

Piwi-associated RNAs (piRNAs), a specific class of 24- to 30-nucleotide-long RNAs produced by the Piwi-type of Argonaute proteins have a specific germline function in repressing transposable elements. This repression is thought to involve heterochromatin formation, transcriptional and posttranscriptional silencing1–6. The piRNA pathway has other essential functions in germline stem cell maintenance7 and in maintaining germline DNA integrity8–10. Here, we uncover an unexpected function of the piRNA pathway in the decay of maternal mRNAs and in translational repression in the early embryo. A subset of maternal mRNAs is degraded in the embryo, at the maternal to zygotic transition. In Drosophila, maternal mRNA degradation depends on the RNA binding protein Smaug (Smg) and the deadenylase CCR411–13, as well as the zygotic expression of a microRNA (miRNA) cluster14. Using mRNA encoding the embryonic posterior morphogen Nanos (Nos) as a paradigm to study maternal mRNA decay, we found that CCR4-mediated deadenylation of nos depends on components of the piRNA pathway including piRNAs complementary to a specific region in nos 3′-UTR. Reduced deadenylation when piRNA-induced regulation is impaired, correlates with nos mRNA stabilization and translational derepression in the embryo, resulting in head development defects. Aubergine (Aub), one of the Argonaute proteins in the piRNA pathway, is present in a complex with Smg, CCR4, nos mRNA and piRNAs that target nos 3′-UTR, in the bulk of the embryo. We propose that piRNAs and their associated proteins act together with Smg to recruit the CCR4 deadenylation complex to specific mRNAs, thus promoting their decay. Because piRNAs involved in this regulation are produced from transposable elements, this identifies a direct developmental function for transposable elements in the regulation of gene expression.
In Drosophila embryos, Nos is expressed as a gradient which emanates from the posterior pole and organizes abdominal segmentation15. The majority of nos mRNA is distributed throughout the bulk cytoplasm, translationally repressed16, and subsequently degraded during the first 2–3 hours of development. This repression is essential for head and thorax segmentation16,17. A small amount of nos transcripts, localised at the posterior pole of the embryo, escapes degradation and is actively translated, giving rise to the Nos protein gradient. nos mRNA decay in the bulk cytoplasm depends on the CCR4-NOT deadenylation complex and its recruitment onto nos by Smg. This contributes to translational repression in the bulk of the embryo and is required for embryonic antero-posterior patterning13.
Smg was suggested not to be the only activator of nos mRNA decay during early embryogenesis11,12. Zygotically-expressed miRNAs have been reported to activate maternal mRNA deadenylation in zebrafish embryos18 and decay in Drosophila embryos14. We investigated the potential involvement of other classes of small RNAs in mRNA deadenylation and decay before zygotic expression. Since piRNAs are expressed maternally in the germline and present in early embryos19,20, we analysed the possible role of the piRNA pathway in maternal mRNA deadenylation. Piwi, Aub and Ago3 are specific Argonaute proteins1,3,21,22, Armitage (Armi) and Spindle-E (Spn-E) are RNA helicases, and Squash (Squ) is a nuclease2,10,23,24 involved in piRNA biogenesis and function. Poly(A) test (PAT) assays were performed to measure nos mRNA poly(A) tail length in embryos spanning one hour intervals during the first four hours of embryogenesis. In contrast to the progressive shortening of nos mRNA poly(A) tails observed in wild-type embryos correlating with mRNA decay during this period, nos poly(A) tail shortening was affected in embryos from females mutant for the piRNA pathway (herein referred to as mutant embryos) (Figure 1a, Supplementary Figures 1a, 2, 12). This defect in deadenylation correlated with higher amounts of nos mRNA in mutant embryos, as quantified by RT-QPCR (Figure 1b). in situ hybridization revealed stabilized nos mRNA in the bulk cytoplasm of mutant embryos where it is normally degraded in the wild-type (Figure 1c, Supplementary Figure 1b). Consistent with previous data showing that nos mRNA deadenylation is required for translational repression13, defective deadenylation in mutant embryos resulted in the presence of ectopic Nos protein throughout the embryo (Figure 1d, Supplementary Figure 1c). The presence of Nos in the anterior region results in the repression of bicoid and hunchback mRNA translation and in affected head skeleton. Consistent with earlier mentioned defects7, we found that the piwi1 mutant embryos which were able to produce a cuticle had strong head defects (Figure 1e).
Figure 1.
