

Abstract
This paper reviews the latest understanding of biological and pharmacological properties of osthole (7-methoxy-8-(3-methyl-2-butenyl)-2H-1-benzopyran-2-one), a natural product found in several medicinal plants such as Cnidium monnieri and Angelica pubescens. In vitro and in vivo experimental results have revealed that osthole demonstrates multiple pharmacological actions including neuroprotective, osteogenic, immunomodulatory, anticancer, hepatoprotective, cardiovascular protective, and antimicrobial activities. In addition, pharmacokinetic studies showed osthole uptake and utilization are fast and efficient in body. Moreover, the mechanisms of multiple pharmacological activities of osthole are very likely related to the modulatory effect on cyclic adenosine monophosphate (cAMP) and cyclic adenosine monophosphate (cGMP) level, though some mechanisms remain unclear. This review aims to summarize the pharmacological properties of osthole and give an overview of the underlying mechanisms, which showcase its potential as a multitarget alternative medicine.
1. Introduction
Osthole (also known as osthol), 7-methoxy-8-(3-methyl-2-butenyl)-2H-1-benzopyran-2-one, is a natural coumarin first derived from Cnidium plant (Figure 1). High content of osthole is found in the mature fruit of Cnidium monnieri (Fructus Cnidii), which is commonly applied in clinical practice of Traditional Chinese Medicine (TCM) (Figure 2), while it is also widely found in other medicinal plants including Angelica, Archangelica, Citrus, Clausena. Fructus Cnidii strengthens immune system and improves male function, relieving rheumatic pain and eliminating dampness; most of these medicinal properties are considered to attribute to one of its major bioactive components, osthole [1, 2]. Modern researches have suggested that osthole exhibits antioxidant, anticancer, anti-inflammatory, and immunomodulatory properties [1, 3, 4]. With multiple bioactivities of osthole reported, developing osthole and derivatives as potential multitarget drug should be encouraged. Therefore, it is meaningful to summarize the pharmacological and biological researches on this coumarin, to review the mechanisms behind the effects and get a comprehensive picture of its miscellaneous functions.
Figure 1.
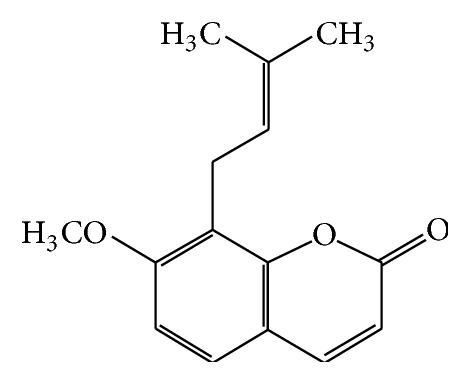
Chemical structure of osthole, the principle component of Cnidium monnieri.
Figure 2.

Illustration of the whole plant Cnidium monnieri with the fruit structure (a) and photo of the upper ground parts of the herb (b) (modified from http://www.google.com/).
2. Biological and Pharmacological Activities of Osthole
2.1. Nootropic and Neuroprotective Effect
The benefits of osthole and Fructus Cnidii (FC) extract on nervous system have been investigated in recent years. Osthole regulates ion channels and G protein-coupled receptor (GPCR) activities influencing neuronal and neuroendocrine function. Evidence suggested that osthole blocked L-type Ca2+ channel and Na+ channels in mouse neuronal cells [5, 6]. Osthole increased the affinity of thyrotropin-releasing hormone (TRH) receptor (one of GPCR), hence decreasing the binding of TRH to its receptor and suppressing TRH-evoked production of triphosphoinositol (IP3) and mobilization of sequestered Ca2+ in rat pituitary GH4C1 cells [7]. In addition, Wang et al. examined the effect of osthole and imperatorin (another coumarin isolated from FC) on glutamate release from rat hippocampal synaptosomes. The results suggested that both chemicals facilitated 4-aminopyridine- (4-AP-) evoked glutamate release by activating N-and P/Q-type Ca2+ channel through a signaling cascade involving protein kinase C (PKC) [8]. Lin et al. then suggested osthole-facilitated glutamate release was related to increased synaptic vesicle availability for exocytosis [9] and to activation of cGMP/PKG-dependent pathway [10]. Osthole was also found to reduce acid-sensing ion channel 3 (ASIC3) expression in rat dorsal root ganglion, which may contribute to its relieving chronic pain from lumbar disc herniation [11]. Moreover, Luszczki et al. reported that osthole showed anticonvulsant effect in maximal electroshock seizure models, suggesting its potential in seizure treatment [12, 13]. Osthole was identified as a modulator of the neurotransmitter gamma-aminobutyric acid (GABA)A receptor in vitro, which provided a possible mechanism explaining such antiseizure effect [14, 15].
On the other hand, osthole exhibited neuroprotective effects against neurodegeneration in both in vitro and in vivo experimental models. Pretreatment of osthole showed significant protective effect on viability of PC12 cells exposed to neurotoxin MPP+, an in vitro model of Parkinson's disease [16]. Moreover, multiple evidences have demonstrated the protective effect of osthole on alleviating brain damage and improving neurobehavioral functions caused by both chronic [17] and acute [18–20] ischemia due to its antioxidative and anti-inflammatory properties, through mitogen-activated protein kinase (MAPK) pathway by prolonged activation of ERK1/2 and suppression of JNK activity [20]. Osthole has also been suggested as a promising herbal component for memory loss therapy [21]. Animal experiments have been conducted in aluminium chloride- (AlCl3-) induced acute senile model [22] and scopolamine-induced amnesia model [23], and results from both studies demonstrated ameliorating effect on memory impairment. In addition, studies demonstrated that osthole was also effective in treating traumatic brain injury by significantly reducing neurological deficits, cerebral edema, and hippocampal neuron loss [24], as well as relieving spatial performance deficits in scopolamine- (SCOP-) treated or ovariectomized (OVX) rats [25], and attenuating autoimmune encephalomyelitis in mice [26].
2.2. Osteogenic Activity
Bone modeling effect is one of the bioactivities osthole showing most promising therapeutic potential. Plenty of in vitro studies have shown that osthole and coumarin extract from FC promoted proliferation and differentiation of osteoblasts [27–30] and suppress formation and activity of osteoclasts [31, 32], hence tipping the balance in favor of bone remodeling and increasing bone density, which makes osthole a potential agent for osteoporosis treatment. Findings from experiments in both ovariectomy and glucocorticoids-induced osteoporosis rat models supported that treatments with osthole and FC coumarin reduced osteoporotic bone loss [33–36] through estrogen-independent pathway, rather than phytoestrogens commonly found in medicinal herb [36]. Kuo et al. studied the mechanism of osthole-mediated cell differentiation in detail with human osteoblast MG-63 and hFOB. The results obtained after application of bone morphogenetic protein- (BMP-) 2 antagonists, p38 inhibitor, and mitogen-activated protein kinase (ERK2), respectively, indicated that BMP-2/p38 pathway was associated with early phase of osthole-induced osteoblast differentiation, whereas ERK2 was involved in later phase of cellular ossification [33]. A recent paper investigated the potential of osthole in treating and preventing bone loss due to estrogen deficiency and studied the underlying mechanism. In vitro findings revealed that osthole promoted osteoblast differentiation by activating Wnt/β-catenin signaling and subsequently increasing BMP2 expression. In vivo periosteal bone-formation assay by local injection of osthole indicated osthole promoted new bone formation; in addition, comparison of bone microarchitecture, histomorphometric parameters, and biomechanical properties in OVX rat treated with or without osthole revealed that treatment with osthole effectively prevented bone loss in OVX rats [37].
