

Abstract
Since 1950, links between intake of saturated fatty acids and heart disease have led to recommendations to limit consumption of saturated fatty acid-rich foods, including beef. Over this time, changes in food consumption patterns in several countries including Canada and the USA have not led to improvements in health. Instead, the incidence of obesity, type II diabetes and associated diseases have reached epidemic proportions owing in part to replacement of dietary fat with refined carbohydrates. Despite the content of saturated fatty acids in beef, it is also rich in heart healthy cis-monounsaturated fatty acids, and can be an important source of long-chain omega-3 (n-3) fatty acids in populations where little or no oily fish is consumed. Beef also contains polyunsaturated fatty acid biohydrogenation products, including vaccenic and rumenic acids, which have been shown to have anticarcinogenic and hypolipidemic properties in cell culture and animal models. Beef can be enriched with these beneficial fatty acids through manipulation of beef cattle diets, which is now more important than ever because of increasing public understanding of the relationships between diet and health. The present review examines recommendations for beef in human diets, the need to recognize the complex nature of beef fat, how cattle diets and management can alter the fatty acid composition of beef, and to what extent content claims are currently possible for beef fatty acids.
Keywords: Beef, Biohydrogenation products, Lipids, n-3 fatty acids, Rumenic acid, Vaccenic acid
Introduction
Quality and price are key factors considered when consumers purchase beef, and a growing segment of medium- to high-income informed consumers now consider the health implications of beef consumption [1, 2]. The present review will cover recent challenges to long-standing recommendations for beef consumption, the content and composition of beef fat, how beef fat composition can be modified through cattle nutrition and practical considerations when beef with enhanced fatty acid profiles reaches consumers’ plates. The overall objective of the review is to provide some insight into how beef and its constituent fatty acids may now, and in the future, fit into the human diet.
Revisiting recommendations for beef consumption
Diet effects on human health are often related to several diseases associated with dietary fat, many of which take years to develop, and often result in changes in quality of life and lifespan. Many developed countries suffer from high incidences of obesity, type II diabetes, coronary heart disease (CHD) and cancer. Efforts to examine associated dietary factors, and make recommendations to improve health, have at times fallen short. Recommendations to decrease consumption have been targeted at foods that contain nutrients singled out as culprits, and changes in dietary patterns have in some cases led to even more difficulties. One of the best examples is the recommendation to substitute foods containing saturated fatty acids (SFA) with trans fatty acid-rich margarines and refined carbohydrates [3]. Years of recommendations to reduce red meat consumption have not been met by dramatic reductions in the incidence of diseases related to dietary fat; on the contrary, the incidence of obesity and type II diabetes has reached epidemic proportions and has been related to refined carbohydrate consumption [4]. Fatty acids singled out in the Nurses’ Health Study as being problematic for CHD are SFA with chain lengths from 14:0 to 18:0, and a stronger association was found when the polyunsaturated fatty acid (PUFA) to SFA ratio was reduced [5].
The current recommendations to reduce SFA intake are based on the findings from studies in mid-20th century that dietary SFA cause an increase in serum total and LDL-cholesterol and therefore increase the risk of heart disease [6]. These earlier studies overlooked other contributing factors as well as the fact that SFA also increase HDL-cholesterol, which is protective against heart disease. Later studies found that the ratio of total serum cholesterol to HDL-cholesterol is a better indicator of heart disease risk than total or LDL-cholesterol [7]. More recently, many studies have started to question the current dietary recommendations against consuming SFA and revealed that SFA intake is not associated with an increased risk of cardiovascular disease [8–10]. In contrast, substitutions of dietary SFA with refined carbohydrates have resulted in increased obesity and worsen blood lipid profiles by increasing serum triacylglycerol and small, dense LDL particles [11, 12]. Reevaluations are required for the existing dietary recommendations which overstate the health risks of SFA and promote their replacement with alternative nutrients such as refined carbohydrates.
Recently, the dogma that meat consumption should be limited in human diets because of its fatty acid composition has come under close scrutiny [13]. In a recent meta-analysis reviewing 20 studies with more than 1 million subjects, Micha et al. [14] found that consumption of red meats was not associated with higher incidence of CHD and type II diabetes, whereas processed meats were associated with increased incidence of both diseases. The authors suggested that other ingredients (e.g., preservatives such as nitrate) used in processed meats, rather than SFA, contributed to the negative disease outcomes. In Europe, current evidence suggests unprocessed lean red meat is safe to consume as a healthy food choice, and recommendations to limit its consumption in substitution for other protein sources including white meat are not justified [15]. In contrast, in the USA, consumption of both unprocessed and processed red meat still reveal associations with disease outcomes, with a greater hazard ratio for unprocessed red meat [16]. However, not all beef is consumed as unprocessed lean beef. In fact, the most consumed beef product in the USA is hamburger [17] which typically contains 10 to 30% fat. Consequently, it would be prudent to shift research focus away from what to do about the SFA in beef towards how beef fat can be used as a vehicle to deliver health-enhancing fatty acids to consumers.
The Global Burden of Disease Study [18] estimated the contribution of risk factors to disease and disability and identified, among other things, that a low intake of omega-3 (n-3) fatty acids is a concern. The high ratio of omega-6 (n-6) to n-3 fatty acids promotes many diseases from cardiovascular disease and arthritis to cancer, whereas lower ratios have suppressive effects [19]. The n-6 to n-3 ratio of diets during human evolution was estimated to be close to 1:1, whereas current Western diets have ratios close to 15:1 [19]. The great amounts n-6 PUFA in the diet promotes the production of eicosanoids (i.e., prostaglandins, thromboxanes, leukotrienes) formed from arachidonic acid (AA) at the expense of those formed from n-3 fatty acids, specifically eicosapentaenoic acid (EPA) [20]. The disproportionate increase in eicosanoids from AA could result in allergic and inflammatory responses such as increase in platelet aggregation, blood viscosity, vasospasm and vasoconstriction as well as reduced bleeding time [21]. Furthermore, an increased n-6 to n-3 ratio could promote or exacerbate atherogenesis [10]. The balance of n-6 to n-3 fatty acids is therefore an important determinant in reducing the risk of inflammatory and autoimmune disorders such as diabetes, CHD, hypertension, diabetes and arthritis.
In China, the n-6 to n-3 fatty acid ratios of red meat were recently found to range from 6/1 to 23/1 [22]. Unless protected from rumen biohydrogenation, beef naturally contains a low content of n-3 fatty acids including α-linolenic acid (ALA; 18:3n-3) and its long-chain (LC) elongation and desaturation products EPA, docoasapentaenenoic acid (DPA) and docosahexaenoic acid (DHA) [23]. The health benefits ascribed to n-3 fatty acids are mostly related to the LC n-3 s typically found at higher concentrations in fish oil (i.e., EPA and DHA), and efforts have been made to establish dietary reference intakes for these [24]. In contrast, the most common LC n-3 fatty acid in beef is DPA, but it can be readily converted to EPA and DHA [25], and should thus be included when calculating LC n-3 s. Consequently, in populations where little or no oily fish is consumed, beef can still be an important source of LC n-3 fatty acids, particularly when DPA is included [26]. The fact that beef fat can be a source of LC n-3 fatty acids is positive, but again when considering health implications of beef fat, it is important not to narrow the scope of consideration to a few individual or related groups of fatty acids. The complexity of beef fat, and that its effects on human health stems both from individual fatty acids and their combined effects, is under appreciated. Consequently beef producers wanting to improve the health profile of beef require information on which fatty acids would be of interest, and how these can be practically and profitably manipulated by diet to reach levels required to be of benefit to consumers.
Beef fat content and composition
Beef and meat from other ruminant species are noted for having complex fatty acid profiles compared to meat from monogastric species. Paradoxically, using diet to modify meat composition is much easier in monogastric than ruminant species. Rumen microbes are responsible for both the complexity of beef fatty acid composition and for its lack of resemblance to dietary fatty acid profiles [27]. Rumen microbes produce branched- and odd-chain fatty acids and their precursors, resulting in their deposition in beef lipids. In addition, rumen microbes produce several PUFA biohydrogenation products (PUFA-BHP) including conjugated trienes, conjugated dienes, non-conjugated dienes and monounsaturated fatty acids (MUFA) with a vast array of double bond locations and cis/trans configurations. Cattle diets typically contain 1-4% lipids, which mainly consist of PUFA including linoleic acid (LA, 18:2n-6) and ALA. When cattle consume feed, dietary lipids are acted upon by microbial lipases in the rumen, releasing mainly free PUFA, which are toxic to rumen microbes [28]. To cope, rumen microbes biohydrogenate PUFA to less toxic SFA, particularly to 18:0, and this process is typically very efficient. Residual PUFA-BHP bypassing the rumen can then be absorbed from the lower gut and incorporated into beef. In a survey of Canadian retail beef (longissimus lumborum from strip loin steaks) conducted by Aldai et al. [29], the three most concentrated fatty acids were cis9-18:1, 16:0 and 18:0 with concentrations of 38%, 24% and 12%, respectively, constituting 74% of total fatty acids (Table 1). The next eight most concentrated fatty acids (1 to 5% of total fatty acids) accounted for 15.2% of total fatty acids. The next 16 most concentrated fatty acids (0.2 to 1% of total fatty acids) contributed 6.4% to total fatty acids, and the final 60 fatty acids (0.0–0.1% of total fatty acids) accounted for 4.4% of total fatty acids with the majority being PUFA-BHP. Beef analyzed in this survey was collected at retail, and in all likelihood would have been from cattle fed barley grain-based diets (75–90% of dry matter).
Table 1.
