

Abstract
Odontogenic tumors contain a heterogeneous collection of lesions that are categorized from hamartomas to benign and malignant neoplasms of inconstant aggressiveness. Odontogenic tumors are usually extraordinary with assessed frequency of short of 0.5 cases/100,000 population for every year. The lesions such as odontogenic tumors are inferred from the components of the tooth-structuring contraption. They are discovered solely inside the maxillary and mandibular bones. This audit speaks to experiences and cooperation of the molecular and genetic variations connected to the development and movement of odontogenic tumors which incorporate oncogenes, tumor-silencer genes, APC gene, retinoblastoma genes, DNA repair genes, onco-viruses, development components, telomerase, cell cycle controllers, apoptosis-related elements, and regulators/conttrollers of tooth development. The reasonable and better understanding of the molecular components may prompt new ideas for their detection and administrating a better prognosis of odontogenic tumors.
Keywords: Genes, Odontogenesis, Odontogenic tumors
Introduction
A tumor can be defined as a swelling of tissues that does not imply a neoplastic process (1). Neoplasia is a biologic phenomenon that is poorly understood and cannot be exactly differentiated from other processes or tissue reactions (1). The lesions originating from the odontogenic epithelium and remaining entrapped either in adjacent soft tissues or within the jaws are considered odontogenic tumors (2). Neoplasia truly implies the methodology of new development or another growth. The famous British oncologist Willis in 1952 quoted: “A neoplasm is an unusual mass of tissue, the development of which surpasses and is uncoordinated with that of the ordinary tissues and holds on in the same over the top way after cessation of the stimuli which evoked the change”. The hereditary progressions permit unreasonable and unregulated expansion that gets independent (autonomous of physiologic development stimuli), in spite of the fact that tumor for the most part stays subject to have the nourishment and blood supply. Ten million fresh incidents of malignancies and 6 million malignancy related deaths have been reported worldwide in the last decade (3).
Ameloblastoma is a benign tumor of epithelial origin found in jaw bones (4-6). Ameloblastoma is a genuine neoplasm that was perceived as the first odontogenic tumor in 19th century. It has been portrayed suitably by Robinson as a tumor that seems to be generally uni-centric, nonfunctional, irregular in development, anatomically benign, and clinically persistent (7). Information of normal fundamental characteristics, for example, age, area, and radiographic appearance of the different odontogenic tumors could be of great importance in creating a clinical differential diagnosis (8). Like any other neoplasm in the body, odontogenic tumors may mimic cells or tissues of origin histologically, or may be similar to hard dental tissues (enamel, dentin and cementum) or soft tissues of enamel organ or dental pulp (8, 9). The clinical and developmental features of odontogenic tumors have been shown (Figures 1 and 2).
Figure 1.
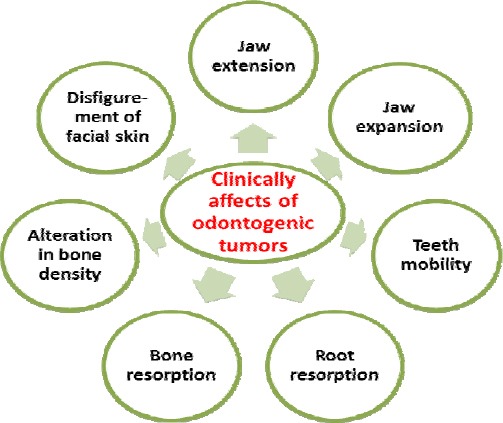
Clinical effects of odontogenic tumors on oral and dental tissues
Figure 2.
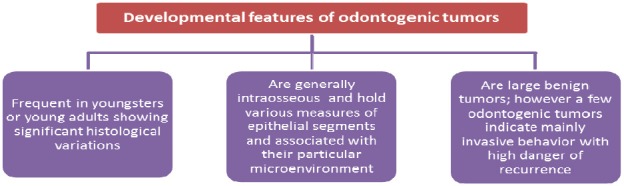
Developmental features of odontogenic tumors
The odontogenic tumors vary from malignant (rare) and benign neoplasms to dental hamartomas. The most common sites of involvement are maxilla and mandible. The sequence of biologic events in the development of neoplasia (carcinogenesis) may occur in odontogenic tumors. Genetic mutations are caused at cellular level, resulting in abnormal control of growth (8, 10). Now, it has been established that genetic damage is a multistep occurrence that may take several years with a long period of time to develop. The aim of this review was to explore information regarding odontogenic tumors. In addition various cell differentiation molecules and tumor genesis mechanisms have been discussed.
