

Abstract
The innate immune system is an integral component of the inflammatory response to pathophysiological stimuli. Toll-like receptors (TLRs) and inflammasomes are the major sensors and pattern recognition receptors (PRRs) of the innate immune system that activate stimulus (signal)-specific pro-inflammatory responses. Chronic activation of PRRs has been found to be associated with the aggressiveness of various cancers and poor prognosis. Involvement of PRRs was earlier considered to be limited to infection- and injury-driven carcinogenesis, where they are activated by pathogenic ligands. With the recognition of damage-associated molecular patterns (DAMPs) as ligands of PRRs, the role of PRRs in carcinogenesis has also been implicated in other non-pathogen-driven neoplasms. Dying (apoptotic or necrotic) cells shed a plethora of DAMPs causing persistent activation of PRRs, leading to chronic inflammation and carcinogenesis. Such chronic activation of TLRs promotes tumor cell proliferation and enhances tumor cell invasion and metastasis by regulating pro-inflammatory cytokines, metalloproteinases, and integrins. Due to the decisive role of PRRs in carcinogenesis, targeting PRRs appears to be an effective cancer-preventive strategy. This review provides a brief account on the association of PRRs with various cancers and their role in carcinogenesis.
Keywords: pattern recognition receptors (PRRs), toll-like receptors (TLRs), damage-associated molecular patterns (DAMPs), inflammasome, HMGB1
Introduction
Many pathological conditions that involve tissue damage, bacterial or viral infections, and metabolic disorders leading to chronic inflammation are known to develop as a consequence of chronic activation of innate immune pattern recognition receptors (PRRs).1 The role of inflammation and innate immune function in cancer initiation and progression is well understood.2,3 Innate immunity is the first line of defense against infection and/or internal tissue injury and recognizes highly conserved sets of molecular structures called pathogen/damage-associated molecular patterns (PAMPs/DAMPs) via PRRs.1,4 Toll-like receptors (TLRs) and nucleotide oligomerization domain (NOD)-like receptors (NLRs) are the innate immune PRRs which, by their downstream signaling, activate multiple pro-inflammatory responses leading to an efficient antigen-specific acquired immunity. TLR signaling drives dendritic cell (DC) maturation, antigen presentation, as well as CD8+ T-cytotoxic effector functions, which are decisive for an efficient antitumor immunity.5 TLRs are also expressed by intestinal epithelial cells (IECs), where they regulate tissue homeostasis by stimulating immune response to bacterial pathogens while attenuating the immune response against favorable microbes.6 Furthermore, TLRs sense the breakdown of the protective intestinal barriers and trigger proliferative signaling.6 An aberrant TLR expression or chronic stimulation can alter homeostasis and cause negative regulation of antitumor immunity, as evidenced by increased immune-suppressive functions such as enhanced regulatory T-cell proliferation.7,8 Similarly, upon activation, NLRs lead to the secretion of mature forms of interleukin (IL)-1β and IL-18, triggering multiple inflammatory signaling pathways, which contribute to the chronic inflamed tumor microenvironment.3 DAMPs such as HMGB1 and ATP as well as microbial macromolecules such as double-stranded DNA (dsDNA) and double-stranded RNA (dsRNA) can act as intracellular or extracellular ligands to the innate PRRs.9,10 TLRs and NLRP3 function as critical sensors of metabolic disturbances caused by obesity, injury, and infection to augment the processes leading to the restoration of tissue homeostasis.11,12 TLRs and NLRs follow a coordinated signaling system that senses pathogenic invasion or tissue damage through various molecular signatures (PAMPs/DAMPs), triggering the expression of specific genes/signaling such as tumor necrosis factor (TNF)-α, IL-6, and IL-1β.1,13 Clinical studies suggest that the expression of PRRs is not limited to the immune cells, but they are also found to be elevated in various human epithelial cancers.14,15 Chronic activation of TLRs promotes carcinogenesis through pro-inflammatory responses, which augments the proliferative, anti-apoptotic, and pro-fibrogenic signals in the tumor microenvironment as well as in the tumor cells.15 Thus PRR homeostasis and their aberrant expression confer a dual role, ie, an immune-enhancing role that potentiates antitumor immunity, and a tumor-promoting role triggered by the aberrant and dysregulated expression. This review presents the current understanding of the functions of PRRs in promoting carcinogenesis and metastasis.
TLRs: From Pathogenic Sensors to Role in Cancer
Pathogenic sensor
TLRs are members of the type-1 transmembrane glycoprotein family that sense extracellular ligands and initiate pro-inflammatory signaling through various leucine-rich repeated sequences and Toll/interleukin-1 receptor (TIR) domains.16 There are five different TIR-containing adaptor proteins, namely, myeloid differentiation factor 88 (MyD88), MyD88 adaptor-like (TIRAP), TIR domain containing adaptor inducing interferon-β (TRIF), TRIF-related adaptor molecule (TRAM), and sterile α and heat-armadillo motifs.17,18 TLRs are expressed on the surface of the myeloid lineage cells, ie, monocytes, macrophages, dendritic cells (DCs), and mast cells as well as other immune cells such as lymphocytes.16 At present, 10 TLRs (TLR1–10) have been identified in humans and 12 (TLR1–9 and TLR11–13) in mice, which sense pathogenic wall components as well as foreign DNA and RNA fragments.17–29 These components are mainly lipid or lipid-derived macromolecules such as lipopeptides (TLR1–6), nucleic acid fragments (TLR3 and TLR7–9), and flagellin (TLR5).17–23 TLR3 specifically recognizes double-stranded RNA (dsRNA) from viruses, while TLR7 and TLR8 recognize single-stranded RNAs (ssRNA).17,22,23 TLR7 has an affinity for immunoglobulin/self-RNA complexes and synthetic ligands such as imiquimod and is associated with autoimmune disease conditions, while TLR9 specifically binds to unmethylated CpG sequences (found in viral DNA), synthetic oligodeoxynucleotides (ODNs), or immunoglobulin-DNA complexes.17,22,30 Detailed information on different TLRs, their ligands, and cellular localization is summarized in Table 1.
Table 1.
Diversity of Toll-like receptors (TLRs), their ligands, and cellular localization.
| TLRs | SPECIES | LIGAND | LOCALIZATION | REFERENCES |
|---|---|---|---|---|
| TLR1/TLR2 | Mice and humans | Peptidoglycans, lipopolysaccharide (LPS) | Cell surface | 17, 18 |
| TLR2/TLR6 | Mice and humans | Diacylated lipoproteins | Cell surface | 18 |
| TLR3 | Mice and humans | ds RNA | Endosomes | 19 |
| TLR4 | Mice and humans | LPS | Cell surface | 20 |
| TLR5 | Mice and humans | Flagellin protein | Cell surface | 21 |
| TLR7 | Mice and humans | ssRNA | Endosomes | 22 |
| TLR8 | Mice and humans | ssRNA | Endosomes | 23 |
| TLR9 | Mice and humans | Unmethylated CpG DNA | Endosomes | 17 |
| TLR10 | Humans | Listeria proteins (no specific ligand) | Endosomes | 24–26 |
| TLR11 | Mice | Bacterial profiling proteins and flagellin | Endosomes | 27 |
| TLR12 | Mice | Bacterial profiling proteins | Endosomes | 28 |
| TLR13 | Mice | 23S rRNA | Endosomes | 29 |
Associaton with cancer
Involvement of TLR4 and MyD88 in tumor progression has been demonstrated in various preclinical models, such as 7,12-dimethylbenz (α) anthracene (DMBA)-initiated and 12-O-tetradecanoylphorbol 13-acetate (TPA)-promoted mouse skin tumorigenesis, azoxymethanedextran sulfate [(AOM)-DS]-induced colon carcinoma, and diethylnitrosamine (DEN)-induced hepato-carcinogenesis.31–33 Besides these classic inflammation-induced carcinogenesis models, TLR4 and MyD88 have also been shown to contribute to tumor immune-editing-driven tumor models such as 3-methylcholanthrene (3-MCA)-induced sarcoma.31,34 TLR5 is shown to be responsible for wound-induced skin cancers, and TLR7 agonist (ssRNA40) leads to the pronounced tumor growth and stromal expansion in K-RAS-overexpressed pancreatic ductal carcinoma.35,36 Human myeloma cancers and myeloma cell lines exhibit differential expression of nearly all the TLRs.37 Various hematological malignancies such as chronic lymphoid leukemia (CLL), acute lymphoblastic leukemia (ALL), mantle cell lymphoma (MCL), and B-cell lymphoma have been found with the elevated TLR expression.38 TLR10, TLR1, and TLR6 have recently been found to be involved in prostate carcinogenesis.39 TLR overexpression has also been correlated with the resistance of tumors to the chemotherapeutic drugs such as paclitaxel.40 Table 2 describes various TLRs and their association with various cancers. All these studies describe the association of TLRs in nearly all cancer types, which therefore could be exploited as potential targets for developing tumor-preventive strategies.
Table 2.