Rank order of fatty acids in Canadian retail strip loin steak (longissiums lumborum)1
| Rank | Group2 | Common Name3 | Fatty Acid4 | Weight, % | SD5 |
|---|---|---|---|---|---|
| 1 | MUFA | Oleic | c9-18:1 | 38.08 | 2.699 |
| 2 | SFA | Palmitic | 16:0 | 23.95 | 1.381 |
| 3 | SFA | Stearic | 18:0 | 12.03 | 1.667 |
| 4 | MUFA | Palmitoleic | c9-16:1 | 3.592 | 0.709 |
| 5 | PUFA | Linoleic | 18:2n-6 | 2.708 | 0.786 |
| 6 | SFA | Myrsitic | 14:0 | 2.289 | 0.392 |
| 7 | MUFA | Asclepic | c11-18:1 | 1.847 | 0.213 |
| 8 | SFA | Margaric | 17:0 | 1.309 | 0.298 |
| 9 | MUFA | Margarolic | c9-17:1 | 1.302 | 0.298 |
| 10 | MUFA | t10-18:1 | 1.100 | 0.688 | |
| 11 | DMA | 16:0DMA | 1.017 | 0.422 | |
| 12 | PUFA | Arachidonic | 20:4n-6 | 0.809 | 0.349 |
| 13 | MUFA | Myristoleic | c9-14:1 | 0.589 | 0.202 |
| 14 | DMA | 18:0DMA | 0.557 | 0.222 | |
| 15 | BCFA/SFA | anteiso17:0 | 0.504 | 0.084 | |
| 16 | MUFA | Vaccenic | t11-18:1 | 0.470 | 0.164 |
| 17 | MUFA | c13-18:1 | 0.452 | 0.121 | |
| 18 | SFA | 15:0 | 0.448 | 0.103 | |
| 19 | BCFA/SFA | iso17:0 | 0.325 | 0.053 | |
| 20 | PUFA | ALA | 18:3n-3 | 0.277 | 0.090 |
| 21 | PUFA | DPA | 22:5n-3 | 0.274 | 0.099 |
| 22 | CLA/PUFA | Rumenic | c9,t11-18:2 | 0.273 | 0.134 |
| 23 | MUFA | t13-t14-/c6-c8-18:1 | 0.263 | 0.111 | |
| 24 | PUFA | 20:3n-6 | 0.252 | 0.082 | |
| 25 | MUFA | Elaidic | t9-18:1 | 0.230 | 0.069 |
| 26 | MUFA | t15-18:1 | 0.208 | 0.051 | |
| 27 | MUFA | Gondoic | c11-20:1 | 0.208 | 0.061 |
| 28 | MUFA | c11-16:1 | 0.189 | 0.060 | |
| 29 | PUFA | c9t13-/t8c12-18:2 | 0.179 | 0.039 | |
| 30 | PUFA | c9c15-18:2 | 0.167 | 0.034 | |
| 31 | MUFA | c7-16:1 | 0.156 | 0.020 | |
| 32 | MUFA | t6-t8-18:1 | 0.156 | 0.078 | |
| 33 | MUFA | c15-18:1 | 0.154 | 0.055 | |
| 34 | BCFA | iso16:0 | 0.136 | 0.035 | |
| 35 | MUFA | c12-18:1 | 0.122 | 0.052 | |
| 36 | BCFA/SFA | anteiso15:0 | 0.121 | 0.036 | |
| 37 | MUFA | t12-18:1 | 0.118 | 0.045 | |
| 38 | PUFA | Adrenic | 22:4n-6 | 0.114 | 0.024 |
| 39 | BCFA/SFA | iso18:0 | 0.113 | 0.047 | |
| 40 | PUFA | EPA | 20:5n-3 | 0.110 | 0.063 |
| 41 | PUFA | t11c15-18:2 | 0.101 | 0.052 | |
| 42 | PUFA | t8c13-18:2 | 0.092 | 0.021 | |
| 43 | MUFA | t16-18:1 | 0.088 | 0.033 | |
| 44 | SFA | 11-cyclohexyl-17:0 | 0.088 | 0.027 | |
| 45 | MUFA | c13-16:1 | 0.085 | 0.024 | |
| 46 | MUFA | Gondoleic | c9-20:1 | 0.084 | 0.018 |
| 47 | BCFA/SFA | iso15:0 | 0.084 | 0.028 | |
| 48 | SFA | Arachidic | 20:0 | 0.080 | 0.021 |
| 49 | SFA | 19:0 | 0.077 | 0.030 | |
| 50 | MUFA | c9-15:1 | 0.076 | 0.026 | |
| 51 | PUFA+MUFA | c9t12-18:2/c16-18:1 | 0.062 | 0.016 | |
| 52 | SFA | Behenic | 22:0 | 0.062 | 0.018 |
| 53 | CLA/PUFA | Yurawic | t7,c9-18:2 | 0.061 | 0.027 |
| 54 | PUFA | 20:2n-6 | 0.051 | 0.013 | |
| 55 | PUFA | Rumelenic | c9t11c15-18:3 | 0.049 | 0.021 |
| 56 | DMA | 18:1DMA | 0.047 | 0.021 | |
| 57 | MUFA | c11-17:1 | 0.043 | 0.016 | |
| 58 | DMA | 16:1DMA | 0.043 | 0.015 | |
| 59 | MUFA | t11/t12-16:1 | 0.043 | 0.008 | |
| 60 | CLA/PUFA | t9c11-18:2 | 0.041 | 0.016 | |
| 61 | CLA/PUFA | Linelaidic | t9t12-18:2 | 0.040 | 0.019 |
| 62 | MUFA | c10-16:1 | 0.039 | 0.011 | |
| 63 | PUFA/MUFA | 20:3n-3/c13-22:1 | 0.037 | 0.020 | |
| 64 | MUFA | c14-18:1 | 0.037 | 0.011 | |
| 65 | MUFA | c5-17:1 | 0.035 | 0.009 | |
| 66 | MUFA | c7-17:1 | 0.034 | 0.009 | |
| 67 | PUFA | t9c12-18:2 | 0.032 | 0.014 | |
| 68 | PUFA | GLA | 18:3n-6 | 0.032 | 0.013 |
| 69 | SFA | Lauric | 12:0 | 0.032 | 0.011 |
| 70 | PUFA | DHA | 22:6n-3 | 0.031 | 0.018 |
| 71 | SFA | Lignoceric | 24:0 | 0.030 | 0.014 |
| 72 | MUFA | t5-18:1 | 0.027 | 0.016 | |
| 73 | MUFA | c12-16:1 | 0.026 | 0.008 | |
| 74 | SFA | 21:0 | 0.023 | 0.006 | |
| 75 | CLA/PUFA | t10,c12-18:2 | 0.011 | 0.006 | |
| 76 | CLA/PUFA | c9,c11-18:2 | 0.011 | 0.003 | |
| 77 | CLA/PUFA | t8,c10-18:2 | 0.011 | 0.003 | |
| 78 | CLA/PUFA | c11,t13-18:2 | 0.009 | 0.003 | |
| 79 | CLA/PUFA | Mangold's | t9,t11-18:2 | 0.009 | 0.003 |
| 80 | CLA/PUFA | t11,c13-18:2 | 0.008 | 0.006 | |
| 81 | CLA/PUFA | t11,t13-18:2 | 0.005 | 0.002 | |
| 82 | CLA/PUFA | Mikusch's | t10,t12-18:2 | 0.004 | 0.002 |
| 83 | CLA/PUFA | c12,t14-18:2 | 0.004 | 0.002 | |
| 84 | CLA/PUFA | t12,c14-18:2 | 0.004 | 0.003 | |
| 85 | CLA/PUFA | t7,t9-18:2 | 0.003 | 0.001 | |
| 86 | CLA/PUFA | t12,t14-18:2 | 0.003 | 0.001 | |
| 87 | CLA/PUFA | t8,t10-18:2 | 0.002 | 0.001 |
130 steaks collected in summer and 30 in winter [29]
2 SFA saturated fatty acids, MUFA monounsaturated fatty acids, BCFA branched-chain fatty acids, PUFA polyunsaturated fatty acids, DMA dimethyl acetal, CLA conjugated linoleic acid
3 ALA α-linolenic acid, DPA docosapentaenoic acid, EPA eicosapentaenoic acid, GLA γ-linolenic acid, DHA docosahexaenoic acid
4 c cis, t trans. Co-eluting fatty acids on gas chromatogram are separated by a slash (/)
5 SD standard deviation
Modifying the fat content and composition of beef has been the subject of several reviews, and, in summary, the amount of fat in beef and its composition can be modified primarily by diet and to a lesser extent by gender and genetics [30–34]. Feeding high-grain diets to cattle leads to fatter carcasses and deposition of intramuscular fat (i.e., marbling), a valued attribute in several markets including Japan, the USA and Canada. In contrast to high-grain diets, reducing dietary energy content, through feeding high-forage diets, reduces carcass fatness, decreases intramuscular fat and increases the proportion of PUFA rich phospholipids relative to SFA rich neutral lipids [35]. Feeding high-forage diets can also lead to what is perceived to be a more healthful beef fatty acid profile, but the trade-off with lower energy diets is increased time to market, and the need to be able to source pasture or conserved forage. In addition, increased proportions of forage in the diet can lead to changes in beef palatability [36] such as decreased tenderness because of increased age at finished weights, and the beef may not be as marketable to consumers that value marbling. In countries like Canada and the USA where feedlot finishing on high-grain diets is the norm, finishing on forage-based diets is limited to a small but growing segment of the market [36]. Future expansion of this market will likely depend on whether fatty acid-associated impacts on human health can be scientifically substantiated.