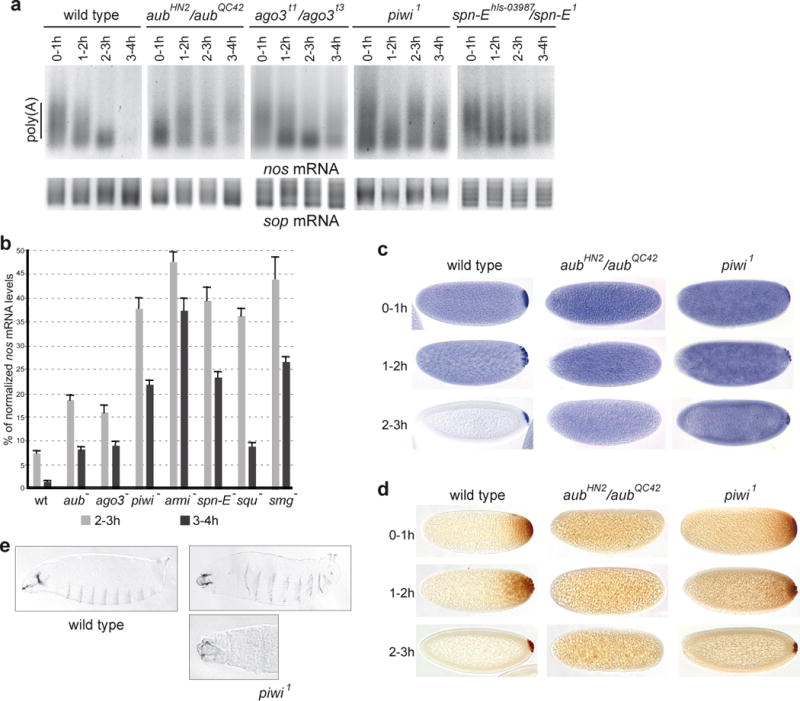
The piRNA pathway is required for nos mRNA deadenylation and decay as well as translational repression in the bulk cytoplasm of the embryo. (a, b) PAT assays and RT-QPCR of nos mRNA. Mutant females of the indicated genotypes were crossed with wild-type males. The sop mRNA was used as a control in a. (b) Levels of nos mRNA in 2–3 hour and 3–4 hour embryos. Mean value of three quantifications, error bars correspond to s.d. (c) In situ hybridizations of nos mRNA. (d) Immunostaining of embryos with anti-Nos antibody. (e) Cuticle preparations of piwi1 embryos showing head defects (rudimentary head skeleton (top panel), head skeleton replaced by a hole (bottom panel)). 2% of embryos from piwi1 germline clones produced a cuticle (n=1060), among those, 22/23 had strong head defects. No embryos from aubN11/aubHN2 (n=1230) or aubQC42/aubHN2 (n=813) females produced a cuticle.
The piRNA pathway plays a role during early oogenesis in preventing DNA damage, possibly through the repression of transposable element transposition. DNA double-strand breaks arising in mutants of the piRNA pathway result in affected embryonic axis specification, and this developmental defect is suppressed by mutations in the Chk2 DNA damage signal transduction pathway9,10. We found that defects in nos mRNA deadenylation and decay observed in aub or armi mutants were not suppressed by Chk2 (mnkP6) mutations, indicating that these defects did not result from activation of the Chk2 pathway earlier during oogenesis (Supplementary Figure 3a–c). Moreover, affected deadenylation of nos mRNA in piRNA pathway mutants did not depend on oskar (Supplementary Figure 3d).
We addressed a potential direct role of the piRNA pathway in the regulation of nos mRNA deadenylation and decay in the embryo. Aub and Piwi accumulate in the pole plasm and in pole cells of the embryo25,26. However, we found lower levels of Aub and Piwi throughout the entire embryo (Figure 2a, Supplementary Figures 4, 5). Ago3 was also present throughout the embryo (Supplementary Figure 6a, c). Aub and Ago3 were cytoplasmic and accumulated in discrete foci, a distribution similar to those of CCR4 and Smg (Figure 2b, Supplementary Figure 6b). CCR4 and Smg were reported to partially colocalise in small cytoplasmic foci13. Aub and Ago3 also partially colocalised with Smg and CCR4 in the bulk of syncytial embryos, in both cytoplasmic foci and a diffusely distributed cytoplasmic pool (Figure 2b, Supplementary Figure 6b). Importantly, the distributions of CCR4 and Smg depended on the piRNA pathway, as they were strongly affected in aub and spn-E mutant embryos. Although global amouts of CCR4 and Smg did not decrease in mutant embryos, CCR4 foci strongly increased in size, whereas Smg foci decreased in size or disappeared (Figure 2c, d). This suggests different functions for a subset of CCR4 and Smg foci and that deadenylation could take place diffusely in the cytoplasm. These results demonstrate a functional link between CCR4-mediated deadenylation and the piRNA pathway.
Figure 2.