Abnormalities in genes regulating cell proliferation underlie neoplastic transformation
There are four phases of malignant tumors progression (11-13): (1) malignant variation in the target cell-transformation; (2) progression of transformed cells; (3) local invasion; (4) distant metastases. The role of various genes in the development of neoplasms has been summarized (Table 1).
Table 1.
Various genes showing expression in different subcategories of odontogenic tumor
| Molecule | Increased expression | Low expression | Reference |
|---|---|---|---|
| Signal transducer Ras (RAS) | Ameloblastoma Ameloblastic fibromas Odontogenic myxomas | (8) | |
| TNF-related apoptosis-inducing ligand (TRAIL) | Granular cell ameloblastoma | Acanthomatous ameloblastoma | (14) |
| single-stranded DNA (ssDNA) | Granular cells of granular cell Ameloblastoma Keratinizing cells of Acanthomatous ameloblastoma | (14) | |
| 53-kilodalton (kDa) protein (P53 gene) | Ameloblastic fibrosarcoma Ameloblastomas, Primary intraosseous carcinomas Malignant ameloblastomas | (18,19) | |
| Fas | Desmoplastic ameloblastoma | Basal cell ameloblastoma | (20) |
| Caspase protein (Caspase 3) | Desmoplastic ameloblastoma | Basal cell ameloblastoma | (20) |
| The transforming growth factor beta (TGF-β) | Epithelial lining of Ameloblastomas Pseudo glandular cells of aots Basal cell ameloblastoma Metastasizing ameloblastoma | Keratinizing cells of acanthomatous ameloblastoma Granular cells of granular cell Ameloblastoma Ccots Ameloblastic carcinomas | (27) |
| Hepatocyte growth factor (HGF) | Basal cell ameloblastoma Pseudo glandular cells of aots Metastasizing ameloblastoma Ccots Ameloblastic carcinomas | (27) | |
| The murine double minute 2 MDM2 | Ameloblastoma | (28,29) | |
| Transforming gene c-Met | Acanthomatous ameloblastoma | (29) |
Oncogenes express inappropriate activity of products under normal circumstances whereas, tumor suppressor genes or anti-oncogenes, under normal circumstances, inhibit cell growth by loss of activity of genes. Generally products produced by over expression of genes; prevent normal cell death hence leading to continued growth of tumors. The products activity of genes is lost under normal circumstances which repairs damaged DNA, hence leading to DNA unsteadiness with the growth of somatic mutations in oncogenes or tumor suppressor genes (8). The genetic reason for neoplastic transformation is variable, for example, the excessive production of oncoproteins result in gene amplification. Similarly, loss of suppressors by point mutations in oncogenes or promoter region may activate oncogenes leading to chromosomal rearrangement.
Cell differentiation molecules for odontogenic tumor genesis
The exact etiology and pathogenesis of odontogenic tumors is unfamiliar, although recent studies have shown several molecular alterations in the formation of odontogenic tumors (8). Advanced research has shown that alteration in key genes is responsible for growth of cells.
Oncogenes
Oncogenes are responsible for autonomous cell evolution in cancer cells and their standard/regular cellular counterparts are known as proto-oncogenes. These genes are responsible for neoplastic transformation by the activation of gene amplification, mutation or translocation (8). The gene products of oncogenes function as growth factor and growth factor receptors such as Epidermal Growth Factor Receptor (EGFR), Fibroblast Growth Factor (FGF), Platelet-Derived Growth Factor (PDGF), HER-2, Ret, non-receptor tyrosine kinases (Src, Abl), signal transducers (Ras), Serine/Threonine kinases (Mos), and transcription factors (Myc, FOS), and play a role in differentiation and proliferation of the cells.
Signal transducer Ras is responsible for over expression for growth factor genes leading to increased production of Tumor Necrotizing Factor α (TNF-α). This growth factor induces proliferation of Epidermal Growth Factor (EGF) by binding to the EGF receptor. The p21Ras are the gene products encoded by Ras gene responsible for transduction of external stimuli prompted by growth factors. In addition, P21Ras expression has been reported in odontogenic epithelial cells in case of ameloblastoma, ameloblastic fibromas and odontogenic myxomas. Increased expression of p21Ras has been also seen in tumors of odontogenic origin when comparing with usual developing dentition (14). Point mutation of Ras gene is reported in a number of tumors like pancreatic, bladder, colorectal and lung carcinomas. K-Ras mutation has been also reported in ameloblastoma.