Toll-like receptors (TLRs) and associated cancers.
| TLRs | CANCER | REFERENCES |
|---|---|---|
| TLR1 | Prostate cancer | 39 |
| TLR2 | Breast cancer, hepatocarcinoma | 142 |
| TLR3 | Esophageal squamous cell carcinoma | 143 |
| TLR4 | Esophageal squamous cell carcinoma, colorectal cancer, bladder cancer, ovarian cancer, prostate cancer, cervical cancer, pancreatic cancer, hepatocellular carcinoma, lung cancer, breast cancer | 5, 39, 143–147 |
| TLR6 | Prostate cancer | 39 |
| TLR7 | Pancreatic carcinogenesis, esophageal squamous cell carcinoma | 143 |
| TLR9 | Lung cancer, esophageal squamous cell carcinoma | 143 |
| TLR10 | Prostate cancer, nasopharyngeal | 39, 120 |
Signaling
Stimulation of TLRs is known to trigger the expression of several genes that are involved in various inflammatory and immune responses. Binding of ligands leads to the dimerization of TLRs, TLR2 forming heterodimers with TLR1 or TLR6 while all other TLRs forming homodimers (Fig. 1).17,41,42 Dimerization of TLRs initiate signaling events (originating from the cytoplasmic TIR domain of the receptor) and progresses through adaptor proteins involving two major pathways (Fig. 1). In the first, the activated MyD88 adapter protein recruits IRAK-4 to TLRs, facilitating IRAK-4-mediated phosphorylation of IRAK-1, which then binds to TRAF-6 leading to the activation of two subpathways. One of them involves MAP kinases leading to the activation of AP-1 and its recruitment to the nucleus, while the other, where the TAK1/TAB complex is activated, enhances the activity of the IKB kinase (IKK) resulting in phosphorylation and degradation of IKB, thereby increasing the localization of NF-κB to the nucleus.17,43 MyD88-dependent TLR signaling involves a second TIR domain molecule, namely TIRAP/Mal. Further, the MyD88-independent TLR signaling involves a third TIR domain involving a TRIF-dependent pathway, where signaling is executed with the recruitment of IRF to the nucleus via TBK-1. IRF-3 regulates the transcription of interferon (IFN)-β and is therefore called TIR domain containing adaptor-inducing IFN-β (TRIF).17 TRIF-dependent pathways are pivotal during the stimulation through TLR3 and TLR4.2,44 All TLRs utilize the MyD88-dependent signaling pathway to induce the expression of pro-inflammatory cytokine genes with the exception of TLR3, which exclusively uses the TRIF pathway.2 Both MyD88-dependent cascades and TRIF pathways have been associated with tumor growth and metastasis.45 Eventually, TLR functions are regulated by the transcription factors AP-1, NF-κB, and IRF-3.2,17,46
Figure 1.

TLR diversity, ligands, and signaling. TLRs are expressed on the epithelial and immune cells on the plasma membrane (TLR1, TLR2, TLR4, and TLR5) and endosomal membrane (TLR3, TLR7, and TLR9). Pathogenic PAMPS are the natural ligands of TLRs: eg, bacterial lipid-derived macromolecules are ligands for TLR1, TLR2, TLR4, and TLR6, while nucleic acid derivatives like dsDNA (TLR3) nucleotide complexes with other macromolecules (TLR7) and the CpG nucleotide sequence (TLR9). TLR signaling in an MyD88-dependent and TRIF-mediated (MyD88-independent) pathway leads to the translocation of AP-1, NF-κB, and IRF-3, respectively, to the nucleus, which induces the transcriptional activation of pro-inflammatory genes.
TLR signaling coupled with other inflammatory pathways contributes to the inflamed tumor microenvironment and leads to the inflammation-driven disease progression; eg, chronic infection by Helicobactor pylori leads to the development of gastric cancer.47 In spite of the inability to stimulate TLR4 on its own, H. pylori actively promotes inflammation by upregulating TLR4 expression via the TLR2 and MEK1/2-ERK1/2 signaling pathway.48,49 IL-6 and other TLR-induced effector cytokines, such as TNF-α and IL-17A, strongly promote mucosal and hepatic cancer. Another associated signaling is the IL-6-mediated STAT3 activation, which is known to be highly pro-oncogenic and also contributes to the radioresistance of tumor cells.50,51 Similarly, STAT3-mediated T-helper type 17 (TH-17) expression has been shown to facilitate tumor development in APC−/+ mice.52 Recent studies have shown the involvement of nearly all the TLRs in increased cancer incidence, disease severity, and poor prognosis, and thus can be exploited as targets for the cancer-preventive approaches.53–55
Inflammasomes: Inflaming Tumors
Inflammasome assembly
Inflammasomes are multiscaffold proteins with an interacting N-terminal homotypic protein–protein interaction motif called caspase-1 activation recruitment domain (CARD), where procaspase-1 undergoes a conformational change required for its cleavage and activation.56 The release of mature IL-1β and IL-18 from myeloid cells is dependent on activation of procaspase-1 to active 10- and 20-kDa caspase-1 peptides.57 Inflammasomes are formed with NLR proteins containing an N-terminal CARD or pyrin domains required for homotypic protein–protein interaction, an intermediate nucleotide binding with self-oligomerization potential, the NACHT domain, and a C-terminal domain containing leucine-rich repeats (LRRs).58 NLRP3 inflammasomes (NACHT, LRR, and pyrin domain-containing protein) are involved in the sensing of DAMPs such as extracellular ATP, monosodium urate (MSU) crystals, asbestos, silica, and β-amyloid.59–62 Inflammasome activation requires the interaction of pyrin domain (PYD) of ASC (apoptosis-associated speck-like protein containing C-terminal CARD) with PYD of NLRP3, forming a functional inflammasome complex through CARD–CARD interaction of ASC with procaspase.63 Classical inflammasome activation has two steps: The first step involves the induction of mRNA followed by the expression of pro-IL-1β and pro-IL-18 followed by NF-κB translocation to the nucleus, which requires either TLR/NOD stimulation or signaling through TNF-α or IL-1β receptor.64 The second and the most critical step is the sensing of PAMPs or DAMPs by NLRs, which leads to the autocatalytic cleavage of caspase-1 (Fig. 2).65
Figure 2.
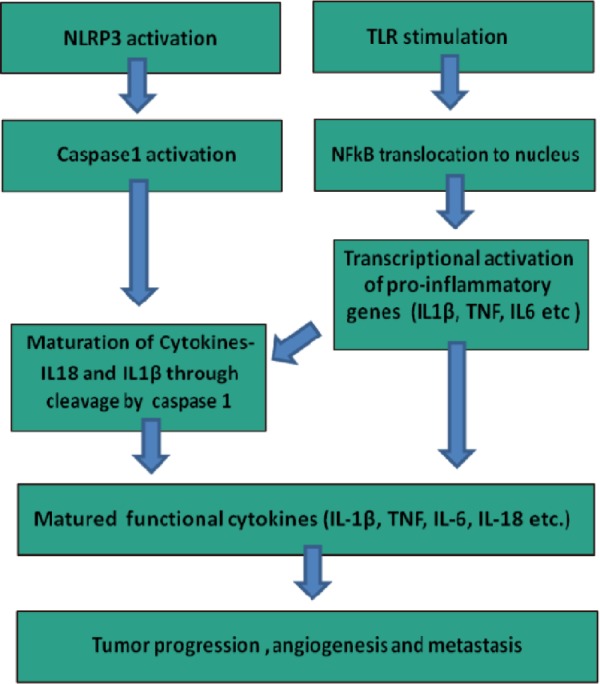
Coordinated action of TLRs and NLRP3 inflammasomes in tumor progression, angiogenesis, and metastasis.
Association with cancer
The major products of inflammasome activation are the pro-inflammatory cytokines IL-1β and IL-18, which are pro-tumorigenic in inflammation-induced gastrointestinal (GI) cancers.3 Chronic inflammation in the stomach due to H. pylori infection or other causes is mediated by the upregulation of pro-inflammatory cytokines, including IL-1β.66,67 While stomach-specific expression of IL-1β in mice induces inflammation and gastric carcinogenesis, in colitis-associated cancer (CAC) IL-1β may promote tumor growth and invasion by inducing an epithelial to mesenchymal transition (EMT) as well as stem cell phenotype in colon tumor cells.68,69 Further, IL-1β stimulates COX-2, IL-6, IL-8, and CCL5 production, which leads to an increase in the proliferative and invasive capabilities of colon cancer cells.70–72
DAMPs as PRR Activators
TLR-mediated carcinogenesis was earlier thought to be associated with organs that directly or indirectly get exposed to the bacterial TLR ligands, such as the GI tract, skin, and liver.73 However, with the discovery of the DAMPs as endogenous TLR ligands, the possible role of TLRs in carcinogenesis has extended beyond the pathogenic ligand-associated organs. DAMP–PRR interaction leads to the smoldering chronic inflammation, which nurtures transformed and tumor cells with mitogenic, anti-apoptotic, and proliferative factors (Fig. 3). DAMPs are either released from the dying cells in the injured/inflamed tissues or actively secreted from the lysosomes.74
Figure 3.

