

Abstract
The change in ancient atmospheric CO2 concentrations provides important clues for understanding the relationship between the atmospheric CO2 concentration and global temperature. However, the lack of CO2 evolution curves estimated from a single terrestrial proxy prevents the understanding of climatic and environmental impacts due to variations in data. Thus, based on the stomatal index of fossilized Metasequoia needles, we reconstructed a history of atmospheric CO2 concentrations from middle Miocene to late Early Pleistocene when the climate changed dramatically. According to this research, atmospheric CO2 concentration was stabile around 330–350 ppmv in the middle and late Miocene, then it decreased to 278–284 ppmv during the Late Pliocene and to 277–279 ppmv during the Early Pleistocene, which was almost the same range as in preindustrial time. According to former research, this is a time when global temperature decreased sharply. Our results also indicated that from middle Miocene to Pleistocene, global CO2 level decreased by more than 50 ppmv, which may suggest that CO2 decrease and temperature decrease are coupled.
Introduction
Carbon dioxide (CO2) is an important greenhouse gas that influences the surface temperature of the Earth [1]. The 5th report of IPCC concluded [2] that the present positive radiative forcing is unequivocally caused by anthropogenic increases in atmospheric CO2 concentration and that it influences the climate [3,4]. Estimating the impact of high CO2 concentration on global environmental systems is the first step to propose solutions for the present global climate change. This impact can be unraveled by a better understanding of the relationship between the paleo-atmospheric CO2 concentration (paleo-[CO2]atm) and ancient climate change.
A lot of research has involved the estimation of paleo-[CO2]atm to understand the correlation between CO2 and global warming. To obtain the paleo-[CO2]atm values three major approaches have been used: (1) geochemical modeling (GCS) [5–7], (2) composition measurements of air trapped in ice cores [8], and (3) various proxies (reviewed in [9]). Geochemical modeling (GCS) can reconstruct paleo-[CO2]atm, but for long geological time scales its resolution cannot be fine enough to show the details of paleo-[CO2]atm fluctuation [10]. Ice core analysis is the most reliable method to measure paleo-[CO2]atm directly, but is only applicable after 0.8 Ma [8]. Several CO2 proxies have been used to estimate paleo-[CO2]atm, such as the carbon isotope composition of phytoplankton, the boron (B) isotope composition of fossil foraminifera, the carbon isotope composition of carbonates in paleosol, and the stomatal parameters of fossil leaves [11]. High resolution records for CO2 can be obtained from marine sediments with the two former proxies, but these do not directly show the paleo-[CO2]atm. The latter two proxies are terrestrial-based proxies that reflect paleo-[CO2]atm directly, although they rarely provide continuous paleo-[CO2]atm records for a long geological time. Therefore, while there is a consensus on the general tendency of the Cenozoic paleo-[CO2]atm changes, the estimated paleo-[CO2]atm values vary greatly [9]. To understand the paleoclimatic system, it is important to reduce uncertainties in the relationships between paleo-[CO2]atm and past climate [12].
Stomatal parameters (SI (stomatal index) and SD (stomatal density)) are reliable proxies to estimate paleo-[CO2]atm. In particular, SI can provide a robust indicator of terrestrial paleo-[CO2]atm as it is independent of other environmental parameters, such as soil moisture supply, atmospheric humidity and temperature [13]. Many studies have already used the SI of different taxa to estimate paleo-[CO2]atm, such as Metasequoia Miki ex Hu et Cheng [12,14], Ginkgo Linn. [15,16], Quercus Linn. [17,18], Laurus Linn. [17,19,20], Platanus Linn. [17,21], and Typha Linn. [22]. As the relationship between the SI and paleo-[CO2]atm is species-specific even within a single family [23] and the response sensitivities to CO2 change are different in various taxa [24], it is necessary to select a single modern taxon that has survived for an extended period to reconstruct atmospheric CO2 over a long geological time.
Metasequoia has exhibited an evolutionary stasis since its appearance in the Late Cretaceous [25], and fossilized Metasequoia can be considered to be conspecific with modern Metasequoia based on the morphology, biochemistry and inferred physiology [26]. Therefore, the paleo-[CO2]atm changes over a long geological time can be determined from a correlation between the SI of Metasequoia needles and the paleo-[CO2]atm concentration [14].
In this study, we use Metasequoia needles from seven localities in China and Japan to reconstruct continuous terrestrial paleo-[CO2]atm changes from the middle Miocene to Pleistocene. Based on the reconstructed paleo-[CO2]atm curve, we discuss the interaction between paleo-[CO2]atm evolution and global environment change since the middle Miocene.
Materials and Methods
Materials
The fossilized needles of Metasequoia (Fig 1) were collected from one locality in SW China (Sanzhangtian) and six localities in central Japan (Kumagaya, Sennan, Hachioji, Konan, Tokamachi, and Ikoma sites) (Fig 2, Table 1). We confirm that our field study did not involve endangered or protected species and none of the localities which provided samples for this study are in protected areas. The Sanzhangtian locality belongs to the National land of the People's Republic of China, and the Land and Resources Bureau of Zhenyuan County gave permission to collect fossils from this locality. The Japanese sites: Kumagaya, Hachioji, Konan, and Tokamachi are on valley floors which are public space, so no permission was required to conduct sampling. The Sennan and Ikoma sites belong to private owners, who gave permission for sampling.
Fig 1. Fossilized Metasequoia from Tokamachi and Kumagaya sites.

Fossilized Metasequoia branchlet and needles from Tokamachi (a) and Kumagaya (b) as examples to show the megafossils of Metasequoia used in this research, compare with a modern Metasequoia branchlet (c). White arrows in (a) indicate the branchlet.
Fig 2. Localities where fossilized Metasequoia were obtained.

Locality map (a) showing the seven fossil sites in China and Japan. Enlarged map (b) illustrating the central area of Japan showing the position of the six localities in Japan: Kumagaya, Sennan, Konan, Hachioji, Tokamachi and Ikoma. Different colors identify the different ages of the localities (Red: Miocene; Green: Pliocene; Blue: Pleistocene).