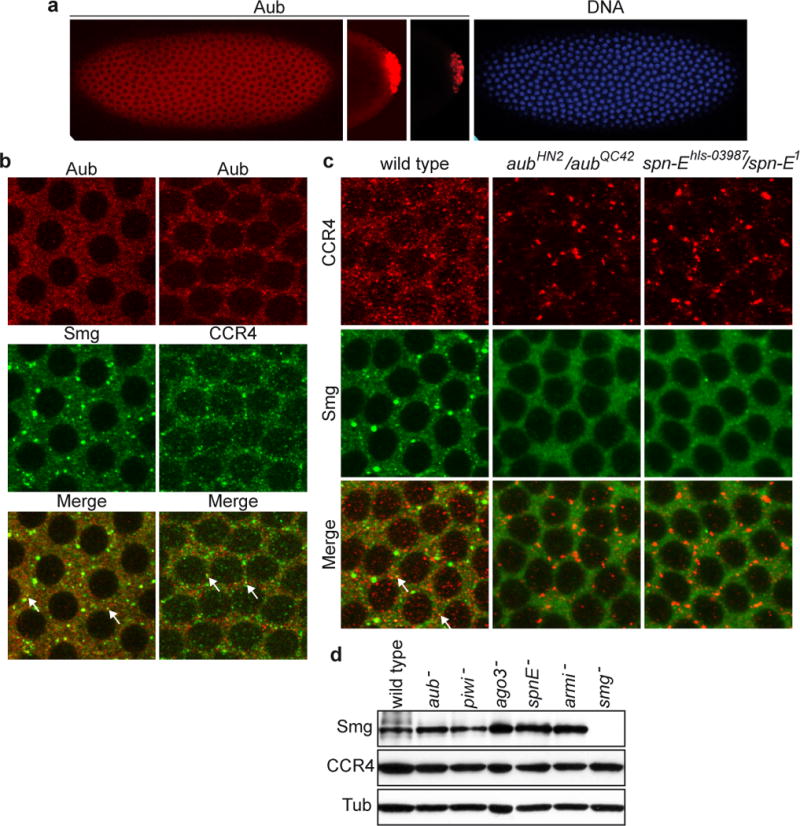
Aub is present in the bulk of the embryo and the piRNA pathway is required for CCR4 and Smg cytoplasmic distributions. (a) Confocal images of cytoplasmic expression of Aub in the embryo. Syncytial blastoderm embryo at nuclear cycle 11, anterior is to the left. Pole cells of the same embryo, at the same setting (middle Aub panel) and at lower intensity (right Aub panel)20,25. DAPI staining (right panel). (b) Double immunostaining of embryos at nuclear cycles 11/12 with anti-Aub and anti-Smg, or anti-Aub and anti-CCR4. Arrows indicate examples of small foci showing colocalisation in b and c. (c) Smg and CCR4 cytoplasmic distributions are affected in aub and spn-E mutant embryos. Double immunostaining of embryos at nuclear cycle 11 with anti-CCR4 and anti-Smg. (d) Western blots of proteins from 0–2 hour embryos revealed with anti-Smg and anti-CCR4. α-tubulin (Tub) was used as a loading control.
Co-immunoprecipitation experiments showed that Aub co-precipitated Smg, CCR4 and Ago3 in the absence of RNA, suggesting the presence of these proteins in a common complex (Figure 3a, Supplementary Figure 7a, b). Smg also co-precipitated CCR4, Aub and Ago3 (Figure 3b, Supplementary Figure 7c). Piwi, however, was not found to co-precipitate Smg or CCR4 (data not shown). Importantly, Smg, CCR4 and Ago3 also co-precipitated with Aub in osk54 mutant embryos that are defective in pole plasm assembly27, indicating the presence of this complex outside the pole plasm (Figure 3a). We next showed that nos mRNA co-precipitated with Aub in both wild-type and osk54 embryos. The amounts of nos mRNA were similar in Aub and Smg immunoprecipitates (Figure 3c).
Figure 3.
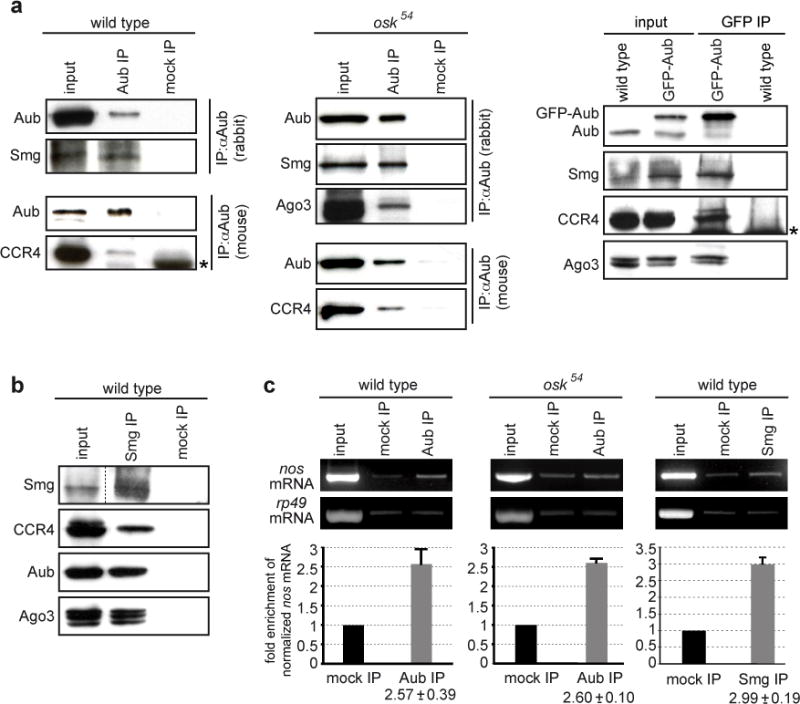
Aub, Ago3, Smg, CCR4 and nos mRNA are present in a common complex in the bulk of the embryo. (a) Co-immunoprecipitations of Smg, CCR4 and Ago3 with Aub in 0–2 hour embryo extracts. Anti-Aub and anti-GFP were used for immunoprecipitations in wild-type, osk54 and GFP-Aub expressing embryos, respectively. (b) Co-immunoprecipitations of CCR4, Aub and Ago3 with Smg in 0–2 hour wild-type embryo extracts. The asterisks indicate immunoglobulins. (c) Quantification of nos mRNA enrichment in Aub and Smg immunoprecipitations. Extracts from 0–2 hour wild-type or osk54 embryos were immunoprecipitated with anti-Aub (rabbit), or anti-Smg. For quantifications performed by RT-QPCR, the ratio of nos mRNA/rp49 mRNA was set to 1 in the mock immunoprecipitation. Mean value of three quantifications, error bars correspond to s.d. rp49 was used as a control mRNA.