The Myc oncogene
The Myc protooncogene is an early response gene usually expressed in all eukaryotic cells. These genes are prompted when quiescent cells get a signal to split (15). The molecular function of Myc in cell replication is still unclear. This Myc protein is translocated to the nucleus called MAX and they are considered to be associated with cell proliferation regulated through nuclear phosphoprotein encoded by c-Myc oncogene. They are expressed predominantly in cancerous cells existing in approximation to the basement membrane. Various tumors of odontogenic origin show high incidence of transgenic mice carrier of Myc and/or H-Ras gene (16, 17).
Tumor suppressor genes
Tumor suppressor genes (for example, p53, patched (PTC), WT-1, adenomatous polyposis coli (APC), and retinoblastoma (RB) genes) assume a key part in the development of tumors. These genes play a role in developing cells or genes inactivation by changing and loss of heterozygosity (LOH) in alleles. P53 gene is the most important altered gene in tumors and its products are responsible for genomic destruction by inducing apoptosis or halting in the cell cycle. Certain pathologies such as ameloblastomas, primary intraosseous carcinomas, malignant ameloblastoma, and ameloblastic fibrosarcoma show increased expression of p53 gene; although many studies have shown that p53 mutations are occasional in ameloblastomas. The MDM2 is a protein which is responsible for inhibiting the role of p53 by initiating its degradation so; it is considered a negative regulator of p53 or an inhibitor of tumor suppressor function of p53. Increased expression of MDM2 protein has been seen in various human malignancies, indicating that the oncogene plays a key role in human carcinogenesis therefore, it is associated with a poor prognosis of several human cancers (8).
Over expression of MDM2 has been reported in ameloblastoma and the results show various labeling index of MDM2 based on WHO classification and cytological pattern of outer layer cells. The highest proliferative activity has been reported in basal ameloblastoma with the highest MDM2 labeling index. Hence it was reported that MDM2, is responsible for high proliferative activity of ameloblastoma, but p53 might be responsible, especially in basal ameloblastoma (18, 19). Increased reactivity of p14ARF, MDM2 and p53 were found both in benign as well as malignant changes of the epithelium of odontogenic origin. The reactivity of these genes in various ameloblastomas might be related with tissue structuring and cyto-differentiation; p53 homologs perform an important part in proliferation and differentiation of odontogenic epithelial cells (18, 19).
Adenomatous polyposis coli (APC gene)
Down-regulation of β-catenin is an essential task of APC protein. In the deficiency of Wnt signaling pathway APC results in degradation of β-catenin and subsequently repressing its accumulation in the cytoplasm. Expression of β-catenin has been found in the membrane of the cell and cytoplasm of benign as well as malignant epithelial cells of tooth forming apparatus but the expression of this in nucleus has been only seen in malignant epithelial cells of odontogenic origin (19, 20). The epithelial cells of dental laminae and enamel organ in tooth germ, demonstrate the expression of β-catenin. Expression of both APC and β-catenin in ameloblastomas and germs of tooth reveals the distortion of the Wnt pathway indicating its role in cyto-differentiation and cogenesis of the epithelium of odontogenic origin by means of deregulation of multiplication of cell (19, 20).
Retinoblastoma gene (Rb)
Retinoblastoma gene (Rb) is the first suppressor gene found in malignant tumors of the retina in childhood, which is located on chromosome 13 and termed RB. Hereditary ameloblastoma is a tumor developed from neural precursor cells in the immature retina; it is a rare childhood cancer. The affected child had somatic inactivation of the remaining Rb allele and inherited the mutated Rb allele leading to tumor growth (8). It has been concluded that RB gene is not expressed in tumors of odontogenic origin, although E2F-1 gene expressed by transgenic mice by the promoter of keratin, has been detected in progression of tumors of skin and ameloblastoma (21, 22). Hence it is concluded that genes that suppress tumors are involved in the development of tumors of the tooth forming apparatus (8).
Genes repair DNA
In reducing mutations and maintaining genomic instability, the human DNA disparity reparation (hMMR) system performs a significant role by correcting short insertions/deletions and base-base mismatches produced as a homologous recombination or a consequence of errors by DNA replication. Exclusively nuclear stain is expressed by hMSH2 and hMLH1 in the outermost epithelial layer of all ameloblastomas. The progression and development of odontogenic tumors do not rely on a deficiency in the hMMR system (23).
Oncoviruses
Various numbers of DNA and RNA are considered to be oncogenic and the etiology of a few varieties of tumors are related to viruses for instance Epstein Barr virus (EBV), Human Papilloma Virus (HPV), and Human T-cell Leukemia Virus type 1 (HTCV-1) (8). HPV plays an important role in oral squamous cell carcinomas (OSCC), and its participation in oral tumors has also been put forward (24, 25). A few studies showed the relationship among HPV and odontogenic tumors and cysts with hybridization methods. A few intrabony and peripheral ameloblas-tomas showing the expression of HPV 18 and HPV 16 have been described in an odontogenic keratocyst. Usually HPV is involved with benign oral lesions and majority of these lesions consist of surface epithelium. Ameloblastoma is an epithelium consequential intrabony lesion. HPV showed expression in 42% of ameloblastoma lesions (26).