The quantity and composition of PUFA-BHP in beef is very much dependent on the supply of PUFA in the diet, and associated dietary and animal factors (e.g., feeding behavior and rumen conditions) which influence the degree of biohydrogenation [37]. In general, pathways used for biohydrogenation of LA and ALA, the major fatty acids in typical cattle diet, are influenced by the forage to concentrate ratio [38]. The most highly characterized pathways for LA and ALA biohydrogenation were elucidated when greater proportions of forage versus concentrate were fed (Fig. 1). Pathways for both LA and ALA are characterized by initial isomerization of the cis double bond at carbon 12 to a trans double bond at carbon 11 resulting in the production of RA and cis9,trans11,cis15-18:3, respectively [39, 28]. In contrast, when feeding diets with increased amounts of readily fermentable carbohydrate (i.e., high-grain diets), isomerization of the cis 9 double bond for LA shifts towards a trans double bond at carbon 10 [28], while isomerization of the cis 12 double bond of ALA shifts towards a trans double bond at carbon 13, resulting in the production of trans10, cis12-18:2 and cis9, trans13, cis15-18:3, respectively [40]. Following this are rounds of hydrogenation and isomerization leading to trans 18:1 isomers (e.g., VA and trans13-18:1) and eventually complete hydrogenation to 18:0. However, pathways for the formation of many BHP found in Table 1 have not been established. In addition, new BHP continue to be found. For example, recently trans10, cis15-18:2 was found to be a BHP of ALA [41], adding one more piece to the puzzle of ALA biohydrogenation pathways. In addition, a great number of BHP of longer chain more highly unsaturated PUFA (e.g., DHA) have also been recently characterized [42].
Fig. 1.
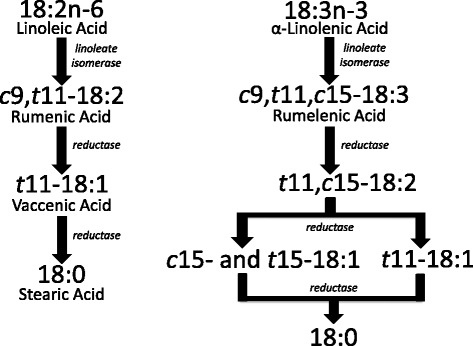
Major pathways for the biohydrogenation of linoleic and α-linolenic acids in the rumen showing isomerization and hydrogenation. Adapted from Harfoot and Hazlewood [39]
The fatty acid composition of beef is complex, but concentrations of many fatty acids can be extremely low. Interestingly, the fatty acids in low concentration including many PUFA-BHP have become of interest because of the finding that some can have potent biological activity. The BHP that have been studied the most are VA and RA, which have been shown to have anticarcinogenic and hypolipidemic properties in cell culture and animal models [43–46]. Still, the effects of many PUFA-BHP have not been studied and finding ways to consistently and meaningfully influence their concentrations is of considerable interest.
Fatty acids of interest and their manipulation by diet
As the effects of fat on ill-health have in part been associated with SFA, logically fat with increased contents of unsaturated fatty acids (UFA), at the expense of SFA, may be more healthful for people to consume. Interestingly, feeding ruminants diets rich in grain are not always associated with greater contents of SFA in meat. In fact, the longer cattle are finished on grain, the greater the delta-9 desaturase activity and conversion of 18:0 to cis9-18:1 in beef [47]. Feeding grain-based diets is, however, also associated with increased trans10-18:1 deposition [48, 49], and consuming trans10-18:1 enriched fats may result in undesirable shifts in plasma cholesterol profiles [50, 51]. Consequently, it would be of importance to determine if the health value of beef enriched withcis9-18:1 is still maintained when different proportions of trans10-18:1 are present.
Cattle feeding practices most frequently associated with increased proportions of PUFA in beef, particularly n-3 fatty acids, are grazing or feeding preserved forages [36, 52]. From a human nutrition perspective, grazing or feeding cattle forages compared to concentrate is appealing as it reduces the fat content of beef and provides several potential improvements in beef fatty acid composition. Forage finishing can increase the percentage of n-3 fatty acids [53], reduce the n-6 to n-3 ratio, reduce the SFA/PUFA ratio, and increase the percentages of specific PUFA-BHP such as VA and RA [23]. These changes in fatty acid composition may exert protective effects against a number of diseases ranging from cancer to cardiovascular disease [33, 37, 45]. It is thus important to consider if improvements in beef fatty acid composition when including more forage in cattle diets, has any human health benefits over and above those related to reductions in total fat content. In addition, it should be determined if human health benefits are consistent when consuming steak (<10% fat) versus ground beef (10–30% fat). Humans consuming red meat (beef and lamb) from grass or concentrate finished animals were found to have no differences in serum lipids, lipoproteins, triacylglycerols or blood pressure [54]. Interestingly, grass fed beef and lamb were able to increase consumer plasma and platelet LC n-3 PUFA status, leading authors to conclude red meat from grass-fed animals may contribute to dietary intakes of LC n-3 PUFA in populations where red meat is habitually consumed. On the other hand, Wagyu steers finished for an extended period on corn grain versus pasture yielded hamburgers enriched with MUFA and SFA, respectively. Consuming SFA-rich hamburgers decreased serum high-density lipoprotein cholesterol (HDL or “good cholesterol”) in mildly hypercholesterolaemic men [55]. Consuming SFA-rich hamburgers did not, however, change serum low-density lipoprotein (LDL) cholesterol, but did reduce LDL particle diameter, and increased triacylglycerols. Consequently, in the future, it will be important to place these and other results into context when considering which beef or beef products to consume, as these may have differing effects on human health, even when coming from the same animal. For example, as it can be interpreted from studies cited above, lean beef from grass-fed cattle may have a fatty acid profile associated with positive effects on human health, but regular hamburger produced from the same beef may be less favorable in terms of the MUFA/SFA ratio.
Beyond strategies to increase amounts of UFA in beef by feeding forage, a more direct possibility can be through supplementing diets with PUFA rich oils or oilseeds. Nevertheless, this dietary strategy is not without difficulty because of the high efficiency of microbial biohydrogenation of PUFA in the rumen, and the influence of diet on routes of biohydrogenation. Supplementing PUFA in cattle diets has, therefore, frequently led to only minor changes in the PUFA or PUFA-BHP content of beef. For example, Gonzalez et al. [56] found very limited accumulation of PUFA or PUFA-BHP in beef when adding 4.5% sunflower, linseed or soybean oil to a concentrate-based diet, and concluded finding ways to protect PUFA from ruminal biohydrogenation would be an important step to increase the PUFA content of beef. Trying to protect PUFA through feed processing [57] or by chemical treatment (e.g., feeding calcium salts or amides of fatty acid) have met with limited success [58, 59]. Significant ruminal bypass of PUFA has been achieved by Scott and co-workers in Australia when using formaldehyde-treated casein to encapsulate oils [60], and more recently this has been extended to include long-chain n-3 fatty acids [61]. The higher content of PUFA can, however, lead to changes in beef sensory characteristics [62], but oxidative stability has been in part offset by vitamin E supplementation [61]. Encapsulating oils has been noted to be expensive, but the process has also been applied to oilseeds to reduce costs [60]. Encapsulation is certainly an area that could benefit from additional investigation, particularly for n-3 fatty acid rich oilseeds such as flaxseed.
Feeding PUFA rich oils or oilseeds in combination with forage versus concentrate-based diets can also have differing effects on the fatty acid composition of beef. Labrune et al. [63] found increased contents of ALA in beef when feeding flaxseed together with corn, which may have been related to effects of low pH on ruminal lipolysis, the first committed step leading to biohydrogenation [64]. In contrast, when Aldai et al. [65, 66] fed 3% soybean oil in a barley grain-based diet with barley straw as the forage source, there was a preferential accumulation of trans10-18:1 in beef at the expense of VA and RA. Supplementing grazing heifers with concentrate fortified with vegetable oils (sunflower or linseed oil) led to remarkable increases in VA and RA in lean beef and adipose tissue [67], but no appreciable increases in VA or RA were found by Kronberg et al. [68] when supplementing steers with flaxseed on pasture. Feeding flaxseed or sunflower seed with either grass-hay or red clover silage-based diets did, however, result in accumulations of VA and RA [69]. In addition, feeding flaxseed supplemented diets resulted in accumulation of BHP specific to ALA, notably trans 13/14-18:1, trans11,cis15-18:2, trans11,cis13-18:2 and cis9,trans11,cis15-18:3. Accumulations of BHP specific to ALA were reduced when feeding flaxseed together with barley silage compared to grass hay [70].
The quantity and type of forage in cattle diets can be keys to increasing BHP with potential influences on human health. Forage-based diets can promote rumen conditions conducive to VA and RA synthesis. Furthermore, they appear to influence the final step in PUFA biohydrogenation to 18:0, resulting in a differences in BHP outflow from the rumen. In addition to adding forage to the diet, there are some indications that the final step in PUFA biohydrogenation to 18:0 can also be influenced by other means. Long-chain n-3 fatty acids found in fish oil or marine microalgae can inhibit the final step in PUFA hydrogenation to 18:0 [71], but the effects may be variable depending on the composition of the basal diet [72, 73]. In addition, some plant secondary metabolites such as tannins [74], saponins [75] and polyphenol oxidase products [76, 77] have potential to interfere with the final step of ruminal biohydrogenation. In the future, there may also be opportunities to influence ruminal biohydrogenation using direct fed microbials, as several bacterial species with biohydrogenation activity have been indentified [78] and several others have recently been associated with deposition of high and low levels of VA in adipose tissue when feeding steers either flaxseed or sunflower seed [79].
