, Coller BS. Immobilized Arg-Gly-Asp (RGD) peptides of varying lengths as structural probes of the platelet glycoprotein IIb/IIIa receptor. Blood. 1992;79:117–128. [PubMed] [Google Scholar]
- 65.Gu F, Zhang L, Teply Ba, Mann N, Wang A, Radovic-Moreno AF, Langer R, Farokhzad OC. Precise engineering of targeted nanoparticles by using self-assembled biointegrated block copolymers. Proc Natl Acad Sci U S A. 2008;105:2586–91. doi: 10.1073/pnas.0711714105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Garg A, Tisdale AW, Haidari E, Kokkoli E. Targeting colon cancer cells using PEGylated liposomes modified with a fibronectin-mimetic peptide. Int J Pharm. 2009;366:201–10. doi: 10.1016/j.ijpharm.2008.09.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Gindy ME, Ji S, Hoye TR, Panagiotopoulos AZ, Prud’homme RK. Preparation of poly(ethylene glycol) protected nanoparticles with variable bioconjugate ligand density. Biomacromolecules. 2008;9:2705–11. doi: 10.1021/bm8002013. [DOI] [PubMed] [Google Scholar]
- 68.Olivier V, Meisen I, Meckelein B, Hirst TR, Peter-Katalinic J, Schmidt MA, Frey A. Influence of targeting ligand flexibility on receptor binding of particulate drug delivery systems. Bioconjug Chem. 2003;14:1203–8. doi: 10.1021/bc034077z. [DOI] [PubMed] [Google Scholar]
- 69.Roche AM, James MF, Bennett-Guerrero E, Mythen MG. A head-to-head comparison of the in vitro coagulation effects of saline-based and balanced electrolyte crystalloid and colloid intravenous fluids. Anesth Analg. 2006;102:1274–1279. doi: 10.1213/01.ane.0000197694.48429.94. [DOI] [PubMed] [Google Scholar]
- 70.Elias DR, Poloukhtine A, Popik V, Tsourkas A. Effect of ligand density, receptor density, and nanoparticle size on cell targeting. Nanomedicine. 2013;9:194–201. doi: 10.1016/j.nano.2012.05.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Shoffstall AJ, Everhart LM, Varley ME, Soehnlen ES, Shick AM, Ustin JS, Lavik EB. Tuning ligand density on intravenous hemostatic nanoparticles dramatically increases survival following blunt trauma. Biomacromolecules. 2013;14:2790–2797. doi: 10.1021/bm400619v. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Coller BS, Kutok JL, Scudder LE, Galanakis DK, West SM, Rudomen GS, Springer KT. Studies of Activated GPllb / llla Receptors on the Luminal Surface of Adherent Platelets Paradoxical Loss of Luminal Receptors When Platelets Adhere to High Density Fibrinogen. 1993;92:2796–2806. doi: 10.1172/JCI116899. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Grannis GF. Plasma fibrinogen: determination, normal values, physiopathologic shifts, and fluctuations. Clin Chem. 1970;16:486–494. [PubMed] [Google Scholar]
- 74.Lord ST. Fibrinogen and fibrin: scaffold proteins in hemostasis. Curr Opin Hematol. 2007;14:236–241. doi: 10.1097/MOH.0b013e3280dce58c. [DOI] [PubMed] [Google Scholar]
- 75.Blombäck B, Carlsson K, Fatah K, Hessel B, Procyk R. Fibrin in human plasma: Gel architectures governed by rate and nature of fibrinogen activation. Thromb Res. 1994;75:521–538. doi: 10.1016/0049-3848(94)90227-5. [DOI] [PubMed] [Google Scholar]
- 76.Wolberg AS. Thrombin generation and fibrin clot structure. Blood Rev. 2007;21:131–142. doi: 10.1016/j.blre.2006.11.001. [DOI] [PubMed] [Google Scholar]
- 77.Brummel-Ziedins KE, Branda RF, Butenas S, Mann KG. Discordant fibrin formation in hemophilia. J Thromb Haemost. 2009;7:825–832. doi: 10.1111/j.1538-7836.2009.03306.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Lord ST. Molecular mechanisms affecting fibrin structure and stability. Arterioscler Thromb Vasc Biol. 2011;31:494–499. doi: 10.1161/ATVBAHA.110.213389. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Lorand L, Parameswaran KN, Murthy SN. A double-headed Gly-Pro-Arg-Pro ligand mimics the functions of the E domain of fibrin for promoting the end-to-end crosslinking of gamma chains by factor XIIIa. Proc Natl Acad Sci U S A. 1998;95:537–541. doi: 10.1073/pnas.95.2.537. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Laudano AP, Doolittle RF. Synthetic peptide derivatives that bind to fibrinogen and prevent the polymerization of fibrin monomers. Proc Natl Acad Sci U S A. 1978;75:3085–3089. doi: 10.1073/pnas.75.7.3085. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Everse SJ, Spraggon G, Veerapandian L, Riley M, Doolittle RF. Articles Crystal Structure of Fragment Double-D from Human Fibrin with Two Different. Biochemistry. 1998;2960:8637–8642. doi: 10.1021/bi9804129. [DOI] [PubMed] [Google Scholar]