Epstein barr virus (EBV)
EBV is associated to the herpes genre and has been involved in the pathogenesis of different types of tumors in human; B-cell lymphomas, the African form of Burkitt’s lymphoma, few cases of Hodgkin’s lymphoma, and nasopharyngeal carcinoma. EBV-DNA showed expression in 15% of non-odontogenic tumors, 48% of ameloblastomas and 92% of normal oral tissue (3).
Growth factors
Various types of growth factors such as EGF, TGF- α, TGF-β, FGF, and a few of their receptors have been found in a few odontogenic tumors. The presence of TGF- β and its receptor has been found during tooth development (27). Epithelial-mesenchymal interactions perform a very vital part in normal progression as well as neoplasia.
TGF-β acts as a mediator of these interactions and plays an important role in epithelial tumors. TGF-β showed strongest positivity in epithelial lining of ameloblastomas, and its receptor was diffusely reactive with neoplastic cells. Keratinizing and granular cells in the subcategories of ameloblastoma showed low expression to TGF-β and high expression to Tβ RI and Tβ RII. These expression arrays of TGF-β signaling associated agent were found to be constant with p27Kip1 immuno-reactivity in acanthomatous and granular cell ameloblastomas. On comparing with other subtypes of ameloblastoma, high expressions of HGF were found to be in basal cell ameloblastoma and TGF-β and its receptors in desmoplastic ameloblastoma. TGF-β, HGF and its receptors expression have been found in malignant cells of COCs, AOTs and CEOTs. Over expression of TGF-β and hepatocyte growth factor have been reported in pseudo glandular cells of AOTs and their receptors in COCs predominately in ghost cells (17, 27).
Hepatocyte growth factor (HGF)
This development component is isolated from platelets and serum. The expression of HGF and TGF-β were found together in epithelial and mesenchymal cells of germs of tooth origin as well as odontogenic tumors of epithelial origin, while the reactivity of their receptors were seen just in cells of epithelial origin. They showed their reactivity in epithelium close to the basement membrane (28). Decreased expression of TGF-β, c-Met and HGF and increased expression of receptors of TGF-β, have been found in acanthomatous ameloblastomas predominately in keratinizing cells, whereas very low or nil expression of TGF-β or HGF and their receptors have been seen in granular cells of granular cell ameloblastoma. TGF-β, HGF and their receptors showed reactivity in malignant cells of CEOTs, AOTs and COCs. Pseudo glandular cells of AOTs and metastasizing ameloblastoma showed increased expression of TGF-β, HGF and its receptors showed expression in ghost’s cells of COCs and benign ameloblastomas. The increased expression of HGF and low expression of TGF-β and its receptors have been observed in CCOTs and ameloblastic carcinomas when compared with benign ameloblastoma lesions (29).
Fibroblast growth factor (FGF)
The fibroblast growth factor (FGF) plays an important role in odontogenesis consisting of repetitive signaling between oral/odontogenic epithelium and ectomesenchyme. So et al (29) hypothesized the presence of FGFs in human odontogenesis and performing an important part in the progression and development of the cysts and tumors of odontogenic origin. Expressions in odontogenic cysts and tumors were found to be similar to the epithelium of customary/standard follicles of tooth germ; expression of FGF2 and FGF2 were discovered to be further predictable and extreme in contrast to FGF1 and FGF3, which show either absence or equivocal staining. Usually they showed cytoplasmic stain but FGF2 showed nuclear staining in a few zones of ameloblastic fibromas and ameloblastic fibro-odontomas in epithelial cells. FGF 1 and FGF 2 showed variable expression in ghost cells of COCs. In odontomas, adequate to extreme cytoplasmic staining for FGF-1 and FGFR3 was recognized in ameloblastic cells next to enamel matrix or spaces. Dental follicle, epithelium of odontogenic tumor and normal mouse enamel organ showed that generalized FGF-2 staining uncovers the procedure of differentiation of odontogenic epithelium instead of a non-physiological change in tumor development (29).