Genetic and metabolic influences on beef fatty acid composition
As previously mentioned, the amount of intramuscular fat influences the fatty acid composition of beef because of increases in SFA deposition as total fat increases [35]. Beyond this, fatty acid composition has been noted to have low to moderate heritability [47, 80, 81, 30], but efforts to use genetic selection to improve beef fatty acid composition have been limited for a number of reasons [30]. First, fatty acid composition is not a single trait and it is not clear at present the type or number of fatty acids or their derived parameters that should be included as criteria in a breeding program. Second, if the PUFA/SFA ratio is one criterion for selection, the favorable correlation with reduced fatness suggests that an improved PUFA/SFA ratio can probably be more easily obtained by selecting for lower fatness versus direct selection for individual fatty acids. Third, measuring fatty acid composition on a large number of animals for breeding value estimation would be expensive using conventional techniques (i.e., gas chromatography). Beyond conventional breeding strategies, however, recent developments in genomic technologies have provided opportunities for marker-assisted selection. Single nucleotide polymorphisms (SNPs) have been found for a number of candidate genes involved in fatty acid metabolism [82–85]. A 54 k single nucleotide polymorphism (SNP) chip has also now been used to investigate possibilities for marker-assisted selection of multiple traits from basic meat quality to nutritional composition including mineral and fatty acid composition [86]. Greater advances in the area may thus be on offer with >54 k chips, and with this, the potential for finding quantitative trait loci (QTL) and identify specific genes associated with variation in fatty acid composition. Rapid and low cost fatty acid analysis is, however, needed to match the pace of development in genomic technologies (higher speed genotyping at lower and lower costs). Along this line, the use of newer non-destructive technologies, such as near infra-red spectroscopy (NIRS), to measure beef fatty acid composition have shown promise [87–89], with the ability to predict the content of a number of fatty acids in beef fat related to human health. Further study of fatty acid synthesis and metabolism in beef cattle at the fundamental biochemical and molecular levels is also required to help explain breed, inter-animal and tissue (e.g., adipose vs. muscle) differences. Understanding these differences would then allow opportunities to identify physiological and nutritional factors that influence gene expression and enzyme activity, providing additional avenues to improve beef fatty acid composition [90].
Regulations, recommendations and delivery of PUFA and PUFA-BHP in beef
In the past 10–15 years, manipulating the PUFA and PUFA-BHP content of beef has been intensively investigated. However, to be of practical importance for the industry (1) the profiles and concentrations needed for health benefits must be defined (2) requirements established and (3) source and health claims generated [37]. More importantly, consumers should be informed about potential health benefits of consuming beef products enriched with PUFA or PUFA-BHP. How nutrient source and health claims are handled vary from country to country, and in some countries basic nutritional labelling of foods is not even required [91]. Some countries have an agency that regulates the use of health claims (for example, Health Canada in Canada, the Food and Drug Administration in the USA, The Ministry of Health, Labour, and Welfare in Japan, the Korean Food and Drug Administration, the State Food and Drug Administration in China, the Food Control Department in Singapore and the Department of Health in South Africa). Historically, some governments permitted health claims but left it up to private interests to regulate their use (United Kingdom and Sweden). Other countries have decided to cooperatively develop regulations together on health and nutrition claims (e.g., the European Union, Australia and New Zealand). Given the between country differences, the present review focuses on fatty acid claims permitted in Canada, USA and the European Union as examples of what similarities and differences exsist between countries even when claims are permitted.
Currently, for fatty acids of greatest interest (i.e.,n-3 and certain PUFA-BHP), source claims can only be made for n-3 fatty acids in Canada, the USA and the European Union. In Canada, a source of n-3 fatty acids has to have at least 300 mg of total n-3 fatty acids per 100 g serving [92]. In the USA, foods with ≥ 160 mg or ≥ 320 mg ALA can be referred to as a “source” or “rich” in ALA, and no claims can be made for EPA or DHA [93]. In the European Union, foods with 300 mg ALA or 40 mg combined EPA and DHA per 100 g can be labeled as a source of n-3 fatty acids, and foods with 600 mg ALA or 80 mg combined EPA and DHA per 100 g can be labeled as rich in n-3 fatty acids [94]. Meeting the label requirements for different markets thus require different production strategies.
Irrespective of requirements for each country, it has been difficult to achieve target amounts of n-3 fatty acids in lean beef [95]. LaBrune et al. [63] reached a high of 2.1% ALA in lean beef when feeding flaxseed in a corn-based diet. Estimating 4–6% fat in lean beef, a yield of 84–126 mg ALA per 100 g serving would have been achieved. In pork chops from pigs fed flaxseed, inclusion of some level of external trim fat is required to meet labeling requirements for an n-3 source claim in Canada [96]. Consequently, with slightly more marbling fat or inclusion of a small amount of external trim fat in a serving, the beef from LaBrune et al. [63] may have been able to reach a source claim in the USA (i.e., 160 mg of ALA per serving). Although most studies have not been able to exceed 2% ALA in lean beef fatty acids even when feeding supplemental flaxseed [97–102, 69, 70, 103], there may still be potential to achieve claims in ground beef and further processed beef products. For example, Nassu et al. [70] estimated regular (30% fat) ground beef from flaxseed fed cows would have contained as much as 339 mg of total n-3 fatty acids per 4 oz (114 g) serving. On the other hand, in Europe, meeting a source claim for combined EPA and DHA would be very difficult without feeding some form of protected long-chain n-3 fatty acids [61, 23], although some success has been achieved when feeding fish meal as opposed to fish oil [104]. Again, there would be some potential for a source claim in the EU for ground beef when feeding flaxseed, but only if DPA could be included with EPA and DHA as a long-chain n-3 fatty acid, as is the case in Australia and New Zealand [105], and South Africa [106]. For example, Nassu et al. [70] estimated regular (30% fat) ground beef from flaxseed fed cows would contain as much as 39.4 mg EPA + DPA + DHA per 4 oz (114 g) serving. Consequently, there are definite possibilities to produce beef capable of entering the n-3 fatty acid-enriched market. However, the economic feasibility will depend on balancing the consumer’s willingness to pay for the enhanced nutritional attributes versus the cost of production [107, 108]. Hence research geared to reliably and cost effectively enhance fatty acid composition is of relevance.
Beyond n-3 fatty acids, the greatest potential for enriching beef with healthful fatty acids is likely with PUFA-BHP, specifically VA and CLA. In 2005, Dhiman et al. [109] estimated a serving (100 g) of beef steak enriched with CLA would provide about 41 mg of CLA, and taken together with other foods (mainly whole milk and cheese) would exceed the 300 mg of CLA per day calculated to be required to reduce the incidence of cancer in humans [110]. The major isomer of CLA is RA with its precursor, VA, having a 19% conversion efficiency in humans [111]. Based on RA equivalents (RA + 0.19*VA), Sofi et al. [112] found humans consuming cheese providing 203 mg RA equivalents per day elicited favorable changes in atherosclerotic markers. Consumption of between 200 and 300 mg RA equivalents, therefore, seems to be a reasonable estimate for the amount of RA needed to elicit positive effects on human health. Consumption of 200–300 mg RA is considerably less than the 3.4 g per day thought to be required to induce a reduction in body fat [113]. Recent results indicate enriched beef may be able to provide substantially more than the 41 mg CLA per day as estimated by Dhiman et al. [109]. Noci et al. [67] supplemented pastured heifers with sunflower oil yielding ~127 mg RA equivalents per 100 g serving of lean beef. Mapiye et al. [69] feeding rolled flaxseed together with red clover silage also produced lean beef with 173 mg RA equivalents per 100 g serving. Using a similar feeding strategy in a follow up study Mapiye et al. [102] only produced 29 mg RA equivalents in lean beef, but from the same experiment, Turner et al. [114] produced hamburgers made with 20% perirenal fat that contained 319 mg RA equivalents per 100 g serving. In addition, these hamburgers contained 49 mg of cis9,trans11,cis15-18:3 and 224 mg of its precursor trans11,cis15-18:2. Such alterations in fatty acid profile could add further value to the hamburgers if the health effects of these fatty acids are similar to plant-derived conjugated linolenic acid isomers [115, 116]. However, for any of the PUFA-BHP, their health value in beef still needs to be recognized by regulatory authorities, and recommended intakes need to be defined before requirements for enrichment levels can be established. Subsequent to this, studies would still be necessary to define/refine cost effective production strategies to produce beef with required and consistent enrichments of various PUFA-BHP.
Conclusions
1) Early investigations linking SFA intake with diet-related diseases in humans led to recommendations that consumption of red meat, including beef, should be reduced. Changes in dietary patterns that ensued did not lead to improvements in health, but instead led to increases in prevalence of obesity and type II diabetes. 2) Recommendations to reduce red meat intake still persist, but some recent evidence indicates this may not always be justified. 3) Beef can be an important source of LC n-3 fatty acids, and the potential to increase these should be a research priority. Research should be at the feeding level, but also at the fundamental level in understanding and potentially capitalizing on differences in pathways for LC n-3 fatty acid synthesis. In addition, recent evidence suggests the recognition of DPA, as well as EPA and DHA, as a dietary source of LC n-3 fatty acids, may be justified. 4) The complexity of beef fat composition may also have untapped potential in the form of PUFA-BHP. Although, the concentration of many PUFA-BHP can be quite low, methods to selectively increase or decrease these fatty acids have not been thoroughly investigated. 5) All told, given the complexity and differences in fatty acid composition within beef carcass fat depots (e.g., intramuscular fat versus subcutaneous fat) and the differences in beef product fat content and source, recommendations for beef consumption should not be generalized. Rather these recommendations need to evolve as our knowledge of individual and combined health effects of beef fatty acids develop.
