

Abstract
Cryptosporidium, a protozoan parasite that can cause severe diarrhea in a wide range of vertebrates including humans, is increasingly recognized as a parasite of a diverse range of wildlife species. However, little data are available regarding the identification of Cryptosporidium species and genotypes in wild aquatic environments, and more particularly in edible freshwater fish. To evaluate the prevalence of Cryptosporidiumspp. in fish from Lake Geneva (Lac Léman) in France, 41 entire fish and 100 fillets (cuts of fish flesh) were collected from fishery suppliers around the lake. Nested PCR using degenerate primers followed by sequence analysis was used. Five fish species were identified as potential hosts of Cryptosporidium: Salvelinus alpinus, Esox lucius, Coregonus lavaretus, Perca fluviatilis, and Rutilus rutilus. The presence of Cryptosporidium spp. was found in 15 out of 41 fish (37%), distributed as follows: 13 (87%) C. parvum, 1 (7%) C. molnari, and 1 (7%) mixed infection (C. parvum and C. molnari). C. molnari was identified in the stomach, while C. parvum was found in the stomach and intestine. C. molnari was also detected in 1 out of 100 analyzed fillets. In order to identify Cryptosporidium subtypes, sequencing of the highly polymorphic 60-kDa glycoprotein (gp60) was performed. Among the C. parvum positive samples, three gp60 subtypes were identified: IIaA15G2R1, IIaA16G2R1, and IIaA17G2R1. Histological examination confirmed the presence of potential developmental stages of C. parvum within digestive epithelial cells. These observations suggest that C. parvum is infecting fish, rather than being passively carried. Since C. parvum is a zoonotic species, fish potentially contaminated by the same subtypes found in terrestrial mammals would be an additional source of infection for humans and animals, and may also contribute to the contamination of the environment with this parasite. Moreover, the risk of human transmission is strengthened by the observation of edible fillet contamination.
Introduction
Cryptosporidium, a protozoan parasite that can cause severe diarrhea in a wide range of vertebrates including humans, is increasingly recognized as a parasite of a diverse range of wildlife species, including mammals, birds, reptiles, amphibians, and fish [1]. Although the epidemiology of cryptosporidiosis has been widely reported worldwide for different groups of animals, little biological, epidemiological and molecular data are available on Cryptosporidium infection in fish, even though the parasite has been already described and genetically characterized in more than 20 species of both freshwater and marine fish. Cryptosporidium molnari, the only currently recognized species infecting fish, was first identified in sea bream (Sparus aurata) and European sea bass (Dicentrarchus labrax) [2]. Cryptosporidium scophthalmi was detected in turbot (Psetta maxima, syn. Scophthalmus maximus) [3], but this species is still considered a nomen nudum due to a lack of genetic data [4].
Cryptosporidium species found in other groups of vertebrates have also been identified in fish, including C. parvum, C. hominis, C. scrofarum and C. xiaoi. Additionally, eight Cryptosporidium fish genotypes, and one Cryptosporidium rat III-like genotype, have been described in fish [4]. Recently, the species name Cryptosporidium huwi has been proposed for the piscine genotype 1 from the guppy (Poecilia reticulata) to reflect its genetic and biological differences from gastric and intestinal Cryptosporidium species [5].
In fish hosts, Cryptosporidium fish species and genotypes are located either in the stomach or intestine, as attested by histological analyses. Moreover, it has been reported that the parasite can cause clinical manifestations, such as emaciation, decrease in growth rate, anorexia, whitish feces, abdominal swelling, and ascites [2,3]. An increase in the mortality rate associated with Cryptosporidium infection has also been reported, particularly in larval and juvenile infected fish [6]. A significant correlation was found between the presence of the parasite and both fish weight and seasonality, the rate of infection being higher in fish weighing less than 100 grams and in the spring [7]. In addition, a relationship was observed between the presence of the parasite and the production stage in farmed fish [7].
It is notable that many results relating to fish Cryptosporidium infection were reported in farmed or aquarium fish [2,7,8]. However, little data are currently available regarding the molecular identification of Cryptosporidium species and genotypes in wild fish populations and, in particular, in edible fish. Indeed, only two studies have been conducted in Australia and Papua New Guinea on wild marine and freshwater fish [9,10].
Therefore, the aim of our study was to evaluate the prevalence of Cryptosporidium species/ genotypes in freshwater edible fish hosts from Lake Geneva in France. Lake Geneva is located between Switzerland and France, and is the largest freshwater reservoir in Western Europe, with a surface area of 580 km2, a volume of 89 km3, and a maximum depth of 309 m (Fig 1). More than 1.5 million people (in France and Switzerland) live around this lake [11]. In addition, the local fish are quite often consumed as raw preparations by the local population at home or in restaurants located around the shores of the lake. Fish are also a source of income, as around 150 professional fishermen are registered as active on the lake.
