

Abstract
Background
RNA interference (RNAi) technology shows a great potential in controlling agricultural pests, despite the difficulty of introducing exogenous dsRNA/siRNA into target pests. Isaria fumosorosea is a common fungal pathogen of the B-biotype Bemisia tabaci (whitefly), which is a widespread pest. Entomopathogenic fungi directly penetrate the cuticle and invade insect hemocoel. Application of I. fumosorosea expressing dsRNA of whitefly immunity-related gene may aid in developing RNAi technology to effectively control whiteflies.
Methods
A dsRNA expression plasmid, psTLR7, was constructed by introducing the Toll-like receptor 7 (TLR7) gene of B-biotype whitefly to the silent vector, pSilent-1. The plasmid psTLR7 was transferred into the protoplast of the I. fumosorosea strain IfB01. Then, the recombinant strain was screened out based on the biological stability and bioactivity against whitefly.
Results
A genetically stable recombinant strain IfB01-TRL7 was screened out. The impact of IfB01-TRL7 against whitefly TRL7 gene was validated by qPCR. Lower expression levels of the TLR7 gene was observed in the whiteflies infected by the recombinant strain. The bioassay results indicated that compared to IfB01 strain, IfB01-TRL7 increased the mortality of whitefly nymphs, and decreased and shortened the values of LC50 and LT50, thus indicating higher virulence of IfB01-TRL7.
Conclusion
The expression of the dsRNA of whitefly TLR7 gene in recombinant I. fumosorosea strain successfully knocked down the host target gene by infecting the nymphs and enhanced the whiteflies mortality. The present study will give insight to new application of RNAi technology for more effective biocontrol of this pests.
Electronic supplementary material
The online version of this article (doi:10.1186/s12896-015-0170-8) contains supplementary material, which is available to authorized users.
Keywords: Isaria fumosorosea, Recombinant strain, dsRNA, Whitefly
Background
Bemisia tabaci, commonly known as the whitefly, is an important species complex, and its B-biotype has been named as a superbug [1, 2]. Whiteflies not only directly affect the growth and development of host plants by sucking sap, but also seriously injure the crop production by spreading begomoviruses, such as Tomato yellow leaf curl virus (TYLCV) and Tomato yellow leaf curl China virus (TYLCCNV) [3–5]. At present, chemical insecticides remain the main mode of pest control. However, the serious problems associated with the increasing use of chemicals, such as environmental pollution, food security, and poisoning, demand an urgent need for new alternative technologies [3].
RNA interference (RNAi) is considered to be a novel technology to control agricultural pests [4–7]. However, the difficulty of introducing exogenous dsRNA/siRNA into target pests blocks its application in fields. To date, the common dsRNA/siRNA delivery methods, including injection and oral feeding, are mainly used under laboratory conditions, and only a few transgenic host plants have been used in the fields after safety evaluation [7–9]. Several researchers have reported about the whitefly RNAi. In one such study, the injection of dsRNA molecules (directed specifically towards genes uniquely expressed in the midgut and salivary glands) into the body cavity of whiteflies caused 70 % reduction in the gene expression levels as compared to the whiteflies injected with buffer or with a green fluorescent protein (GFP)-specific dsRNA [10]. By oral route, the ds/siRNAs of actin ortholog, ADP/ATP translocase, alpha-tubulin, ribosomal protein L9 (RPL9), and V-ATPase A subunit caused 29–97 % mortality in the whitefly [11]. Recently, transgenic tobacco lines were developed for the expression of long dsRNA precursors of siRNA and for knocking down the V-ATPase A mRNA in whiteflies. When these pests fed on the transgenic plants, their transcript level of V-ATPase A was reduced down to 62 % [12]. In our previous studies, the dsRNAs of five genes (TLR7, GNBP1, integrin alpha-PS1-like isoform 3, C-type lectin-like precursor and phenoloxidase subunit A3-like) related to innate immunity were delivered into whitefly adults through oral and contact treatments. The 68–96 % reduction of gene expressions and 71–97 % mortalities of whitefly adults were yielded in the oral treatments, at a dosage of 100 μg/mL. Among them, the Toll-like receptor 7 (TLR7) was found to be the most susceptible gene. However, the contact treatments failed to deliver any impact (Hu et al., unpublished data). These findings suggest that RNAi technology can be further developed to control the spread of whiteflies.
Entomopathogenic fungi are often used as myco-insecticides to control sucking sap pests. Entomogenous fungi mainly invade insects through the cuticle and morphologically change into blastospores after entering the host’s hemocoel to adapt the hemolymph immunity [13, 14]. Toll pathway plays important role in defending fungal infection. The induction of the Toll pathway by fungi leads to the activation of cellular immunity as well as the systemic production of certain antimicrobial peptides. The Toll receptors are essential for immunity. They are activated when the proteolytically cleaved ligand Spaetzle binds to the receptors, eventually leading to the activation of the NF-kB factors Dorsal-related immunity factor or Dorsal. In total, nine Toll receptors are encoded in the Drosophila genome [15, 16]. However, in Drosophila, Toll-7 is considered an antiviral gene for pattern recognition receptors of vesicular stomatitis virus [17]. Many Toll-like receptors (TLRs) were discovered in other insects, but their exact functions are not clear yet [18, 19].