These findings show that the Argonaute proteins Aub and Ago3 associate with Smg and the CCR4 deadenylase complex to directly regulate nos mRNA in the bulk cytoplasm of early embryos.
nos 3′-UTR contains Smg binding sites (SRE) located in its 5′-most region (referred to as TCE)16. We searched for piRNAs, sequenced from early embryos, presumed capable of targeting nos 3′-UTR based on their sequence complementarity. Strikingly, a specific region located in the 3′-most part of the 3′-UTR could be targeted by over 200 copies of piRNAs originating from two transposable elements, 412 and roo (Figure 4a, Supplementary Figure 8). piRNAs complementary to nos 3′-UTR were visualized by northern blots. In addition, piRNAs predicted to target nos 3′-UTR co-immunoprecipitated with Aub (Figure 4b). We used nos genomic transgenes deleted for different parts of the 3′-UTR16 to address the requirement of the corresponding regions for nos mRNA deadenylation. We have shown previously that the TCE (nt 1–184) is required for nos mRNA poly(A) tail shortening, consistent with the role of Smg in this process13. Deletion of region 184–403 (nos(Δ1)) had no effect, whereas poly(A) tails from the transgene deleted of the region 403–618 (nos(Δ2)) were elongated in 3–4 hour embryos (Figure 4c, Supplementary Figure 12). This could indicate a regulation by the miRNA predicted by miRBase to target this region. Deletion of 618–844 in nos 3′-UTR (nos(Δ3)) had a strong effect on nos deadenylation (Figure 4c, Supplementary Figures 9, 12). Consistent with this, nos mRNA levels produced by this transgene remained mostly stable (Figure 4d). This resulted in defects in embryo patterning: A total of 35% (n=1894) of embryos from nos(Δ3) females did not hatch and among them 86% (n=28) showed head skeleton defects (Figure 3e). We next deleted specific sequences complementary to 412 (15 nt) and roo (11 nt) retrotransposon piRNAs (Supplementary Figure 8). These short deletions, either independently or in combination, affected nos mRNA deadenylation (Figure 4f, Supplementary Figure 12).
Figure 4.
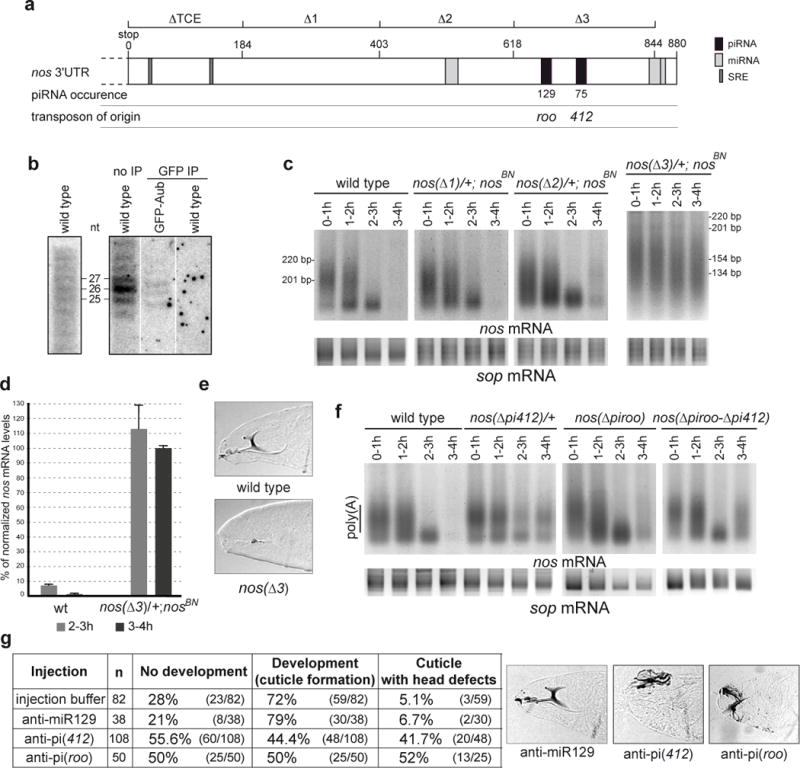
piRNAs target a specific region in nos 3′-UTR which is required for nos mRNA deadenylation. (a) Schematic representation of nos 3′-UTR. The regions deleted in nos genomic transgenes (nos(Δ)) are indicated on the upper line16. SRE, piRNA and miRNA target sites are indicated. Predictions of miRNA targeted regions are from miRBase (miR-31a, miR-314 and miR-263b from proximal to distal). piRNA occurrences in the data sets19,20 are indicated. (b) Northern blots of 0–2 hour embryos probed with riboprobes corresponding to sense nos 3′-UTR (position 403–844) (left panel) and to antisense 412 piRNA (right panel). Anti-GFP immunoprecipitations (GFP IP) were performed using wild-type and GFP-Aub expressing embryos. (c) nos PAT assays. For nos(Δ3) the fragment amplified in the PAT assay is shorter than the fragment amplified in the other nos PAT assays (Supplementary Figure 9). (d) Quantification of nos mRNA levels from the nos(Δ3) transgene by RT-QPCR. Mean value of three quantifications, error bars correspond to s.d. (e) Cuticle preparations of embryos from nos(Δ3) females (lack of head skeleton). (f) PAT assays of embryos with nos genomic transgenes in which sequences complementary to 412 piRNA, roo piRNA, or both sequences have been deleted. The sop mRNA was used as a control in c and f. (g) Injection of 2′ O-methyl anti-piRNA in embryos. Control injections were with injection buffer alone or with the irrelevant anti-miR129. Examples of cuticles following injections of anti-miR129 (wild-type head skeleton), anti-pi(412) and anti-pi(roo) (affected head skeleton).