Fig 1. Map of the study area and sampling site (Thônon-les-Bains).

Effluent from wastewater treatment plants discharged into the Lake Geneva catchment area (CIPEL: Commission Internationale pour la Protection des Eaux du Léman).
Materials and Methods
Fish sampling
A total of 41 adult fish were purchased directly on the shores of the lake from local fishermen of Thônon-les-Bains (geographic coordinates: 46° 22' 0" North, 6° 29' 0" East), or Sechex, a small village located eight kilometers West of Thônon-les-Bains, in November 2011 (fall) and April 2013 (spring) (Fig 1). The weight, size, sex, origin and sexual maturity of each individual were determined (Table 1). For each fish, scratchings of the gastric and intestinal epithelia were performed after dissection, and the cells were preserved in the fixative RCL2 and stored at -20°C. A section of the stomach and bowel were also fixed in 10% buffered formalin. One hundred additional fillets (only cuts of fish flesh without viscera) of European perch (Perca fluviatilis) were purchased from the fishermen of Thônon-les-Bains, or St Gingolph (27 km Eeast of Thônon-les-Bains near the Swiss border, geographical coordinates: 46° 23' 0" North, 6° 40' 0" East) to evaluate potential contamination with Cryptosporidium spp. at this location. Slices of 2–3 mm were sampled and stored at—20°C in RCL2. No approval from Institutional Animal Care and Use Committee or ethics committee was necessary as no experiments that involved alive fish were performed. All fish examined were bought dead from professional fishermen, fishmongers and supermarkets selling fresh fish for consumption. Therefore, no sacrificial method was required. No fish sampled in this work was captured in a protected area and consequently, our sampling protocol did not need any specific permission for the location. Finally, no specimen included in the present work involved endangered or protected species.
Table 1. Freshwater fish specimens collected in Lake Geneva.
| Fish code | Fish species | Common name | Size (cm) | Weight (g) | Sexual maturity | Sex | Presence of other parasites |
|---|---|---|---|---|---|---|---|
| 5301 | Salvelinus alpinus | Arctic char | 40 | 610 | ND | Male | Cestoda |
| 5302 | Salvelinus alpinus | Arctic char | 37 | 504 | ND | Male | Cestoda |
| 5303 | Salvelinus alpinus | Arctic char | 37 | 530 | ND | Male | Cestoda |
| 5304 | Salvelinus alpinus | Arctic char | 38 | 548 | ND | Male | Cestoda |
| 5305 | Salvelinus alpinus | Arctic char | 29 | 224 | ND | Male | Cestoda |
| 5306 | Salvelinus alpinus | Arctic char | 38 | 580 | ND | Male | Cestoda |
| 5307 | Esox lucius | Northern pike | 37 | 328 | ND | Female | Cestoda |
| 5308 | Esox lucius | Northern pike | 35 | 328 | No | Female | Cestoda |
| 5309 | Lota lota | Burbot | 29 | 134 | ND | Male | Cestoda |
| 5310 | Lota lota | Burbot | 24 | 96 | No | Female | Nematoda |
| 5311 | Coregonus lavaretus | European whitefish | 31 | 276 | No | Female | No |
| 5312 | Coregonus lavaretus | European whitefish | 33 | 264 | ND | Male | No |
| 5313 | Coregonus lavaretus | European whitefish | 29 | 232 | ND | Male | Cestoda |
| 5314 | Coregonus lavaretus | European whitefish | 33 | 266 | ND | Male | Cestoda |
| 5315 | Coregonus lavaretus | European whitefish | 31 | 220 | ND | Male | Cestoda |
| 5316 | Coregonus lavaretus | European whitefish | 22 | 84 | ND | ND | Cestoda |
| 5317 | Perca fluviatilis | European perch | 11 | 16 | ND | Male | Cestoda |
| 5318 | Perca fluviatilis | European perch | 11 | 18 | ND | Male | Cestoda |
| 5319 | Perca fluviatilis | European perch | 11 | 16 | ND | Male | Cestoda |
| 5320 | Perca fluviatilis | European perch | 11 | 16 | ND | ND | Cestoda |
| 5321 | Perca fluviatilis | European perch | 42 | 1500 | Yes | Female | Acantocephala |
| 5322 | Perca fluviatilis | European perch | 29 | 318 | No | Female | Trematoda digenea |
| 5323 | Perca fluviatilis | European perch | 26 | 220 | Yes | Female | Cestoda |
| 5324 | Perca fluviatilis | European perch | 21 | 124 | No | Female | Trematoda digenea |
| 5325 | Rutilus rutilus | Roach | 18 | 42 | No | Female | Cestoda |
| 5326 | Lota lota | Burbot | 31 | 222 | No | Female | No |