Isaria fumosorosea is a species complex of entomopathogenic fungus with wide geographical distribution and extensive range of host insects, including the orders, Hemiptera, Lepidoptera, Diptera, Coleoptera, and Hymenoptera [20, 21]. I. fumosorosea is also the common fungal species infecting the whitefly nymphs. In a previous research, we screened an I. fumosorosea strain, IfB01, which performed as a good control against whiteflies [22]. With the advent of gene modification techniques in entomopathogenic fungus [23–25] and the development of RNAi technology, the present study aimed at constructing a recombinant strain, which expresses dsRNA to knock down the whitefly immune-related gene, TLR7, so as to improve the insecticidal activity of the fungal strain.
Materials and methods
Rearing of B-biotype B. tabaci
The B-biotype B. tabaci population used in this experiment was reared for three years and more than twenty generations in a greenhouse with the host plant, Hibiscus rosasinensis Linn. During the bioassay, the adults of whiteflies were moved onto the pot-planted H. rosasinensis for 24 h for laying eggs. Subsequently, the pot plants with eggs were cultured in an incubator at 25 °C, RH 70 %, and under a photoperiod of 12 L:12D.
Culturing of I. fumosorosea strain, IfB01
The I. fumosorosea strain, IfB01, was used as the parent strain. IfB01 was isolated by our research group and reserved at the China Centre for Type Culture Collection (CCTCC, Wuhan, China), with an accession number, M2012800. The slant of IfB01 strain was inoculated on Czapek-Dox plate (peptone, 5 g; NaNO3, 3 g; K2HPO4, 1 g; MgSO4 · 7H2O, 0.5 g; KCl, 0.5 g; FeSO4 · 7H2O, 0.01 g; sucrose, 30 g; agar, 15–20 g; ddH2O, 1000 mL) and incubated at 25 °C for two weeks. The conidia were subsequently collected from the plates and suspended with 0.05 % Tween 80 to attain a concentration of 108 spores/mL. For liquid culture, 1 mL conidial suspension was inoculated into 100 mL Czapek Dox broth and cultured at 25 °C, 150 rpm for 30 h to collect the mycelia for preparation of the protoplast.
Construction of recombinant plasmid
The total RNA from the adult B-biotype of B. tabaci was extracted using Trizol Total RNA Isolation Kit (Takara, Japan) according to the manufacturer’s protocol. The cDNA was synthesized using the M-MLV Reverse Transcriptase (Takara, Japan) following the manufacturer’s protocol. The primers, TLR7F/TLR7R, were designed based on the whitefly TLR7 gene sequence as the target gene (from transcriptome unigene sequence, Unigene20701_B4592195; Additional file 1). The PCR products were detected using 0.8 % agarose gel electrophoresis, and sequenced at the Guangzhou Yingjun Biotechnological Company (Guangzhou, China).
The PCR primers, TLR7F-1/TLR7R-1 and TLR7F-2/TLR7R-2 (Table 1), with restriction enzyme sites were used to the forward and reverse ends of the target fragment. The expected size of the target fragment was 548 bp (Additional file 2). The PCR amplification was performed as following: pre-denaturation at 94 °C for 4 min; 35 cycles of denaturation at 94 °C for 1 min, annealing at 56 °C for 30 s, extension at 72 °C for 45 s; and a final extension at 72 °C for 10 min. PCR products were separated on 0.8 % agarose gels and visualized by ethidium bromide staining. The forward and reverse fragments of the target gene were recovered and confirmed by electrophoresis and sequencing.
Table 1.
List of primers used in PCR
| Name | Sequence | Used for |
|---|---|---|
| TLR7F | 5′-CAC GCC GAA AGT TTC ATC TA-3′ | cDNA PCR |
| TLR7R | 5′-TTG TCG TTC AAA AGG AGGG-3′ | cDNA PCR |
| TLR7F-1 | 5′-CCG CTCGAG CAC GCC GAA AGT TTC ATC TA-3′ XhoI | Forward fragment PCR |
| TLR7R-1 | 5′-AGC AAGCTT TTG TCG TTC AAA AGG AGGG-3′ HindIII | Forward fragment PCR |
| TLR7F-2 | 5′-CGG GGTACC CAC GCC GAA AGT TTC ATC TA-3′ KpnI | Reverse fragment PCR |
| TLR7R-2 | 5′-CAT GCATGC TTG TCG TTC AAA AGG AGGG-3′ SphI | Reverse fragment PCR |
| HygB-F | 5′-CGA CAG CGT CTC CGA CCTGA-3′ | PCR of hygromycin B phosphotransferase gene |
| HygB-R | 5′-TTC GAT GAT GCA GCT TGG GCG-3′ | PCR of hygromycin B phosphotransferase gene |
| TLR7-VF | 5′-GTCACCGACGAAATCC-3′ | Validation for expression of TLR7 gene of whitefly |
| TLR7-VR | 5′-AGAGTCCCAGCCTTGTT-3′ | Validation for expression of TLR7 gene of whitefly |
| β-actin-F | 5′-TCACCACCACAGCTGAGAGA-3′ | Validation for expression of TLR7 gene of whitefly |
| β-actin-R | 5′-CTCGTGGATACCGCAAGATT-3′ | Validation for expression of TLR7 gene of whitefly |
The vector, pSilent-1, was kindly provided by the Fungal Genetic Store Center of America (University of Missouri, Kansas City, MO, USA).