2.3. Immunomodulation and Anti-Inflammatory Activity
In late 1990s, a German research group first reported osthole's selective inhibitory effect on 5-lipoxygenase (5-LO) and cyclooxygenase- (COX-) 1 [38, 39]. Both enzymes are critical in the process of inflammation, and inhibition of COX is shown to relieve pain and inflammatory symptoms [40], whereas 5-LO is considered a target for pharmaceutical intervention in various kinds of diseases aside from inflammatory diseases, like cardiovascular diseases, cancer, and osteoporosis [41]. In cell culture, osthole suppressed the immune response of lipopolysaccharide-stimulated macrophages by decreasing reactive oxygen species (ROS) release and inhibiting enzymes including inducible nitric oxide synthase (iNOS), mitogen-activated protein kinases (MAPK), and COX-2 [42, 43]. Osthole also suppressed interleukin- (IL-) 4 and tumor necrosis factor- (TNF-) α induced eotaxin expression in bronchial epithelial BEAS-2B cells [44] and inhibited hypertrophic scar fibroblasts through inducing apoptosis and downregulating TGF-β expression [45]. Similar anti-inflammatory activates of osthole have also been found in rat peritoneal cells and human peripheral blood mononuclear cells [46]. Meanwhile, anti-inflammatory effect of osthole was also confirmed in animal models. Results from carrageenan-induced hind paw edema in rats suggested that osthole suppressed production of prostaglandin (PG), nitric oxide (NO), and malondialdehyde (MDA), as well as downregulated activity of NOS, which was likely to associate with the elevation of cyclic adenosine monophosphate (cAMP) and cyclic adenosine monophosphate (cGMP) levels [47]. Osthole also inhibited the expression of COX-2 and NOS in dorsal root ganglion and relieved mechanical allodynia in rat model of sciatica induced by lumbar disc herniation [48]. The chemical was also reported to have antiallergic effect in passive cutaneous anaphylaxis (PCA), 2, 4-dinitrofluorobenzene (DNFB), and picryl chloride- (PC-) induced contact dermatitis in experimental animals [49].
2.4. Anticancer Effect
Accumulating experimental evidences have shown that osthole exhibited antiproliferative properties and induced apoptosis in various kinds of tumor cells [2, 50], including leukemia HL-60 [51], L1210 [52]; prostatic cancer cell LNCaP, PC3, and DU145 [52]; cervical cancer cell Hela [53]; ovarian cancer cell SKOV3 [54]; lung cancer cell A549 [55]; SK-LU-1 [56]; epidermal cancer cell KB [56]; breast cancer cell MCF-7 [54, 56, 57], MDA-MB-231 [54, 57], and 4T1 [57]; and hepatocellular carcinoma (HCC) HepG2 [56], SMMC-7721, and Hepa1-6 [58]. The antitumor activities of osthole have also been supported with in vivo results showing prolonged survival days of P-388 D1 tumor-bearing CDF mice [53] and suppressed tumor growth in HCC tumor models established by injection of SMMC-7721 or Hepa1-6 cells [58]. It is noteworthy that osthole successfully inhibited the migration and invasion of metastatic cancer cells. Yang et al. firstly reported that osthole inhibited migration of MCF-7 cells and invasion of MDA-MB-231 cells and suggested that suppression of matrix metalloproteinases (MMP) enzyme activities might be the possible mechanism [57]. Prevention of cell migration and invasion was also reported in human lung adenocarcinoma CL1-5, H1299, and A549 [59]. Additionally, combination of osthole and other nature products showed synergetic effect on inhibition of tumor cell proliferation and invasion [60–62].
The molecular mechanism of anticancer effect of osthole still remains unclear. It is highly likely to be combinatory effects on carcinogenesis and tumor progression. Osthole has been suggested to modulate PI3K/Akt signaling pathway leading to G2/M arrest and apoptosis in lung cancer A549 cells [55]. It downregulates FASN, which is highly expressed in many solid tumors, in HER2-overexpressing breast cancer cells through inhibiting the c-Met/Akt/mTOR pathway [54, 63]. Osthole also normalized plasma alanine aminotransferase (ALT) which has been proved to be a strategy for preventing the development of HCC [64]. Moreover, osthole inhibited cancer cells invasion and transition through suppression of MMP-2 and MMP-9 which were induced by a serial of inflammatory factors [57, 59, 61, 62]. Besides, osthole was demonstrated as promising therapeutic agent for cancer treatment due to histone deacetylase inhibition [65, 66].
2.5. Hepatoprotective Effect and Benefits on Metabolic Diseases
Liver is regarded as one of the vital organs that functions as a center for metabolism of nutrients and excretion of wastes. Osthole exerts protective effects against hepatitis. It suppressed the secretion of hepatitis B virus (HBV) in cell culture [67] and prevented hepatitis in mice induced by concanavalin A or anti-Fas antibody [68–70]. Osthole also exhibited therapeutic effect on both hyperlipidemic [71–77] and alcoholic fatty liver animals [71, 75, 78, 79]. Mechanism studies revealed that osthole modulated expression of multiple lipogenic genes [77, 80], increased adiponectin release, and hence improved insulin resistance [76] via activation of PPARα/γ pathway. In addition, antifibrotic activity of osthole on HSC-T6 hepatic stellate cell lines was also reported [81].
Osthole is not only beneficial to liver but also beneficial to endocrine system and metabolism of the whole body. Screening for allosteric modulators of melanocortin-4 receptor, a target for obesity and cachexia therapy, reported osthole and other two FC coumarins as potential modulators [82]. Osthole is regarded as a potential antidiabetic agent as well. In vitro and in vivo experiments demonstrated that osthole alleviated hyperglycemia by activating PPARα/γ, AMP-activated protein kinase (AMPK), and downstream acetyl CoA carboxylase [83, 84]. In addition, the Yang tonifying effect of osthole has been explored in modern experimental way. Oral administration of osthole significantly increased androgen, gonadotropin production, and NOS activity in immature castrate male rats [85]. Administration of osthole and FC coumarin extract improved immunological function in kidney yang deficiency animal models induced by hydrocortisone acetate [86], via elevated function of pituitary-thyroid [87], and pituitary-adrenocortex axis [88].
2.6. Vasorelaxant Properties and Cardiovascular Benefits
Osthole exhibits protective effect on heart and circulatory system. Abnormal vascular smooth muscle cell proliferation is a major component of vascular disease including atherosclerosis, vein graft occlusion, and restenosis after angioplasty, whereas osthole treatment selectively inhibits the proliferation of those vascular smooth muscle cells [89]. Osthole showed vasorelaxant properties due to its Ca2+-channel antagonistic effect and upregulation of cGMP level in vascular smooth muscle [90–92]. Osthole was also proposed to prevent isoprenaline-induced fibrosis through activating of PPARα/γ and subsequent suppression of NF-κB production in myocardial tissues [93]. Additionally, accelerative effect on β-oxidation of hepatic fatty acids in hypertensive rats suggested it might be useful for prevention of atherosclerosis [80]. Osthole suppresses platelet aggregation via inhibition of thromboxane formation and phosphoinositides breakdown, thus making it a prospective antithrombotic agent [94, 95]. Osthole exerts relaxant effect not only on blood vessels, but also on other tissues such as isolated rodent ileum and taeniae coli [96]; trachea [97]; and corpus cavernosum, which partially explain long history of using FC as a herbal medicine to improve male sexual dysfunction [98, 99].
2.7. Antimicrobial and Antiparasitic Effect
Osthole exerts antifungal properties on numbers of fungi species. Experimental evidences showed that osthole inhibited hypha growth of Fusarium graminearum, a parasite found on common weeds and cereal crops, through glucose starvation [100], and controlled powdery mildew caused by Sphaerotheca fuliginea via inhibiting spore germination and mycelia growth [101]. Osthole derivatives exhibited curative effect on pepper blight caused by Phytophthora capsici [102]. Other antibacterial activities on both gram positive and gram negative bacteria were also reported [103, 104]. Osthole was also found to exhibit antiviral activity not only on HBV, but also on HIV-1 by inhibiting Rev-export, which is critical in HIV-1 entails replication [105]. In addition, anthelmintic activity of osthole was also noted in goldfish against Dactylogyrus intermedius [106, 107].