Activation of TLRs and inflammasomes by DAMPs released by apoptotic and necrotic cells. DAMPs from nucleus (HSPs, histones, HMGB1), cytosol (uric acid, ATP), and mtDNA activate PRR signaling in tumor and myeloid cells, which leads to the secretion of pro-inflammatory mediators such as TNF-α, IL-1-β, and IL-6. TLR signaling eventually promotes the transcriptional activation of pro-inflammatory cytokines via localization of NF-κB, while inflammasome activation guides caspases-1 to cleave pro IL-1-β to mature IL-1β, altering integrin and chemokine expression leading to the tumor growth and metastasis by promoting angiogenesis and tumor cell migration.
HMGB1 is known to induce skin, liver, and pancreatic tumors.75 Along with TLR9, TLR2, and TLR4, it can start a pro-inflammatory signaling, which further leads to the chronic inflamed tumor milieu.76–78 Cytoplasmic DAMP, uric acid, can also activate macrophages through TLR2 and TLR4 and also act as an NLR activator.79,80 The cell wall components bi-glycan and decorin also bind to TLR2 and TLR4 and activate pro-inflammatory signaling.81,82 The ability of S100A8 and S100A9 (members of S100 class of proteins) to promote pre-metastatic niches in lungs also requires TLR4-mediated signaling.83 Table 3 describes the various DAMPs and their source organelle. Several studies have shown the involvement of DMAPs in NLRP3 activation. DAMPs such as MSU and ATP are known to activate inflammasomes, leading to the huge burst of reactive oxygen species (ROS) and K+ efflux.84 While ATP activates the inflammasomes through purinergic receptors, MSU also functions by triggering the ATP release from the nearby cells and then leading to an ATP-dependent activation of inflammasomes.85 DAMPs play a critical role in carcinogenesis by continuous activation of PRR signaling, leading to the chronic/sustained inflammation.85 Hypoxic and necrotic regions in tumors continuously release loads of DAMPs to the tumor microenvironment, stimulating TLRs as well as factors such as matrix metalloproteinase (MMP)-9 and vascular endothelial growth factor (VEGF), making the tumor more aggressive and invasive.86
Table 3.
Sources and receptors of endogenous pattern recognition receptor (PRR) ligands.
| PRR LIGANDS (DAMPS) | SOURCE | RECEPTORS | REFERENCES |
|---|---|---|---|
| Uric acid (monosodium urate) | Cytosol | NLRP3 | 61, 80 |
| S100 proteins | Cytosol | TLR2, TLR4 | 83 |
| Heat shock proteins | Cytosol, nucleus | TLR2, TLR4 | 84 |
| Syndicans | Plasma membrane | TLR4 | 84 |
| Glypicans | Plasma membrane | TLR9 | 84 |
| mtDNA | Mitochondria | TLR3 | 9 |
| mROS | Mitochondria | NLRP3 | 9 |
| HMGB1 | Nucleus, cytosol | TLR4 | 9, 76 |
| Histones | Nucleus | TLR-4 | 9 |
| DNA | Nucleus | TLR3 | 9 |
| ATP | Autophagosomes, cytoplasm | P2X7-NLRP3 | 9 |
PRRs in Tumor Growth and Metastasis
PRR expression in tumors has been linked to increased severity and metastasis. PRR signaling promotes tumor growth and metastasis by altering the integrins and tumor cell motility. PRR activation leads to immune tolerance, which helps in evading the antitumor immunity. Furthermore, release of angiogenic factors leads to the neo-vascularization and favors tumor cell growth and migration.
Invasion and metastasis
TLRs along with intracellular signaling (NF-κB) regulates the integrin expression (B1) and motility in the tumor cells.87–89 TLR4 stimulation by lipopolysaccharide (LPS) has been shown to induce EMT and invasive phenotypes in human breast cancer cell lines such as MCF-7 and MDA-MB-231.89,90 Metastatic invasion involves the chemokine-mediated active transport of metastatic cells through the subcapsular sinus epithelium.91,92 TLR activation is known to induce the expression of CCR7 and CCR8, which are major chemokines involved in the tumor-mediated lymphatic remodeling of peritumoral lymph vessels and draining lymph nodes, thereby promoting metastasis.93,94 Many metastatic tumors appear to use the expression of these chemokine molecules to colonize in a new organ/tissue, while defective expression of the homing receptors such as CXCR3, CXCR4/CXCR7, and CCR6 has been observed in liver and lung metastases of colon cancer.95,96 Further, the ligands of CXCL19, SDF-1, and CCL20 receptors are highly expressed in the liver and lungs of metastatic colorectal cancer (CRC) patients.97,98 Integrin signaling is another metastatic mechanism, that is altered by the activation of TLRs. Integrins are required to facilitate immune cell trafficking in melanoma and other tumors.99 TLR activation leads to the acquisition of various integrins, chemokine expression, and trafficking properties similar to the leukocytes.100 Overexpression of PD-L1, TLR, TGF-β, CCL4, and CCL5 has been found to facilitate invasion and proliferation of such leukocytic tumor cells within the lymph nodes.100 Leukocytic tumors show upregulated E/P-selectins, L-selectin ligands, alpha-4B1, ICAM-1, and VCAM-1 required for trafficking and homing to a distal tissue.6,100,101 IL-18 and IL-1β are inflammasome effector pro-inflammatory cytokines, which upregulate the expression of adhesion molecules and therefore play a role in hepatic metastasis.102
Immune evasion
TLRs also help tumor cells in immune evasion, which in turn leads to the increased aggressiveness of the tumor, and increased TLR expression in different cancers appear to confer advantages of both immune evasion and immune suppression.103,104 TLR signaling alterations lead to the production of IL-10 and TGF-β, which are the major immunosuppressive cytokines.105,106 Further, TLR activation is accompanied by an increased expression of suppressive costimulatory molecules such as PD-L1 and HLA-G, which leads to the development of the immune tolerance.107–109 These suppressive functions regulate the hyperactive immune system during auto-immune disorders. TLR4 signaling leads to the increased expression of suppressive costimulatory markers B7-H1 and B7-H2 and inducible nitric oxide synthase (iNOS) by providing resistance against cytotoxic T-cell (CTL) attack.109 TLR-stimulated tumor cell supernatant has been shown to inhibit both T-cell proliferation and natural killer (NK) cell activity, while blocking of the TLR4 pathway reverses the tumor-mediated suppression of T-cell proliferation and NK cell activity and delays tumor growth.107
Angiogenesis
The role of inflammasomes in metastasis and cancer progression is demonstrated by the role of IL-1β and IL-18 (matured by activated NLRP3 inflammasome), which are shown to facilitate tumor angiogenesis. Elevated levels of active IL-1β in melanoma cells have been shown to act both in paracrine and autocrine manners, promoting angiogenesis, and thus contributing to invasion and metastasis.110 IL-1β induces the expression of angiogenic factors such as VEGF and IL-8, which promote tumor growth.110 Lewis lung carcinoma (LLC) cells overexpressing with IL-1β displayed increased angiogenesis and secretion of VEGF, and macrophage inflammatory protein-2 (MIP-2/CXCL2) has been correlated with faster tumor growth, while IL-1β-KO mice showed decreased angiogenesis and growth of melanoma tumors.111,112 Further, NLRP3 activation by the DAMPs in the tumor milieu is also known to induce the production of active IL-1β continuously, thereby contributing to the neovascularization.113 IL-1β signaling is also involved in the release of several angiogenic factors such as VEGF and IL-8 from tumor and stromal cells, thereby further promoting tumor growth.113
PRRs as Cancer Biomarkers
PRR expression has been found to be upregulated in many cancers and can be exploited as a biomarker. TLR4 expression has been shown as a cancer stem cell marker in hepatocellular carcinoma.96 The predictive role of TLR2, TLR4, and TLR9 has been established in gastric tumors and oral tongue squamous cell carcinoma.55 Immunohistochemical analysis and expression studies in adenocarcinoma patients have suggested that the high expression of TLR3, TLR4, and TLR9 levels is associated with prostate cancer recurrence.114,115 NLRP3 products IL-1β and IL-18 in body fluids are correlated with a high risk of pancreatic carcinoma and poor prognosis.116 IL-1β in cyst fluid appears to be a good predictor of enhanced risk of intraductal papillary mucinous pancreatic neoplasms.116 Plasma IL-1β expression is generally elevated in advanced cancers and correlates well with clinical features such as infirmity, sarcopenia, and loss of appetite as well as weight.117 Similarly, increased IL-18 levels in ovarian carcinoma patients appear to be a good predictor of disease severity.118