The forward fragment and plasmid pSilent-1 were digested with the double enzymes, XhoI and HindIII, respectively. After purifying, they were linked with T4 ligase enzyme, incubated overnight at 4 °C, and then, transformed into competent E. coli DH5α cells to form the plasmid, pSilent-1-TLR7F. This plasmid and the reverse fragment were then digested with SphI and KpnI, linked T4 ligase enzyme again, and transformed into E. coli DH5α. Finally, the new plasmid was obtained and validated by double enzyme digest with XhoI and HindIII, and SphI and KpnI.
Transformation of IfB01 strain
The preparation and regeneration of protoplasts from the IfB01 strain were carried out according to the protocols described by Li et al. [26]. The protoplasts were obtained from 30 h-old mycelia following treatment with 1 % snailase, 2 % cellulose, and 0.7 mol/L NaCl pH 6.0 (an osmotic stabilizer), and incubation at 30 °C and 100 rpm for 8 h. The protoplasts were suspended in 1 mL STC solution (1.2 mol/L Sorbitol, 10 mmol/L Tris–HCl pH 7.5, 50 mmol/L CaCl2; autoclaved at 121 °C for 25 min); then, 10 μL plasmid pSilent-1 was added to the prepared protoplast suspension and mixed carefully. After holding it at 4 °C for 1 h, 200 μL PTC solution (40 % PEG4000, 10 mmol/L Tris–HCl pH 7.5, 50 mmol/L CaCl2, sterilized with 0.45 μm bacteria filter) were added to the mixture and stored at room temperature for 20 min. Subsequently, 800 μL PTC solution was added drop wise for shock culturing at 50 rpm for 20 min, then 5 mL liquid regeneration medium was added [26] to the culture and incubated overnight at 25 °C and 150 rpm. The recombinants were isolated from the plates of 200 μL protoplast regeneration medium [26] containing 200 μg/mL hygromycin B, cultured with the pellets of the aforementioned mixture at 25 °C for 30 h.
Screening of the recombinant strain
The fungal plaques were picked out and transferred from the regeneration medium plates to PDA plates, and were cultured at 25 °C for a week. The mycelia were then cut with a punch and transplanted into PD broth with Amp (Ampicillin Sodium Salt), and were continually cultured at 25 °C and 150 rpm for 2 days. The mycelia were collected from the broth and used for extracting the recombinant DNA with a DNA Extraction Kit (Bioteke, Beijing, China), using the manufacturer’s protocol. The quality of the extracted DNA was assessed by 0.8 % agarose gel electrophoresis and was later stored at−20 °C. For further PCR validation, the primers TLR7F/TLR7R and HygB-F/HygB-R were used to detect the target fragment and hygromycin B phosphotransferase gene (Hphr), respectively, under the same cycling conditions as described earlier. The PCR products were recovered using 0.8 % agarose gel electrophoresis and subsequently sequenced at the Beijing Liuhe Huada Gene Technology Company (Beijing, China). The strains simultaneously containing fragments of TLR7 and Hphr genes were determined as recombinant strains. The positive strains were inoculated from the broths to PDA plates, and then, ten plaques were picked out for further screening of genetic stable recombinants. They were subjected to three successive transfers of cultures on PDA plates containing 200 μg/mL hygromycin B; then, transferred into PDA plates for three consecutive cultures; and were later transferred into PDA plates containing 200 μg/mL hygromycin B. After the transferring cultures, the conidia were collected for the PCR detection of target fragment and Hphr gene. The best stable recombinant was screened and subjected to studying the colony morphology and analyzed for their biological characteristics.
Bioassay of virulence of IfB01 and IfB01-TRL7 strains against whitefly
The conidia of IfB01 and IfB01-TRL7 strains were collected from Czapek-Dox plates and were suspended in 0.05 % Tween-80 to obtain a final stock concentration of 1 × 108 spores/mL. Further, the working suspensions of concentrations−2 × 107, 1 × 107, 5 × 106, 2.5 × 106, and 1.25 × 106 spores/mL were prepared. The leaf immersion method (China standard NY/T 1154.14-2008) was employed to the bioassay. Before treatment, the potted plants of H. rosasinensis were checked to ensure that approximately 100 sec instar nymphs remained on each leaf. During the treatment, the leaf with whitefly nymphs was dipped into each working suspension for 30 s. After drying, the potted plants were cultured at 25 °C under a photoperiod of 14 L:10D. Four leaves were utilized for each treatment. The control treatment used was 0.05 % Tween-80. The experiment was replicated two times. The cumulate mortality at 4, 6, 8, 10, and 12 d post-treatment was investigated. The calibrated mortalities, LC-p, LT-p curves, and the values of LC50 and LT50 were evaluated by means of SPSS Statistics software version 19 (IBM, USA).