Table 1. Metasequoia samples used for reconstructing paleo-CO2 including fossil sites, ages, latitude, and longitude.
| Fossil site | Locality | Latitude/ Longitude | Geologic setting | Epoch | Absolute age | Dating method | Voucher specimens # | Remark | Reference |
|---|---|---|---|---|---|---|---|---|---|
| Sanzhangtian | Yunnan, China | 24°06′ N, 101°13′ E | Dajie Formation | middle Miocene | 10–16 Ma | Stratigraphic study | SZT077,SZT156,SZT115,SZT127,SZT123,SZT126 | [27–29] | |
| Kumagaya | Saitama Prefecture, Japan | 36°08′ N, 139°18′ E | Yagii Formation in the Matsuyama Group | early late Miocene | 9–10 Ma | Zircon fission track dating | YJ003,YJ005 | Includes marine bed | [30] |
| Sennan | Osaka Prefecture, Japan | 34°24' N, 135°28' E | Lower than the Habutaki I Tephra, Osaka Group | Late Pliocene | 2.8–3.0 Ma | Magnetostratigraphy and calcareous nanoplankton stratigraphy | FT001 | Included in sediments in fluvial backmarsh | [31–33] |
| Hachioji | Tokyo, Japan. | 35° 40′ N, 139°18′ E | Kasumi Formation (below the Gauss and Matuyama Chron boundary) | Late Pliocene | 2.6–2.7 Ma | Magnetostratigraphy and calcareous nanoplankton stratigraphy | BQC001 | Includes marine bed | [34,35] |
| Konan | Shiga Prefecture, Japan | 34°59′ N, 136°6′ E | Horizon correlated with the Kamide I tephra bed in Kobiwako Group (just below the Gauss and Matuyama Chron boundary) | Late Pliocene | 2.6 Ma | Magnetostratigraphy and calcareous nanoplankton stratigraphy | SG001,SG002 | Included in sediments in fluvial backmarsh | [36] |
| Tokamachi | Niigata Prefecture, Japan. | 37°07'N, 138°48'E | Middle part of the Uonuma Group lower part of Olduvai paleomagnetic chron | middle Early Pleistocene | 1.85 Ma | Magnetostratigraphy and calcareous nanoplankton stratigraphy | 156u01 | Includes marine bed | [32,37,38] |
| Ikoma | Nara Prefecture, Japan | 34°44'N, 135°43'E | Peat layer just below the Ma 2 Marine Clay bed (MIS 25) in the Osaka Group | latest Early Pleistocene | 0.95 Ma | Magnetostratigraphy and calcareous nanoplankton stratigraphy | NR001 | Includes marine bed | [39,40] |
Metasequoia fossils had previously been reported from all the fossil localities. Their ages were estimated based on stratigraphic studies (Sanzhangtian site), zircon fission-track methods (Kumagaya site), and regional stratigraphic correlation using magnetostratigraphy and calcareous nanoplankton stratigraphy (Sennan, Hachioji, Konan, Tokamachi, and Ikoma sites) (Table 1). For the samples, two were from the Miocene (Sanzhangtian and Kumagaya), three from the upper Pliocene (Sennan, Hachioji, and Konan), and two from the lower Pleistocene (Tokamachi and Ikoma) (Table 1). At least six different needles from different branchlets were used in the studies from each site, and the exact amount depends on the total amount of materials at each fossil site (Table 2).
Table 2. Sample size of localities.
| Fossil site | Epoch | Sample size (no.) |
|---|---|---|
| Sanzhangtian | middle Miocene | 25 |
| Kumagaya | early late Miocene | 11 |
| Sennan | Late Pliocene | 6 |
| Hachioji | Late Pliocene | 8 |
| Konan | Late Pliocene | 7 |
| Tokamachi | middle Early Pleistocene | 17 |
| Ikoma | latest Early Pleistocene | 7 |
Voucher specimens from the Sanzhangtian, Kumagaya, Hachioji, Konan, and Tokamachi sites are housed in the Herbarium of Kunming Institute of Botany (KUN), Chinese Academy of Science. Specimens from the Sennan and Ikoma sites are housed in the Graduate School of Horticulture, Chiba University, Japan.
Methods
Pretreatment of the fossilized needles
To remove the inorganic compounds adhering to the fossilized needles, the material was first immersed in 10%–25% Hydrochloric Acid (HCl) for two hours, then in 40% Hydrofluoric Acid (HF) for 12 hours, and in 10%–25% HCl for at least one hour. The needles were then rinsed with distilled water and divided into three parts, and the central piece (when available) used to obtain the cuticle.
Cleaned cuticular membrane maceration
For the material from the Sanzhangtian, Kumagaya, and Konan sites, we followed the methods of Kerp [41] to isolate the lower cuticle of the fossilized needles. (1) The specimens were first macerated with 70% Nitric acid (HNO3) for between a few minutes to an hour until they turned yellowish-brown. (2) Once it had been rinsed with distilled water several times, (3) the upper and the lower epidermis were separated using a needle. (4) Then, the epidermis was treated with a 3%–5% Sodium Hypochlorite (NaClO) solution for around 10 minutes to remove the remnants of the mesophyll, vascular bundle, hypodermal layer, and epidermal cell walls. (5) According to the state of the material, 5%-10% Aqueous Ammonia (NH3·H2O) or 30% Hydrogen Peroxide (H2O2) can be used instead of the 3%–5% NaClO. (6) Finally glycerol was used to mount the separated cuticles for observation.
Cuticle observation and photography
The separated cuticles of the material from the Sanzhangtian, Kumagaya, and Konan sites were observed using a transmitted light microscope (Zeiss Axio Imager A2) and photographed with a digital camera (Zeiss AxioCam MRc). For the materials from the Sennan, Hachioji, Tokamachi, and Ikoma sites, pretreated fossilized leaves were mounted with water on slides and the lower sides of the needles were directly scanned by a confocal laser scanning microscope (Zeiss LSM710, Imager. Z2, Ar Lasser 488nm). Each field-of-view was larger than 0.03mm2 [42]. Photoshop (version CS6, Adobe Systems; Mountain View, CA) was used to merge 6–12 serial images that were taken of the same area but at different focal levels.