Telomerase
A specific converse transcriptase that blends telomeric DNA at the end of chromosomes and adjusts for its compensating to respectively one cell division, taking part in cell deification is known as telomerase (8). Telomerase activity may be connected with oncogenesis of odontogenic epithelium. The outer columnar or cuboidal cells and certain central polyhedral cells of ameloblastomas demonstrated the declaration/expression of telomerase converse transcriptase (TERT), suggesting that the telomerase movement and activity is associated with the proliferative capability of ameloblastoma cells. Nuclei of neoplastic cells of ameloblastoma showed the immune-histochemical reactivity for TERT (30-32). Peripheral columnar or cuboidal cell and some focal polyhedral cells demonstrated the TERT articulation in follicular and plexiform ameloblastomas and granular cell ameloblastoma; predominantly granular cells indicated decreased telomerase converse transcriptase expression compared to non-granular neoplastic cells. TERT showed no expression towards stromal cells (23, 30, 31).
Cell cycle regulators
Cell proliferation trails development by the cell cycle in orderly manner, which is regulated by various elements containing cyclin-dependent kinases (CDKs), cyclins, CDK inhibitors (CKIs), and other critical regulator (8). PCNA is related to DNA synthesis in S phase with proliferation of cells, and is a nuclear protein. Positive PCNA cells have been noticed in dental lamina like odontogenic epithelium especially in case of ameloblastic fibroma, odontogenic ameloblastoma and mesenchymal tissue resembling dental papilla (33).
Cyclin D1 is intensely involved in progression of cell cycle by controlling the move from G1 to S phase, whereas CDK inhibitors, p16 INK4a, p21 WAFI/CipL, and p27KipL hinder progression of cell cycle by cyclin D1 suppression. In ameloblastoma, reactivity of cyclin D1, p16 INK4a, p21 WAFI/CipL, and p27KipL is generally protected recommending that epithelial cells proliferation of odontogenic origin has firmly been regulated using regulators of cell cycle (8). A small number of studies have been accounted for on odontogenic tumors. Ameloblastic fibro-odontoma is for the most part accepted to be a benign tumor; however its histogenesis is questionable (34).
Factors related to apoptosis
In a few epithelial odontogenic tumors, apoptotic cells have been seen by terminal deoxynucleotidyl transferase (TdT) mediated dUTP biotin nick end-labelling (TUNEL) assay, used to perceive those cells which undergo apoptosis and immunohisto-chemistry utilizing single strands of DNA (ssDNA) antibody, concluding that the death of cell assumes a paramount part in oncogenesis and cell differentia-tion of odontogenic epithelium (8). Kumamoto et al (17) observed that bodies left after apoptosis were distinguished in typical and malignant epithelial cells of odontogenic origin disconnected from the basement membrane. Immuno-reactivity for ssDNA was more in granular cell ameloblastoma, especially in granular cells, than in other neoplastic cells. Apoptotic bodies combative with anti-ssDNA antibody were identified as minute circular specks on the nuclei of both normal and malignant odontogenic epithelial cells (14, 35, 36). Scattered apoptotic bodies were seen in tooth germ chiefly in stratum intermedium. Expression of scattered ssDNA reaction shows transition from outer cuboidal or columnar cells to central polyhedral cells in plexiform and Follicular ameloblastomas (20, 32).
Increased reactivity of ssDNA was observed in acanthomatous ameloblastoma especially in keratinizing cells and numerous apoptotic bodies. Basal cell ameloblastoma and desmoplastic ameloblastoma demonstrated comparable to that of follicular ameloblastomas, though negligible apoptotic bodies were seen in ameloblastic carcinoma (20, 32). According to Kumamanto et al (35), the normal and neoplastic odontogenic epithelial cells show the immunoreactions for Fas and FasL. Down regulation of Fas and up regulation of FasL expression were observed in neoplastic ameloblastomas as matched with benign ameloblastoma, signifying the release from death of cells attacked by immune cells. Basal cell ameloblastoma showed low expression, and desmoplastic ameloblastoma showed high expression of Fas and caspase-3 (20, 32).
Expression of TNFα in dental follicles controls progressions included in tooth eruption. Most normal and neoplastic odontogenic tissue displayed the expression of TRAIL and its death receptors. Expression of TRAIL was relatively recurrent in epithelial components of tooth forming apparatus, as paralleled with TNFα expression, and expression was apparent in cells neighboring the basement membrane in ameloblastomas and tooth germs. Low TRAIL expressions were noted of acanthomatous ameloblastoma predominately in keratinizing cells and high in granular cell ameloblastomas (granular cells) (14, 32). The Bcl-2 protein was acknowledged to assume a part in normal tooth development and was located in the tooth germ. Regulation of apoptosis is related to oncogenesis in the epithelium of mucosa. Presence of Bcl-2 protein expression is concluded in ameloblastomas, predominately in the cells of tumor, neighboring the basement membranes (37). In a study by Mitsuyasu et al (38) the protein of Bcl-2 was spotted in the outer layer cells of tumor and in between cells. In basal cell ameloblastoma, there were no account of morphological differences between outer and inner cell layers; all cells expressed the Bcl-2 proteins (38).