The ability of TLRs in predicting various cancers appears to be linked to the single-nucleotide polymorphisms (SNPs) at the loci of PRRs and their regulator genes.119 For example, patients carrying specific polymorphisms (enhanced TLR10 expression) show higher risk of prostate and nasopharyngeal carcinoma.120 Further, serum or ascite concentrations of IL-1RA, as well as the SNPs affecting these concentrations, can be used as important biomarkers.121 While SNPs in regulatory elements of NLRP3 determine susceptibility to inflammatory bowel disease, polymorphisms in IL-18 and IL-18R encoding genes can be used as biomarkers for Crohn’s disease.122,123
PRR Signaling as Potential Targets to Inhibit Carcinogenesis
Chronic stimulation of PRR-induced signaling by PAMPs or DAMPs leads to mitogenic and anti-apoptotic signals, which aid the tumor-promoting inflammatory events that ultimately result in malignant transformation. Modifiers of PRR signaling can be useful in targeting the process of carcinogenesis. Although PRR antagonists could be useful as tumor-preventive anti-inflammatory agents, only a few of them such as TLR4 antagonists CRX-526 (synthetic lipid A mimetic) and 1A6 (monoclonal antibody to TLR) have been investigated for preventing murine colitis.124,125 While TLR blockade as a strategy has failed to reduce the chronic T-cell transfer colitis, administration of 1A6 during the recovery phase of colitis has been found to prevent tumor development in the azoxymethane/dextran sulfate sodium (AOM-DSS) model.125,126 TLR4 antagonists are also being evaluated in clinical trials for treating sepsis.127,128 Inhibitors of STAT3 (downstream to PRR activation) such as OPB-31121 are already in clinical trials as therapeutics, suggesting that targeting secondary signaling to PRR could also be a potential antitumor therapy.129,130 In vivo manipulation of NLRs has been shown to delay the onset of colitis and cure the associated carcinogenesis. Glyburide (glibenclamide), a sulfonylurea drug used for the treatment of type-2 diabetes, is known to inhibit ATP-sensitive K+ channels and can thus prevent NLRP3 inflammasome activation.131,132 Compounds that target TLR signaling and inflammasomes, such as IL-1β neutralizing antibodies, recombinant IL-1R antagonist (anakinra), and IL-18 binding proteins, have already been established as therapeutics, while cancer-preventive strategies using small-molecule inhibitors that target caspase-1 needs to be established.133,134 ST2825, a carboxyamide derivative and novel inhibitor of MyD88 dimerization, is currently under evaluation in preclinical studies for the treatment of chronic inflammatory diseases.135 Other peptide inhibitors such as hydrocinnamoyl-l-valyl pyrrolidine (compound 4a) and Pephinh-MyD88 are also being developed to target MyD88 dimerization for the treatment of lymphoma with MyD88 mutations.136 Selective inhibition of NF-κB in cancer cells can block the stimulatory effect of TNF and markedly increase susceptibility to TRAIL-induced cell death, resulting in tumor regression.137 NF-κB inhibition and anti-TNF therapy, together with the administration of IFN or TRAIL, might offer an attractive strategy for immune-modulation-based cancer therapy. Anti-IL-1 monoclonal antibodies, soluble IL-1RII, and caspase-1 inhibitors are currently being exploited for the treatment of various inflammatory disorders, and can also be employed in tumor preventive strategies. PRR induction is fueled by various metabolites and dietary components.138,139 Metabolic interventions by dietary modification and manipulation of metabolism using modifiers such as metformin and 2-deoxy-d-glucose (2-DG) can also be useful as an effective preventive strategy against PRR-induced carcinogenesis. Indeed, dietary 2-DG has been shown to impair inflammation (DMBA-TPA)-induced tumorigenesis and radiation as well as tumor-induced angiogenesis, while enhanced local tumor control has been found to be associated with the stimulation of inflammation following therapeutic administration of 2-DG.140–143
Although TLRs and inflammasomes appear to be promising targets for cancer therapy and prevention, further studies are required to determine the context-specific functions of PRRs during cancer development and progression. Efficacy of therapies targeting inflammasomes will depend on our ability to determine when such therapies will be beneficial and when they will be detrimental due to the context-specific function of the inflammasomes in cancer.
Footnotes
ACADEMIC EDITOR: Marc D. Basson, Editor in Chief
PEER REVIEW: Nine peer reviewers contributed to the peer review report. Reviewers’ reports totaled 2,135 words, excluding any confidential comments to the academic editor.
FUNDING: Authors disclose no funding sources.
COMPETING INTERESTS: Authors disclose no potential conflicts of interest.
Paper subject to independent expert blind peer review. All editorial decisions made by independent academic editor. Upon submission manuscript was subject to anti-plagiarism scanning. Prior to publication all authors have given signed confirmation of agreement to article publication and compliance with all applicable ethical and legal requirements, including the accuracy of author and contributor information, disclosure of competing interests and funding sources, compliance with ethical requirements relating to human and animal study participants, and compliance with any copyright requirements of third parties. This journal is a member of the Committee on Publication Ethics (COPE). Provenance: the authors were invited to submit this paper.
Author Contributions
Prepared the preliminary draft of the manuscript: SP. Contributed to the writing of the manuscript: SS and VA. Jointly developed the structure and arguments for the paper: SP, BSD and ANB. Made critical revisions and approved final version: BSD, ANB and KN. All authors reviewed and approved the final manuscript.
REFERENCES
- 1.Takeuchi O, Akira S. Pattern recognition receptors and inflammation. Cell. 2010;140(6):805–820. doi: 10.1016/j.cell.2010.01.022. [DOI] [PubMed] [Google Scholar]
- 2.Pradere JP, Dapito DH, Schwabe RF. The Yin and Yang of toll-like receptors in cancer. Oncogene. 2014;33(27):3485–3495. doi: 10.1038/onc.2013.302. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Kolb R, Liu GH, Janowski AM, Sutterwala FS, Zhang W. Inflammasomes in cancer: a double-edged sword. Protein Cell. 2014;5(1):12–20. doi: 10.1007/s13238-013-0001-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Kumar H, Kawai T, Akira S. Pathogen recognition by the innate immune system. Int Rev Immunol. 2011;30(1):16–34. doi: 10.3109/08830185.2010.529976. [DOI] [PubMed] [Google Scholar]
- 5.Schreibelt G, Tel J, Sliepen KH, et al. Toll-like receptor expression and function in human dendritic cell subsets: implications for dendritic cell-based anti-cancer immunotherapy. Cancer Immunol Immunother. 2010;59(10):1573–1582. doi: 10.1007/s00262-010-0833-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Luddy KA, Robertson-Tessi M, Tafreshi NK, Soliman H, Morse DL. The role of toll-like receptors in colorectal cancer progression: evidence for epithelial to leucocytic transition. Front Immunol. 2014;5:429. doi: 10.3389/fimmu.2014.00429. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Conroy H, Marshall NA, Mills KH. TLR ligand suppression or enhancement of Treg cells? A double-edged sword in immunity to tumours. Oncogene. 2008;27(2):168–180. doi: 10.1038/sj.onc.1210910. [DOI] [PubMed] [Google Scholar]