Measurement of SI and paleo-[CO2]atm concentration
Image J (1.43μ, Wayne Rasband, http://rsb.info.nih.gov/ij/) was used to calculate the number of epidermal cells and stomatal complexes (stomatal pore + guard cells). Then, the SI was calculated using Eq 1 [43].
| Equation 1 |
The SI data were used to estimate the paleo-[CO2]atm from the middle Miocene to Pleistocene by using the species-specific, nonlinear negative correlation between atmospheric CO2 partial pressure and SI (Eq 2) based on Royer et al. [14].
| Equation 2 |
The significant differences between the mean variance of the SI from different ages were statistically tested using the two tailed one-way ANOVA with the “LSD” option in IBM SPSS Statistics (Version 20.0).
Results
Fossilized Metasequoia needles from the early late Miocene Kumagaya site had the lowest SI value (SI = 9.80 ± 0.65) and those from the middle Miocene Sanzhangtian site had the second lowest (SI mean = 10.43 ± 0.99). Their calculated paleo-[CO2]atm values were 351 ± 24.8 ppmv and 334 ± 24.8 ppmv, respectively (Fig 3, Table 3, for more details see S1 Table).
Fig 3. Lower cuticles of the Metasequoia needle samples from different localities.

Lower cuticles of Metasequoia needles from a: Sanzhangtian; b: Ikoma; c: Kumagaya; d: Tokamachi; e: Konan; f: Sennan; and g: Hachioji. (Scale Bar = 100μm)
Table 3. Fossilized Metasequoia stomatal index and paleo-[CO2]atm concentration estimates during Cenozoic.
| Fossil site | Epoch | SI (%) | paleo-[CO2]atm (ppmv) | ||||
|---|---|---|---|---|---|---|---|
| Mean ± sd | Max | Min | Mean ± sd | Max | Min | ||
| Sanzhangtian | middle Miocene | 10.4±0.99 | 12.5 | 9.09 | 334±24.9 | 382 | 298 |
| Kumagaya | early late Miocene | 9.80±0.65 | 11.0 | 8.97 | 351±24.8 | 392 | 317 |
| Sennan | Late Pliocene | 17.1±2.30 | 19.8 | 14.8 | 280±5.16 | 285 | 274 |
| Hachioji | Late Pliocene | 17.2±1.65 | 19.4 | 14.8 | 279±3.74 | 285 | 275 |
| Konan | Late Pliocene | 15.2±1.70 | 18.4 | 13.7 | 285±5.15 | 290 | 276 |
| Tokamachi | middle Early Pleistocene | 17.1±2.52 | 21.7 | 13.3 | 280±6.23 | 293 | 272 |
| Ikoma | latest Early Pleistocene | 17.9±1.90 | 20.2 | 15.5 | 278±3.86 | 282 | 273 |
The SI of the Pliocene and Pleistocene samples were higher (SI mean = 15.2–17.9) than the SI of the Miocene samples. The SI of the samples from the middle Early Pleistocene Ikoma site had the highest SI value (SI = 17.9 ± 1.9) and give out the lowest CO2 level of 278 ± 3.86 ppmv. SI of the fossilized leaves from the Sennan, Hachioji, Konan, and Tokamachi sites were 17.1, 17.2, 15.2, and 17.1, respectively. The reconstructed paleo-[CO2]atm from the Pliocene and Pleistocene samples in the Sennan, Hachioji, Konan, Tokamachi, and Ikoma sites were 280 ± 5.16, 279 ± 3.74, 285 ± 5.15, 280 ± 6.23, and 278 ± 3.86 ppmv, respectively (Fig 3, Table 3, for more details see S1 Table).
The significant differences between the mean variance of the stomatal index from different fossil localities were statistically tested (F = 54.016, p<0.001) by one-way ANOVA with the “LSD” option in SPSS Statistics (Version 20.0). The result showed there was no significant difference between the SI data from Sanzhangtian locality (middle Miocene) and Kumagaya locality (late Miocene), but the SI data of these two localities were significantly different from the SI data from late Pliocene and Pleistocene localities. SI data of Konan locality (Late Pliocene) was significantly different from all other localities, but no significant difference has been detected among Sennan (Late Pliocene), Hachioji (Late Pliocene), Tokamachi (middle Early Pleistocene) and Ikoma localities (latest Early Pleistocene) (Table 4).
Table 4. Mean difference of the least significant different (LSD) on stomatal index of fossil localities.
| Locality | Sanzhangtian | Kumagaya | Sennan | Hachioji | Konan | Tokamachi |
|---|---|---|---|---|---|---|
| Kumagaya | 0.63 | |||||
| Sennan | -6.70 *** | -7.33 *** | ||||
| Hachioji | -6.80 *** | -7.42 *** | -0.10 | |||
| Konan | -4.80 *** | -5.42 *** | 1.90 * | 2.00 * | ||
| Tokamachi | -6.68 *** | -7.31 *** | 0.02 | 0.12 | 1.88 * | |
| Ikoma | -7.42 *** | -8.05 *** | -0.73 | -0.63 | -2.63 ** | -0.75 |
The sign of the significance is indicated as
* p<0.05
** p<0.01
*** p<0.001.
Discussion
Middle and late Miocene paleo-[CO2]atm change
The paleo-[CO2]atm changes reconstructed in previous research generally indicate a peak during the middle Miocene Climatic Optimum (MCO; 17–15 Ma) [44] and a decline during the later stage of the middle Miocene (ca. 15–11.5Ma), although the reconstructed paleo-[CO2]atm values and timing of fluctuation were different among proxies (Fig 4B). The most prominent fluctuation was exhibited in the paleosol carbonate records, which showed a spike (ca. 800 ppmv) at 15.6 Ma, drop to 116–310 ppmv at 14.7–13.8 Ma, and increase to 433–519 ppmv around 12.8–13.1 Ma [45]. The stomatal records from fossilized Quercus leaves [23] also indicated a prominent change from the highest value (469–555 ppmv) at 15.7±0.7Ma to the lower value at 13.0 Ma (ca.290 ppmv) and 11.6 Ma (ca.330 ppmv) during the late middle Miocene. Additionally, the stomatal proxies from North America indicate lower paleo-[CO2]atm values and moderate changes during the earlier stage of the middle Miocene: 396 ppmv from Ginkgo leaves at ca. 16.5 Ma and 310–316 ppmv from Metasequoia needles around 15.2–15.3 Ma [14].