- 82.Soon AS, Lee CS, Barker TH. Modulation of fibrin matrix properties via knob:hole affinity interactions using peptide-PEG conjugates. Biomaterials. 2011;32:4406–4414. doi: 10.1016/j.biomaterials.2011.02.050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Stabenfeldt SE, Aboujamous NM, Soon AS, Barker TH. A new direction for anticoagulants: Inhibiting fibrin assembly with PEGylated fibrin knob mimics. Biotechnol Bioeng. 2011;108:2424–33. doi: 10.1002/bit.23184. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Kolodziej AF, Nair SA, Graham P, McMurry TJ, Ladner RC, Wescott C, Sexton DJ, Caravan P. Fibrin specific peptides derived by phage display: characterization of peptides and conjugates for imaging. Bioconjug Chem. 2012;23:548–556. doi: 10.1021/bc200613e. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Kolodziej AF, Zhang Z, Overoye-Chan K, Jacques V, Caravan P. Peptide optimization and conjugation strategies in the development of molecularly targeted magnetic resonance imaging contrast agents. Methods Mol Biol. 2014;1088:185–211. doi: 10.1007/978-1-62703-673-3_13. [DOI] [PubMed] [Google Scholar]
- 86.Vymazal J, Spuentrup E, Cardenas-Molina G, Wiethoff AJ, Hartmann MG, Caravan P, Parsons EC., Jr Thrombus imaging with fibrin-specific gadolinium-based MR contrast agent EP-2104R: results of a phase II clinical study of feasibility. Invest Radiol. 2009;44:697–704. doi: 10.1097/RLI.0b013e3181b092a7. [DOI] [PubMed] [Google Scholar]
- 87.Overoye-Chan K, Koerner S, Looby RJ, Kolodziej AF, Zech SG, Deng Q, Chasse JM, McMurry TJ, Caravan P. EP-2104R: a fibrin-specific gadolinium-Based MRI contrast agent for detection of thrombus. J Am Chem Soc. 2008;130:6025–6039. doi: 10.1021/ja800834y. [DOI] [PubMed] [Google Scholar]
- 88.Chan LW, Wang X, Wei H, Pozzo LD, White NJ, Pun SH. A Synthetic Fibrin-Crosslinking Polymer for Modulating Clot Properties and Inducing Hemostasis. Sci Transl Med. 2015 doi: 10.1126/scitranslmed.3010383. In press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Brohi K, Singh J, Heron M, Coats T. Acute traumatic coagulopathy. J Trauma. 2003;54:1127–1130. doi: 10.1097/01.TA.0000069184.82147.06. [DOI] [PubMed] [Google Scholar]
- 90.Raza I, Davenport R, Rourke C, Platton S, Manson J, Spoors C, Khan S, De’Ath HD, Allard S, Hart DP, Pasi KJ, Hunt BJ, Stanworth S, MacCallum PK, Brohi K. The incidence and magnitude of fibrinolytic activation in trauma patients. J Thromb Haemost. 2013;11:307–314. doi: 10.1111/jth.12078. [DOI] [PubMed] [Google Scholar]
- 91.Brown AC, Stabenfeldt SE, Ahn B, Hannan RT, Dhada KS, Herman ES, Stefanelli V, Guzzetta N, Alexeev A, Lam WA, Lyon LA, Barker TH. Ultrasoft microgels displaying emergent platelet-like behaviours. Nat Mater. 2014;13:1108–14. doi: 10.1038/nmat4066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Osdoit S, Rosa JP. Fibrin Clot Retraction by Human Platelets Correlates with α IIbβ3 Integrin-dependent Protein Tyrosine Dephosphorylation. J Biol Chem. 2001;276:6703–6710. doi: 10.1074/jbc.M008945200. [DOI] [PubMed] [Google Scholar]
- 93.Teramura Y, Okamura Y, Takeoka S, Tsuchiyama H, Narumi H, Kainoh M, Handa M, Ikeda Y, Tsuchida E. Hemostatic effects of polymerized albumin particles bearing rGPIa/IIa in thrombocytopenic mice. Biochem Biophys Res Commun. 2003;306:256–260. doi: 10.1016/s0006-291x(03)00943-4. [DOI] [PubMed] [Google Scholar]