To further support the role of retrotransposon piRNAs in nos mRNA regulation, we blocked 412 and roo piRNAs by injecting specific 2′ O-methyl anti-piRNA in embryos28, and recorded cuticles as a functional assay of Nos ectopic synthesis at the anterior pole. Injection of anti-piRNA(412) or anti-piRNA(roo) resulted in specific head development defects (Figure 4g).
Together, these results provide strong evidence that an interaction between piRNAs and nos mRNA is required for nos mRNA deadenylation and translational repression in the first hours of embryogenesis.
We have identified a new function of the piRNA pathway in the regulation of maternal mRNAs. Recently, piRNAs derived from 3′-UTR of cellular transcripts have been identified in gonadal somatic cells, although their biological role has not been clarified29,30. Here, we propose that piRNAs, in complex with Piwi-type Argonaute proteins Aub and Ago3, target nos maternal mRNAs and recruit or stabilize the CCR4-NOT deadenylation complex together with Smg (Supplementary Figure 10). These interactions induce rapid mRNA deadenylation and decay. Thus, activation of mRNA deadenylation represents a new direct mechanism of action for the piRNA pathway with an essential developmental function during the first steps of embryogenesis.
Smg is a general factor for mRNA decay during early embryogenesis12. Because Aub and Ago3 are present in a complex with Smg in early embryos, a proportion of Smg mRNA targets could be regulated by the piRNA pathway. Consistent with this, other maternal mRNAs that are destabilized during early embryogenesis, are targeted by abundant piRNAs and their deadenylation depends on the piRNA pathway (Supplementary Figure 11).
These piRNAs involved in gene regulation are generated from transposable element sequences. Although transposable elements have been described to be essential for genome dynamics and evolution, their immediate function within an organism has remained rather elusive. This study provides evidence for a co-evolution between transposable elements and the host genome and reveals the direct developmental function of transposable elements in embryonic patterning, through the regulation of gene expression.
Methods summary
RNA and proteins were manipulated using methods described previously and reported in the Methods section. Embryo injections. Injections of embryos were performed laterally with 400 μM of 2′ O-methyl oligonucleotides as reported previously28. Injection buffer was 0.5 mM NaPO4, 5 mM KCl. Sequences of 2′ O-methyl oligonucleotides are indicated in Methods. Bioinformatics. A total of 29,108,987 piRNAs sequenced from 0–1 hour embryos (GSM286613 and GSM286604 data sets19) and from 0–2 hour embryos (GSM327625, GSM327626, GSM327627, GSM327628 and GSM327629 data sets20) was blasted against nos 3′-UTR using the following parameters: NCBI blast with an E value of 100 and a 14-nt match and WU-blast with an E value of 10 and a 11-nt match. Regions potentially targeted by piRNAs with an occurrence of less than 10 were not considered.
Methods
Drosophila stocks and genetics
The w1118 stock was used as a control. Mutant stocks were w; Sp aubN11 bw/CyO, aubHN2 cn1 bw1/CyO, aubQC42 cn1 bw1/CyO31, mnkP6, mnkP6 aubHN2/CyO, mnkP6 aubQC42/CyO9, mnkP6; armi1/SM6-TM6B, mnkP6; armi72.1/SM6-TM6B9, piwi neo-FRT40A/CyO7, spn-E1/TM3, ry506 spn-Ehls-03987 e/TM332, armi1/TM3, armi72.1/TM323, cn1 bw1 squHE47/CyO, cn1 bw1 squPP32/CyO10, bw; st ago3t1/TM6B Tb, bw; st ago3t3/TM6B Tb22, osk54 spn-E1/TM3, osk54 spn-Ehls-03987 e/TM3. piwi1 mutant embryos are from germline clones that were induced with two 1.5-hour heat shocks at 37°C during the second- and third-instar larval stage, using the FLP recombinase-dominant female sterile technique33. smg mutants were smg1 and a deficiency overlapping smg, Df(ScfR6)17. nosBN mutant does not produce nos mRNA34. osk54 is a null allele. GFP-Aub was expressed following crosses between the germline driver nos-Gal4:VP1635 and UASp-GFP-Aub25. nos(Δ) stocks are transgenic lines containing a nos genomic transgene in which different parts of the 3′UTR have been removed16. nos(Δ) stocks were a gift from R. Wharton. The nos(Δpi412) and nos(Δpiroo) transgenes, in which 15 nt (TATATTTATTCAATT) and 11 nt (AACACACATAT) have been deleted, respectively were generated as follows. The pBSKS-R5561 (containing a 5.7 kb nos genomic fragment, gift from R. Wharton) was used as a template