Fig 4. Trend of paleo-[CO2]atm during late Cenozoic.
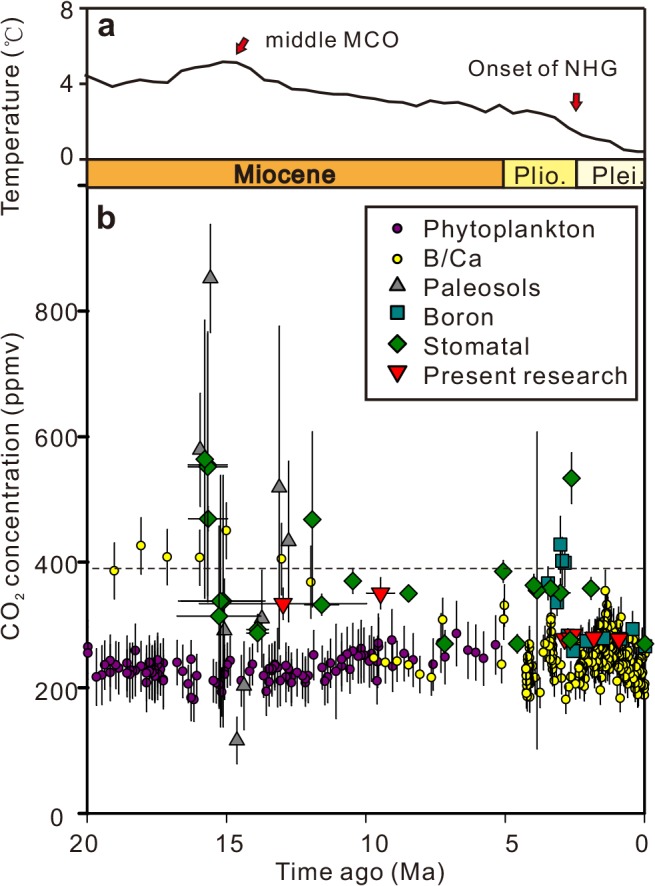
(a) Deep-sea temperatures estimated from δ18O since 20 Ma [46]; (b) atmospheric CO2 reconstructed from terrestrial and marine proxies following recent revisions (S2 Table). Vertical error bars: standard deviation of paleo-[CO2]atm values, and horizontal error bars: standard deviation of materials’ age. The current atmospheric CO2 concentration (390 ppmv) is indicated by the horizontal dashed line.
In general, the values of the middle Miocene [CO2]atm estimated from marine proxies are lower than those from terrestrial records. Boron/Calcium (B/Ca) ratios of surface-dwelling foraminifera give a paleo-[CO2]atm of ca. 420 ppmv during the MCO that declined gradually to ca. 200 ppmv in the earliest late Miocene [47]. B isotope (δ11B)-based paleo-[CO2]atm from ODP761 changed from ca. 400 ppmv in the MCO to ca. 280 ppmv in the late middle Miocene [48]. A stable paleo-[CO2]atm curve with slight changes around 210 ppmv from the MCO to late Miocene was drawn based on phytoplankton δ13C alkene analysis [49,50]. The paleo-[CO2]atm value (334 ppmv) reconstructed from the fossilized leaves of the middle Miocene Sanzhangtian site was similar to the late middle Miocene values based on Quercus leaves [51] and between the results based on Ginkgo (16.5 Ma) and Metasequoia (15.2–15.3 Ma) leaves in the early middle Miocene [14].
The late Miocene stomatal data based on fossilized Quercus exhibited a decreasing paleo-[CO2]atm tendency: ca. 370 ppmv at ca. 10.5 Ma, ca. 350 ppmv at ca. 8.5 Ma, and ca. 270 ppmv at ca. 7.2 Ma [51]. This was related to climatic cooling in the later late Miocene [18,52]. When using B/Ca [53] and phytoplankton [49,50] from marine proxies, they showed fluctuating values that were mostly less than 300 ppmv (Fig 4B). The estimated paleo-[CO2]atm values for 10–9 Ma (351 ppmv) from this work are almost the same as the value from ca. 8.5 Ma from Quercus leaves [18]. Our data showed little change between the middle Miocene (334 ppmv) and the early late Miocene (351 ppmv) that confirmed the stable paleo-[CO2]atm condition during this time as indicated by the phytoplankton record [49,50].
Late Pliocene to Pleistocene paleo-[CO2]atm change
In most of the previous research, paleo-[CO2]atm values are distributed between 200 and 400 ppmv during the Pliocene to Pleistocene (Fig 5A). B/Ca and B data have been used to determine the paleo-[CO2]atm of this period, as there is a lack of data from the stomatal method. The paleo-[CO2]atm curve based on B/Ca from surface-dwelling foraminifera exhibited a peak of ca. 300 ppmv at ca. 3.4 Ma, this decreased to 181 ppmv at ca. 2.9 Ma, and then increased to 332 ppmv at ca. 1.4 Ma [47]. The downward shift in its fluctuation range was observed in the Early Pleistocene (Fig 5A), and the lowest value of 188 ppmv was recorded in the last glacial maximum (0.02Ma) [47]. The paleo-[CO2]atm recorded in the B isotopes indicates a higher level than that in B/Ca record during the Late Pliocene, that is, ca. 340 ppmv at ca. 3.4 Ma and ca. 400 ppmv at ca. 3.0 Ma [54]. However, it decreases to the same level (ca. 270 ppmv) as the B/Ca record in the late Late Pliocene and Early Pleistocene (ca. 2.8–1.0 Ma) [54]. The paleo-[CO2]atm level estimated from Quercus [18] and Cupressaceae [55] stomata indicates a higher level (ca. 350 ppmv) during the early Late Pliocene (3–3.4 Ma) and a lower value (276 ppmv) at 2.7 Ma. While paleo-[CO2]atm based on the SI of Typha at the Plio-Pleistocene boundary (2.65 Ma) exhibits a much higher value (534 ppmv) than the other results [22].