Validation for expression of TLR7 gene of whitefly
The qPCR method [27] was used to validate the expression of TLR7 gene in whitefly. Three cadavers of 3rd–4th instar nymphs infected by IfB01 and IfB01-TRL7 strains were collected for RNA extraction. Three living 3rd–4th instar nymphs were used as controls. The experiment was repeated three times. The primers, TLR7-VF and TLR7-VR (Table 1), were employed for PCR amplification of the TLR7 gene of whitefly. The β-actin gene, considered as an internal reference gene, was amplified with the primers, β-actin-F and β-actin-R (Table 1). The qPCR was performed on Bio-Rad CFX Connect™ Real-time Thermal Cycler (Bio-Rad, USA). The reaction mixture contained target genes DNA template, 1 μL; TLR7-VF, 0.5 μL; TLR7-VR, 0.5 μL; iTaqTM Universal SYBR® Green Supermix, (Bio-Rad, USA), 10 μL; and ddH2O, 8 μL. The cycling conditions involved pre-denaturation for 2 min at 94 °C, and 35 cycles of denaturation for 30 s at 94 °C and annealing for 30 s at 51 °C. The expressions of target genes were quantified by evaluating the values of 2-ΔΔCt.
Results
Construction of dsRNA expression plasmid targeting TLR7 gene of whitefly
The dsRNA expression plasmid targeting TLR7 gene of whitefly was successfully constructed and was named as psTLR7 (Fig. 1). The electrophoretic results indicated that psTLR7 had the three conformations of open loop, close loop and superhelix (Fig. 2a) and had two hydrolysates of ~7500 pb and ~550 pb by double enzymes digestion of XhoI and HindIII or SphI and KpnI (Fig. 2b). The ~550 pb hydrolysate was then sequenced and validated its identity of target fragment (548 bp) (Additional file 2). The size of psTLR7 was about 8 kb, which corroborated the expected 6.9 kb of psilent-1 plus 548 bp each of forward and reverse TLR7 fragments. It was conformed that the expression cassette of hairpin loop dsRNA was successfully cloned into the psilent-1 plasmid. Subsequently, psTLR7 was introduced into the parent strain of I. fumosorosea, IfB01.
Fig. 1.
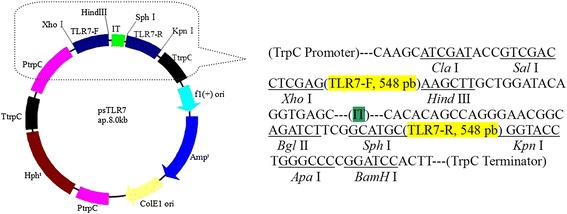
Construction of plasmid psTLR7. IT: intron; TLR7-F: forward fragment of TLR7 gene; TLR7-R: Reverse fragment of TLR7 gene; TtrpC: terminator; PtrpC: promoter; Ampr: Ampicillin resistance gene; Hphr: hygromycin B phosphotransferase resistance gene; XhoI, HindIII, SphIand KpnIare the site for cutting of DNA restrictive enzymes
Fig. 2.
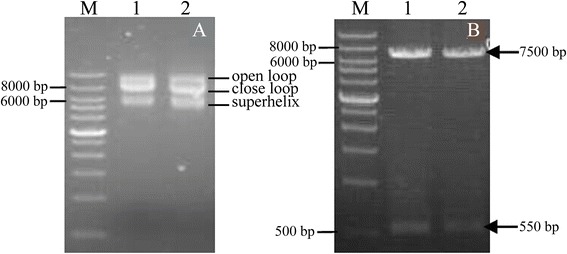
Electrophoresis profile of psTLR7. a: Electrophoresis of psTLR7. lane M, wide range DNA marker, lane 1–2, psTLR7 with the three conformations of open loop, close loop and superhelix. b: Double enzyme digestion of psTLR7. lane M, wide range DNA marker; lane 1, double digestion of psTLR7 by XhoI and HindIII; lane 2, double digestion of psTLR7 by SphI and KpnI
Development of transgenic I. fumosorosea lines
One stably inherited recombinant strain expressing dsRNA of the whitefly TLR7 gene was selected and named as IfB01-TLR7. The strain showed good genetic stability. After transferring for eight generations, the strain still possessed the TLR7 gene fragment (Fig. 3). The target fragment size was ~550 bp, while the Hphr gene fragment was ~850 bp. The sequencing results indicated that the target fragment in IfB01-TLR7 was in agreement with the fragment of psTLR7.
Fig. 3.