The health effects of some fatty acids (e.g., n-3) are known, and recommended intakes have been defined, leading to opportunities to make enrichment claims in beef. For other fatty acids (e.g., PUFA-BHP), the need for these in the human diet still needs to be accepted by regulatory authorities and source claims developed. Once source claims are possible, production of PUFA-BHP enriched beef could move from proof of concept towards development/refinement of economically feasible production strategies.
Acknowledgements
Research from MERD’s laboratory cited in the review has been supported by the Alberta Meat and Livestock Agency (ALMA) and the Agriculture and Agri-Food Canada (AAFC) Peer Review Program. PV acknowledges NSERC post-doctoral funding provided by the AAFC Peer Review program. NP would like to thank the Alberta Crop Industry Development Fund (ACIDF) for funding support.
Abbreviations
- ALA
α-linolenic acid
- CHD
Coronary heart disease
- DPA
Docoasapentaenenoic acid
- DHA
Docosahexaenoic acid
- EPA
Eicosapentaenoic acid
- HDL
High density lipoprotein
- LA
Linoleic acid
- LC
Long-chain
- LDL
Low density lipoprotein
- MUFA
Monounsaturated fatty acids
- PUFA
Polyunsaturated fatty acid
- PUFA-BHP
PUFA biohydrogenation products
- RA
Rumenic acid
- SFA
Saturated fatty acids
- UFA
Unsaturated fatty acids
- VA
Vaccenic acid
Footnotes
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
PV wrote the review with input and editorial assistance of MERD, TAM and JLA. CM, NP and DCR contributed to research published from MERD’s laboratory and critically reviewed the manuscript. All authors read and approved the final manuscript.
Contributor Information
Payam Vahmani, Email: payam.vahmani@agr.gc.ca.
Cletos Mapiye, Email: cmapiye@sun.ac.za.
Nuria Prieto, Email: nuria.prieto@ualberta.ca.
David C. Rolland, Email: david.rolland@agr.gc.ca
Tim A. McAllister, Email: Jtim.mcallister@agr.gc.ca
Jennifer L. Aalhus, Email: jennifer.aalhus@agr.gc.ca
Michael E. R. Dugan, Email: mike.dugan@agr.gc.ca
References
- 1.Kallas Z, Realini CE, Gil JM. Health information impact on the relative importance of beef attributes including its enrichment with polyunsaturated fatty acids (omega-3 and conjugated linoleic acid) Meat Sci. 2014;97:497–503. doi: 10.1016/j.meatsci.2014.03.015. [DOI] [PubMed] [Google Scholar]
- 2.Realini CE, Kallas Z, Pérez-Juan M, Gómez I, Olleta JL, Beriain MJ, et al. Relative importance of cues underlying Spanish consumers’ beef choice and segmentation, and consumer liking of beef enriched with n-3 and CLA fatty acids. Food Qual Prefer. 2014;33:74–85. [Google Scholar]
- 3.Hu FB. Are refined carbohydrates worse than saturated fat? Am J Clin Nutr. 2010;91:1541–1542. doi: 10.3945/ajcn.2010.29622. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Gross LS, Li L, Ford ES, Liu S. Increased consumption of refined carbohydrates and the epidemic of type 2 diabetes in the United States: an ecologic assessment. Am J Clin Nutr. 2004;79:774–779. doi: 10.1093/ajcn/79.5.774. [DOI] [PubMed] [Google Scholar]
- 5.Hu FB, Stampfer MJ, Manson JE, Ascherio A, Colditz GA, Speizer FE, et al. Dietary saturated fats and their food sources in relation to the risk of coronary heart disease in women. Am J Clin Nutr. 1999;70:1001–1008. doi: 10.1093/ajcn/70.6.1001. [DOI] [PubMed] [Google Scholar]
- 6.Steinberg D. Thematic review series: the pathogenesis of atherosclerosis. An interpretive history of the cholesterol controversy: part II: the early evidence linking hypercholesterolemia to coronary disease in humans. J Lipid Res. 2005;46:179–190. doi: 10.1194/jlr.R400012-JLR200. [DOI] [PubMed] [Google Scholar]
- 7.Mensink RP, Zock PL, Kester AD, Katan MB. Effects of dietary fatty acids and carbohydrates on the ratio of serum total to HDL cholesterol and on serum lipids and apolipoproteins: a meta-analysis of 60 controlled trials. Am J Clin Nutr. 2003;77:1146–1155. doi: 10.1093/ajcn/77.5.1146. [DOI] [PubMed] [Google Scholar]
- 8.Puaschitz NG, Strand E, Norekvål TM, Dierkes J, Dahl L, Svingen GFT, et al. Dietary intake of saturated fat is not associated with risk of coronary events or mortality in patients with established coronary artery disease. J Nutr. 2015 doi: 10.3945/jn.114.203505. [DOI] [PubMed] [Google Scholar]
- 9.Chowdhury R, Warnakula S, Kunutsor S, Crowe F, Ward HA, Johnson L, et al. Association of dietary, circulating, and supplement fatty acids with coronary risk: a systematic review and meta-analysis. Ann Intern Med. 2014;160:398–406. doi: 10.7326/M13-1788. [DOI] [PubMed] [Google Scholar]
- 10.Lawrence GD. Dietary fats and health: dietary recommendations in the context of scientific evidence. Adv Nutr. 2013;4:294–302. doi: 10.3945/an.113.003657. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Dreon DM, Fernstrom HA, Campos H, Blanche P, Williams PT, Krauss RM. Change in dietary saturated fat intake is correlated with change in mass of large low-density-lipoprotein particles in men. Am J Clin Nutr. 1998;67:828–836. doi: 10.1093/ajcn/67.5.828. [DOI] [PubMed] [Google Scholar]
- 12.Nelson GJ, Schmidt PC, Kelley DS. Low-fat diets do not lower plasma cholesterol levels in healthy men compared to high-fat diets with similar fatty acid composition at constant caloric intake. Lipids. 1995;30:969–976. doi: 10.1007/BF02536280. [DOI] [PubMed] [Google Scholar]
- 13.Barendse W. Should animal fats be back on the table? A critical review of the human health effects of animal fat. Anim Prod Sci. 2014;54:831–855. [Google Scholar]
- 14.Micha R, Wallace SK, Mozaffarian D. Red and processed meat consumption and risk of incident coronary heart disease, stroke, and diabetes mellitus: a systematic review and meta-analysis. Circulation. 2010;121:2271–2283. doi: 10.1161/CIRCULATIONAHA.109.924977. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Rohrmann S, Overvad K, Bueno-de-Mesquita HB, Jakobsen MU, Egeberg R, Tjønneland A, et al. Meat consumption and mortality-results from the European prospective investigation into cancer and nutrition. BMC Med. 2013;11:63. doi: 10.1186/1741-7015-11-63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Pan A, Sun Q, Bernstein AM, Schulze MB, Manson JE, Stampfer MJ, et al. Red meat consumption and mortality: results from 2 prospective cohort studies. Arch Intern Med. 2012;172:555–563. doi: 10.1001/archinternmed.2011.2287. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.NCBA. Natoinal Cattlemen’s Beef Association: Average annual per capita consumption beef cuts and ground beef. 2012. http://www.beefusa.org/CMDocs/BeefUSA/Resources/Statistics/averageannualpercapitaconsumptionbeefcutsandgroundbeef559.pdf. Accessed 20 Mar 2015.