for PCR reactions to produce the deletion. For each construct, two PCR reactions were performed using the following primers: For nos(Δpi412), 5′CATTCCGATCAAAGCTGGGTTAACC (Primer 1) and 5′AAATTGATCAATGGTAAACAATAACATATATATAT which contains the 15 nt deletion, and 5′TATATATATATATATATGTTATTGTTTACCATTGATCAATTT which contains the 15 nt deletion and 5′CTCCACCGCGGTGGCGGCCGC (Primer 2). For nos(Δpiroo), Primer 1 and 5′TATATATATATATATATATAGGAAATGAATACTTGCGATACA which contains the 11 nt deletion, and 5′TGTATCGCAAGTATTCATTTCCTATATATATATATATATATA which contains the 11 nt deletion and Primer 2. For each construct, the two PCR products were annealed and used as a template for a third PCR reaction using Primers 1 and 2. This third PCR product containing either the 15 nt or the 11 nt deletion surrounded by the restriction sites BglII and NotI was cloned into the TAcloning vector (pCRII) (Invitrogen) and sequenced. For the nos(Δpiroo-Δpi412) transgene, the PCR generating the Δpiroo deletion was done using pCRII containing the nosΔpi412 deletion as a template. The BglII-NotI fragment containing the deletion was used to replace the BglII-NotI fragment in the original pBSKS-R5561. An EcoRI-NotI fragment containing the whole genomic fragment with the deletion was cloned into the pCaSpeR4. Transformant stocks were produced by BestGene Inc.
Embryo injections
Sequences of 2′ O-methyl oligonucleotides were as follows. anti-pi(412): UCGGGCUGACAUAUAUUUAUUCAAUU anti-pi(roo): UCCAAACACACAUAUAUAUAUAAAUA anti-miR129-1: GCAAGCCCAGACCGCAAAAAG (human miR129-1 is not conserved in Drosophila).
RNA
PAT assays, RT-PCR and RT-QPCR were performed as described previously13,36, and were made from two to four independent RNA preparations. For nos(Δ3) transgene, a different nos specific primer was used (5′GTCGTCGGCTACGCATTCATTGT), as the region normally amplified in nos PAT assays is deleted in this transgene. We verified by sequencing that the poly(A) site used in mRNA from this transgene is identical to the one used in nos endogenous mRNA. Real-time PCR (QPCR) were performed with the LightCycler System (Roche Molecular Biochemical) using rp49 as a control mRNA13. For quantification of nos mRNA in 2–3 hour and 3–4 hour embryos, the levels were normalized with the levels of nos mRNA in 0–1 hour embryos that were set to 100% for each genotype. Northern blots were performed as described previously37. The sequence of the riboprobe specific to 412 piRNA was 5′GGGCUGACAUAUAUUUAUUCAAUU.
RNA in situ hybridization and cuticle preparations
Whole mount in situ hybridization and cuticle preparations were performed by standard methods. The probe for in situ hybridization was an antisense RNA made from the pN5 nos cDNA clone.
Antibodies, western blots, immunostaining and immunoprecipitations
Immunoprecipitations were performed as described previously13 using 0–2 hour embryos, and mouse anti-Aub (4D1021), mock IPs: mouse anti-HA (12CA5 Developmental Studies Hybridoma Bank, for wild-type embryos) and mouse IgG (sc-2025 Santa Cruz Biotechnology, for osk54 embryos), rabbit anti-Aub (Abcam, ab17724), mock IPs: rabbit IgG (sc-2027 Santa Cruz Biotechnology), mouse anti-GFP (mAb 3E6 Invitrogen), guinea pig anti-Smg (gift from C. Smibert), mock IP: guinea pig pre-immune serum. Protein coimmunoprecipitations were performed in the presence of 0.1 mg/ml RNase A. Western blots and immunostaining were performed as reported38,39. Antibodies for western blots were used at the following dilutions: guinea pig anti-Smg 1/5000, anti-CCR4 1/100040, anti-Piwi 1/20 (P4D21), anti-Aub 1/1500 (4D1021) and anti-Ago3 1/500 (9G321). Antibodies for immunostaining were used at the following dilutions: rabbit anti-Nos 1/1000 (gift from A. Nakamura), guinea pig anti-Smg 1/1000, anti-CCR4 1/300, anti-Aub 1/1500 (4D10), anti-Piwi 1/1 (P4D2) and anti-Ago3 1/300 (9G3).
Supplementary Material
Acknowledgments
We are very grateful to A. Nakamura, M. and H. Siomi, C. Smibert, H. Lin, P. Macdonald, T. Schüpbach, W. Theurkauf, R. Wharton and P. Zamore, for their gifts of antibodies or Drosophila stocks. We thank M. Benkirane for the gift of 2′ O-methyl anti-miR129. This work was supported by the CNRS UPR1142, ANR Blanche (CONTRA), FRM (Equipe FRM 2007) and ARC Libre 2009 to M.S. and by the NIH to E.C.L. C.R., A.C.M. and B.F. held salaries from ANR Blanche.
Footnotes
Supplementary Information is linked to the online version of the paper at www.nature.com/nature.