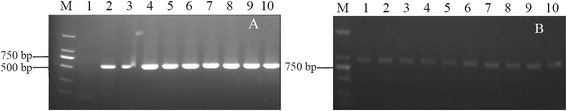
Electrophoresis profile of PCR products of IfB01-TLR7. a: PCR of target fragment; (b): PCR of hygromycin phosphotransferase gene. Lane M: DL2000 Marker, lane 1: plasmid pSilent-1, lane 2: psTLR7, lane 3–10: generation 1–8 of IfB01-TLR7. The objective fragment size was about 550 bp, while the hygromycin B phosphotransferase gene fragment about 850 bp
Although the morphological features of the parent and the recombinant strains were similar, the IfB01-TLR7 showed purple colored colonies and the IfB01 strain presented light yellow colonies. Additionally, the strain IfB01-TLR7 possessed shorter phialides and its conidia were different from the parent strain, IfB01 (Fig. 4).
Fig. 4.

Morphological profile of strain IfB01 and IfB01-TLR7. a: the colony of strain IfB01 on Czapek Dox plate, (b): the sporulation structure and conidia of strain IfB01, c: the colony of strain IfB01-TLR7 on Czapek Dox plate, d: the sporulation structure and conidia of strain IfB01-TLR7
Virulence of IfB01-TLR7 and IfB01 strains against B-biotype B. tabaci
The cumulative calibrated mortalities of whitefly were various in different treatments. Totally, the larger mortalities were observed in the treatments with higher concentrations of spores (Fig. 5). In comparison to the parent strain IfB01, the insecticidal effects of the recombinant strain IfB01-TLR7 were seen to improve evidently in different treatments. After 12 d of treatment at the concentration 2.0 × 107 spores/mL, IfB01-TLR7 obtained a whitefly mortality of 90.33 % compared with 76.00 % of IfB01. Meanwhile, IfB01-TLR7 and IfB01 yielded 42.67 % and 37.67 % mortality rates, respectively, in the treatment with 1.25 × 106 spores/mL on day 4.
Fig. 5.
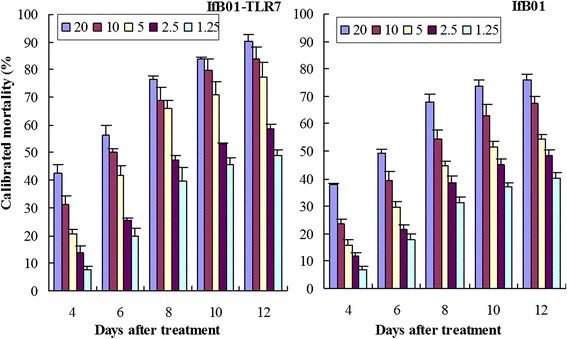
The mortalities of 2nd instar nymphs of B-biotype Bemisia tabaci treated with IfB01-TLR7 and IfB01 strain. The different shaded bars represent different spore concentrations (×106 spores/mL), The error bars represent standard errors
The smaller LC50 values of IfB01-TLR7 indicated that the recombinant strain had higher virulence than the parent strain, IfB01 (Table 2). For instance, on the 8th day of post-treatment, IfB01-TLR7 revealed an LC50 value of 2.37 × 106 spores/mL as compared to a value of 6.18 × 106 spores/mL obtained by the IfB01 strain. In addition, treatment with the concentration of 5 × 106 spores/mL yielded LT50 of 9.35 and 7.07 d in IfB01-TLR7 and IfB01 strains, respectively, indicating shorter periods of LT50 for IfB01-TLR7 (Table 3).
Table 2.
Equations of LC-p and LC50s of fungal strain/dsRNA against whitefly
| Strain | Intercept | Slope | χ 2 | p | LC50 (95 % confidence interval) (×106spores/mL) | |
|---|---|---|---|---|---|---|
| IfB01 | 4d | −1.615 | 0.958 | 1.853 | 0.604 | 48.611 (33.454–83.933) |
| 6d | −1.061 | 0.789 | 0.753 | 0.861 | 22.083 (16.413–33.808) | |
| 8d | −0.609 | 0.771 | 2.764 | 0.429 | 6.175 (5.063–7.618) | |
| 10d | −0.451 | 0.797 | 1.372 | 0.712 | 3.681 (2.935–4.473) | |
| 12d | −0.363 | 0.797 | 1.455 | 0.693 | 2.854 (2.196–3.516) | |
| IfB01-TLR7 | 4d | −1.521 | 1.029 | 0.145 | 0.986 | 30.065 (22.751–44.145) |
| 6d | −0.920 | 0.878 | 4.035 | 0.258 | 11.137 (9.148–14.245) | |
| 8d | −0.311 | 0.828 | 5.594 | 0.061 | 2.374 (1.532–3.150) | |
| 10d | −0.328 | 1.088 | 5.071 | 0.079 | 2.004 (1.403–2.570) | |
| 12d | -0.265 | 1.245 | 4.148 | 0.126 | 1.632 (1.141–2.101) | |
Table 3.