- 18.Lim SS, Vos T, Flaxman AD, Danaei G, Shibuya K, Adair-Rohani H, et al. A comparative risk assessment of burden of disease and injury attributable to 67 risk factors and risk factor clusters in 21 regions, 1990–2010: a systematic analysis for the global burden of disease study 2010. Lancet. 2012;380:2224–2260. doi: 10.1016/S0140-6736(12)61766-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Simopoulos A. Evolutionary aspects of diet, the omega-6/omega-3 ratio and genetic variation: nutritional implications for chronic diseases. Biomed Pharmacother. 2006;60:502–507. doi: 10.1016/j.biopha.2006.07.080. [DOI] [PubMed] [Google Scholar]
- 20.Simopoulos AP. Omega-3 fatty acids in health and disease and in growth and development. Am J Clin Nutr. 1991;54:438–463. doi: 10.1093/ajcn/54.3.438. [DOI] [PubMed] [Google Scholar]
- 21.Simopoulos AP. The importance of the omega-6/omega-3 fatty acid ratio in cardiovascular disease and other chronic diseases. Exp Biol Med. 2008;233:674–688. doi: 10.3181/0711-MR-311. [DOI] [PubMed] [Google Scholar]
- 22.Yu M, Gao Q, Wang Y, Zhang W, Li L, Wang Y, et al. Unbalanced omega-6/omega-3 ratio in red meat products in China. J Biomed Mater Res. 2013;27:366–371. doi: 10.7555/JBR.27.20130066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Scollan ND, Dannenberger D, Nuernberg K, Richardson I, MacKintosh S, Hocquette JF, et al. Enhancing the nutritional and health value of beef lipids and their relationship with meat quality. Meat Sci. 2014;97:384–394. doi: 10.1016/j.meatsci.2014.02.015. [DOI] [PubMed] [Google Scholar]
- 24.Flock MR, Harris WS, Kris‐Etherton PM. Long‐chain omega‐3 fatty acids: time to establish a dietary reference intake. Nutr Rev. 2013;71:692–707. doi: 10.1111/nure.12071. [DOI] [PubMed] [Google Scholar]
- 25.Miller E, Kaur G, Larsen A, Loh SP, Linderborg K, Weisinger HS, et al. A short-term n-3 DPA supplementation study in humans. Eur J Clin Nutr. 2013;52:895–904. doi: 10.1007/s00394-012-0396-3. [DOI] [PubMed] [Google Scholar]
- 26.Howe P, Meyer B, Record S, Baghurst K. Dietary intake of long-chain omega-3 polyunsaturated fatty acids: contribution of meat sources. Nutrition. 2006;22:47–53. doi: 10.1016/j.nut.2005.05.009. [DOI] [PubMed] [Google Scholar]
- 27.Dugan M, Aldai N, Aalhus J, Rolland D, Kramer J. Review: Trans-forming beef to provide healthier fatty acid profiles. Can J Anim Sci. 2011;91:545–556. [Google Scholar]
- 28.Jenkins T, Wallace R, Moate P, Mosley E. Board-invited review: Recent advances in biohydrogenation of unsaturated fatty acids within the rumen microbial ecosystem. J Anim Sci. 2008;86:397–412. doi: 10.2527/jas.2007-0588. [DOI] [PubMed] [Google Scholar]
- 29.Aldai N, Dugan ME, Rolland D, Kramer JK. Survey of the fatty acid composition of Canadian beef: backfat and longissimus lumborum muscle. Can J Anim Sci. 2009;89:315–329. [Google Scholar]
- 30.De Smet S, Raes K, Demeyer D. Meat fatty acid composition as affected by fatness and genetic factors: a review. Anim Res. 2004;53:81–98. [Google Scholar]
- 31.Raes K, De Smet S, Demeyer D. Effect of dietary fatty acids on incorporation of long chain polyunsaturated fatty acids and conjugated linoleic acid in lamb, beef and pork meat: a review. Anim Feed Sci Tech. 2004;113:199–221. [Google Scholar]
- 32.Scollan ND, Choi NJ, Kurt E, Fisher AV, Enser M, Wood JD. Manipulating the fatty acid composition of muscle and adipose tissue in beef cattle. Br J Nutr. 2001;85:115–124. doi: 10.1079/bjn2000223. [DOI] [PubMed] [Google Scholar]
- 33.Shingfield KJ, Bonnet M, Scollan ND. Recent developments in altering the fatty acid composition of ruminant-derived foods. Animal. 2013;7:132–162. doi: 10.1017/S1751731112001681. [DOI] [PubMed] [Google Scholar]
- 34.Woods VB, Fearon AM. Dietary sources of unsaturated fatty acids for animals and their transfer into meat, milk and eggs: A review. Livest Sci. 2009;126:1–20. [Google Scholar]
- 35.Wood JD, Enser M, Fisher AV, Nute GR, Sheard PR, Richardson RI, et al. Fat deposition, fatty acid composition and meat quality: A review. Meat Sci. 2008;78:343–358. doi: 10.1016/j.meatsci.2007.07.019. [DOI] [PubMed] [Google Scholar]
- 36.Daley CA, Abbott A, Doyle PS, Nader GA, Larson S. A review of fatty acid profiles and antioxidant content in grass-fed and grain-fed beef. Nutr J. 2010;9:10. doi: 10.1186/1475-2891-9-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Mapiye C, Aldai N, Turner TD, Aalhus JL, Rolland DC, Kramer JKG, et al. The labile lipid fraction of meat: From perceived disease and waste to health and opportunity. Meat Sci. 2012;92:210–220. doi: 10.1016/j.meatsci.2012.03.016. [DOI] [PubMed] [Google Scholar]
- 38.Chilliard Y, Glasser F, Ferlay A, Bernard L, Rouel J, Doreau M. Diet, rumen biohydrogenation and nutritional quality of cow and goat milk fat. Eur J Lipid Sci Tech. 2007;109:828–855. [Google Scholar]
- 39.Harfoot CG, Hazelwood GP. Lipid metabolism in the rumen. In: Hobson PN, Stewart CS, editors. The Rumen Microbial Ecosystem. London, UK: Blackie Academic and Professional; 1998. pp. 382–426. [Google Scholar]
- 40.Juarez M, Dugan ME, Aalhus JL, Aldai N, Basarab JA, Baron VS, et al. Effects of vitamin E and flaxseed on rumen-derived fatty acid intermediates in beef intramuscular fat. Meat Sci. 2011;88:434–440. doi: 10.1016/j.meatsci.2011.01.023. [DOI] [PubMed] [Google Scholar]
- 41.Alves SP, Bessa RJ. The trans-10, cis-15 18: 2: A missing intermediate of trans-10 shifted rumen biohydrogenation pathway? Lipids. 2014;49:527–541. doi: 10.1007/s11745-014-3897-4. [DOI] [PubMed] [Google Scholar]
- 42.Kairenius P, Toivonen V, Shingfield KJ. Identification and ruminal outflow of long-chain fatty acid biohydrogenation intermediates in cows fed diets containing fish oil. Lipids. 2011;46:587–606. doi: 10.1007/s11745-011-3561-1. [DOI] [PubMed] [Google Scholar]
- 43.De La Torre A, Debiton E, Juaneda P, Durand D, Chardigny JM, Barthomeuf C, et al. Beef conjugated linoleic acid isomers reduce human cancer cell growth even when associated with other beef fatty acids. Br J Nutr. 2006;95:346–352. doi: 10.1079/bjn20051634. [DOI] [PubMed] [Google Scholar]
- 44.Dilzer A, Park Y. Implication of conjugated linoleic acid (CLA) in human health. Crit Rev Food Sci Nutr. 2012;52:488–513. doi: 10.1080/10408398.2010.501409. [DOI] [PubMed] [Google Scholar]
- 45.Field CJ, Blewett HH, Proctor S, Vine D. Human health benefits of vaccenic acid. Appl Physiol Nutr Metab. 2009;34:979–991. doi: 10.1139/H09-079. [DOI] [PubMed] [Google Scholar]
- 46.Wang Y, Jacome-Sosa MM, Ruth MR, Goruk SD, Reaney MJ, Glimm DR, et al. Trans-11 vaccenic acid reduces hepatic lipogenesis and chylomicron secretion in JCR: LA-cp rats. J Nutr. 2009;139:2049–2054. doi: 10.3945/jn.109.109488. [DOI] [PubMed] [Google Scholar]
- 47.Smith SB, Gill CA, Lunt DK, Brooks MA. Regulation of fat and fatty acid composition in beef cattle. Asian Austral J Anim Sci. 2009;22:1225–1233. [Google Scholar]
- 48.Dugan M, Kramer J, Robertson W, Meadus W, Aldai N, Rolland D. Comparing subcutaneous adipose tissue in beef and muskox with emphasis on trans 18:1 and conjugated linoleic acids. Lipids. 2007;42:509–518. doi: 10.1007/s11745-007-3051-7. [DOI] [PubMed] [Google Scholar]
- 49.Leheska JM, Thompson LD, Howe JC, Hentges E, Boyce J, Brooks JC, et al. Effects of conventional and grass-feeding systems on the nutrient composition of beef. J Anim Sci. 2008;86:3575–3585. doi: 10.2527/jas.2007-0565. [DOI] [PubMed] [Google Scholar]
- 50.Bauchart D, Roy A, Lorenz S, Chardigny JM, Ferlay A, Gruffat D, et al. Butters varying in trans 18:1 and cis-9, trans-11 conjugated linoleic acid modify plasma lipoproteins in the hypercholesterolemic rabbit. Lipids. 2007;42:123–133. doi: 10.1007/s11745-006-3018-0. [DOI] [PubMed] [Google Scholar]
- 51.Roy A, Chardigny JM, Bauchart D, Ferlay A, Lorenz S, Durand D, et al. Butters rich either in trans-10-C18: 1 or in trans-11-C18: 1 plus cis-9, trans-11 CLA differentially affect plasma lipids and aortic fatty streak in experimental atherosclerosis in rabbits. Animal. 2007;1:467–476. doi: 10.1017/S175173110770530X. [DOI] [PubMed] [Google Scholar]
- 52.Purchas RW, Knight TW, Busboom JR. The effect of production system and age on concentrations of fatty acids in intramuscular fat of the longissimus and triceps brachii muscles of Angus-cross heifers. Meat Sci. 2005;70:215–221. doi: 10.1016/j.meatsci.2004.12.020. [DOI] [PubMed] [Google Scholar]
- 53.Freitas AK, Lobato JFP, Cardoso LL, Tarouco JU, Vieira RM, Dillenburg DR, et al. Nutritional composition of the meat of Hereford and Braford steers finished on pastures or in a feedlot in southern Brazil. Meat Sci. 2014;96:353–360. doi: 10.1016/j.meatsci.2013.07.021. [DOI] [PubMed] [Google Scholar]