Fig 5. Reconstructed paleo-[CO2]atm of Pliocene to Pleistocene compared with reconstructed paleo-temperature and benthic δ18O record.

(a) Reconstructed paleo-[CO2]atm based on terrestrial and marine proxies following recent revisions (S2 Table) along with our data. Vertical error bars: standard deviation of paleo-[CO2]atm values, and horizontal error bars: standard deviation of ages of materials. The current atmospheric CO2 concentration (390 ppmv) is indicated by the horizontal dashed line. (b) SST records for the last 3.5 Ma from southern South China Sea [56]. (c) Global oxygen isotopes of benthic foraminifera shells [57]. The vertical color bands in (a), (b), and (c) indicate the periods considered by this research, and same period is marked by the same color.
Our data showed that the paleo-[CO2]atm was maintained in the range between 280 and 285 ppmv in the Pliocene and Pleistocene (Figs 4B and 5A), which is about 150 ppmv lower than the results estimated from B isotopes [54], and about 70 ppmv higher than the results estimated from the B/Ca proxy [47]. Our data are consistent with the results estimated from Quercus stomata [18], but are much lower than the data estimated from Typha from sediment at the Plio-Pleistocene boundary [22]. While different proxies [22,47,54] have recorded fluctuations accompanying climate changes (Fig 5B), the paleo-[CO2]atm value of this study stabilized at around 280 ppmv. Seiki et al. concluded that the Pliocene CO2 levels determined by numerous methods agreed well with each other [9,54]. The present research suggests that some disagreements still remain in the results between our stomatal data and B, B/Ca records in the Pliocene, while the Pleistocene proxies give more consistent CO2 levels (than the Pliocene).
Paleo-[CO2]atm change and late Cenozoic climatic deterioration
The overall climate cooling reconstructed for the past 20 Ma has generally been attributed to changes in CO2 concentration in the atmosphere [46,58]. According to the marine oxygen isotope record, global temperature peaked at around 16 Ma (middle MCO) (Fig 4A), and the later part of the middle Miocene is characterized by climate cooling with expansion of the East Arctic ice sheet [59,60]. However, the middle Miocene paleo-[CO2]atm reconstructed in this study (around 334 ppmv) was just slightly lower than the present level, which was also the level maintained during the late Miocene (around 354 ppmv). That means that before the global temperature decrease, paleo-[CO2]atm had already achieved a stable low level. The Miocene paleo-[CO2]atm estimated based on alkenones also showed that paleo-[CO2]atm was similar during middle Miocene and late Miocene [61]. The δ13C record from foraminifera and B/Ca ratios in the foraminifera suggest that paleo-[CO2]atm decreases were apparently synchronous with major episodes of glacial expansion during the middle Miocene [53,62, 63], but this synchronization was not observed in our data. This study supports the view that Miocene climate change was not only influenced by paleo-[CO2]atm changes, but also by increases in seasonality and ocean circulation changes [50,64,65], and these accelerated the cooling in the late middle Miocene that also acted to decrease the paleo-[CO2]atm [62]. Also, climate sensitivity to paleo-[CO2]atm may have been greater than previously thought [66]. The impact of high latitude vegetation on Earth’s albedo may have also played an important role in the Earth’s energy budget in the Miocene [67].
After termination of the mid-Pliocene warmth at ca. 2.9 Ma, cooling trends continued until the onset of major expansion of the Northern Hemisphere ice sheet at ca. 2.7 Ma, which culminated at ca. 2.5 Ma in the earliest Pleistocene [68–70]. However, present results show that the lower paleo-[CO2]atm level started around 2.8–3.0 Ma and lasted until the late Early Pleistocene. Therefore, we consider that the transition to the icehouse world was possibly induced by a decrease of the paleo-[CO2]atm, which already dropped to their lowest levels during the complete Cenozoic before the major expansion of the Northern Hemisphere ice sheets. During the Pliocene to Pleistocene, our data are very stable, but the global temperature estimated from the marine oxygen isotope record [56,57] shows drastic fluctuations (Fig 5). However, our middle and late Miocene data are significantly higher than our Pliocene and Pleistocene data. The oxygen isotope record confirms that the temperature in the Pliocene and Pleistocene was much lower than that of the middle and late Miocene [44,46]. Therefore, we can conclude that the decrease of paleo-[CO2]atm level is coupled with temperature decrease during middle Miocene to Pleistocene.
Conclusions
We used the stomatal index of Metasequoia Miki ex Hu et Cheng as a proxy to reconstruct the paleo-[CO2]atm evolution from the middle Miocene to late Early Pleistocene for the first time. Our results indicate that: (1) From middle to late Miocene the atmospheric CO2 level stabilized around 350 ppmv which is slightly lower than today. (2) The CO2 level during the Pliocene to Pleistocene was similar to the pre-industrial level and no fluctuation can be detected by this research. (3) The Pleistocene CO2 level estimated by different proxies agree well with each other. (4) From middle Miocene to Pleistocene, when the global temperature decreased sharply, the global CO2 level decreased by more than 50 ppmv, which may suggest that CO2 decrease and temperature decrease are coupled.
Supporting Information
(DOC)
(DOC)
Acknowledgments
The authors would like to thank the Central Laboratory of Xishuangbanna Tropical Botanical Garden, CAS for providing cuticle observing equipment (light microscope, confocal laser scanning microscope, and SEM) and advice; Ito A, Kobayashi H, Otao T, and Kawakami T for collecting fossil samples in Japan; Zhang ST for helping to analyze the stratigraphical information; Spicer RA and Liang XQ for suggestions and comments on the manuscript; Li SF and Huang J for advice about the use of the ArcGIS software; Ferguson DJ for polishing the manuscript; and Jia LB for the picture of modern Metasequoia branchlet. We are grateful to our academic editor Wang Q and reviewers Quan C, Gaurav Srivastava BSIP,.Kürschner WM and two anonymous reviewers for their constructive suggestions and thorough comments on the manuscript. This is a contribution to NECLIME (Neogene Climate Evolution in Eurasia).
Data Availability
All relevant data are within the paper and its Supporting Information files.