- 8.Caramalho I, Lopes-Carvalho T, Ostler D, Zelenay S, Haury M, Demengeot J. Regulatory T cells selectively express toll-like receptors and are activated by lipopolysaccharide. J Exp Med. 2003;197(4):403–411. doi: 10.1084/jem.20021633. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Schaefer L. Complexity of danger: the diverse nature of damage-associated molecular patterns. J Biol Chem. 2014;289(51):35237–35245. doi: 10.1074/jbc.R114.619304. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Nakamura T, Ito T, Igarashi H, et al. Cytosolic double-stranded DNA as a damage-associated molecular pattern induces the inflammatory response in rat pancreatic stellate cells: a plausible mechanism for tissue injury-associated pancreatitis. Int J Inflam. 2012;2012:504128. doi: 10.1155/2012/504128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Tannahill GM, O’Neill LA. The emerging role of metabolic regulation in the functioning of toll-like receptors and the NOD-like receptor Nlrp3. FEBS Lett. 2011;585(11):1568–1572. doi: 10.1016/j.febslet.2011.05.008. [DOI] [PubMed] [Google Scholar]
- 12.De Nardo D, Latz E. NLRP3 inflammasomes link inflammation and metabolic disease. Trends Immunol. 2011;32(8):373–379. doi: 10.1016/j.it.2011.05.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Fukata M, Vamadevan AS, Abreu MT. Toll-like receptors (TLRs) and Nod-like receptors (NLRs) in inflammatory disorders. Semin Immunol. 2009;21(4):242–253. doi: 10.1016/j.smim.2009.06.005. [DOI] [PubMed] [Google Scholar]
- 14.Sato Y, Goto Y, Narita N, Hoon DS. Cancer cells expressing toll-like receptors and the tumor microenvironment. Cancer Microenviron. 2009;2(suppl 1):205–214. doi: 10.1007/s12307-009-0022-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Castaño-Rodríguez N, Kaakoush NO, Mitchell HM. Pattern-recognition receptors and gastric cancer. Front Immunol. 2014;5:336. doi: 10.3389/fimmu.2014.00336. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Akira S, Yamamoto M, Takeda K. Role of adapters in toll-like receptor signalling. Biochem Soc Trans. 2003;31(pt 3):637–642. doi: 10.1042/bst0310637. [DOI] [PubMed] [Google Scholar]
- 17.Takeda K, Akira S. Toll-like receptors in innate immunity. Int Immunol. 2005;17(1):1–14. doi: 10.1093/intimm/dxh186. [DOI] [PubMed] [Google Scholar]
- 18.Kawai T, Akira S. The role of pattern-recognition receptors in innate immunity: update on toll-like receptors. Nat Immunol. 2010;11(5):373–384. doi: 10.1038/ni.1863. [DOI] [PubMed] [Google Scholar]
- 19.Bernard JJ, Cowing-Zitron C, Nakatsuji T, et al. Ultraviolet radiation damages self noncoding RNA and is detected by TLR3. Nat Med. 2012;18(8):1286–1290. doi: 10.1038/nm.2861. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Erridge C. Endogenous ligands of TLR2 and TLR4: agonists or assistants? J Leukoc Biol. 2010;87(6):989–999. doi: 10.1189/jlb.1209775. [DOI] [PubMed] [Google Scholar]
- 21.Akira S, Uematsu S, Takeuchi O. Pathogen recognition and innate immunity. Cell. 2006;124(4):783–801. doi: 10.1016/j.cell.2006.02.015. [DOI] [PubMed] [Google Scholar]
- 22.Mancuso G, Gambuzza M, Midiri A, et al. Bacterial recognition by TLR7 in the lysosomes of conventional dendritic cells. Nat Immunol. 2009;10(6):587–594. doi: 10.1038/ni.1733. [DOI] [PubMed] [Google Scholar]
- 23.Guiducci C, Gong M, Cepika AM, et al. RNA recognition by human TLR8 can lead to autoimmune inflammation. J Exp Med. 2013;210(13):2903–2919. doi: 10.1084/jem.20131044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Lee SM, Kok KH, Jaume M, et al. Toll-like receptor 10 is involved in induction of innate immune responses to influenza virus infection. Proc Natl Acad Sci U S A. 2014;111(10):3793–3798. doi: 10.1073/pnas.1324266111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Regan T, Nally K, Carmody R, et al. Identification of TLR10 as a key mediator of the inflammatory response to Listeria monocytogenes in intestinal epithelial cells and macrophages. J Immunol. 2013;191(12):6084–6092. doi: 10.4049/jimmunol.1203245. [DOI] [PubMed] [Google Scholar]
- 26.Barton GM, Medzhitov R. Toll-like receptor signaling pathways. Science. 2003;300(5625):1524–1525. doi: 10.1126/science.1085536. [DOI] [PubMed] [Google Scholar]
- 27.Raetz M, Kibardin A, Sturge CR, et al. Cooperation of TLR12 and TLR11 in the IRF8-dependent IL-12 response to Toxoplasma gondii profilin. J Immunol. 2013;191(9):4818–4827. doi: 10.4049/jimmunol.1301301. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Ghosh D, Stumhofer JS. Do you see what I see: recognition of protozoan parasites by toll-like receptors. Curr Immunol Rev. 2013;9(3):129–140. doi: 10.2174/1573395509666131203225929. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Oldenburg M, Krüger A, Ferstl R, et al. TLR13 recognizes bacterial 23S rRNA devoid of erythromycin resistance-forming modification. Science. 2012;337(6098):1111–1115. doi: 10.1126/science.1220363. [DOI] [PubMed] [Google Scholar]
- 30.Hemmi H, Kaisho T, Takeuchi O, et al. Small anti-viral compounds activate immune cells via the TLR7 MyD88-dependent signaling pathway. Nat Immunol. 2002;3(2):196–200. doi: 10.1038/ni758. [DOI] [PubMed] [Google Scholar]
- 31.Jarnicki A, Putoczki T, Ernst M. Stat3: linking inflammation to epithelial cancer—more than a “gut” feeling? Cell Div. 2010;5:14. doi: 10.1186/1747-1028-5-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Fukata M, Chen A, Vamadevan AS, et al. Toll-like receptor-4 promotes the development of colitis-associated colorectal tumors. Gastroenterology. 2007;133(6):1869–1881. doi: 10.1053/j.gastro.2007.09.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Naugler WE, Sakurai T, Kim S, et al. Gender disparity in liver cancer due to sex differences in MyD88-dependent IL-6 production. Science. 2007;317(5834):121–124. doi: 10.1126/science.1140485. [DOI] [PubMed] [Google Scholar]
- 34.Salcedo R, Cataisson C, Hasan U, Yuspa SH, Trinchieri G. MyD88 and its divergent toll in carcinogenesis. Trends Immunol. 2013;34(8):379–389. doi: 10.1016/j.it.2013.03.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Hoste E, Arwert EN, Lal R, et al. Innate sensing of microbial products promotes wound-induced skin cancer. Nat Commun. 2015;6:5932. doi: 10.1038/ncomms6932. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Ochi A, Graffeo CS, Zambirinis CP, et al. Toll-like receptor 7 regulates pancreatic carcinogenesis in mice and humans. J Clin Invest. 2012;122(11):4118–4129. doi: 10.1172/JCI63606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Abdi J, Garssen J, Redegeld F. Toll-like receptors in human multiple myeloma: new insight into inflammation-related pathogenesis. Curr Mol Med. 2014;14(4):423–431. doi: 10.2174/1566524014666140414204100. [DOI] [PubMed] [Google Scholar]
- 38.Isaza-Correa JM, Liang Z, van den Berg A, Diepstra A, Visser L. Toll-like receptors in the pathogenesis of human B cell malignancies. J Hematol Oncol. 2014;7:57. doi: 10.1186/s13045-014-0057-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Stevens VL, Hsing AW, Talbot JT, et al. Genetic variation in the toll-like receptor gene cluster (TLR10-TLR1-TLR6) an prostate cancer risk. Int J Cancer. 2008;123(11):2644–2650. doi: 10.1002/ijc.23826. [DOI] [PubMed] [Google Scholar]
- 40.Kelly MG, Alvero AB, Chen R, et al. TLR-4 signaling promotes tumor growth and paclitaxel chemoresistance in ovarian cancer. Cancer Res. 2006;66(7):3859–3868. doi: 10.1158/0008-5472.CAN-05-3948. [DOI] [PubMed] [Google Scholar]
- 41.Trinchieri G, Sher A. Cooperation of toll-like receptor signals in innate immune defence. Nat Rev Immunol. 2007;7(3):179–190. doi: 10.1038/nri2038. [DOI] [PubMed] [Google Scholar]
- 42.Asea A, Rehli M, Kabingu E, et al. Novel signal transduction pathway utilized by extracellular HSP70: role of toll-like receptor (TLR) 2 and TLR4. J Biol Chem. 2002;277(17):15028–15034. doi: 10.1074/jbc.M200497200. [DOI] [PubMed] [Google Scholar]
- 43.Kawai T, Akira S. Signaling to NF-kappaB by toll-like receptors. Trends Mol Med. 2007;13(11):460–469. doi: 10.1016/j.molmed.2007.09.002. [DOI] [PubMed] [Google Scholar]
- 44.Weighardt H, Jusek G, Mages J, et al. Identification of a TLR4 and TRIF-dependent activation program of dendritic cells. Eur J Immunol. 2004;34(2):558–564. doi: 10.1002/eji.200324714. [DOI] [PubMed] [Google Scholar]