Regulators of tooth development
Growth of the teeth is mediated through the genetic control of regulators and position and morphology of the teeth is determined by several genes such as Barx-1, Dlx-2, Msx-1, Msx-2, and Pax-9. SHH signaling molecules such as SMO, PTC, SHH, and GLI1, has been observed in a number of odontogenic tumors of (8). Benign and metastasizing ameloblastomas show the reactivity of SHH predominantly in malignant cells of these tumors. Barreto et al have recognized expression of PTC protein in ameloblastomas, uncovering that the signaling pathway by SHH may be initiated in tumors of odontogenic origin (39). According to Kumamoto et al finding SMO and GLI 1 expression was noted in neoplastic and stromal cells of benign and metastasizing ameloblastoma, and the expression of GLI1 was found to be more obvious in malignant cells than the stromal cells of these tumors. Low SMO expression was noted in these epithelial odontogenic tumors, in contrast the dental laminae and stromal GLI1 expression in ameloblastoma were significantly greater than in the mesenchymal portion of thr developing tooth germ (40).
Conclusion
Alteration in various types of genes and molecules affects the development and progression of odontogenic tumors. Comparative analysis provides visions into the mechanisms of the differentiation of physiological odontogenesis and pathological tumors. The odontogenic tumor epithelial cells’ capability to reiterate genetic programs expressed during normal odontogenesis is well known, yet the unusual representation of these genes is expressed in tumor cells, which needs further research.
Footnotes
Conflict of interest
The authors declare that they have no conflicts of interest in conducting this research.
References
- 1.Rajendran R. India: Elsevier; 2009. Shafer's textbook of oral pathology. [Google Scholar]
- 2.Regezi JA, Sciubba JJ, Jordan RC. Elsevier Health Sciences; 2012. Oral pathology: clinical pathologic correlations. [Google Scholar]
- 3.Kumar V, Abbas AK, Fausto N, Aster JC. Elsevier Health Sciences; 2009. Robbins and Cotran Pathologic Basis of Disease, Professional Edition: Expert Consult-Online. [Google Scholar]
- 4.Reichart P, Philipsen H, Sonner S. Ameloblastoma: biological profile of 3677 cases. Eur J Cancer Part B: Oral Oncol. 1995;31:86–99. doi: 10.1016/0964-1955(94)00037-5. [DOI] [PubMed] [Google Scholar]
- 5.Nakamura N, Mitsuyasu T, Higuchi Y, Sandra F, Ohishi M. Growth characteristics of ameloblastoma involving the inferior alveolar nerve: a clinical and histopathologic study. Oral Surg Oral Med Oral Pathol Oral Radiol Endod. 2001;91:557–562. doi: 10.1067/moe.2001.113110. [DOI] [PubMed] [Google Scholar]
- 6.Becelli R, Carboni A, Cerulli G, Perugini M, Iannetti G. Mandibular ameloblastoma: analysis of surgical treatment carried out in 60 patients between 1977 and 1998. J Craniofac Surg. 2002;13:395–400. doi: 10.1097/00001665-200205000-00006. [DOI] [PubMed] [Google Scholar]
- 7.Stevens A, Lowe J. New York: Mosbay; 2000. Pathology. [Google Scholar]
- 8.Kumamoto H. Molecular pathology of odontogenic tumors. J Oral Pathol Med. 2006;35:65–74. doi: 10.1111/j.1600-0714.2006.00380.x. [DOI] [PubMed] [Google Scholar]
- 9.Gazal G, Fareed WM, Zafar MS, Al-Samadani KH. Pain and anxiety management for pediatric dental procedures using various combinations of sedative drugs: A review. Saudi Pharm J. 2014 doi: 10.1016/j.jsps.2014.04.004. Published ahead of print. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Cawson RA. 7th ed. Edinburgh: Churchill Livingstone; 2002. Cawson's Essentials of Oral Pathology and Oral Medicine. [Google Scholar]