Funding Statement
The work was supported by the following: 1. 973 programme of Ministry of Science and Technology of the People's Republic of China (2012CB821901), Receiver: ZZK, URL: http://www.973.gov.cn/Default_3.aspx; 2. National Natural Science Foundation of China (NSFC, No. 1300188), Receiver: WL, URL: http://www.nsfc.gov.cn/; 3. Chinese Academy of Sciences 135 program (XTBG-F01), Receiver: ZZK, URL: http://www.xtbg.ac.cn/yisanwu/; and 4. State Key Laboratory of Palaeobiology and Stratigraphy (Nanjing Institute of Geology and Palaeontology, CAS) (No. 123108), Receiver: WL, URL: http://159.226.74.1/site/list?catid=25&page=3&size=12.
References
- 1. Lacis AA, Schmidt GA, Rind D, Ruedy RA. Atmospheric CO2: principal control knob governing Earth's temperature. Science. 2010; 330: 356–359. 10.1126/science.1190653 [DOI] [PubMed] [Google Scholar]
- 2. Stocker TF, Qin D, Plattner GK, Tignor MMB, Allen SK, Boschung J et al. Climate Change 2013: The Physical Science Basis Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. 1st ed. Cambridge, United Kingdom and New York, NY, USA: Cambridge University Press; 2013. [Google Scholar]
- 3. Raupach MR, Canadell JG. Carbon and the Anthropocene. Curr Opin Environ Sustain. 2010; 2: 210–218. [Google Scholar]
- 4. Peng J, Dan L, Huang M. Sensitivity of global and regional terrestrial carbon storage to the direct CO2 effect and climate change based on the CMIP5 model intercomparison. PLoS ONE. 2014; 9: e95282 10.1371/journal.pone.0095282 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Berner RA. GEOCARB II: a revised model of atmospheric CO2 over Phanerozoic time. American Journal of Science. 1994; 294: 56–91. [DOI] [PubMed] [Google Scholar]
- 6. Berner RA. GEOCARBSULF: A combined model for Phanerozoic atmospheric O2 and CO2 . Geochim Cosmochim Acta. 2006; 70: 5653–5664. [Google Scholar]
- 7. Berner RA, Kothavala Z. GEOCARB III: A revised model of atmospheric CO2 over phanerozoic time. American Journal of Science. 2001; 301: 182–204. [Google Scholar]
- 8. Monnin E, Indermühle A, Dällenbach A, Flückiger J, Stauffer B, Stocker TF et al. Atmospheric CO2 Concentrations over the Last Glacial Termination. Science. 2001; 291: 112–114. [DOI] [PubMed] [Google Scholar]
- 9. Beerling DJ, Royer DL. Convergent Cenozoic CO2 history. Nat Geosci. 2011; 4: 418–420. [Google Scholar]
- 10. Royer DL. CO2-forced climate thresholds during the Phanerozoic. Geochim Cosmochim Acta. 2006; 70: 5665–5675. [Google Scholar]
- 11. Solomon S, Qin D, Martin M, Marquis M, Averyt K, Tignor MMB et al. Climate Change 2007: The Physical Science Basis. Contribution of Working Group I to the Forth Assessment Report of the Intergovernmental Panel on Climate Change. 1st ed. Cambridge, United Kingdom and New York, NY, USA: Cambridge University Press; 2007. [Google Scholar]
- 12. Doria G, Royer DL, Wolfe AP, Fox A, Westgate JA, Beerling D. Declining Atmospheric CO2 during the Late Middle Eocene Climate Transition. American Journal of Science. 2011; 311: 63–75. [Google Scholar]
- 13. Beerling DJ. Stomatal density and index: Theory and application In: Jones TP, Rowe NP, editors. Fossil plants and spores: modern techniques. London: Geological Society of London; 1999. pp. 251–256. [Google Scholar]
- 14. Royer DL, Wing SL, Beerling DJ, Jolley DW, Koch PL, Hickey L et al. Paleobotanical evidence for near present-day levels of atmospheric CO2 during part of the tertiary. Science. 2001; 292: 2310–2313. [DOI] [PubMed] [Google Scholar]
- 15. Quan C, Sun CL, Sun YW, Sun G. High resolution estimates of paleo-CO2 levels through the Campanian (Late Cretaceous) based on Ginkgo cuticles. Cretaceous Research. 2009; 30: 424–428. [Google Scholar]
- 16. Smith RY, Greenwood DR, Basinger JF. Estimating paleoatmospheric pCO2 during the Early Eocene Climatic Optimum from stomatal frequency of Ginkgo, Okanagan Highlands, British Columbia, Canada. Palaeogeogr Palaeoclimatol Palaeoecol. 2010; 293: 120–131. [Google Scholar]
- 17. Grein M, Oehm C, Konrad W, Utescher T, Kunzmann L, Roth-Nebelsick A. Atmospheric CO2 from the late Oligocene to early Miocene based on photosynthesis data and fossil leaf characteristics. Palaeogeogr Palaeoclimatol Palaeoecol. 2013; 374: 41–51. [Google Scholar]
- 18. Kürschner WM, Burgh J, Visscher H, Dilcher DL. Oak leaves as biosensors of late Neogene and early Pleistocene paleoatmospheric CO2 concentrations. Mar Micropaleontol, 1996; 27: 299–312. [Google Scholar]
- 19. Sun B, Ding S, Wu J, Dong C. Carbon isotope and stomatal data of Late Pliocene Betulaceae leaves from SW China: implications for palaeoatmospheric CO2-levels. Turkish Journal of Earth Sciences, 2012; 21: 237–250. [Google Scholar]
- 20. Wagner F, Bohncke SJ, Dilcher DL, Kürschner WM, Geel B, Visscher H. Century-scale shifts in early Holocene atmospheric CO2 concentration. Science. 1999; 284: 1971–1973. [DOI] [PubMed] [Google Scholar]
- 21. Roth-Nebelsick A, Oehm C, Grein M, Utescher T, Kunzmann L, Friedrich JP et al. Stomatal density and index data of Platanus neptuni leaf fossils and their evaluation as a CO2 proxy for the Oligocene. Rev Palaeobot Palynol. 2014; 206: 1–9. [Google Scholar]
- 22. Bai YJ, Chen LQ, Ranhotra PS, Wang Q, Wang YF, Li CS. Reconstructing atmospheric CO2 during the Plio-Pleistocene transition by fossil Typha . Glob Chang Biol. 2014; 21: 874–881. 10.1111/gcb.12670 [DOI] [PubMed] [Google Scholar]
- 23. Kürschner WM, Kvacek Z, Dilcher DL. The impact of Miocene atmospheric carbon dioxide fluctuations on climate and the evolution of terrestrial ecosystems. PNAS. 2008; 105: 449–453. 10.1073/pnas.0708588105 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Royer DL. Stomatal density and stomatal index as indicators of paleoatmospheric CO2 concentration. Rev Palaeobot Palynol. 2001; 114: 1–28. [DOI] [PubMed] [Google Scholar]