- 45.Ochi A, Nguyen AH, Bedrosian AS, et al. MyD88 inhibition amplifies dendritic cell capacity to promote pancreatic carcinogenesis via Th2 cells. J Exp Med. 2012;209(9):1671–1687. doi: 10.1084/jem.20111706. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Kawai T, Takeuchi O, Fujita T, et al. Lipopolysaccharide stimulates the MyD88-independent pathway and results in activation of IFN-regulatory factor 3 and the expression of a subset of lipopolysaccharide-inducible genes. J Immunol. 2001;167(10):5887–5894. doi: 10.4049/jimmunol.167.10.5887. [DOI] [PubMed] [Google Scholar]
- 47.Fukata M, Abreu MT. Role of toll-like receptors in gastrointestinal malignancies. Oncogene. 2008;27(2):234–243. doi: 10.1038/sj.onc.1210908. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Yokota S, Okabayashi T, Rehli M, Fujii N, Amano K. Helicobacter pylori lipopolysaccharides upregulate toll-like receptor 4 expression and proliferation of gastric epithelial cells via the MEK1/2-ERK1/2 mitogen-activated protein kinase pathway. Infect Immun. 2010;78(1):468–476. doi: 10.1128/IAI.00903-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Yokota S, Amano K, Nishitani C, Ariki S, Kuroki Y, Fujii N. Implication of antigenic conversion of Helicobacter pylori lipopolysaccharides that involve interaction with surfactant protein D. Infect Immun. 2012;80(8):2956–2962. doi: 10.1128/IAI.00345-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Demaria M, Misale S, Giorgi C, et al. STAT3 can serve as a hit in the process of malignant transformation of primary cells. Cell Death Differ. 2012;19(8):1390–1397. doi: 10.1038/cdd.2012.20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.You S, Li R, Park D, et al. Disruption of STAT3 by niclosamide reverses radio-resistance of hu,an lung cancer. Mol Cancer Ther. 2014;13(3):606–616. doi: 10.1158/1535-7163.MCT-13-0608. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Li Y, Kundu P, Seow SW, et al. Gut microbiota accelerate tumor growth via c-jun and STAT3 phosphorylation in APC Min/+ mice. Carcinogenesis. 2012;33(6):1231–1238. doi: 10.1093/carcin/bgs137. [DOI] [PubMed] [Google Scholar]
- 53.Yesudhas D, Gosu V, Anwar MA, Choi S. Multiple roles of toll-like receptor 4 in colorectal cancer. Front Immunol. 2014;5:334. doi: 10.3389/fimmu.2014.00334. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Zhao S, Zhang Y, Zhang Q, Wang F, Zhang D. Toll-like receptors and prostate cancer. Front Immunol. 2014;5:352. doi: 10.3389/fimmu.2014.00352. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Mäkinen LK, Atula T, Häyry V, et al. Predictive role of toll-like receptors 2, 4, and 9 in oral tongue squamous cell carcinoma. Oral Oncol. 2015;51(1):96–102. doi: 10.1016/j.oraloncology.2014.08.017. [DOI] [PubMed] [Google Scholar]
- 56.Lamkanfi M, Dixit VM. Inflammasomes: guardians of cytosolic sanctity. Immunol Rev. 2009;227(1):95–105. doi: 10.1111/j.1600-065X.2008.00730.x. [DOI] [PubMed] [Google Scholar]
- 57.Franchi L, Eigenbrod T, Muñoz-Planillo R, Nuñez G. The inflammasome: a caspase-1-activation platform that regulates immune responses and disease pathogenesis. Nat Immunol. 2009;10(3):241–247. doi: 10.1038/ni.1703. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Martinon F, Mayor A, Tschopp J. The inflammasomes: guardians of the body. Annu Rev Immunol. 2009;27:229–265. doi: 10.1146/annurev.immunol.021908.132715. [DOI] [PubMed] [Google Scholar]
- 59.Halle A, Hornung V, Petzold GC, et al. The NALP3 inflammasome is involved in the innate immune response to amyloid-beta. Nat Immunol. 2008;9(8):857–865. doi: 10.1038/ni.1636. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Mariathasan S, Weiss DS, Newton K, et al. Cryopyrin activates the inflammasome in response to toxins and ATP. Nature. 2006;440(7081):228–232. doi: 10.1038/nature04515. [DOI] [PubMed] [Google Scholar]
- 61.Martinon F, Pétrilli V, Mayor A, Tardivel A, Tschopp J. Gout-associated uric acid crystals activate the NALP3 inflammasome. Nature. 2006;440(7081):237–241. doi: 10.1038/nature04516. [DOI] [PubMed] [Google Scholar]
- 62.Hillegass JM, Miller JM, MacPherson MB, et al. Asbestos and erionite prime and activate the NLRP3 inflammasome that stimulates autocrine cytokine release in human mesothelial cells. Part Fibre Toxicol. 2013;10:39. doi: 10.1186/1743-8977-10-39. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Mariathasan S, Newton K, Monack DM, et al. Differential activation of the inflammasome by caspase-1 adaptors ASC and Ipaf. Nature. 2004;430(6996):213–218. doi: 10.1038/nature02664. [DOI] [PubMed] [Google Scholar]
- 64.Nunes T, de Souza HS. Inflammasome in intestinal inflammation and cancer. Mediators Inflamm. 2013;2013:654963. doi: 10.1155/2013/654963. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Cassel SL, Sutterwala FS. Sterile inflammatory responses mediated by the NLRP3 inflammasome. Eur J Immunol. 2010;40(3):607–611. doi: 10.1002/eji.200940207. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Basso D, Scrigner M, Toma A, et al. Helicobacter pylori infection enhances mucosal interleukin-1 beta, interleukin-6, and the soluble receptor of interleukin-2. Int J Clin Lab Res. 1996;26(3):207–210. doi: 10.1007/BF02592984. [DOI] [PubMed] [Google Scholar]
- 67.McNamara D, El-Omar E. Helicobacter pylori infection and the pathogenesis of gastric cancer: a paradigm for host-bacterial interactions. Dig Liver Dis. 2008;40(7):504–509. doi: 10.1016/j.dld.2008.02.031. [DOI] [PubMed] [Google Scholar]
- 68.Tu S, Bhagat G, Cui G, et al. Overexpression of interleukin-1beta induces gastric inflammation and cancer and mobilizes myeloid-derived suppressor cells in mice. Cancer Cell. 2008;14(5):408–419. doi: 10.1016/j.ccr.2008.10.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Li Y, Wang L, Pappan L, Galliher-Beckley A, Shi J. IL-1β promotes stemness and invasiveness of colon cancer cells through Zeb1 activation. Mol Cancer. 2012;11:87. doi: 10.1186/1476-4598-11-87. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Zhu Y, Zhu M, Lance P. IL1β-mediated Stromal COX-2 signaling mediates proliferation and invasiveness of colonic epithelial cancer cells. Exp Cell Res. 2012;318(19):2520–2530. doi: 10.1016/j.yexcr.2012.07.021. [DOI] [PubMed] [Google Scholar]
- 71.Dinarello CA. Immunological and inflammatory functions of the interleukin-1 family. Annu Rev Immunol. 2009;27:519–550. doi: 10.1146/annurev.immunol.021908.132612. [DOI] [PubMed] [Google Scholar]
- 72.Boone BA, Lotze MT. Targeting damage-associated molecular pattern molecules (DAMPs) and DAMP receptors in melanoma. Methods Mol Biol. 2014;1102:537–552. doi: 10.1007/978-1-62703-727-3_29. [DOI] [PubMed] [Google Scholar]
- 73.Huang B, Zhao J, Unkeless JC, Feng ZH, Xiong H. TLR signaling by tumor and immune cells: a double-edged sword. Oncogene. 2008;27(2):218–224. doi: 10.1038/sj.onc.1210904. [DOI] [PubMed] [Google Scholar]
- 74.Pöllänen R, Sillat T, Pajarinen J, Levón J, Kaivosoja E, Konttinen YT. Microbial antigens mediate HLA-B27 diseases via TLRs. J Autoimmun. 2009;32(3–4):172–177. doi: 10.1016/j.jaut.2009.02.010. [DOI] [PubMed] [Google Scholar]
- 75.Lee SA, Kwak MS, Kim S, Shin JS. The role of high mobility group box 1 in innate immunity. Yonsei Med J. 2014;55(5):1165–1176. doi: 10.3349/ymj.2014.55.5.1165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Yu M, Wang H, Ding A, et al. HMGB1 signals through toll-like receptor TLR4 and TLR2. Shock. 2006;26(2):174–179. doi: 10.1097/01.shk.0000225404.51320.82. [DOI] [PubMed] [Google Scholar]
- 77.Ivanov S, Dragoi AM, Wang X, et al. A novel role for HMGB1 in TLR9-mediated inflammatory responses to CpG-DNA. Blood. 2007;110(6):1970–1981. doi: 10.1182/blood-2006-09-044776. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Park JS, Svetkauskaite D, He Q, et al. Involvement of toll-like receptors 2 and 4 in cellular activation by high mobility group box 1 protein. J Biol Chem. 2004;279(9):7370–7377. doi: 10.1074/jbc.M306793200. [DOI] [PubMed] [Google Scholar]
- 79.Liu-Bryan R, Pritzker K, Firestein GS, Terkeltaub R. TLR2 signaling in chondrocytes drives calcium pyrophosphate dihydrate and monosodium urate crystal-induced nitric oxide generation. J Immunol. 2005;174(8):5016–5023. doi: 10.4049/jimmunol.174.8.5016. [DOI] [PubMed] [Google Scholar]