- 11.Klein CA. Selection and adaptation during metastatic cancer progression. Nature. 2013;501:365–372. doi: 10.1038/nature12628. [DOI] [PubMed] [Google Scholar]
- 12.Beerenwinkel N, Antal T, Dingli D, Traulsen A, Kinzler KW, Velculescu VE, et al. Genetic progression and the waiting time to cancer. PLoS Comput Biol. 2007;3:e225. doi: 10.1371/journal.pcbi.0030225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Michor F, Iwasa Y, Nowak MA. Dynamics of cancer progression. Nat Rev Cancer. 2004;4:197–205. doi: 10.1038/nrc1295. [DOI] [PubMed] [Google Scholar]
- 14.Kumamoto H, Ooya K. Expression of tumor necrosis factor α, TNF-related apoptosis-inducing ligand, and their associated molecules in ameloblastomas. J Oral Pathol Med. 2005;34:287–294. doi: 10.1111/j.1600-0714.2005.00311.x. [DOI] [PubMed] [Google Scholar]
- 15.Bishop JM. Molecular themes in oncogenesis. Cell. 1991;64:235–248. doi: 10.1016/0092-8674(91)90636-d. [DOI] [PubMed] [Google Scholar]
- 16.Kumamoto H, Takahashi N, Ooya K. K-Ras gene status and expression of Ras/mitogen-activated protein kinase (MAPK) signaling molecules in ameloblastomas. J Oral Pathol Med. 2004;33:360–367. doi: 10.1111/j.1600-0714.2004.00141.x. [DOI] [PubMed] [Google Scholar]
- 17.Kumamoto H, Ooya K. Detection of mitochondria-mediated apoptosis signaling molecules in ameloblastomas. J Oral Pathol Med. 2005;34:565–572. doi: 10.1111/j.1600-0714.2005.00354.x. [DOI] [PubMed] [Google Scholar]
- 18.Kumamoto H, Izutsu T, Ohki K, Takahashi N, Ooya K. p53 gene status and expression of p53, MDM2, and p14ARF proteins in ameloblastomas. J Oral Pathol Med. 2004;33:292–299. doi: 10.1111/j.0904-2512.2004.00044.x. [DOI] [PubMed] [Google Scholar]
- 19.Kumamoto H, Ohki K, Ooya K. Expression of p63 and p73 in ameloblastomas. J Oral Pathol Med. 2005;34:220–226. doi: 10.1111/j.1600-0714.2005.00284.x. [DOI] [PubMed] [Google Scholar]
- 20.Kumamoto H, Kimi K, Ooya K. Immunohistochemical analysis of apoptosis-related factors (Fas, Fas ligand, caspase-3 and single-stranded DNA) in ameloblastomas. J Oral Pathol Med. 2001;30:596–602. doi: 10.1034/j.1600-0714.2001.301004.x. [DOI] [PubMed] [Google Scholar]
- 21.Wunder JS, Czitrom AA, Kandel R, Andrulis IL. Analysis of alterations in the retinoblastoma gene and tumor grade in bone and soft-tissue sarcomas. J Natl Cancer Inst. 1991;83:194–200. doi: 10.1093/jnci/83.3.194. [DOI] [PubMed] [Google Scholar]
- 22.Benedict WF, Murphree AL, Banerjee A, Spina CA, Sparkes MC, Sparkes RS. Patient with 13 chromosome deletion: evidence that the retinoblastoma gene is a recessive cancer gene. Science. 1983;219:973–975. doi: 10.1126/science.6336308. [DOI] [PubMed] [Google Scholar]
- 23.Castrilli G, Piantelli M, Artese L, Perfetti G, Rubini C, Fioroni M, et al. Expression of hMSH2 and hMLH1 proteins of the human DNA mismatch repair system in ameloblastoma. J oral path & med. 2001;30:305–308. doi: 10.1034/j.1600-0714.2001.300508.x. [DOI] [PubMed] [Google Scholar]
- 24.Mork J, Lie AK, Glattre E, Clark S, Hallmans G, Jellum E, et al. Human papillomavirus infection as a risk factor for squamous-cell carcinoma of the head and neck. N Engl J Med. 2001;344:1125–1131. doi: 10.1056/NEJM200104123441503. [DOI] [PubMed] [Google Scholar]
- 25.Kreimer AR, Clifford GM, Boyle P, Franceschi S. Human papillomavirus types in head and neck squamous cell carcinomas worldwide: a systematic review. Cancer Epidemiol Biomarkers Prev. 2005;14:467–475. doi: 10.1158/1055-9965.EPI-04-0551. [DOI] [PubMed] [Google Scholar]
- 26.Sand L, Jalouli J, Larsson P, Magnusson B, Hirsch J. Presence of human papilloma viruses in intraosseous ameloblastoma. J Oral Maxillofac Surg. 2000;58:1129–1134. doi: 10.1053/joms.2000.9573. [DOI] [PubMed] [Google Scholar]