- 54.McAfee A, McSorley E, Cuskelly G, Fearon A, Moss B, Beattie J, et al. Red meat from animals offered a grass diet increases plasma and platelet n-3 PUFA in healthy consumers. Br J Nutr. 2011;105:80–89. doi: 10.1017/S0007114510003090. [DOI] [PubMed] [Google Scholar]
- 55.Adams TH, Walzem RL, Smith DR, Tseng S, Smith SB. Hamburger high in total, saturated and trans-fatty acids decreases HDL cholesterol and LDL particle diameter, and increases TAG, in mildly hypercholesterolaemic men. Br J Nutr. 2010;103:91–98. doi: 10.1017/S0007114509991516. [DOI] [PubMed] [Google Scholar]
- 56.Gonzalez L, Moreno T, Bispo E, Dugan ME, Franco D. Effect of supplementing different oils: linseed, sunflower and soybean, on animal performance, carcass characteristics, meat quality and fatty acid profile of veal from “Rubia Gallega” calves. Meat Sci. 2014;96:829–836. doi: 10.1016/j.meatsci.2013.09.027. [DOI] [PubMed] [Google Scholar]
- 57.McNiven M, Duynisveld J, Turner T, Mitchell A. Ratio of n-6/n-3 in the diets of beef cattle: Effect on growth, fatty acid composition, and taste of beef. Anim Feed Sci Tech. 2011;170:171–181. [Google Scholar]
- 58.Jenkins TC, Bridges WC. Protection of fatty acids against ruminal biohydrogenation in cattle. Eur J Lipid Sci Technol. 2007;109:778–789. [Google Scholar]
- 59.Oliveira EA, Sampaio AAM, Henrique W, Pivaro TM, Rosa BL, Fernandes ARM, et al. Quality traits and lipid composition of meat from Nellore young bulls fed with different oils either protected or unprotected from rumen degradation. Meat Sci. 2012;90:28–35. doi: 10.1016/j.meatsci.2011.05.024. [DOI] [PubMed] [Google Scholar]
- 60.McDonald IW, Scott TW. Foods of ruminant origin with elevated content of polyunsaturated fatty acids. World Rev Nutr Diet. 1977;26:144–207. doi: 10.1159/000428538. [DOI] [PubMed] [Google Scholar]
- 61.Dunne PG, Rogalski J, Childs S, Monahan FJ, Kenny DA, Moloney AP. Long chain n-3 polyunsaturated fatty acid concentration and color and lipid stability of muscle from heifers offered a ruminally protected fish oil supplement. J Agric Food Chem. 2011;59:5015–5025. doi: 10.1021/jf200232e. [DOI] [PubMed] [Google Scholar]
- 62.Ford AL, Park RJ, Ratcliff D. Effect of protected lipid supplement on flavor properties of beef. J Food Sci. 1976;41:94–96. [Google Scholar]
- 63.LaBrune H, Reinhardt C, Dikeman M, Drouillard J. Effects of grain processing and dietary lipid source on performance, carcass characteristics, plasma fatty acids, and sensory properties of steaks from finishing cattle. J Anim Sci. 2008;86:167–172. doi: 10.2527/jas.2007-0011. [DOI] [PubMed] [Google Scholar]
- 64.Van Nevel C, Demeyer D. Influence of pH on lipolysis and biohydrogenation of soybean oil by rumen contents in vitro. Reprod Nutr Dev. 1996;36:53–63. doi: 10.1051/rnd:19960105. [DOI] [PubMed] [Google Scholar]
- 65.Aldai N, Dugan ME, Juarez M, Martinez A, Osoro K. Double-muscling character influences the trans-18:1 and conjugated linoleic acid profiles in concentrate-fed yearling bulls. Meat Sci. 2010;85:59–65. doi: 10.1016/j.meatsci.2009.12.004. [DOI] [PubMed] [Google Scholar]
- 66.Aldai N, Nájera A, Dugan M, Celaya R, Osoro K. Characterisation of intramuscular, intermuscular and subcutaneous adipose tissues in yearling bulls of different genetic groups. Meat Sci. 2007;76:682–691. doi: 10.1016/j.meatsci.2007.02.008. [DOI] [PubMed] [Google Scholar]
- 67.Noci F, French P, Monahan FJ, Moloney AP. The fatty acid composition of muscle fat and subcutaneous adipose tissue of grazing heifers supplemented with plant oil-enriched concentrates. J Anim Sci. 2007;85:1062–1073. doi: 10.2527/jas.2006-105. [DOI] [PubMed] [Google Scholar]
- 68.Kronberg SL, Scholljegerdes EJ, Lepper AN, Berg EP. The effect of flaxseed supplementation on growth, carcass characteristics, fatty acid profile, retail shelf life, and sensory characteristics of beef from steers finished on grasslands of the northern Great Plains. J Anim Sci. 2011;89:2892–2903. doi: 10.2527/jas.2011-4058. [DOI] [PubMed] [Google Scholar]
- 69.Mapiye C, Turner T, Rolland D, Basarab J, Baron V, McAllister T, et al. Adipose tissue and muscle fatty acid profiles of steers fed red clover silage with and without flaxseed. Livest Sci. 2013;151:11–20. [Google Scholar]
- 70.Nassu RT, Dugan ME, He ML, McAllister TA, Aalhus JL, Aldai N, et al. The effects of feeding flaxseed to beef cows given forage based diets on fatty acids of longissimus thoracis muscle and backfat. Meat Sci. 2011;89:469–477. doi: 10.1016/j.meatsci.2011.05.016. [DOI] [PubMed] [Google Scholar]
- 71.Lee MR, Tweed JK, Moloney AP, Scollan ND. The effects of fish oil supplementation on rumen metabolism and the biohydrogenation of unsaturated fatty acids in beef steers given diets containing sunflower oil. Anim Sci. 2005;80:361–367. [Google Scholar]
- 72.Duckett SK, Gillis MH. Effects of oil source and fish oil addition on ruminal biohydrogenation of fatty acids and conjugated linoleic acid formation in beef steers fed finishing diets. J Anim Sci. 2010;88:2684–2691. doi: 10.2527/jas.2009-2375. [DOI] [PubMed] [Google Scholar]
- 73.Vahmani P, Fredeen AH, Glover KE. Effect of supplementation with fish oil or microalgae on fatty acid composition of milk from cows managed in confinement or pasture systems. J Dairy Sci. 2013;96:6660–6670. doi: 10.3168/jds.2013-6914. [DOI] [PubMed] [Google Scholar]
- 74.Vasta V, Makkar HPS, Mele M, Priolo A. Ruminal biohydrogenation as affected by tannins in vitro. Br J Nutr. 2009;102:82–92. doi: 10.1017/S0007114508137898. [DOI] [PubMed] [Google Scholar]
- 75.Brogna D, Nasri S, Salem HB, Mele M, Serra A, Bella M, et al. Effect of dietary saponins from Quillaja saponaria L. on fatty acid composition and cholesterol content in muscle Longissimus dorsi of lambs. Animal. 2011;5:1124–1130. doi: 10.1017/S1751731111000048. [DOI] [PubMed] [Google Scholar]
- 76.Lee MR, Evans P, Nute G, Richardson RI, Scollan N. A comparison between red clover silage and grass silage feeding on fatty acid composition, meat stability and sensory quality of the M. Longissimus muscle of dairy cull cows. Meat Sci. 2009;81:738–744. doi: 10.1016/j.meatsci.2008.11.016. [DOI] [PubMed] [Google Scholar]
- 77.Van Ranst G, Lee M, Fievez V. Red clover polyphenol oxidase and lipid metabolism. Animal. 2011;5:512–521. doi: 10.1017/S1751731110002028. [DOI] [PubMed] [Google Scholar]
- 78.Lourenço M, Ramos-Morales E, Wallace RJ. The role of microbes in rumen lipolysis and biohydrogenation and their manipulation. Animal. 2010;4:1008–1023. doi: 10.1017/S175173111000042X. [DOI] [PubMed] [Google Scholar]
- 79.Petri RM, Mapiye C, Dugan ME, McAllister TA. Subcutaneous adipose fatty acid profiles and related rumen bacterial populations of steers fed red clover or grass hay diets containing flax or sunflower-seed. PLoS One. 2014;9:e104167. doi: 10.1371/journal.pone.0104167. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Pitchford WS, Deland M, Siebert BD, Malau-Aduliand A, Bottema CDK. Genetic variation in fatness and fatty acid composition of crossbred cattle. J Anim Sci. 2002;80:2825–2832. doi: 10.2527/2002.80112825x. [DOI] [PubMed] [Google Scholar]
- 81.Kelly MJ, Tume RK, Newman S, Thompson JM. Genetic variation in fatty acid composition of subcutaneous fat in cattle. Anim Prod Sci. 2013;53:129–133. [Google Scholar]
- 82.Li C, Aldai N, Vinsky M, Dugan ME, McAllister TA. Association analyses of single nucleotide polymorphisms in bovine stearoyl-CoA desaturase and fatty acid synthase genes with fatty acid composition in commercial cross-bred beef steers. Anim Genet. 2012;43:93–97. doi: 10.1111/j.1365-2052.2011.02217.x. [DOI] [PubMed] [Google Scholar]
- 83.Morris CA, Cullen NG, Glass BC, Hyndman DL, Manley TR, Hickey SM, et al. Fatty acid synthase effects on bovine adipose fat and milk fat. Mamm Genome. 2007;18:64–74. doi: 10.1007/s00335-006-0102-y. [DOI] [PubMed] [Google Scholar]
- 84.Roy R, Ordovas L, Zaragoza P, Romero A, Moreno C, Altarriba J, et al. Association of polymorphisms in the bovine FASN gene with milk-fat content. Anim Genet. 2006;37:215–218. doi: 10.1111/j.1365-2052.2006.01434.x. [DOI] [PubMed] [Google Scholar]
- 85.Taniguchi M, Utsugi T, Oyama K, Mannen H, Kobayashi M, Tanabe Y, et al. Genotype of stearoyl-coA desaturase is associated with fatty acid composition in Japanese Black cattle. Mamm Genome. 2004;15:142–148. doi: 10.1007/s00335-003-2286-8. [DOI] [PubMed] [Google Scholar]
- 86.Reecy J, Tait R, VanOverbeke D, Garmyn A, Mateescu R, Van Eenennaam A et al., editors. Use of genomics to improve healthfulness and quality of meat. Proceedings of the Ninth World Congress on Genetics applied to Livestock Production: 1-6 August 2010; Leipzig; 2010. Http://www.kongressband.de/wcgalp2010/assets/pdf/0053.pdf.