3. Pharmacokinetics and Metabolism of Osthole
Pharmacokinetics of osthole in rat plasma after oral or intravenous administration was studied using HPLC method, yielding concentration/time curve with rapid distribution followed by a slower elimination phase [108–111]. Intestinal absorption of osthole was studied with HPLC in rat single pass intestine perfusion (SPIP) model, where results showed osthole absorption was a passive diffusion process in whole intestinal sections [112]. Osthole metabolism after oral administration was studied in male SD rats and 10 phase I and 3 phase II metabolites were isolated and identified from urine. The major phase I metabolic reactions were hydroxylation, demethylation, and hydrogenation, while glucuronidation contributed to phase II metabolism [113]. Absorption and metabolism of osthole were also investigated in human colorectal Caco-2 cell model. Osthole demonstrated high absorptive permeability and accumulation in Caco-2 cells; major phase I metabolites were desmethyl-osthol and its multiple isomers [114, 115].
4. Discussion
4.1. Composite Bioactivities of Osthole on Body System
Osthole exerts a broad spectrum of biological and pharmacological activities. While effects of osthole are categorized under different biological activities in this paper, a lot of connections can actually be found among them (Figure 3). Osthole exhibits immunomodulatory and anti-inflammatory properties, by regulating the expression of a series of key factors, including TNF-α, NF-κB, TGF-β, COX, NO, ERK, and JNK, involved in the process of immune response and other metabolic and biological processes. For instance, NF-κB plays important role in modulating immunological response, and disturbance of NF-κB expression has been linked to some autoimmune diseases, cancer, and many other diseases. Likewise, TGF-β is a cytokine involving in several key pathways which are related to the development of numerous diseases. Modulation by osthole on those critical factors probably contributes to its various benefits on systemic level. Principally, in various tissues and organs, the anti-inflammatory and antioxidative activities of osthole induce and magnify its anticancer properties and protective effects on other systems. Similarly, osthole showed antifibrotic effect in both hepatic and myocardial tissues, which contribute to the protective effect on liver and cardiovascular system. Osthole has also been suggested as a natural activator of defective DeltaF508-cystic fibrosis transmembrane conductance regulator (CFTR) chloride channel gating and thus may be a lead compound for cystic fibrosis therapies [116, 117]. In addition, liver plays a critical role in metabolism including catabolism, storage, and anabolism. Hence, the numerous benefits on liver certainly contribute to better metabolic system of human body. Moreover, the positive influence of osthole to male sexual dysfunction has been found related to vasorelaxant activity which was also reported in other tissues such as ileum and taeniae coli and thoracic aorta.
Figure 3.

Multiple systemic pharmacological and beneficial effects and related experimental results.
4.2. cAMP and cGMP Level
Accumulating evidence in studies for biological activities of osthole revealed that osthole exerts a nonspecific elevation of intracellular and tissue cAMP and cGMP, which is likely involved in the underlying mechanism of some bioactivities of osthole. cAMP and cGMP, derived from ATP and GTP, respectively, are second messenger prominent in many biological processes. cAMP mainly works by activating protein kinase A (PKA), as well as directly affecting ion channels and growth hormone. cGMP is also a regulator of ion channels related to cell cycle arrest, apoptosis, and smooth muscle tissue relaxing as well. Osthole increases cAMP and cGMP level by inhibiting cAMP and cGMP phosphodiesterases (PDEs) which hydrolyze cAMP and cGMP by degradation of the phosphodiester bond [95, 97]. The inhibitory effect on vascular smooth muscle cell was found to associate with osthole-induced elevation of cAMP and cGMP level [89], whereas osthole-facilitated glutamate release in hippocampal synaptosomes was associated with activation of cGMP/PKG-dependent pathway [10]. Osthole-mediated suppression of inflammatory factors in carrageenan-induced model was noted to attribute to cGMP elevation [47]. Moreover, researchers suggested that vasorelaxant property of osthole was linked to increased cAMP and cGMP levels caused by osthole treatment [92, 97, 99] (Figure 4).
Figure 4.

Possible interrelation between effect of osthole on intracellular ion channels, cyclic adenosine monophosphate (cAMP), and cyclic guanosine monophosphate (cGMP) levels with some of its pharmacological activities. (Hypotheses unconfirmed in studies of osthole are indicated with dashed line.)
4.3. Ion Channel Regulator
Osthole has been found to influence membrane potential and some types of ion channels in various cells and tissues, including sodium channel, acid-sensing ion channel, chloride channel of CFTR, and especially calcium channel. Effect of osthole on these ion channels is closely related to bioactivities of osthole as vasorelaxant, immunomodulatory, antifibrotic effects, and, in particular, its neuronal and neuroendocrine functions including neuroprotective, anticonvulsant, and pain relieving properties. On the other hand, calcium concentration regulates the osteoprogenitors behavior which play important roles in both bone homeostasis and regeneration [117, 118], thereby making calcium channel a prospective direction to study the mechanism of osteogenic effect of osthole. cAMP and cGMP have been well documented as important regulators of L-type Ca2+ channel and some other ion channels [119–121]. Therefore, effect of osthole on ion channels is at least partially induced by modulation of cAMP and cGMP level (Figure 4).
4.4. 5-Lipoxygenase Inhibitor
5-lipoxygenase (5-LO) is a rate-limiting dioxygenase in the process of leukotrienes (LTs) biosynthesis from the precursor arachidonic acid. As LTs are key mediators in immune and inflammatory responses in pathophysiology of numbers of respiratory and cardiovascular diseases, 5-LO is regarded as a target in developing therapy of related diseases and 5-LO inhibitors are being developed as a treatment approach. Osthole has been reported to be a 5-LO inhibitor in previous studies. Meanwhile, researchers proposed that signaling through cAMP/PKA results in phosphorylation and cytoplasmic sequestration of 5-LO and inhibition of LTs synthesis. It has been found that many cAMP-elevating agents such as isoproterenol, prostaglandin E, and prostaglandins attenuate 5-LO translocation and LTs biosynthesis [122–124]. Therefore, inhibitory effect of osthole on 5-LO may also be associated with osthole-mediated cAMP elevation.
5. Conclusion
Plenty of experimental results demonstrated that osthole exhibits a variety of pharmacological benefits including neuroprotection, osteogenesis, immunomodulation, and cancer-combating properties, making it a potential multitarget complementary medicine and functional food. The mechanisms underlying these properties have not been fully understood, yet the regulatory effect of osthole on cAMP and cGMP level and some ion channels can be seen as contributing to several of those properties. Further studies are needed to enrich the knowledge of the pharmacological effects and address the safety issues of osthole in order to develop this natural product and its derivatives as an agent for disease prevention and therapeutics in human.
Acknowledgments
This project is supported by General Research Fund, Hong Kong Research Grant Council (ref. no. 461113) and Research Committee Funding (Direct Grants) (ref. nos. 4053024 and 2030445).
Conflict of Interests
The authors declare that there is no conflict of interests regarding the publication of this paper.