Author Contributions C.R. and C.P. designed and performed the experiments, analysed the data and contributed equally to the study. A.C.M. contributed to the generation of DNA constructs, B.F. contributed to PAT assays in Fig. 1. A.B., N.R. and E.C.L. performed the bioinformatic analyses. A.P. performed northern blots. M.S. designed the study, analysed data and wrote the paper. All authors discussed the results and commented on the manuscript.
References
- 1.Saito K, et al. Specific association of Piwi with rasiRNAs derived from retrotransposon and heterochromatic regions in the Drosophila genome. Genes Dev. 2006;20:2214–22. doi: 10.1101/gad.1454806. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Vagin VV, et al. A distinct small RNA pathway silences selfish genetic elements in the germline. Science. 2006;313:320–4. doi: 10.1126/science.1129333. [DOI] [PubMed] [Google Scholar]
- 3.Brennecke J, et al. Discrete small RNA-generating loci as master regulators of transposon activity in Drosophila. Cell. 2007;128:1089–103. doi: 10.1016/j.cell.2007.01.043. [DOI] [PubMed] [Google Scholar]
- 4.Yin H, Lin H. An epigenetic activation role of Piwi and a Piwi-associated piRNA in Drosophila melanogaster. Nature. 2007;450:304–8. doi: 10.1038/nature06263. [DOI] [PubMed] [Google Scholar]
- 5.Aravin AA, et al. A piRNA pathway primed by individual transposons is linked to de novo DNA methylation in mice. Mol Cell. 2008;31:785–99. doi: 10.1016/j.molcel.2008.09.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Lim AK, Tao L, Kai T. piRNAs mediate posttranscriptional retroelement silencing and localization to pi-bodies in the Drosophila germline. J Cell Biol. 2009;186:333–42. doi: 10.1083/jcb.200904063. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Cox DN, et al. A novel class of evolutionarily conserved genes defined by piwi are essential for stem cell self-renewal. Genes Dev. 1998;12:3715–27. doi: 10.1101/gad.12.23.3715. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Chen Y, Pane A, Schupbach T. Cutoff and aubergine mutations result in retrotransposon upregulation and checkpoint activation in Drosophila. Curr Biol. 2007;17:637–42. doi: 10.1016/j.cub.2007.02.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Klattenhoff C, et al. Drosophila rasiRNA pathway mutations disrupt embryonic axis specification through activation of an ATR/Chk2 DNA damage response. Dev Cell. 2007;12:45–55. doi: 10.1016/j.devcel.2006.12.001. [DOI] [PubMed] [Google Scholar]
- 10.Pane A, Wehr K, Schupbach T. zucchini and squash encode two putative nucleases required for rasiRNA production in the Drosophila germline. Dev Cell. 2007;12:851–62. doi: 10.1016/j.devcel.2007.03.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Semotok JL, et al. Smaug recruits the CCR4/POP2/NOT deadenylase complex to trigger maternal transcript localization in the early Drosophila embryo. Curr Biol. 2005;15:284–94. doi: 10.1016/j.cub.2005.01.048. [DOI] [PubMed] [Google Scholar]
- 12.Tadros W, et al. SMAUG is a major regulator of maternal mRNA destabilization in Drosophila and its translation is activated by the PAN GU kinase. Dev Cell. 2007;12:143–55. doi: 10.1016/j.devcel.2006.10.005. [DOI] [PubMed] [Google Scholar]
- 13.Zaessinger S, Busseau I, Simonelig M. Oskar allows nanos mRNA translation in Drosophila embryos by preventing its deadenylation by Smaug/CCR4. Development. 2006;133:4573–83. doi: 10.1242/dev.02649. [DOI] [PubMed] [Google Scholar]
- 14.Bushati N, Stark A, Brennecke J, Cohen SM. Temporal reciprocity of miRNAs and their targets during the maternal-to-zygotic transition in Drosophila. Curr Biol. 2008;18:501–6. doi: 10.1016/j.cub.2008.02.081. [DOI] [PubMed] [Google Scholar]
- 15.Gavis ER, Lehmann R. Translational regulation of nanos by RNA localization. Nature. 1994;369:315–8. doi: 10.1038/369315a0. [DOI] [PubMed] [Google Scholar]
- 16.Dahanukar A, Wharton RP. The Nanos gradient in Drosophila embryos is generated by translational regulation. Genes Dev. 1996;10:2610–20. doi: 10.1101/gad.10.20.2610. [DOI] [PubMed] [Google Scholar]
- 17.Dahanukar A, Walker JA, Wharton RP. Smaug, a novel RNA-binding protein that operates a translational switch in Drosophila. Mol Cell. 1999;4:209–18. doi: 10.1016/s1097-2765(00)80368-8. [DOI] [PubMed] [Google Scholar]
- 18.Giraldez AJ, et al. Zebrafish MiR-430 Promotes Deadenylation and Clearance of Maternal mRNAs. Science. 2006 doi: 10.1126/science.1122689. [DOI] [PubMed] [Google Scholar]