- 87.Prieto N, Dugan ME, Lopez-Campos O, Aalhus JL, Uttaro B. At line prediction of PUFA and biohydrogenation intermediates in perirenal and subcutaneous fat from cattle fed sunflower or flaxseed by near infrared spectroscopy. Meat Sci. 2013;94:27–33. doi: 10.1016/j.meatsci.2012.12.014. [DOI] [PubMed] [Google Scholar]
- 88.Prieto N, Dugan ME, Lopez-Campos O, McAllister TA, Aalhus JL, Uttaro B. Near infrared reflectance spectroscopy predicts the content of polyunsaturated fatty acids and biohydrogenation products in the subcutaneous fat of beef cows fed flaxseed. Meat Sci. 2012;90:43–51. doi: 10.1016/j.meatsci.2011.05.025. [DOI] [PubMed] [Google Scholar]
- 89.Prieto N, Lopez-Campos O, Aalhus JL, Dugan ME, Juarez M, Uttaro B. Use of near infrared spectroscopy for estimating meat chemical composition, quality traits and fatty acid content from cattle fed sunflower or flaxseed. Meat Sci. 2014;98:279–288. doi: 10.1016/j.meatsci.2014.06.005. [DOI] [PubMed] [Google Scholar]
- 90.Gruffat D, Cherfaoui M, Bonnet M, Thomas A, Bauchart D, Durand D. Breed and dietary linseed affect gene expression of enzymes and transcription factors involved in n-3 long chain polyunsaturated fatty acids synthesis in longissimus thoracis muscle of bulls. J Anim Sci. 2013;91:3059–3069. doi: 10.2527/jas.2012-6112. [DOI] [PubMed] [Google Scholar]
- 91.Malla S, Hobbs JE, Sogah EK. Functional foods and natural health products regulations in Canada and around the world: nutrition labels and health claims. Saskatoon, Saskatchewan, Canada: Report prepared for the Canadian Agricultural Innovation and Regulation Network; 2013. [Google Scholar]
- 92.Agency CFI. Omega-3 and Omega-6 Polyunsaturated Fatty Acid Claims. 2014. http://www.inspection.gc.ca/food/labelling/food-labelling-for-industry/nutrient-content/specific-claim-requirements/eng/1389907770176/1389907817577?chap=7. Accessed 20 Apr 2015.
- 93.Dept-Health-and-Human-Services Food labeling: nutrient content claims; alpha-linolenic acid, eicosapentaenoic acid, and docosahexaenoic acid omega-3 fatty acids. Fed Regist. 2014;79:23262–23273. [Google Scholar]
- 94.Authority EUFS. Scientific opinion: Labelling reference intake values for n-3 and n-6 polyunsaturated fatty acids. EPSA. 2009;1176:1–11. [Google Scholar]
- 95.Moloney AP, Scollan N, Miles L. Enrichment of n‐3 fatty acids and conjugated linoleic acid in beef: ProSafeBeef. Nutr Bull. 2008;33:374–381. [Google Scholar]
- 96.Turner TD, Mapiye C, Aalhus JL, Beaulieu AD, Patience JF, Zijlstra RT, et al. Flaxseed fed pork: n-3 fatty acid enrichment and contribution to dietary recommendations. Meat Sci. 2014;96:541–547. doi: 10.1016/j.meatsci.2013.08.021. [DOI] [PubMed] [Google Scholar]
- 97.Aharoni Y, Orlov A, Brosh A. Effects of high-forage content and oilseed supplementation of fattening diets on conjugated linoleic acid (CLA) and trans fatty acids profiles of beef lipid fractions. Anim Feed Sci Tech. 2004;117:43–60. [Google Scholar]
- 98.Archibeque SL, Lunt DK, Gilbert CD, Tume RK, Smith SB. Fatty acid indices of stearoyl-CoA desaturase do not reflect actual stearoyl-CoA desaturase enzyme activities in adipose tissues of beef steers finished with corn-, flaxseed-, or sorghum-based diets. J Anim Sci. 2005;83:1153–1166. doi: 10.2527/2005.8351153x. [DOI] [PubMed] [Google Scholar]
- 99.Barton L, Marounek M, Kudrna V, Bures D, Zahradkova R. Growth performance and fatty acid profiles of intramuscular and subcutaneous fat from Limousin and Charolais heifers fed extruded linseed. Meat Sci. 2007;76:517–523. doi: 10.1016/j.meatsci.2007.01.005. [DOI] [PubMed] [Google Scholar]
- 100.Hollo G, Ender K, Loki K, Seregi J, Hollo I, Nuernberg K. Carcass characteristics and meat quality of Hungarian Simmental young bulls fed different forage to concentrate ratios with or without linseed supplementation. Archiv fur Tierzucht. 2008;51:517–530. [Google Scholar]
- 101.Kim CM, Kim JH, Chung TY, Park KK. Effects of flaxseed diets on fattening response of Hanwoo cattle: 2. Fatty acid composition of serum and adipose tissues. Asian Austral J Anim Sci. 2004;17:1246–1254. [Google Scholar]
- 102.Mapiye C, Aalhus JL, Turner TD, Rolland DC, Basarab JA, Baron VS, et al. Effects of feeding flaxseed or sunflower-seed in high-forage diets on beef production, quality and fatty acid composition. Meat Sci. 2013;95:98–109. doi: 10.1016/j.meatsci.2013.03.033. [DOI] [PubMed] [Google Scholar]
- 103.Raes K, Haak L, Balcaen A, Claeys E, Demeyer D, De Smet S. Effect of linseed feeding at similar linoleic acid levels on the fatty acid composition of double-muscled Belgian Blue young bulls. Meat Sci. 2004;66:307–315. doi: 10.1016/S0309-1740(03)00105-0. [DOI] [PubMed] [Google Scholar]
- 104.Mandell IB, Buchanan-Smith JG, Holub BJ. Enrichment of beef with omega 3 fatty acids. World Rev Nutr Diet. 1998;83:144–159. doi: 10.1159/000059659. [DOI] [PubMed] [Google Scholar]
- 105.Capra S. Nutrient reference values for Australia and New Zealand: Including recommended dietary intakes. Commonwealth of Australia: Australian Dept of Health and Aging and the New Zealand Ministry of Health; 2006. [Google Scholar]
- 106.Department of Health. South African Government Gazette 32975. Regulations relating to the labelling and advertising of foodstuffs. 2010. http://www.danone.co.za/upload/R146%20of%201%20March%202010.pdf. Accessed 20 April 2015.
- 107.Grunert KG. Food quality and safety: consumer perception and demand. Eur Rev Agric Econ. 2005;32:369–391. [Google Scholar]
- 108.Xue H, Mainville D, You W, Nayga RM., Jr . Nutrition knowledge, sensory characteristics and consumers’ willingness to pay for pasture-fed beef. Milwaukee, WI: Agricultural & Applied Economics Association’s 2009 AAEA & ACCI Joint Annual Meeting; 2009. [Google Scholar]
- 109.Dhiman TR, Nam S-H, Ure AL. Factors affecting conjugated linoleic acid content in milk and meat. Crit Rev Food Sci Nutr. 2005;45:463–482. doi: 10.1080/10408390591034463. [DOI] [PubMed] [Google Scholar]
- 110.McGuire MA, McGuire MK. Conjugated linoleic acid (CLA): A ruminant fatty acid with beneficial effects on human health. J Anim Sci. 2000;77:1–8. [Google Scholar]
- 111.Turpeinen AM, Mutanen M, Aro A, Salminen I, Basu S, Palmquist DL, et al. Bioconversion of vaccenic acid to conjugated linoleic acid in humans. Am J Clin Nutr. 2002;76:504–510. doi: 10.1093/ajcn/76.3.504. [DOI] [PubMed] [Google Scholar]
- 112.Sofi F, Buccioni A, Cesari F, Gori AM, Minieri S, Mannini L, et al. Effects of a dairy product (pecorino cheese) naturally rich in cis-9, trans-11 conjugated linoleic acid on lipid, inflammatory and haemorheological variables: A dietary intervention study. Nutr Metab Cardiovasc Dis. 2010;20:117–124. doi: 10.1016/j.numecd.2009.03.004. [DOI] [PubMed] [Google Scholar]
- 113.Blankson H, Stakkestad JA, Fagertun H, Thom E, Wadstein J, Gudmundsen O. Conjugated linoleic acid reduces body fat mass in overweight and obese humans. J Nutr. 2000;130:2943–2948. doi: 10.1093/jn/130.12.2943. [DOI] [PubMed] [Google Scholar]
- 114.Turner T, Aalhus J, Mapiye C, Rolland D, Larsen I, Basarab J, et al. Effects of diets supplemented with sunflower or flax seeds on quality and fatty acid profile of hamburgers made with perirenal or subcutaneous fat. Meat Sci. 2015;99:123–131. doi: 10.1016/j.meatsci.2014.08.006. [DOI] [PubMed] [Google Scholar]
- 115.Hennessy AA, Ross RP, Devery R, Stanton C. The health promoting properties of the conjugated isomers of α-linolenic acid. Lipids. 2011;46:105–119. doi: 10.1007/s11745-010-3501-5. [DOI] [PubMed] [Google Scholar]
- 116.Koba K, Belury MA, Sugano M. Potential health benefits of conjugated trienoic acids. Lipid Tech. 2007;19:200–203. [Google Scholar]