References
- 1.You L., Feng S., An R., Wang X. Osthole: a promising lead compound for drug discovery from a traditional Chinese medicine (TCM) Natural Product Communications. 2009;4(2):297–302. [PubMed] [Google Scholar]
- 2.Zhou Z.-W., Liu P.-X. Progress in study of chemical constituents and anti-tumor activities of Cnidium monnieri . Zhongguo Zhongyao Zazhi. 2005;30(17):1309–1313. [PubMed] [Google Scholar]
- 3.Hoult J. R. S., Payá M. Pharmacological and biochemical actions of simple coumarins: natural products with therapeutic potential. General Pharmacology. 1996;27(4):713–722. doi: 10.1016/0306-3623(95)02112-4. [DOI] [PubMed] [Google Scholar]
- 4.Wang R., Kong J., Wang D., Lien L. L.-M., Lien E. J.-C. A survey of Chinese herbal ingredients with liver protection activities. Chinese Medicine. 2007;2, article 5 doi: 10.1186/1749-8546-2-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Wu S.-N., Lo Y.-K., Chen C.-C., Li H.-F., Chiang H.-T. Inhibitory effect of the plant-extract osthole on L-type calcium current in NG108-15 neuronal cells. Biochemical Pharmacology. 2002;63(2):199–206. doi: 10.1016/S0006-2952(01)00873-5. [DOI] [PubMed] [Google Scholar]
- 6.Leung Y.-M., Kuo Y.-H., Chao C.-C., et al. Osthol is a use-dependent blocker of voltage-gated Na+ channels in mouse neuroblastoma N2A cells. Planta Medica. 2010;76(1):34–40. doi: 10.1055/s-0029-1185941. [DOI] [PubMed] [Google Scholar]
- 7.Ojala T., Vuorela P., Vuorela H., Törnquist K. The coumarin osthol attenuates the binding of thyrotropin-releasing hormone in rat pituitary GH4C1 cells. Planta Medica. 2001;67(3):236–239. doi: 10.1055/s-2001-12010. [DOI] [PubMed] [Google Scholar]
- 8.Wang S.-J., Lin T.-Y., Lu C.-W., Huang W.-J. Osthole and imperatorin, the active constituents of Cnidium monnieri (L.) Cusson, facilitate glutamate release from rat hippocampal nerve terminals. Neurochemistry International. 2008;53(6–8):416–423. doi: 10.1016/j.neuint.2008.09.013. [DOI] [PubMed] [Google Scholar]
- 9.Lin T.-Y., Lu C.-W., Huang W.-J., Wang S.-J. Osthole or imperatorin-mediated facilitation of glutamate release is associated with a synaptic vesicle mobilization in rat hippocampal glutamatergic nerve endings. Synapse. 2010;64(5):390–396. doi: 10.1002/syn.20738. [DOI] [PubMed] [Google Scholar]
- 10.Lin T. Y., Lu C. W., Huang W.-J., Wang S.-J. Involvement of the cGMP pathway in the osthole-facilitated glutamate release in rat hippocampal nerve endings. Synapse. 2012;66(3):232–239. doi: 10.1002/syn.21505. [DOI] [PubMed] [Google Scholar]
- 11.He Q.-L., Chen Y., Qin J., et al. Osthole, a herbal compound, alleviates nucleus pulposus-evoked nociceptive responses through the suppression of overexpression of acid-sensing ion channel 3 (ASIC3) in rat dorsal root ganglion. Medical Science Monitor. 2012;18(6):BR229–BR236. doi: 10.12659/MSM.882899. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Luszczki J. J., Wojda E., Andres-Mach M., et al. Anticonvulsant and acute neurotoxic effects of imperatorin, osthole and valproate in the maximal electroshock seizure and chimney tests in mice: a comparative study. Epilepsy Research. 2009;85(2-3):293–299. doi: 10.1016/j.eplepsyres.2009.03.027. [DOI] [PubMed] [Google Scholar]
- 13.Luszczki J. J., Andres-Mach M., Cisowski W., Mazol I., Glowniak K., Czuczwar S. J. Osthole suppresses seizures in the mouse maximal electroshock seizure model. European Journal of Pharmacology. 2009;607(1–3):107–109. doi: 10.1016/j.ejphar.2009.02.022. [DOI] [PubMed] [Google Scholar]
- 14.Zaugg J., Eickmeier E., Rueda D. C., Hering S., Hamburger M. HPLC-based activity profiling of Angelica pubescens roots for new positive GABAA receptor modulators in Xenopus oocytes. Fitoterapia. 2011;82(3):434–440. doi: 10.1016/j.fitote.2010.12.001. [DOI] [PubMed] [Google Scholar]
- 15.Singhuber J., Baburin I., Ecker G. F., Kopp B., Hering S. Insights into structure–activity relationship of GABAA receptor modulating coumarins and furanocoumarins. European Journal of Pharmacology. 2011;668(1-2):57–64. doi: 10.1016/j.ejphar.2011.06.034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Liu W.-B., Zhou J., Qu Y., et al. Neuroprotective effect of osthole on MPP+-induced cytotoxicity in PC12 cells via inhibition of mitochondrial dysfunction and ROS production. Neurochemistry International. 2010;57(3):206–215. doi: 10.1016/j.neuint.2010.05.011. [DOI] [PubMed] [Google Scholar]
- 17.Ji H.-J., Hu J.-F., Wang Y.-H., Chen X.-Y., Zhou R., Chen N.-H. Osthole improves chronic cerebral hypoperfusion induced cognitive deficits and neuronal damage in hippocampus. European Journal of Pharmacology. 2010;636(1–3):96–101. doi: 10.1016/j.ejphar.2010.03.038. [DOI] [PubMed] [Google Scholar]
- 18.Chao X., Zhou J., Chen T., et al. Neuroprotective effect of osthole against acute ischemic stroke on middle cerebral ischemia occlusion in rats. Brain Research. 2010;1363:206–211. doi: 10.1016/j.brainres.2010.09.052. [DOI] [PubMed] [Google Scholar]
- 19.Mao X., Yin W., Liu M., et al. Osthole, a natural coumarin, improves neurobehavioral functions and reduces infarct volume and matrix metalloproteinase-9 activity after transient focal cerebral ischemia in rats. Brain Research. 2011;1385:275–280. doi: 10.1016/j.brainres.2011.02.015. [DOI] [PubMed] [Google Scholar]
- 20.Chen T., Liu W., Chao X., et al. Neuroprotective effect of osthole against oxygen and glucose deprivation in rat cortical neurons: involvement of mitogen-activated protein kinase pathway. Neuroscience. 2011;183:203–211. doi: 10.1016/j.neuroscience.2011.03.038. [DOI] [PubMed] [Google Scholar]
- 21.Wu L., Cheng S.-Y., Wang Q., Chen Y.-B. Advances in study on the pharmacological effects of active components of Chinese herbs on Alzheimer's disease. Zhongguo Zhongyao Zazhi. 2004;29(5):388–389. [PubMed] [Google Scholar]
- 22.Shen L.-X., Jin L.-Q., Zhang D.-S., Xue G.-P. Effect of osthol on memory impairment of mice in AlCl3-induced acute senile model. Yao Xue Xue Bao. 2002;37(3):178–180. [PubMed] [Google Scholar]
- 23.Wu C.-R., Lin L.-W., Hsieh C.-L., Wang W.-H., Lin Y.-T., Hsieh M.-T. Petroleum ether extract of Cnidium monnieri ameliorated scopolamine-induced amnesia through adrenal gland-mediated mechanism in male rats. Journal of Ethnopharmacology. 2008;117(3):403–407. doi: 10.1016/j.jep.2008.02.013. [DOI] [PubMed] [Google Scholar]
- 24.He Y., Qu S., Wang J., et al. Neuroprotective effects of osthole pretreatment against traumatic brain injury in rats. Brain Research. 2012;1433:127–136. doi: 10.1016/j.brainres.2011.11.027. [DOI] [PubMed] [Google Scholar]