- 25. Basinger JF. The Vegetative Body of Metasequoia-Milleri from the Middle Eocene of Southern British-Columbia. Can J Bot. 1981; 59: 2379–2410. [Google Scholar]
- 26. LePage BA, Williams CJ, Yang H. The geobiology and ecology of Metasequoia Dordrecht: Springer; 2005. [Google Scholar]
- 27. Ge HR, Li DY. Cenozoic coal-bearing basins and coal-forming regularity in West Yunnan Kunming: Yunnan Science and Technology Press; 1999. [Google Scholar]
- 28. Zhang QQ, Ferguson DK, Mosbrugger V, Wang YF, Li CS. Vegetation and climatic changes of SW China in response to the uplift of Tibetan Plateau. Palaeogeogr Palaeoclimatol Palaeoecol. 2012; 363: 23–36. [Google Scholar]
- 29.Wang L, Zhou ZK, Xing YW, Su T, Jacques FMB, Liu YS. Miocene Metasequoia from Yunnan, southwest China and its biological implications. Japanese Journal of Palynology (Special Issue: Abstract Issue for the Joint Meeting of 13th International Palynological Congress and 9th International Organisation of Palaeobotany Conference). 2012; 58: 251.
- 30. Kobayashi M., Saito T., Okitsu S. Zircon fission-track ages of the Miocene Yagii Formation, Saitama Prefecture, central Japan, and their palaeoecological significance. The journal of the Geological Society of Japan. 2011; 117 632–636. [Google Scholar]
- 31. Itihara M, Ichikawa K, Yamada N. Geology of the Kishiwada district with geological sheet map at 1:50,000 Tsukuba: Geological Survey of Japan; 1986. [Google Scholar]
- 32. Satoguchi Y, Nagahashi Y. Tephrostratigraphy of the Pliocene to Middle Pleistocene Series in Honshu and Kyushu Islands, Japan. Island Arc. 2012; 21: 149–169. [Google Scholar]
- 33. Tomita Y, Kurokawa K. A widespread volcanic ash layer of about 2.7 Ma in central Japan: correlation of the Habutaki I (Osaka Group), the MT2 (Himi Group) and the Arg-2 (Nishiyama Formation) ash layers. Journal of the Geological Society of Japan. 1999; 105: 63–71. [Google Scholar]
- 34. Horiuchi J. Neogene Flora of the Kanto District. Science Reports of the Institute of Geoscience Geological Sciences, Tsukuba University, Section B, Geological Sciences. 1996; 17: 109–208. [Google Scholar]
- 35. Kimura T, Ohana T, Yoshiyama H. Fossil plants from the Tama and Azuyama Hills, Southern Kwanto, Japan. Transactions and Proceedings of the Paleontological Society of Japan. 1984; 122: 87–104. [Google Scholar]
- 36. Satoguchi Y, Yamakawa C and Takahashi K. The old and newly defined Pliocene-Pleistocene boundary sites of the Kobiwako Group, central Japan. Journal of Geological Society. 2012; 118: 70–78. [Google Scholar]
- 37. Niigata Fossil Plant Research Group and Niigata Pollen Research Group. Plant megafossils and pollen fossils from the Uonuma Group, Niigata Prefecture, central Japan In: Uonuma Hills Collaborative Research Group, editor. The Uonuma Group. Tokyo: The Association for the Geological Collaboration in Japan; 1983. [Google Scholar]
- 38. Yanagisawa Y, Kayahara K, Suzuki Y, Uemura T, Kodama K, Kato T. Geology of the Tokamachi District with geological sheet map at 1:50,000 Tsukuba: Geological Survey of Japan; 1985. [Google Scholar]
- 39. Mitamura M. Stratigraphy and Geologic Structure of the Osaka Group (Pliocene and Pleistocene) in Keihanna Hills, Kinki District, Japan. The Quaternary Research. 1992; 31: 159–177. [Google Scholar]
- 40. Yoshikawa S, Mitamura M. Quaternary stratigraphy of the Osaka Plain, central Japan and its correlation with oxygen isotope record from deep sea cores. The Geological Society of Japan. 1999; 105: 332–340. [Google Scholar]
- 41. Kerp H. The study of fossil gymnosperms by means of cuticular analysis. Palaios. 1990; 5: 548–569. [Google Scholar]
- 42. Jones TP, Rowe NP, editors. Fossil plants and spores: modern techniques London: Geological Society of London; 1999. [Google Scholar]
- 43. Salisbury EJ. On the causes and ecological significance of stomatal frequency, with special reference to the woodland flora In: Philosophical Transactions of the Royal Society of London (vol. 216). London: The Royal Society; 1982. pp. 1–65. [Google Scholar]
- 44. Zachos J, Pagani M, Sloan LC, Thomas E, Billups K. Trends, rhythms, and aberrations in global climate 65 Ma to present. Science. 2001; 292: 686–693. [DOI] [PubMed] [Google Scholar]
- 45. Retallack GJ. Refining a pedogenic-carbonate CO2 paleobarometer to quantify a middle Miocene greenhouse spike. Palaeogeogr Palaeoclimatol Palaeoecol. 2009; 281: 57–65. [Google Scholar]
- 46. Zachos JC, Dickens GR, Zeebe RE. An early Cenozoic perspective on greenhouse warming and carbon-cycle dynamics. Nature. 2008; 451: 279–283. 10.1038/nature06588 [DOI] [PubMed] [Google Scholar]
- 47. Tripati AK, Roberts CD, Eagle RA, Li G (2011) A 20 million year record of planktic foraminiferal B/Ca ratios: Systematics and uncertainties in pCO2 reconstructions. Geochim Cosmochim Acta. 2011; 75: 2582–2610. [Google Scholar]
- 48. Foster GL, Lear CH, Rae JW. The evolution of pCO2, ice volume and climate during the middle Miocene. Earth Planet Sci Lett. 2012; 341: 243–254. [Google Scholar]
- 49. Henderiks J, Pagani M. Coccolithophore cell size and the Paleogene decline in atmospheric CO2 . Earth Planet Sci Lett. 2008; 269: 576–584. [Google Scholar]
- 50. Pagani M, Lemarchand D, Spivack A, Gaillardet J. A critical evaluation of the boron isotope-pH proxy: The accuracy of ancient ocean pH estimates. Geochim Cosmochim Acta. 2005; 69: 953–961. [Google Scholar]
- 51.Kürschner WM. Leaf stomata as biosensors of paleoatmospheric CO2 levels. PhD Thesis, Utrecht University. 1996. Available: http://library.wur.nl/WebQuery/clc/94370.