Equations of LT-p and LT50s of fungal strain against whitefly
| Strain (×106spores/mL) | Intercept | Slope | χ 2 | p | LT50 (95 % confidence interval) (d) | |
|---|---|---|---|---|---|---|
| IfB01 | 10 | −1.364 | 0.176 | 2.493 | 0.288 | 7.77 (7.35–8.24) |
| 5 | −1.615 | 0.173 | 3.567 | 0.168 | 9.35 (8.81–10.05) | |
| IfB01-TLR7 | 10 | −1.327 | 0.220 | 1.459 | 0.482 | 6.04 (5.66–6.39) |
| 5 | −1.659 | 0.235 | 12.204 | 0.002 | 7.07 (4.30–9.91) | |
Validation of expression of the whitefly TLR7 gene
In the qPCR validation experiment, the relative expression level of the whitefly TLR7 gene infected by the recombinant strain IfB01-TLR7 was recorded as 0.15, which was lower than that infected by the IfB01 parent strain (0.66) and the control (1.01) (Fig. 6).
Fig. 6.
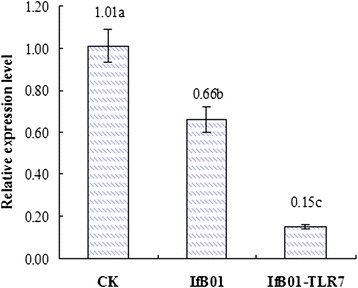
The relative expression level of TLR7 gene of whitefly infected by strain IfB01-TLR7 and IfB01
Discussion
RNAi is recognized as a promising new pest control technology, but there are several problems associated with introducing the exogenous dsRNA/siRNA into target pests, which restrict its practical application in the fields.
To the best of our knowledge, this is the first study wherein a recombinant I. fumosorosea strain (IfB01-TLR7) was constructed for the expression of the dsRNA targeting the immune-related gene, TLR7 of B-biotype B. tabaci. The target gene, TLR7, was successfully knocked down after whitefly nymphs were infected with the IfB01-TLR7 strain. Meanwhile, the insecticidal activity of IfB01-TLR7 against whitefly was apparently increased. Our study have suggested that dsRNA can be delivered into target pests through infection of entomopathogenic fungus, and that the expression of specific dsRNA in entomopathogenic fungus may result in the development of a new RNAi methodology for pest control.
Although the virulence of our new genetically modified fungal strain was not extremely improved in comparison to the parent strain, the increase of more than 2-fold LC50 is still commendable for recombinant strain because the parent strain was selected from lots of wild strains and has excellent effectiveness to whitefly. What is the reason of the genetically modified strain? For the total insecticidal activity of modified strain, it is because of the combination of gene knock down and fungal infection. However, for the increased toxicity (LC50 decreased more than 2-fold), it should be contributed to gene knock down. In the future, it is maybe practicable that introduce two or more exogenous dsRNAs to get more effective genetically modified fungul strain.
The plasmid, pSilent-1, has been commonly used for silencing fungal genes [28]. However, there is no published report on the use of pSilent-1 in developing a transgenic entomogenous fungus strain in order to knock down the insect (host) target gene. The current experiment has, therefore, opened up a new avenue for the application of this gene silencing plasmid.
It is difficult to understand the mechanism of that the dsRNAs produced by cells of IfB01-TLR7 release and enter into the whitefly cells, because there are not sufficient information available. Many researchers have reported that entomogenous fungi penetrate the cuticle of the host and enter the hemocoel to morphologically transform from mycelia to blastospores (yeast-like hyphae). The type of blastospores benefits the fungus spread to whole body and adaptation for the hemolymph immunity [13, 14]. However, the insect host has effective innate immunity system to defend the invasion of fungus. Blastospores are attacked by hemocytes and humoral immunity. Generally, hemocytes damage and digest the blastospores by means of encapsulation, nodulation, and phagocytosis. In the process, the dsRNA may be released from the fungal cells and get absorbed by the hemocytes. This phenomenon may be considered as one of the mechanisms associated with the entry of specific dsRNA into the host cells. Secondly, we supposed that the fungal cells release the plasmid into the host hemocoel or other tissues. This might be attributed to the structure of the blastospore, which has a thick cell wall and hydrophilic cell surfaces [29]. Nevertheless, either of the speculated mechanisms requires further research to elucidate the course of entry of the specific dsRNA into the host cells.
There are still many aspects of this study, which remain unclear; like, how is the systemic RNA interference in host insect arisen. Therefore, further research is required to provide greater insights into the use of RNAi technology for the successful development of myco-insecticides and pest biocontrol.
Conclusion
The expression of the dsRNA of whitefly TLR7 gene in recombinant I. fumosorosea strain successfully knocked down the host target gene by infecting the nymphs and enhanced the whiteflies mortality. The present study will give insight to new application of RNAi technology for more effective biocontrol of this pests.
Acknowledgment
This research was supported by the National High Technology Research and Development Program (‘863’ Program) of China (2012AA101505).