- 27.Eberhart CG, Argani P. Wnt signaling in human development: beta-catenin nuclear translocation in fetal lung, kidney, placenta, capillaries, adrenal, and cartilage. Pediatri Dev Pathol. 2001;4:351–357. doi: 10.1007/s10024001-0037-y. [DOI] [PubMed] [Google Scholar]
- 28.Kumamoto H, Ooya K. Expression of bone morphogenetic proteins and their associated molecules in ameloblastomas and adenomatoid odontogenic tumors. Oral Dis. 2006;12:163–170. doi: 10.1111/j.1601-0825.2005.01177.x. [DOI] [PubMed] [Google Scholar]
- 29.So F, Daley TD, Jackson L, Wysocki GP. Immunohistochemical localization of fibroblast growth factors FGF-1 and FGF-2, and receptors FGFR2 and FGFR3 in the epithelium of human odontogenic cysts and tumors. J Oral Pathol Med. 2001;30:428–433. doi: 10.1034/j.1600-0714.2001.300708.x. [DOI] [PubMed] [Google Scholar]
- 30.Kumamoto H, Kinouchi Y, Ooya K. Telomerase activity and telomerase reverse transcriptase (TERT) expression in ameloblastomas. J Oral Pathol Med. 2001;30:231–236. doi: 10.1034/j.1600-0714.2001.300407.x. [DOI] [PubMed] [Google Scholar]
- 31.Allsopp RC, Weissman IL. Replicative senescence of hematopoietic stem cells during serial transplantation: does telomere shortening play a role? Oncogene. 2002;21:3270–3273. doi: 10.1038/sj.onc.1205314. [DOI] [PubMed] [Google Scholar]
- 32.Kumamoto H, Ooya K. Immunohistochemical detection of β-catenin and adenomatous polyposis coli in ameloblastomas. J Oral Pathol Med. 2005;34:401–406. doi: 10.1111/j.1600-0714.2005.00328.x. [DOI] [PubMed] [Google Scholar]
- 33.Yamamoto K, Yoneda K, Yamamoto T, Ueta E, Osaki T. An immunohistochemical study of odontogenic mixed tumours. Eur J Cancer Part B: Oral Oncol. 1995;31:122–128. doi: 10.1016/0964-1955(94)00046-7. [DOI] [PubMed] [Google Scholar]
- 34.Sekine J, Kitamura A, Ueno K, Sano K, Inokuchi T, Takahashi H, et al. Cell kinetics in mandibular ameloblastic fibro-odontoma evaluated by bromodeoxyuridine and proliferating cell nuclear antigen immunohistochemistry: case report. Br J Oral Maxillofac Surg. 1996;34:450–453. doi: 10.1016/s0266-4356(96)90106-0. [DOI] [PubMed] [Google Scholar]
- 35.Kumamoto H, Ooya K. Immunohistochemical and ultrastructural investigation of apoptotic cell death in granular cell ameloblastoma. J Oral Pathol Med. 2001;30:245–250. doi: 10.1034/j.1600-0714.2001.300409.x. [DOI] [PubMed] [Google Scholar]
- 36.Zafar MS, Ahmed N. Nano-mechanical evaluation of dental hard tissues using indentation technique. W App Sci J. 2013;28:1393–1399. [Google Scholar]
- 37.Kumamoto H. Detection of apoptosis-related factors and apoptotic cells in ameloblastomas: analysis by immunohistochemistry and an in situ DNA nick end-labelling method. J Oral Pathol Med. 1997;26:419–425. doi: 10.1111/j.1600-0714.1997.tb00242.x. [DOI] [PubMed] [Google Scholar]
- 38.Mitsuyasu T, Harada H, Higuchi Y, Kimura K, Nakamura N, Katsuki T, et al. Immunohistochemical demonstration of bcl-2 protein in ameloblastoma. J Oral Pathol Med. 1997;26:345–358. doi: 10.1111/j.1600-0714.1997.tb00228.x. [DOI] [PubMed] [Google Scholar]
- 39.Kumamoto H, Ohki K, Ooya K. Expression of Sonic hedgehog (SHH) signaling molecules in ameloblastomas. J Oral Pathol Med. 2004;33:185–190. doi: 10.1111/j.0904-2512.2004.00070.x. [DOI] [PubMed] [Google Scholar]
- 40.Sauk JJ, Nikitakis NG, Scheper MA. Are we on the brink of nonsurgical treatment for ameloblastoma? Oral Surg Oral Med Oral Pathol Oral Radiol Endod. 2010;110:68–78. doi: 10.1016/j.tripleo.2010.01.024. [DOI] [PubMed] [Google Scholar]