- 25.Hsieh M.-T., Hsieh C.-L., Wang W.-H., Chen C.-S., Lin C.-J., Wu C.-R. Osthole improves aspects of spatial performance in ovariectomized rats. American Journal of Chinese Medicine. 2004;32(1):11–20. doi: 10.1142/S0192415X04001758. [DOI] [PubMed] [Google Scholar]
- 26.Chen X., Pi R., Zou Y., et al. Attenuation of experimental autoimmune encephalomyelitis in C57 BL/6 mice by osthole, a natural coumarin. European Journal of Pharmacology. 2010;629(1–3):40–46. doi: 10.1016/j.ejphar.2009.12.008. [DOI] [PubMed] [Google Scholar]
- 27.Zhang W., Ma D., Zhao Q., Ishida T. The effect of the major components of Fructus Cnidii on osteoblasts in vitro. JAMS Journal of Acupuncture and Meridian Studies. 2010;3(1):32–37. doi: 10.1016/s2005-2901(10)60005-2. [DOI] [PubMed] [Google Scholar]
- 28.Ming L. G., Ge B. F., Chen K. M., et al. Effect of Osthol on the proliferation and differentiate of osteoblasts in vitro. Zhongguo Gu Shang. 2010;23(9):688–691. [PubMed] [Google Scholar]
- 29.Qin L.-P., Zhang Q.-Y., Tian Y.-P., Zheng H.-C., Huang M., Huang B.-K. Total coumarins from fruits of Cnidium monnieri inhibit formation and differentiation of multinucleated osteoclasts of rats. Acta Pharmacologica Sinica. 2003;24(2):181–186. [PubMed] [Google Scholar]
- 30.Ming L.-G., Zhou J., Cheng G.-Z., Ma H.-P., Chen K.-M. Osthol, a coumarin isolated from common cnidium fruit, enhances the differentiation and maturation of osteoblasts in vitro . Pharmacology. 2011;88(1-2):33–43. doi: 10.1159/000328776. [DOI] [PubMed] [Google Scholar]
- 31.Zhang Q., Qin L., He W., et al. Coumarins from Cnidium monnieri and their antiosteoporotic activity. Planta Medica. 2007;73(1):13–19. doi: 10.1055/s-2006-951724. [DOI] [PubMed] [Google Scholar]
- 32.Ming L.-G., Wang M.-G., Chen K.-M., Zhou J., Han G.-Q., Zhu R.-Q. Effect of osthol on apoptosis and bone resorption of osteoclasts cultured in vitro . Yaoxue Xuebao. 2012;47(2):174–179. [PubMed] [Google Scholar]
- 33.Li Q. N., Liang N. C., Wu T., et al. Effects of total coumarins of Fructus cnidii on skeleton of ovariectomized rats. Zhongguo Yao Li Xue Bao. 1994;15(6):528–532. [PubMed] [Google Scholar]
- 34.Li C. Y., Wu T., Li Q. N., et al. Quantitative study on the effect of osthole on proximal tibiae in ovariectomized(OVX) rats. Yaoxue Xuebao. 1996;31(5):327–332. [PubMed] [Google Scholar]
- 35.Liao J.-M., Zhu Q.-A., Lu H.-J., Li Q.-N., Wu T., Huang L.-F. Effects of total coumarins of Cnidium monnieri on bone density and biomechanics of glucocorticoids-induced osteoporosis in rats. Acta Pharmacologica Sinica. 1997;18(6):519–521. [PubMed] [Google Scholar]
- 36.Xiao X. L., Hara I., Matsumiya T. Effects of osthole on postmenopausal osteoporosis using ovariectomized rats; comparison to the effects of estradiol. Biological & Pharmaceutical Bulletin. 2002;25(6):738–742. doi: 10.1248/bpb.25.738. [DOI] [PubMed] [Google Scholar]
- 37.Tang D.-Z., Hou W., Zhou Q., et al. Osthole stimulates osteoblast differentiation and bone formation by activation of beta-catenin-BMP signaling. Journal of Bone and Mineral Research. 2010;25(6):1234–1245. doi: 10.1002/jbmr.21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Liu J. H., Zschocke S., Reininger E., Bauer R. Inhibitory effects of Angelica pubescens f. biserrata on 5-lipoxygenase and cyclooxygenase. Planta Medica. 1998;64(6):525–529. doi: 10.1055/s-2006-957507. [DOI] [PubMed] [Google Scholar]
- 39.Resch M., Steigel A., Chen Z.-L., Bauer R. 5-lipoxygenase and cyclooxygenase-1 inhibitory active compounds from Atractylodes lancea . Journal of Natural Products. 1998;61(3):347–350. doi: 10.1021/np970430b. [DOI] [PubMed] [Google Scholar]
- 40.Seibert K., Zhang Y., Leahy K., et al. Pharmacological and biochemical demonstration of the role of cyclooxygenase 2 in inflammation and pain. Proceedings of the National Academy of Sciences of the United States of America. 1994;91(25):12013–12017. doi: 10.1073/pnas.91.25.12013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Pergola C., Werz O. 5-Lipoxygenase inhibitors: a review of recent developments and patents. Expert Opinion on Therapeutic Patents. 2010;20(3):355–375. doi: 10.1517/13543771003602012. [DOI] [PubMed] [Google Scholar]
- 42.Nakamura T., Kodama N., Arai Y., et al. Inhibitory effect of oxycoumarins isolated from the Thai medicinal plant Clausena guillauminii on the inflammation mediators, iNOS, TNF-alpha, and COX-2 expression in mouse macrophage RAW 264.7. Journal of Natural Medicines. 2009;63(1):21–27. doi: 10.1007/s11418-008-0277-5. [DOI] [PubMed] [Google Scholar]
- 43.Liao P.-C., Chien S.-C., Ho C.-L., et al. Osthole regulates inflammatory mediator expression through modulating NF-KB, mitogen-activated protein kinases, protein kinase C, and reactive oxygen species. Journal of Agricultural and Food Chemistry. 2010;58(19):10445–10451. doi: 10.1021/jf102812t. [DOI] [PubMed] [Google Scholar]
- 44.Chiu P.-R., Lee W.-T., Chu Y.-T., Lee M.-S., Jong Y.-J., Hung C.-H. Effect of the Chinese herb extract osthol on IL-4-induced eotaxin expression in BEAS-2B cells. Pediatrics and Neonatology. 2008;49(4):135–140. doi: 10.1016/s1875-9572(08)60028-5. [DOI] [PubMed] [Google Scholar]
- 45.Hou X.-H., Cao B., Liu H.-Q., Wang Y.-Z., Bai S.-F., Chen H. Effects of osthole on apoptosis and TGF-β1 of hypertrophic scar fibroblasts. Journal of Asian Natural Products Research. 2009;11(7):663–669. doi: 10.1080/10286020902975772. [DOI] [PubMed] [Google Scholar]
- 46.Zimecki M., Artym J., Cisowski W., Mazol I., Włodarczyk M., Gleńsk M. Immunomodulatory and anti-inflammatory activity of selected osthole derivatives. Zeitschrift fur Naturforschung—Section C. 2009;64(5-6):361–368. doi: 10.1515/znc-2009-5-610. [DOI] [PubMed] [Google Scholar]
- 47.Liu J., Zhang W., Zhou L., Wang X., Lian Q. Anti-inflammatory effect and mechanism of osthole in rats. Journal of Chinese Medicinal Materials. 2005;28(11):1002–1006. [PubMed] [Google Scholar]
- 48.Wei M., Zhang J.-J., He Q.-L., et al. Effects and mechanisms of osthole on sciatica induced by lumber disc herniation. Zhong Yao Cai. 2011;34(5):746–750. [PubMed] [Google Scholar]
- 49.Matsuda H., Tomohiro N., Ido Y., Kubo M. Anti-allergic effects of cnidii monnieri fructus (dried fruits of Cnidium monnieri) and its major component, osthol. Biological & Pharmaceutical Bulletin. 2002;25(6):809–812. doi: 10.1248/bpb.25.809. [DOI] [PubMed] [Google Scholar]
- 50.Kawaii S., Tomono Y., Ogawa K., et al. Antiproliferative effect of isopentenylated coumarins on several cancer cell lines. Anticancer Research. 2001;21(3):1905–1911. [PubMed] [Google Scholar]
- 51.Yang L.-L., Wang M.-C., Chen L.-G., Wang C.-C. Cytotoxic activity of coumarins from the fruits of Cnidium monnieri on leukemia cell lines. Planta Medica. 2003;69(12):1091–1095. doi: 10.1055/s-2003-45188. [DOI] [PubMed] [Google Scholar]