Abbreviations
- Amp
Ampicillin
- cDNA
Complementary DNA
- ddH2O
Double distilled H2O
- DNA
Deoxyribonucleic acid
- dsRNA
Double-stranded ribonucleic Acid
- Hphr
Hygromycin B phosphotransferase gene
- LB
Luria-Bertani
- mRNA
Messenger ribonucleic acid
- PDA
Potato dextrose agar
- PCR
Polymerase chain reaction
- PEG
Polyethylene glycol
- qPCR
Quantitative polymerase chain reaction
- RNA
Ribonucleic acid
- RNAi
Ribonucleic acid interference
- siRNA
Small interfering RNA
- Tris
Tris (Hydroxymethyl) aminomethane
- TLR7
Toll-like receptor 7
Additional files
Unigene20701_B4592195 (TLR7 gene fragment) seguence.
The sequence of fragment in plasmid pSilent-1-TLR7F corresponding to whitefly’s TLR7 gene.
Footnotes
Xiurun Chen and Lin Li contributed equally to this work.
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
XC performed a part of experiments, wrote and partly revised the MS. LL performed the main experiments. QH designed the experiments, partly wrote and revised MS. BZ completed the validation test. WW partly performed the bioassay. FJ and JJ partly designed and revised the paper. All authors read and approved the final paper.
Contributor Information
Xiurun Chen, Email: 2409323925@qq.com.
Lin Li, Email: 578942719@qq.com.
Qiongbo Hu, Email: hqbscau@126.com.
Bowen Zhang, Email: 757510384@qq.com.
Wei Wu, Email: 1028212450@qq.com.
Fengliang Jin, Email: 690646250@qq.com.
Junxi Jiang, Email: 1564739051@qq.com.
References
- 1.Culotta E. Biological Immigrants Under Fire. Science. 1991;6:444–1447. doi: 10.1126/science.254.5037.1444. [DOI] [PubMed] [Google Scholar]
- 2.De Barro PJ, Liu SS, Boykin LM, Dinsdale AB. Bemisia tabaci: A Statement of Species Status. Annu Rev Entomol. 2011;56:1–19. doi: 10.1146/annurev-ento-112408-085504. [DOI] [PubMed] [Google Scholar]
- 3.Wan FH, Zhang GF, Liu SS, Luo C, Chu D, Zhang YJ, et al. Invasive mechanism and management strategy of Bemisia tabaci (Gennadius) biotype B: Progress report of 973 Program on invasive alien species in China. Sci China Ser C. 2009;52:88–95. doi: 10.1007/s11427-008-0135-4. [DOI] [PubMed] [Google Scholar]
- 4.Dong Y, Friedrich M. Nymphal RNAi: systemic RNAi mediated gene knockdown in juvenile grasshopper. BMC Biotechnology 2005;5:25 doi:10.1186/1472-6750-5-25. [DOI] [PMC free article] [PubMed]
- 5.Zhou XG, Wheeler MM, Oi FM, Scharf ME. RNA interference in the termite Reticulitermes flavipes through ingestion of double-stranded RNA. Insect Biochem Mol. 2008;38:805–15. doi: 10.1016/j.ibmb.2008.05.005. [DOI] [PubMed] [Google Scholar]
- 6.Murphy M. Agrochemicals - RNAi for pest control. Chem Ind (London). 2008;11:10–0. [Google Scholar]
- 7.Tian HG, Peng H, Yao Q, Chen HX, Xie Q, Tang B, et al. Developmental control of a lepidopteran pest Spodoptera exigua by ingestion of bacteria expressing dsRNA of a non-midgut gene. PloS One. 2009;13;4(7):e6225. doi:10.1371/journal.pone.0006225. [DOI] [PMC free article] [PubMed]
- 8.Chen J, Zhang D, Yao Q, Zhang J, Dong X, Tian H, et al. Feeding-based RNA interference of a trehalose phosphate synthase gene in the brown planthopper, Nilaparvata lugens. Insect Mol Biol. 2010;19:777–86. doi: 10.1111/j.1365-2583.2010.01038.x. [DOI] [PubMed] [Google Scholar]
- 9.Huvenne H, Smagghe G. Mechanisms of dsRNA uptake in insects and potential of RNAi for pest control: A review. J Insect Physiol. 2010;56:227–35. doi: 10.1016/j.jinsphys.2009.10.004. [DOI] [PubMed] [Google Scholar]
- 10.Ghanim M, Kontsedalov S, Czosnek H. Tissue-specific gene silencing by RNA interference in the whitefly Bemisia tabaci (Gennadius) Insect Biochem Mol. 2007;37:732–8. doi: 10.1016/j.ibmb.2007.04.006. [DOI] [PubMed] [Google Scholar]
- 11.Upadhyay SK, Chandrashekar K, Thakur N, Verma PC, Borgio JF, Singh PK, et al. RNA interference for the control of whiteflies (Bemisia tabaci) by oral route. J Biosci. 2011;36:153–61. doi: 10.1007/s12038-011-9009-1. [DOI] [PubMed] [Google Scholar]