- 52. Burgh J, Visscher H, Dilcher DL, Kurschner WM. Paleoatmospheric signatures in Neogene fossil leaves. Science. 1993; 260: 1788–1790. [DOI] [PubMed] [Google Scholar]
- 53. Tripati AK, Roberts CD, Eagle RA. Coupling of CO2 and ice sheet stability over major climate transitions of the last 20 million years. Science. 2009; 326: 1394–1397. 10.1126/science.1178296 [DOI] [PubMed] [Google Scholar]
- 54. Seki O, Foster GL, Schmidt DN, Mackensen A, Kawamura K, Pancost RD. Alkenone and Boron-based Pliocene pCO2 records. Earth Planet Sci Lett. 2010; 292: 201–211. [Google Scholar]
- 55. Stults DZ, Wagner-Cremer F, Axsmith BJ. Atmospheric paleo-CO2 estimates based on Taxodium distichum (Cupressaceae) fossils from the Miocene and Pliocene of Eastern North America. Palaeogeogr Palaeoclimatol Palaeoecol. 2011; 309: 327–332. [Google Scholar]
- 56. Li L, Li Q, Tian J, Wang P, Wang H, Liu ZH. A 4-Ma record of thermal evolution in the tropical western Pacific and its implications on climate change. Earth Planet Sci Lett. 2011; 309: 10–20. [Google Scholar]
- 57. Lisiecki LE, Raymo ME. A Pliocene-Pleistocene stack of 57 globally distributed benthic δ18O records. Paleoceanography. 2005; 20: PA1003. [Google Scholar]
- 58. Ruddiman WF. Orbital insolation, ice volume, and greenhouse gases. Quat Sci Rev. 2003; 22: 1597–1629. [Google Scholar]
- 59. Flower BP, Kennett JP. The middle Miocene climatic transition: East Antarctic ice sheet development, deep ocean circulation and global carbon cycling. Palaeogeogr Palaeoclimatol Palaeoecol. 1994; 108: 537–555. [Google Scholar]
- 60. Shevenell AE, Kennett JP, Lea DW. Middle Miocene ice sheet dynamics, deep-sea temperatures, and carbon cycling: A Southern Ocean perspective. Geochemistry, Geophysics, Geosystems. 2008; 9: Q02006. [Google Scholar]
- 61. Zhang YG, Pagani M, Liu Z, Bohaty SM, DeConto R. (2013) A 40-million-year history of atmospheric CO2 . 2013; 373: 1–20. [DOI] [PubMed] [Google Scholar]
- 62. Vincent E, Berger WH. Carbon dioxide and polar cooling in the Miocene: The Monterey hypothesis. The Carbon cycle and atmospheric CO2: Natural Variations Archean to Present. 1985; 32: 455–468. [Google Scholar]
- 63. Badger MPS, Lear CH, Pancost RD, Foster GL, Bailey TR, Leng MG et al. CO2 drawdown following the middle Miocene expansion of the Antarctic Ice Sheet. Paleoceanography. 2013; 28: 12. [Google Scholar]
- 64. Mosbrugger V, Utescher T, Dilcher DL. Cenozoic continental climatic evolution of Central Europe. PNAS. 2005; 102: 14964–14969. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65. Shevenell AE, Kennett JP, Lea DW. Middle Miocene Southern Ocean cooling and Antarctic cryosphere expansion. Science. 2004; 305: 1766–1770. [DOI] [PubMed] [Google Scholar]
- 66. Pearson PN, Ditchfield PW, Singano J, Harcourt-Brown KG, Nicholas CJ, Olsson RK et al. Warm tropical sea surface temperatures in the Late Cretaceous and Eocene epochs. Nature. 2001; 414: 470–470. [DOI] [PubMed] [Google Scholar]
- 67. Knorr G, Butzin M, Micheels A, Lohmann G. A warm Miocene climate at low atmospheric CO2 levels. Geophys Res Lett. 2011; 38: 5. [Google Scholar]
- 68. Schepper SD, Groeneveld J, Naafs BDA, Renterghem CV, Hennissen J, Head MJ. Northern hemisphere glaciation during the globally warm early late Pliocene. PLoS ONE. 2013; 8: e81508 10.1371/journal.pone.0081508 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69. Domingo L, Koch PL, Fernández HM, Fox DL, Domingo MS, Alberdi MT et al. Late Neogene and early Quaternary paleoenvironmental and paleoclimatic Conditions in Southwestern Europe: Isotopic analyses on mammalian taxa. PLoS ONE. 2013; 8: e63739 10.1371/journal.pone.0063739 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70. Raymo M. The initiation of Northern Hemisphere glaciation. Annual Review of Earth and Planetary Sciences. 1994; 22: 353–383. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
(DOC)
(DOC)
Data Availability Statement
All relevant data are within the paper and its Supporting Information files.