- 52.Riviere C., Goossens L., Pommery N., Fourneau C., Delelis A., Henichart J. P. Antiproliferative effects of isopentenylated coumarins isolated from Phellolophium madagascariense Baker. Natural Product Research. 2006;20(10):909–916. doi: 10.1080/14786410500277787. [DOI] [PubMed] [Google Scholar]
- 53.Chou S.-Y., Hsu C.-S., Wang K.-T., Wang M.-C., Wang C.-C. Antitumor effects of osthol from Cnidium monnieri: an in vitro and in vivo study. Phytotherapy Research. 2007;21(3):226–230. doi: 10.1002/ptr.2044. [DOI] [PubMed] [Google Scholar]
- 54.Lin V. C.-H., Chou C.-H., Lin Y.-C., et al. Osthole suppresses fatty acid synthase expression in HER2-overexpressing breast cancer cells through modulating Akt/mTOR pathway. Journal of Agricultural and Food Chemistry. 2010;58(8):4786–4793. doi: 10.1021/jf100352c. [DOI] [PubMed] [Google Scholar]
- 55.Xu X., Zhang Y., Qu D., Jiang T., Li S. Osthole induces G2/M arrest and apoptosis in lung cancer A549 cells by modulating PI3K/Akt pathway. Journal of Experimental & Clinical Cancer Research. 2011;30(1, article 33) doi: 10.1186/1756-9966-30-33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Dien P. H., Nhan N. T., Le Thuy H. T., Quang D. N. Main constituents from the seeds of Vietnamese Cnidium monnieri and cytotoxic activity. Natural Product Research. 2012;26(22):2107–2111. doi: 10.1080/14786419.2011.619186. [DOI] [PubMed] [Google Scholar]
- 57.Yang D., Gu T., Wang T., Tang Q., Ma C. Effects of osthole on migration and invasion in breast cancer cells. Bioscience, Biotechnology and Biochemistry. 2010;74(7):1430–1434. doi: 10.1271/bbb.100110. [DOI] [PubMed] [Google Scholar]
- 58.Zhang L., Jiang G., Yao F., et al. Growth inhibition and apoptosis induced by osthole, a natural coumarin, in Hepatocellular Carcinoma. PLoS ONE. 2012;7(5) doi: 10.1371/journal.pone.0037865.e37865 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Kao S.-J., Su J.-L., Chen C.-K., et al. Osthole inhibits the invasive ability of human lung adenocarcinoma cells via suppression of NF-κB-mediated matrix metalloproteinase-9 expression. Toxicology and Applied Pharmacology. 2012;261(1):105–115. doi: 10.1016/j.taap.2012.03.020. [DOI] [PubMed] [Google Scholar]
- 60.Han X.-H., Ye Y.-Y., Guo B.-F., Liu S. Effects of platycodin D in combination with different active ingredients of Chinese herbs on proliferation and invasion of 4T1 and MDA-MB-231 breast cancer cell lines. Journal of Chinese Integrative Medicine. 2012;10(1):67–75. doi: 10.3736/jcim20120111. [DOI] [PubMed] [Google Scholar]
- 61.Guo B.-F., Liu S., Ye Y.-Y., Han X.-H. Inhibitory effects of osthole, psoralen and aconitine on invasive activities of breast cancer MDA-MB-231BO cell line and the mechanisms. Journal of Chinese Integrative Medicine. 2011;9(10):1110–1117. doi: 10.3736/jcim20111012. [DOI] [PubMed] [Google Scholar]
- 62.Sheng L., Wu C. Y., Chen X. F. Inhibitory acting mechanism of psoralen-osthole on bone metastasis of breast cancer—an expatiation viewing from OPG/RANKL/RANK system. Zhongguo Zhong Xi Yi Jie He Za Zhi. 2011;31(5):684–689. [PubMed] [Google Scholar]
- 63.Hung C.-M., Kuo D.-H., Chou C.-H., Su Y.-C., Ho C.-T., Way T.-D. Osthole suppresses hepatocyte growth factor (HGF)-induced epithelial-mesenchymal transition via repression of the c-Met/Akt/mTOR pathway in human breast cancer cells. Journal of Agricultural and Food Chemistry. 2011;59(17):9683–9690. doi: 10.1021/jf2021489. [DOI] [PubMed] [Google Scholar]
- 64.Okamoto T., Kobayashi T., Yoshida S. Chemical aspects of coumarin compounds for the prevention of hepatocellular carcinomas. Current Medicinal Chemistry: Anti-Cancer Agents. 2005;5(1):47–51. doi: 10.2174/1568011053352622. [DOI] [PubMed] [Google Scholar]
- 65.Huang W.-J., Chen C.-C., Chao S.-W., et al. Synthesis of n-hydroxycinnamides capped with a naturally occurring moiety as inhibitors of histone deacetylase. ChemMedChem. 2010;5(4):598–607. doi: 10.1002/cmdc.200900494. [DOI] [PubMed] [Google Scholar]
- 66.Huang W.-J., Chen C.-C., Chao S.-W., et al. Synthesis and evaluation of aliphatic-chain hydroxamates capped with osthole derivatives as histone deacetylase inhibitors. European Journal of Medicinal Chemistry. 2011;46(9):4042–4049. doi: 10.1016/j.ejmech.2011.06.002. [DOI] [PubMed] [Google Scholar]
- 67.Huang R.-L., Chen C.-C., Huang Y.-L., et al. Osthole increases glycosylation of hepatitis B surface antigen and suppresses the secretion of hepatitis B virus in vitro . Hepatology. 1996;24(3):508–515. doi: 10.1053/jhep.1996.v24.pm0008781315. [DOI] [PubMed] [Google Scholar]
- 68.Okamoto T., Yoshida S., Kobayashi T., Okabe S. Inhibition of concanavalin A-induced mice hepatitis by coumarin derivatives. Japanese Journal of Pharmacology. 2001;85(1):95–97. doi: 10.1254/jjp.85.95. [DOI] [PubMed] [Google Scholar]
- 69.Okamoto T., Kawasaki T., Hino O. Osthole prevents anti-Fas antibody-induced hepatitis in mice by affecting the caspase-3-mediated apoptotic pathway. Biochemical Pharmacology. 2003;65(4):677–681. doi: 10.1016/S0006-2952(02)01606-4. [DOI] [PubMed] [Google Scholar]
- 70.Okamoto T., Kobayashi T., Yoshida S. Synthetic derivatives of osthole for the prevention of hepatitis. Medicinal Chemistry. 2007;3(1):35–44. doi: 10.2174/157340607779317607. [DOI] [PubMed] [Google Scholar]
- 71.Song F., Xie M.-L., Zhu L.-J., Zhang K.-P., Xue J., Gu Z.-L. Experimental study of osthole on treatment of hyperlipidemic and alcoholic fatty liver in animals. World Journal of Gastroenterology. 2006;12(27):4359–4363. doi: 10.3748/wjg.v12.i27.4359. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Zhang Y., Xie M.-L., Zhu L.-J., Gu Z.-L. Therapeutic effect of osthole on hyperlipidemic fatty liver in rats. Acta Pharmacologica Sinica. 2007;28(3):398–403. doi: 10.1111/j.1745-7254.2007.00533.x. [DOI] [PubMed] [Google Scholar]
- 73.Zhang Y., Xie M., Xue J., Gu Z. Osthole improves fat milk-induced fatty liver in rats: modulation of hepatic PPAR-alpha/gamma-mediated lipogenic gene expression. Planta Medica. 2007;73(8):718–724. doi: 10.1055/s-2007-981552. [DOI] [PubMed] [Google Scholar]
- 74.Zhang Y., Xie M.-L., Xue J., Gu Z.-L. Osthole regulates enzyme protein expression of CYP7A1 and DGAT2 via activation of PPARalpha/gamma in fat milk-induced fatty liver rats. Journal of Asian Natural Products Research. 2008;10(7-8):807–812. doi: 10.1080/10286020802102303. [DOI] [PubMed] [Google Scholar]
- 75.Sun F., Xie M.-L., Xue J., Wang H.-B. Osthol regulates hepatic PPARα-mediated lipogenic gene expression in alcoholic fatty liver murine. Phytomedicine. 2010;17(8-9):669–673. doi: 10.1016/j.phymed.2009.10.021. [DOI] [PubMed] [Google Scholar]