- 12.Thakur N, Upadhyay SK, Verma PC, Chandrashekar K, Tuli R, Singh PK. Enhanced whitefly resistance in transgenic tobacco plants expressing double stranded RNA of v-ATPase A gene. PLoS One. 2014;9(3):e87235. doi:10.1371/journal.pone.0087235. [DOI] [PMC free article] [PubMed]
- 13.Hajek AE, St Leger RJ. Interaction between fungal pathogens and insect hosts. Annu Rev Entomol. 1994;39:293–322. doi: 10.1146/annurev.en.39.010194.001453. [DOI] [Google Scholar]
- 14.Boomsma JJ, Jensen AB, Meyling NV, Eilenberg J. Evolutionary interaction networks of insect pathogenic fungi. Annu Rev Entomol. 2014;59:467–85. doi: 10.1146/annurev-ento-011613-162054. [DOI] [PubMed] [Google Scholar]
- 15.Ashok Y. Drosophila Toll pathway: the new model. Sci Signal. 2009;1. doi:10.1126/scisignal.252jc1. [DOI] [PubMed]
- 16.Valanne S, Wang J-H, Ramet M. The drosophila Toll signaling pathway. J Immunol. 2011;186:649–56. doi: 10.4049/jimmunol.1002302. [DOI] [PubMed] [Google Scholar]
- 17.Nakamoto M, Moy RH, Xu J, Bambina S, Yasunaga A, Shelly SS, et al. Virus Recognition by Toll-7 Activates Antiviral Autophagy in Drosophila. Immunity. 2012;36:658–67. doi: 10.1016/j.immuni.2012.03.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Cheng TC, Zhang YL, Liu C, Xu PZ, Gao ZH, Xia QY, et al. Identification and analysis of Toll-related genes in the domesticated silkworm, Bombyx mori. Dev Comp Immunol. 2008;32:464–75. doi: 10.1016/j.dci.2007.03.010. [DOI] [PubMed] [Google Scholar]
- 19.Luna C, Wang XL, Huang YM, Zhang JA, Zheng LB. Characterization of four Toll related genes during development and immune responses in Anopheles gambiae. Ins Biochem Mol Biol. 2002;32:1171–9. doi: 10.1016/S0965-1748(02)00053-X. [DOI] [PubMed] [Google Scholar]
- 20.Zimmermann G. The entomopathogenic fungi Isaria farinosa (formerly Paecilomyces farinosus) and the Isaria fumosorosea species complex (formerly Paecilomyces fumosoroseus): biology, ecology and use in biological control. Biocontrol Sci Tech. 2008;18:865–901. doi: 10.1080/09583150802471812. [DOI] [Google Scholar]
- 21.Meyling NV, Schmidt NM, Eilenberg J. Occurrence and diversity of fungal entomopathogens in soils of low and high Arctic Greenland. Polar Biol. 2012;35:1439–45. doi: 10.1007/s00300-012-1183-6. [DOI] [Google Scholar]
- 22.Zou CH, Li L, Dong TY, Zhang BW, Hu QB. Joint action of the entomopathogenic fungus Isaria fumosorosea and four chemical insecticides against the whitefly Bemisia tabaci. Biocontrol Sci Tech. 2014;24:315–24. doi: 10.1080/09583157.2013.860427. [DOI] [Google Scholar]
- 23.Wang CS, St Leger RJ. A scorpion neurotoxin increases the potency of a fungal insecticide. Nat Biotechnol. 2007;25:1455–6. doi: 10.1038/nbt1357. [DOI] [PubMed] [Google Scholar]
- 24.Wang CS, Feng MG. Advances in fundamental and applied studies in China of fungal biocontrol agents for use against arthropod pests. Biol Control. 2014;68:129–35. doi: 10.1016/j.biocontrol.2013.06.017. [DOI] [Google Scholar]
- 25.Wang SB, O’Brien TR, Pava-Ripoll M, St Leger RJ. Local adaptation of an introduced transgenic insect fungal pathogen due to new beneficial mutations. Proc Natl Acad Sci U S A. 2011;108:20449–54. doi: 10.1073/pnas.1113824108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Li L, Chen X, Cui C, Jiang J, Hu Q. Studies on conditions for protoplast preparation and transformation of Isaria fumosorosea strain IfB01. Acta Agricul Univ Jiangxiensis. 2014; in press.
- 27.Livak KJ, Schmittgen TD. Analysis of rlative gene expression data using real-time quantitative PCR and the 22DDCT method. Methods. 2001;25:402–8. doi: 10.1006/meth.2001.1262. [DOI] [PubMed] [Google Scholar]
- 28.Nakayashiki H, Hanada S, Quoc NB, Kadotani N, Tosa Y, Mayama S. RNA silencing as a tool for exploring gene function in ascomycete fungi. Fungal Genet Biol. 2005;42:275–83. doi: 10.1016/j.fgb.2005.01.002. [DOI] [PubMed] [Google Scholar]
- 29.Holder DJ, Kirkland BH, Lewis MW, Keyhani NO. Surface characteristics of the entomopathogenic fungus Beauveria (Cordyceps) bassiana. Microbiol Sgm. 2007;153:3448–57. doi: 10.1099/mic.0.2007/008524-0. [DOI] [PubMed] [Google Scholar]