- 80.Ghaemi-Oskouie F, Shi Y. The role of uric acid as an endogenous danger signal in immunity and inflammation. Curr Rheumatol Rep. 2011;13(2):160–166. doi: 10.1007/s11926-011-0162-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Neill T, Schaefer L, Iozzo RV. Decorin: a guardian from the matrix. Am J Pathol. 2012;181(2):380–387. doi: 10.1016/j.ajpath.2012.04.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Babelova A, Moreth K, Tsalastra-Greul W, et al. Biglycan, a danger signal that activates the NLRP3 inflammasome via toll-like and P2X receptors. J Biol Chem. 2009;284(36):24035–24048. doi: 10.1074/jbc.M109.014266. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Ehrchen JM, Sunderkötter C, Foell D, Vogl T, Roth J. The endogenous toll-like receptor 4 agonist S100A8/S100A9 (calprotectin) as innate amplifier of infection, autoimmunity, and cancer. J Leukoc Biol. 2009;86(3):557–566. doi: 10.1189/jlb.1008647. [DOI] [PubMed] [Google Scholar]
- 84.Feldman N, Rotter-Maskowitz A, Okun E. DAMPs as mediators of sterile inflammation in aging-related pathologies. Ageing Res Rev. 2015;(15):S1568–1637. 00005–7. doi: 10.1016/j.arr.2015.01.003. [DOI] [PubMed] [Google Scholar]
- 85.Gombault A, Baron L, Couillin I. ATP release and purinergic signaling in NLRP3 inflammasome activation. Front Immunol. 2013;3:414. doi: 10.3389/fimmu.2012.00414. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Tafani M, Russo A, Di Vito M, et al. Up-regulation of pro-inflammatory genes as adaptation to hypoxia in MCF-7 cells and in human mammary invasive carcinoma microenvironment. Cancer Sci. 2010;101(4):1014–1023. doi: 10.1111/j.1349-7006.2010.01493.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Killeen SD, Wang JH, Andrews EJ, Redmond HP. Bacterial endotoxin enhances colorectal cancer cell adhesion and invasion through TLR-4 and NF-kappaB-dependent activation of the urokinase plasminogen activator system. Br J Cancer. 2009;100(10):1589–1602. doi: 10.1038/sj.bjc.6604942. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Merrell MA, Ilvesaro JM, Lehtonen N, et al. Toll-like receptor 9 agonists promote cellular invasion by increasing matrix metalloproteinase activity. Mol Cancer Res. 2006;4(7):437–447. doi: 10.1158/1541-7786.MCR-06-0007. [DOI] [PubMed] [Google Scholar]
- 89.Wang JH, Manning BJ, Wu QD, Blankson S, Bouchier-Hayes D, Redmond HP. Endotoxin/lipopolysaccharide activates NF-kappa B and enhances tumor cell adhesion and invasion through a beta 1 integrin-dependent mechanism. J Immunol. 2003;170(2):795–804. doi: 10.4049/jimmunol.170.2.795. [DOI] [PubMed] [Google Scholar]
- 90.Sandholm J, Kauppila JH, Pressey C, et al. Estrogen receptor-α and sex steroid hormones regulate toll-like receptor-9 expression and invasive function in human breast cancer cells. Breast Cancer Res Treat. 2012;132(2):411–419. doi: 10.1007/s10549-011-1590-3. [DOI] [PubMed] [Google Scholar]
- 91.Das S, Sarrou E, Podgrabinska S, et al. Tumor cell entry into the lymph node is controlled by CCL1 chemokine expressed by lymph node lymphatic sinuses. J Exp Med. 2013;210(8):1509–1528. doi: 10.1084/jem.20111627. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Ben-Baruch A. Organ selectivity in metastasis: regulation by chemokines and their receptors. Clin Exp Metastasis. 2008;25(4):345–356. doi: 10.1007/s10585-007-9097-3. [DOI] [PubMed] [Google Scholar]
- 93.Monteleone I, Platt AM, Jaensson E, Agace WW, Mowat AM. IL-10-dependent partial refractoriness to toll-like receptor stimulation modulates gut mucosal dendritic cell function. Eur J Immunol. 2008;38(6):1533–1547. doi: 10.1002/eji.200737909. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Fusi A, Liu Z, Kümmerlen V, Nonnemacher A, Jeske J, Keilholz U. Expression of chemokine receptors on circulating tumor cells in patients with solid tumors. J Transl Med. 2012;10:52. doi: 10.1186/1479-5876-10-52. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Paschos KA, Canovas D, Bird NC. The role of cell adhesion molecules in the progression of colorectal cancer and the development of liver metastasis. Cell Signal. 2009;21(5):665–674. doi: 10.1016/j.cellsig.2009.01.006. [DOI] [PubMed] [Google Scholar]
- 96.Nimmagadda S. Differential expression of chemokine receptors and their roles in cancer imaging. Front Oncol. 2012;2:46. doi: 10.3389/fonc.2012.00046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Ghadjar P, Coupland SE, Na IK, et al. Chemokine receptor CCR6 expression level and liver metastases in colorectal cancer. J Clin Oncol. 2006;24(12):1910–1916. doi: 10.1200/JCO.2005.04.1822. [DOI] [PubMed] [Google Scholar]
- 98.Iwasa S, Yanagawa T, Fan J, Katoh R. Expression of CXCR4 and its ligand SDF-1 in intestinal-type gastric cancer is associated with lymph node and liver metastasis. Anticancer Res. 2009;29(11):4751–4758. [PubMed] [Google Scholar]
- 99.Lefrançois L, Parker CM, Olson S, et al. The role of beta7 integrins in CD8 T cell trafficking during an antiviral immune response. J Exp Med. 1999;189(10):1631–1638. doi: 10.1084/jem.189.10.1631. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Erreni M, Bianchi P, Laghi L, et al. Expression of chemokines and chemokine receptors in human colon cancer. Methods Enzymol. 2009;460:105–121. doi: 10.1016/S0076-6879(09)05205-7. [DOI] [PubMed] [Google Scholar]
- 101.Strell C, Entschladen F. Extravasation of leukocytes in comparison to tumor cells. Cell Commun Signal. 2008;6:10. doi: 10.1186/1478-811X-6-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Vidal-Vanaclocha F, Fantuzzi G, Mendoza L, et al. IL-18 regulates IL-1beta-dependent hepatic melanoma metastasis via vascular cell adhesion molecule-1. Proc Natl Acad Sci U S A. 2000;97(2):734–739. doi: 10.1073/pnas.97.2.734. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Ridnour LA, Cheng RY, Switzer CH, et al. Molecular pathways: toll-like receptors in the tumor microenvironment—poor prognosis or new therapeutic opportunity. Clin Cancer Res. 2013;19(6):1340–1346. doi: 10.1158/1078-0432.CCR-12-0408. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Ray A, Chakraborty K, Ray P. Immunosuppressive MDSCs induced by TLR signaling during infection and role in resolution of inflammation. Front Cell Infect Microbiol. 2013;3:52. doi: 10.3389/fcimb.2013.00052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Jin Y, Wi HJ, Choi MH, Hong ST, Bae YM. Regulation of anti-inflammatory cytokines IL-10 and TGF-β in mouse dendritic cells through treatment with Clonorchis sinensis crude antigen. Exp Mol Med. 2014;46:e74. doi: 10.1038/emm.2013.144. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Huang B, Zhao J, Li H, et al. Toll-like receptors on tumor cells facilitate evasion of immune surveillance. Cancer Res. 2005;65(12):5009–5014. doi: 10.1158/0008-5472.CAN-05-0784. [DOI] [PubMed] [Google Scholar]
- 107.Yu L, Wang L, Chen S. Dual character of toll-like receptor signaling: pro-tumorigenic effects and anti-tumor functions. Biochim Biophys Acta. 2013;1835(2):144–154. doi: 10.1016/j.bbcan.2012.10.006. [DOI] [PubMed] [Google Scholar]
- 108.Qian Y, Deng J, Geng L, et al. TLR4 signaling induces B7-H1 expression through MAPK pathways in bladder cancer cells. Cancer Invest. 2008;26(8):816–821. doi: 10.1080/07357900801941852. [DOI] [PubMed] [Google Scholar]
- 109.Pulko V, Liu X, Krco CJ, et al. TLR3-stimulated dendritic cells up-regulate B7-H1 expression and influence the magnitude of CD8 T cell responses to tumor vaccination. J Immunol. 2009;183(6):3634–3641. doi: 10.4049/jimmunol.0900974. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Saijo Y, Tanaka M, Miki M, et al. Proinflammatory cytokine IL-1 beta promotes tumor growth of Lewis lung carcinoma by induction of angiogenic factors: in vivo analysis of tumor-stromal interaction. J Immunol. 2002;169(1):469–475. doi: 10.4049/jimmunol.169.1.469. [DOI] [PubMed] [Google Scholar]
- 111.Carmi Y, Dotan S, Rider P, et al. The role of IL-1β in the early tumor cell-induced angiogenic response. J Immunol. 2013;190(7):3500–3509. doi: 10.4049/jimmunol.1202769. [DOI] [PubMed] [Google Scholar]
- 112.Lewis AM, Varghese S, Xu H, Alexander HR. Interleukin-1 and cancer progression: the emerging role of interleukin-1 receptor antagonist as a novel therapeutic agent in cancer treatment. J Transl Med. 2006;4:48. doi: 10.1186/1479-5876-4-48. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Dunn JH, Ellis LZ, Fujita M. Inflammasomes as molecular mediators of inflammation and cancer: potential role in melanoma. Cancer Lett. 2012;314(1):24–33. doi: 10.1016/j.canlet.2011.10.001. [DOI] [PubMed] [Google Scholar]