- 76.Qi Z., Xue J., Zhang Y., Wang H., Xie M. Osthole ameliorates insulin resistance by increment of adiponectin release in high-fat and high-sucrose-induced fatty liver rats. Planta Medica. 2011;77(3):231–235. doi: 10.1055/s-0030-1250268. [DOI] [PubMed] [Google Scholar]
- 77.Du R., Xue J., Wang H.-B., Zhang Y., Xie M.-L. Osthol ameliorates fat milk-induced fatty liver in mice by regulation of hepatic sterol regulatory element-binding protein-1c/2-mediated target gene expression. European Journal of Pharmacology. 2011;666(1–3):183–188. doi: 10.1016/j.ejphar.2011.05.014. [DOI] [PubMed] [Google Scholar]
- 78.Zhang J., Xue J., Wang H., Zhang Y., Xie M. Osthole improves alcohol-induced fatty liver in mice by reduction of hepatic oxidative stress. Phytotherapy Research. 2011;25(5):638–643. doi: 10.1002/ptr.3315. [DOI] [PubMed] [Google Scholar]
- 79.Sun F., Xie M. L., Zhu L. J., Xue J., Gu Z. L. Inhibitory effect of osthole on alcohol-induced fatty liver in mice. Digestive and Liver Disease. 2009;41(2):127–133. doi: 10.1016/j.dld.2008.01.011. [DOI] [PubMed] [Google Scholar]
- 80.Ogawa H., Sasai N., Kamisako T., Baba K. Effects of osthol on blood pressure and lipid metabolism in stroke-prone spontaneously hypertensive rats. Journal of Ethnopharmacology. 2007;112(1):26–31. doi: 10.1016/j.jep.2007.01.028. [DOI] [PubMed] [Google Scholar]
- 81.Shin E., Lee C., Sung S. H., Kim Y. C., Hwang B. Y., Lee M. K. Antifibrotic activity of coumarins from Cnidium monnieri fruits in HSC-T6 hepatic stellate cells. Journal of Natural Medicines. 2011;65(2):370–374. doi: 10.1007/s11418-010-0485-7. [DOI] [PubMed] [Google Scholar]
- 82.Pantel J., Williams S. Y., Mi D., et al. Development of a high throughput screen for allosteric modulators of melanocortin-4 receptor signaling using a real time cAMP assay. European Journal of Pharmacology. 2011;660(1):139–147. doi: 10.1016/j.ejphar.2011.01.031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Liang H.-J., Suk F.-M., Wang C.-K., et al. Osthole, a potential antidiabetic agent, alleviates hyperglycemia in db/db mice. Chemico-Biological Interactions. 2009;181(3):309–315. doi: 10.1016/j.cbi.2009.08.003. [DOI] [PubMed] [Google Scholar]
- 84.Lee W.-H., Lin R.-J., Lin S.-Y., Chen Y.-C., Lin H.-M., Liang Y.-C. Osthole enhances glucose uptake through activation of AMP-activated protein kinase in skeletal muscle cells. Journal of Agricultural and Food Chemistry. 2011;59(24):12874–12881. doi: 10.1021/jf2036559. [DOI] [PubMed] [Google Scholar]
- 85.Yuan J., Xie J., Li A., Zhou F. Effects of osthol on androgen level and nitric oxide synthase activity in castrate rats. Zhong Yao Cai. 2004;27(7):504–506. [PubMed] [Google Scholar]
- 86.Qin L. P., Wang H. B., Zhang J. Q. Effects of osthol and total-coumarins from Cnidium monnieri on immunological function in kidney yang deficiency mice. Zhongguo Zhong Xi Yi Jie He Za Zhi. 1995;15(9):547–549. [PubMed] [Google Scholar]
- 87.Qin L. P., Zhang H. M., Zhang W. D. Effect of osthol and total coumarins of fructus cnidii on thyroid hormone and thyrotropic hormone in kidney-yang deficiency rats. Zhongguo Zhong Xi Yi Jie He Za Zhi. 1996;16(9):552–553. [PubMed] [Google Scholar]
- 88.Qin L. P., Zhang J. Q., Shi H. P. Effects of coumarins from cnidium monnieri on the function of pituitary-adrenocortical axis in kidney yang deficiency rats. Zhongguo Zhong Xi Yi Jie He Za Zhi. 1997;17(4):227–229. [PubMed] [Google Scholar]
- 89.Guh J.-H., Yu S.-M., Ko F.-N., Wu T.-S., Teng C.-M. Antiproliferative effect in rat vascular smooth muscle cells by osthole, isolated from Angelica pubescens . European Journal of Pharmacology. 1996;298(2):191–197. doi: 10.1016/0014-2999(95)00812-8. [DOI] [PubMed] [Google Scholar]
- 90.Li L., Zhuang F.-E., Yang L., Zhang C.-L., Zhao G.-S., Zhao D.-K. Effects of osthole on isolated guinea pig heart atria. Zhongguo Yao Li Xue Bao. 1995;16(3):251–254. [PubMed] [Google Scholar]
- 91.Fusi F., Sgaragli G., Ha L. M., Cuong N. M., Saponara S. Mechanism of osthole inhibition of vascular Cav1.2 current. European Journal of Pharmacology. 2012;680(1–3):22–27. doi: 10.1016/j.ejphar.2012.01.038. [DOI] [PubMed] [Google Scholar]
- 92.Ko F.-N., Wu T.-S., Liou M.-J., Huang T.-F., Teng C.-M. Vasorelaxation of rat thoracic aorta caused by osthole isolated from Angelica pubescens . European Journal of Pharmacology. 1992;219(1):29–34. doi: 10.1016/0014-2999(92)90576-p. [DOI] [PubMed] [Google Scholar]
- 93.Chen R., Xue J., Xie M.-L. Reduction of isoprenaline-induced myocardial TGF-β1 expression and fibrosis in osthole-treated mice. Toxicology and Applied Pharmacology. 2011;256(2):168–173. doi: 10.1016/j.taap.2011.08.005. [DOI] [PubMed] [Google Scholar]
- 94.Ko F.-N., Wu T.-S., Liou M.-J., Huang T.-F., Teng C.-M. Inhibition of platelet thromboxane formation and phosphoinositide breakdown by osthole from Angelica pubescens. Thrombosis and Haemostasis. 1989;62(3):996–999. [PubMed] [Google Scholar]
- 95.Teng C.-M., Ko F.-N., Wang J.-P., et al. Antihaemostatic and antithrombotic effect of some antiplatelet agents isolated from Chinese herbs. The Journal of Pharmacy and Pharmacology. 1991;43(9):667–669. doi: 10.1111/j.2042-7158.1991.tb03561.x. [DOI] [PubMed] [Google Scholar]
- 96.Li L., Zhuang F. E., Zhao G. S., Zhao D. K. Effects of osthole on the isolated guinea-pig ileum and taeniae coli. Acta Pharmaceutica Sinica. 1993;28(12):899–904. [PubMed] [Google Scholar]
- 97.Teng C.-M., Lin C.-H., Ko F.-N., Wu T.-S., Huang T.-F. The relaxant action of osthole isolated from Angelica pubescens in guinea-pig trachea. Naunyn-Schmiedeberg's Archives of Pharmacology. 1994;349(2):202–208. doi: 10.1007/bf00169838. [DOI] [PubMed] [Google Scholar]
- 98.Chen J., Chiou W.-F., Chen C.-C., Chen C.-F. Effect of the plant-extract osthole on the relaxation of rabbit corpus cavernosum tissue in vitro. The Journal of Urology. 2000;163(6):1975–1980. doi: 10.1016/s0022-5347(05)67613-6. [DOI] [PubMed] [Google Scholar]
- 99.Chiou W.-F., Huang Y.-L., Chen C.-F., Chen C.-C. Vasorelaxing effect of coumarins from Cnidium monnieri on rabbit corpus cavernosum. Planta Medica. 2001;67(3):282–284. doi: 10.1055/s-2001-12013. [DOI] [PubMed] [Google Scholar]
- 100.Shi Z., Shen S., Zhou W., Wang F., Fan Y. Fusarium graminearum growth inhibition due to glucose starvation caused by osthol. International Journal of Molecular Sciences. 2008;9(3):371–382. doi: 10.3390/ijms9030371. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Wang C.-M., Zhou W., Li C.-X., Chen H., Shi Z.-Q., Fan Y.-J. Efficacy of osthol, a potent coumarin compound, in controlling powdery mildew caused by Sphaerotheca fuliginea . Journal of Asian Natural Products Research. 2009;11(9):783–791. doi: 10.1080/10286020903158964. [DOI] [PubMed] [Google Scholar]