- 19.Chung WJ, Okamura K, Martin R, Lai EC. Endogenous RNA interference provides a somatic defense against Drosophila transposons. Curr Biol. 2008;18:795–802. doi: 10.1016/j.cub.2008.05.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Brennecke J, et al. An epigenetic role for maternally inherited piRNAs in transposon silencing. Science. 2008;322:1387–92. doi: 10.1126/science.1165171. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Gunawardane LS, et al. A slicer-mediated mechanism for repeat-associated siRNA 5′ end formation in Drosophila. Science. 2007;315:1587–90. doi: 10.1126/science.1140494. [DOI] [PubMed] [Google Scholar]
- 22.Li C, et al. Collapse of germline piRNAs in the absence of Argonaute3 reveals somatic piRNAs in flies. Cell. 2009;137:509–21. doi: 10.1016/j.cell.2009.04.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Cook HA, Koppetsch BS, Wu J, Theurkauf WE. The Drosophila SDE3 homolog armitage is required for oskar mRNA silencing and embryonic axis specification. Cell. 2004;116:817–29. doi: 10.1016/s0092-8674(04)00250-8. [DOI] [PubMed] [Google Scholar]
- 24.Malone CD, et al. Specialized piRNA pathways act in germline and somatic tissues of the Drosophila ovary. Cell. 2009;137:522–35. doi: 10.1016/j.cell.2009.03.040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Harris AN, Macdonald PM. Aubergine encodes a Drosophila polar granule component required for pole cell formation and related to eIF2C. Development. 2001;128:2823–32. doi: 10.1242/dev.128.14.2823. [DOI] [PubMed] [Google Scholar]
- 26.Megosh HB, Cox DN, Campbell C, Lin H. The role of PIWI and the miRNA machinery in Drosophila germline determination. Curr Biol. 2006;16:1884–94. doi: 10.1016/j.cub.2006.08.051. [DOI] [PubMed] [Google Scholar]
- 27.Lehmann R, Nusslein-Volhard C. The maternal gene nanos has a central role in posterior pattern formation of the Drosophila embryo. Development. 1991;112:679–91. doi: 10.1242/dev.112.3.679. [DOI] [PubMed] [Google Scholar]
- 28.Leaman D, et al. Antisense-mediated depletion reveals essential and specific functions of microRNAs in Drosophila development. Cell. 2005;121:1097–108. doi: 10.1016/j.cell.2005.04.016. [DOI] [PubMed] [Google Scholar]
- 29.Saito K, et al. A regulatory circuit for piwi by the large Maf gene traffic jam in Drosophila. Nature. 2009;461:1296–9. doi: 10.1038/nature08501. [DOI] [PubMed] [Google Scholar]
- 30.Robine N, et al. A broadly conserved pathway generates 3′UTR-directed primary piRNAs. Curr Biol. 2009;19:2066–76. doi: 10.1016/j.cub.2009.11.064. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Schupbach T, Wieschaus E. Female sterile mutations on the second chromosome of Drosophila melanogaster. II. Mutations blocking oogenesis or altering egg morphology. Genetics. 1991;129:1119–36. doi: 10.1093/genetics/129.4.1119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Gillespie DE, Berg CA. Homeless is required for RNA localization in Drosophila oogenesis and encodes a new member of the DE-H family of RNA-dependent ATPases. Genes Dev. 1995;9:2495–508. doi: 10.1101/gad.9.20.2495. [DOI] [PubMed] [Google Scholar]
- 33.Chou TB, Perrimon N. The autosomal FLP-DFS technique for generating germline mosaics in Drosophila melanogaster. Genetics. 1996;144:1673–1679. doi: 10.1093/genetics/144.4.1673. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Wang C, Dickinson LK, Lehmann R. Genetics of nanos localization in Drosophila. Dev Dyn. 1994;199:103–15. doi: 10.1002/aja.1001990204. [DOI] [PubMed] [Google Scholar]
- 35.Rorth P. Gal4 in the Drosophila female germline. Mech Dev. 1998;78:113–118. doi: 10.1016/s0925-4773(98)00157-9. [DOI] [PubMed] [Google Scholar]
- 36.Juge F, Zaessinger S, Temme C, Wahle E, Simonelig M. Control of poly(A) polymerase level is essential to cytoplasmic polyadenylation and early development in Drosophila. EMBO J. 2002;21:6603–6613. doi: 10.1093/emboj/cdf633. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Pelisson A, Sarot E, Payen-Groschene G, Bucheton A. A novel repeat-associated small interfering RNA-mediated silencing pathway downregulates complementary sense gypsy transcripts in somatic cells of the Drosophila ovary. J Virol. 2007;81:1951–60. doi: 10.1128/JVI.01980-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Benoit B, et al. An essential cytoplasmic function for the nuclear poly(A) binding protein, PABP2, in poly(A) tail length control and early development in Drosophila. Dev Cell. 2005;9:511–522. doi: 10.1016/j.devcel.2005.09.002. [DOI] [PubMed] [Google Scholar]
- 39.Benoit B, et al. The Drosophila poly(A)-binding protein II is ubiquitous throughout Drosophila development and has the same function in mRNA polyadenylation as its bovine homolog in vitro. Nucleic Acids Res. 1999;27:3771–3778. doi: 10.1093/nar/27.19.3771. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Temme C, Zaessinger S, Meyer S, Simonelig M, Wahle E. A complex containing the CCR4 and CAF1 proteins is involved in mRNA deadenylation in Drosophila. EMBO J. 2004;23:2862–2871. doi: 10.1038/sj.emboj.7600273. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.