- 114.Weng PH, Huang YL, Page JH, et al. Polymorphisms of an innate immune gene, toll-like receptor 4, and aggressive prostate cancer risk: a systematic review and meta-analysis. PLoS One. 2014;9(10):e110569. doi: 10.1371/journal.pone.0110569. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.González-Reyes S, Fernández JM, González LO, et al. Study of TLR3, TLR4, and TLR9 in prostate carcinomas and their association with biochemical recurrence. Cancer Immunol Immunother. 2011;60(2):217–226. doi: 10.1007/s00262-010-0931-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Maker AV, Katabi N, Qin LX, et al. Cyst fluid interleukin-1beta (IL1beta) levels predict the risk of carcinoma in intraductal papillary mucinous neoplasms of the pancreas. Clin Cancer Res. 2011;17(6):1502–1508. doi: 10.1158/1078-0432.CCR-10-1561. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Scheede-Bergdahl C, Watt HL, Trutschnigg B, et al. Is IL-6 the best pro-inflammatory biomarker of clinical outcomes of cancer cachexia? Clin Nutr. 2012;31(1):85–88. doi: 10.1016/j.clnu.2011.07.010. [DOI] [PubMed] [Google Scholar]
- 118.Orengo AM, Fabbi M, Miglietta L, et al. Interleukin (IL)-18, a biomarker of human ovarian carcinoma, is predominantly released as biologically inactive precursor. Int J Cancer. 2011;129(5):1116–1125. doi: 10.1002/ijc.25757. [DOI] [PubMed] [Google Scholar]
- 119.Weber AN, Försti A. Toll-like receptor genetic variants and colorectal cancer. Oncoimmunology. 2014;3(1):e27763. doi: 10.4161/onci.27763. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Zhou XX, Jia WH, Shen GP, et al. Sequence variants in toll-like receptor 10 are associated with nasopharyngeal carcinoma risk. Cancer Epidemiol Biomarkers Prev. 2006;15(5):862–866. doi: 10.1158/1055-9965.EPI-05-0874. [DOI] [PubMed] [Google Scholar]
- 121.Dinarello CA. Why not treat human cancer with interleukin-1 blockade? Cancer Metastasis Rev. 2010;29(2):317–329. doi: 10.1007/s10555-010-9229-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Zhernakova A, Festen EM, Franke L, et al. Genetic analysis of innate immunity in Crohn’s disease and ulcerative colitis identifies two susceptibility loci harboring CARD9 and IL18RAP. Am J Hum Genet. 2008;82(5):1202–1210. doi: 10.1016/j.ajhg.2008.03.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Villani AC, Lemire M, Fortin G, et al. Common variants in the NLRP3 region contribute to Crohn’s disease susceptibility. Nat Genet. 2009;41(1):71–76. doi: 10.1038/ng285. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Fort MM, Mozaffarian A, Stöver AG, et al. A synthetic TLR4 antagonist has anti-inflammatory effects in two murine models of inflammatory bowel disease. J Immunol. 2005;174(10):6416–6423. doi: 10.4049/jimmunol.174.10.6416. [DOI] [PubMed] [Google Scholar]
- 125.Ungaro R, Fukata M, Hsu D, et al. A novel toll-like receptor 4 antagonist antibody ameliorates inflammation but impairs mucosal healing in murine colitis. Am J Physiol Gastrointest Liver Physiol. 2009;296(6):G1167–G1179. doi: 10.1152/ajpgi.90496.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Fukata M, Shang L, Santaolalla R, et al. Constitutive activation of epithelial TLR4 augments inflammatory responses to mucosal injury and drives colitis-associated tumorigenesis. Inflamm Bowel Dis. 2011;17(7):1464–1473. doi: 10.1002/ibd.21527. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Raja SG, Dreyfus GD. Eritoran: the evidence of its therapeutic potential in sepsis. Core Evid. 2008;2(3):199–207. [PMC free article] [PubMed] [Google Scholar]
- 128.Fukata M, Arditi M. The role of pattern recognition receptors in intestinal inflammation. Mucosal Immunol. 2013;6(3):451–463. doi: 10.1038/mi.2013.13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Hayakawa F, Sugimoto K, Harada Y, et al. A novel STAT inhibitor, OPB-31121, has a significant antitumor effect on leukemia with STAT-addictive oncokinases. Blood Cancer J. 2013;3:e166. doi: 10.1038/bcj.2013.63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Brambilla L, Genini D, Laurini E, et al. Hitting the right spot: mechanism of action of OPB-31121, a novel and potent inhibitor of the signal transducer and activator of transcription 3 (STAT3) Mol Oncol. 2015;9(6):1194–1206. doi: 10.1016/j.molonc.2015.02.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Watanabe T, Asano N, Murray PJ, et al. Muramyl dipeptide activation of nucleotide-binding oligomerization domain 2 protects mice from experimental colitis. J Clin Invest. 2008;118(2):545–559. doi: 10.1172/JCI33145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Lamkanfi M, Mueller JL, Vitari AC, et al. Glyburide inhibits the cryopyrin/Nalp3 inflammasome. J Cell Biol. 2009;187(1):61–70. doi: 10.1083/jcb.200903124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Dinarello CA, Simon A, van der Meer JW. Treating inflammation by blocking interleukin-1 in a broad spectrum of diseases. Nat Rev Drug Discov. 2012;11(8):633–652. doi: 10.1038/nrd3800. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Mallat Z, Corbaz A, Scoazec A, et al. Interleukin-18/interleukin-18 binding protein signaling modulates atherosclerotic lesion development and stability. Circ Res. 2001;89(7):E41–E45. doi: 10.1161/hh1901.098735. [DOI] [PubMed] [Google Scholar]
- 135.Wang JQ, Jeelall YS, Ferguson LL, Horikawa K. Toll-like receptors and cancer: MYD88 mutation and inflammation. Front Immunol. 2014;5:367. doi: 10.3389/fimmu.2014.00367. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Loiarro M, Ruggiero V, Sette C. Targeting TLR/IL-1R signalling in human diseases. Mediators Inflamm. 2010;2010:674363. doi: 10.1155/2010/674363. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Kim YS, Schwabe RF, Qian T, Lemasters JJ, Brenner DA. TRAIL-mediated apoptosis requires NF-kappaB inhibition and the mitochondrial permeability transition in human hepatoma cells. Hepatology. 2002;36(6):1498–1508. doi: 10.1053/jhep.2002.36942. [DOI] [PubMed] [Google Scholar]
- 138.O’Neill LA. A critical role for citrate metabolism in LPS signalling. Biochem J. 2011;438(3):e5–e6. doi: 10.1042/BJ20111386. [DOI] [PubMed] [Google Scholar]
- 139.Mills E, O’Neill LA. Succinate: a metabolic signal in inflammation. Trends Cell Biol. 2014;24(5):313–320. doi: 10.1016/j.tcb.2013.11.008. [DOI] [PubMed] [Google Scholar]
- 140.Singh S, Oberoi R, Khanna S, Bhatt AN, Saluja D, Dwarakanath BS. Energy restriction caused by the glycolytic inhibitor, 2-deoxy-D-glucose inhibits chemical induced carcinogenesis in mice. Cancer Res. 2012;72(8 suppl):3229–3229. [Google Scholar]
- 141.Singh S, Chawla AS, Pandey S, Soni R, Saluja D, Dwarakanath BS. Inhibition of radiation induced angiogenesis by 2-deoxy-D-glucose. J Cancer Res Therap. 2012;8(3):476. [Google Scholar]
- 142.Singh S, Chawla AS, Pandey S, Soni R, Saluja D, Dwarakanath BS. Glycolytic inhibitor 2-deoxy-D-glucose (2-DG) inhibits tumor associated angiogenesis. Cancer Res Therap. 2012;8(3):486. [Google Scholar]
- 143.Farooque A, Singh S, Verma A, Afrin F, Adhikari JS, Dwarakanath BS. Activation of macrophages by the glycolytic inhibitor 2-deoxy-D-glucose (2-DG): implications for tumor therapy. Cancer Res. 2009;69:3176. [Google Scholar]
- 144.Scheeren FA, Kuo AH, van Weele LJ, et al. A cell-intrinsic role for TLR2-MYD88 in intestinal and breast epithelia and oncogenesis. Nat Cell Biol. 2014;16(12):1238–1248. doi: 10.1038/ncb3058. [DOI] [PubMed] [Google Scholar]
- 145.Sheyhidin I, Nabi G, Hasim A, et al. Overexpression of TLR3, TLR4, TLR7 and TLR9 in esophageal squamous cell carcinoma. World J Gastroenterol. 2011;17(32):3745–3751. doi: 10.3748/wjg.v17.i32.3745. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Wang YH, Cao YW, Yang XC, et al. Effect of TLR4 and B7-H1 on immune escape of urothelial bladder cancer and its clinical significance. Asian Pac J Cancer Prev. 2014;15(3):1321–1326. doi: 10.7314/apjcp.2014.15.3.1321. [DOI] [PubMed] [Google Scholar]
- 147.Dapito DH, Mencin A, Gwak GY, et al. Promotion of hepatocellular carcinoma by the intestinal microbiota and TLR4. Cancer Cell. 2012;21:504–516. doi: 10.1016/j.ccr.2012.02.007. [DOI] [PMC free article] [PubMed] [Google Scholar]