- 102.Wang C.-M., Guan W., Fang S., et al. Antifungal activity of the osthol derivative JS-B against Phytophthora capsici . Journal of Asian Natural Products Research. 2010;12(8):672–679. doi: 10.1080/10286020.2010.495331. [DOI] [PubMed] [Google Scholar]
- 103.Figueroa M., Rivero-Cruz I., Rivero-Cruz B., Bye R., Navarrete A., Mata R. Constituents, biological activities and quality control parameters of the crude extract and essential oil from Arracacia tolucensis var. multifida . Journal of Ethnopharmacology. 2007;113(1):125–131. doi: 10.1016/j.jep.2007.05.015. [DOI] [PubMed] [Google Scholar]
- 104.Rosselli S., Maggio A., Bellone G., et al. Antibacterial and anticoagulant activities of coumarins isolated from the flowers of Magydaris tomentosa . Planta Medica. 2007;73(2):116–120. doi: 10.1055/s-2006-951772. [DOI] [PubMed] [Google Scholar]
- 105.Tamura S., Fujitani T., Kaneko M., Murakami N. Prenylcoumarin with Rev-export inhibitory activity from Cnidii Monnieris Fructus. Bioorganic & Medicinal Chemistry Letters. 2010;20(12):3717–3720. doi: 10.1016/j.bmcl.2010.04.081. [DOI] [PubMed] [Google Scholar]
- 106.Wang G., Zhou Z., Cheng C., Yao J., Yang Z. Osthol and isopimpinellin from Fructus cnidii for the control of Dactylogyrus intermedius in Carassius auratus . Veterinary Parasitology. 2008;158(1-2):144–151. doi: 10.1016/j.vetpar.2008.07.034. [DOI] [PubMed] [Google Scholar]
- 107.Wang K.-Y., Yao L., Du Y.-H., Xie J.-B., Huang J.-L., Yin Z.-Q. Anthelmintic activity of the crude extracts, fractions, and osthole from Radix angelicae pubescentis against Dactylogyrus intermedius in goldfish (Carassius auratus) in vivo. Parasitology Research. 2011;108(1):195–200. doi: 10.1007/s00436-010-2058-9. [DOI] [PubMed] [Google Scholar]
- 108.Tsai T. H., Tsai T. R., Chen C. C., Chen C. F. Pharmacokinetics of osthole in rat plasma using high-performance liquid chromatography. Journal of Pharmaceutical and Biomedical Analysis. 1996;14(6):749–753. doi: 10.1016/0731-7085(95)01692-9. [DOI] [PubMed] [Google Scholar]
- 109.Li Y., Meng F., Xiong Z., Liu H., Li F. HPLC determination and pharmacokinetics of osthole in rat plasma after oral administration of fructus cnidii extract. Journal of Chromatographic Science. 2005;43(8):426–429. doi: 10.1093/chromsci/43.8.426. [DOI] [PubMed] [Google Scholar]
- 110.Zhou J., Wang S. W., Sun X. L. Determination of osthole in rat plasma by high-performance liquid chromatograph using cloud-point extraction. Analytica Chimica Acta. 2008;608(2):158–164. doi: 10.1016/j.aca.2007.12.029. [DOI] [PubMed] [Google Scholar]
- 111.Zhou J., Zeng P., Cheng Z. H., Liu J., Wang F. Q., Qian R. J. Application of hollow fiber liquid phase microextraction coupled with high-performance liquid chromatography for the study of the osthole pharmacokinetics in cerebral ischemia hypoperfusion rat plasma. Journal of Chromatography B: Analytical Technologies in the Biomedical and Life Sciences. 2011;879(23):2304–2310. doi: 10.1016/j.jchromb.2011.06.022. [DOI] [PubMed] [Google Scholar]
- 112.Wu Y.-N., Luan L.-B. In situ rats single pass perfusion intestinal absorption of the effective components in Radix Angelicae Pubescentis. Yao Xue Xue Bao. 2008;43(1):102–107. [PubMed] [Google Scholar]
- 113.Lv X., Wang C.-Y., Hou J., et al. Isolation and identification of metabolites of osthole in rats. Xenobiotica. 2012;42(11):1120–1127. doi: 10.3109/00498254.2012.689887. [DOI] [PubMed] [Google Scholar]
- 114.Yang X.-W., Guo Q.-M., Wang Y. Absorption and transport of 6 coumarins isolated from the roots of Angelica pubescens f. biserrata in human Caco-2 cell monolayer model. Zhong Xi Yi Jie He Xue Bao. 2008;6(4):392–398. doi: 10.3736/jcim20080413. [DOI] [PubMed] [Google Scholar]
- 115.Yuan Z., Xu H., Wang K., Zhao Z., Hu M. Determination of osthol and its metabolites in a phase I reaction system and the Caco-2 cell model by HPLC-UV and LC-MS/MS. Journal of Pharmaceutical and Biomedical Analysis. 2009;49(5):1226–1232. doi: 10.1016/j.jpba.2008.12.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Xu L.-N., Na W.-L., Liu X., et al. Identification of natural coumarin compounds that rescue defective ΔF508-CFTR chloride channel gating. Clinical and Experimental Pharmacology and Physiology. 2008;35(8):878–883. doi: 10.1111/j.1440-1681.2008.04943.x. [DOI] [PubMed] [Google Scholar]
- 117.Yang H., Xu L. N., Sui Y. J., et al. Stimulation of airway and intestinal mucosal secretion by natural coumarin CFTR activators. Frontiers in Pharmacology. 2011;2, article 52 doi: 10.3389/fphar.2011.00052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Barradas A. M. C., Fernandes H. A. M., Groen N., et al. A calcium-induced signaling cascade leading to osteogenic differentiation of human bone marrow-derived mesenchymal stromal cells. Biomaterials. 2012;33(11):3205–3215. doi: 10.1016/j.biomaterials.2012.01.020. [DOI] [PubMed] [Google Scholar]
- 119.Chik C. L., Liu Q.-Y., Li B., Karpinski E., Ho A. K. cGMP Inhibits L-type Ca2+ channel currents through protein phosphorylation in rat pinealocytes. Journal of Neuroscience. 1995;15(4):3104–3109. doi: 10.1523/JNEUROSCI.15-04-03104.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Kamp T. J., Hell J. W. Regulation of cardiac L-type calcium channels by protein kinase A and protein kinase C. Circulation Research. 2000;87(12):1095–1102. doi: 10.1161/01.RES.87.12.1095. [DOI] [PubMed] [Google Scholar]
- 121.Ay B., Iyanoye A., Sieck G. C., Prakash Y. S., Pabelick C. M. Cyclic nucleotide regulation of store-operated Ca2+ influx in airway smooth muscle. The American Journal of Physiology: Lung Cellular and Molecular Physiology. 2006;290(2):L278–L283. doi: 10.1152/ajplung.00188.2005. [DOI] [PubMed] [Google Scholar]
- 122.Harizi H., Juzan M., Moreau J.-F., Gualde N. Prostaglandins inhibit 5-lipoxygenase-activating protein expression and leukotriene B4 production from dendritic cells via an IL-10-dependent mechanism. Journal of Immunology. 2003;170(1):139–146. doi: 10.4049/jimmunol.170.1.139. [DOI] [PubMed] [Google Scholar]
- 123.Flamand N., Surette M. E., Picard S., Bourgoin S., Borgeat P. Cyclic AMP-mediated inhibition of 5-lipoxygenase translocation and leukotriene biosynthesis in human neutrophils. Molecular Pharmacology. 2002;62(2):250–256. doi: 10.1124/mol.62.2.250. [DOI] [PubMed] [Google Scholar]
- 124.Kambayashi T., Wallin R. P. A., Ljunggren H.-G. cAMP-elevating agents suppress dendritic cell function. Journal of Leukocyte Biology. 2001;70(6):903–910. [PubMed] [Google Scholar]