| 5327 | Esox lucius | Northern pike | 53 | 1800 | Yes | Female | Cestoda |
| 5328 | Lota lota | Burbot | 25 | 97 | ND | Male | Microsporidia |
| 5329 | Coregonus lavaretus | European whitefish | 40 | 54 | Yes | Female | Cestoda |
| 5330 | Coregonus lavaretus | European whitefish | 33 | 54 | ND | Male | Cestoda |
| 5331 | Coregonus lavaretus | European whitefish | 36 | 54 | ND | Male | Cestoda |
| 5332 | Coregonus lavaretus | European whitefish | 40 | 54 | Yes | Female | No |
| 5333 | Coregonus lavaretus | European whitefish | 39 | 54 | Yes | Female | No |
| 5334 | Esox lucius | Northern pike | 69 | 2600 | ND | Male | Monogenea |
| 5335 | Squalius cephalus | European chub | 52 | 2300 | Yes | Female | Monogenea |
| 5336 | Esox lucius | Northern pike | 60 | 2000 | Yes | Female | Monogenea |
| 5337 | Perca fluviatilis | European perch | 27 | 30 | Yes | Female | Acantocephala |
| 5338 | Abramis brama | Common bream | 50 | 2000 | ND | Male | Trematoda digenea |
| 5339 | Perca fluviatilis | European perch | 15 | 45 | Yes | Female | Trematoda digenea |
| 5340 | Perca fluviatilis | European perch | 16 | 47 | ND | Male | Cestoda |
| 5341 | Perca fluviatilis | European perch | 16 | 56 | Yes | Female | No |
Fish from 5301 to 5326 were purchased from the fishermen of Thônon-les-Bains; Fish from 5327–5341 were purchased from the fishermen of Sechex
ND: Not determined.
DNA extraction
Genomic DNA extraction was performed on 96-well plates, using the NucleoSpin Kit (Macherey-Nagel, GmbH & Co KG, Germany) according to the manufacturer’s protocol. The final DNA elution was 100 μl.
Primer design and nested PCR
An alignment of the 18S rRNA gene sequences obtained from Cryptosporidium isolates characterized in fish (GenBank accession numbers: FJ769050, HM243547, HM243548, HM243549, HM243550, JF285332, JF285333, AY524773, HM989832, HM989833, HM989834, HM991857, GQ925452) [7] was performed using the BioEdit v7.0.1 package (http://www.mbio.ncsu.edu:BioEdit/bioedit.html). After identification of a target DNA fragment for nested 18S PCR common to all sequences, two sets of generate primers were selected within the hypervariable region. These degenerate primers were modified from those proposed by Ryan et al [12]. The external primer pair JerExtF (5’-GACATATCWTTYAAGTTTCTGACC-3’) (base pair position 292) and JerExtR (5’-CTGAAGGAGTAAGGAACAACC-3’) (base pair position 1007) amplified a DNA fragment of 784 bp. The internal primer pair JerIntF (5’-CCTATCAGCTTTMGACGGTAGG-3’) (base pair position 289) and JerIntR (5’-TCTAAGAATTTCACCTCTGACTG-3’) (base pair position 851) resulted in the amplification of a DNA fragment of 588 bp. For the first round of amplification, the PCR mixture contained 10 μl of DNA, 1x HotStarTaq Plus buffer, 2 mM MgCl2, 0.4 μM for each primer, 200μM dNTP each and 1.5U HotStarTaq Plus DNA polymerase (Qiagen Inc., Valencia, California) in a final volume of 50 μl. The PCR conditions were as follows: a denaturation step at 94°C for 10 min, followed by 40 cycles of 94°C for 45 sec, annealing at 67°C for 45 sec, and extension at 72°C for 1 min. The post-extension was completed at 72°C for 5 min. The second PCR amplification was performed in a 50 μl reaction volume containing 2 μl of the primary PCR product, 1xHotStarTaq Plus buffer, 3 mM MgCl2, 0.4 μM for each primer, 200 μM dNTP each and 1.5 U HotStarTaq Plus DNA polymerase. The PCR conditions were identical to those in the first round. Nested 18S PCR reactions were conducted using a PTC 200 thermocycler (MJ Research, Waltham, USA). The PCR products were analyzed on a 2% agarose gel and visualized by ethidium bromide staining.
DNA sequencing and analysis
To identify Cryptosporidium species at the molecular level, positive nested 18S PCR products were purified and sequenced directly on both strands, using the forward and reverse primers from the second round, by the company Genoscreen (Institut Pasteur de Lille, France). The sequences obtained were aligned using the BioEdit v7.0.1 package, and then compared with the sequences of Cryptosporidium published on the NCBI server (http://www.ncbi.nlm.nih.gov/BLAST/) using the basic local alignment search tool (BLAST) program. Isolates genotyped as C. parvum were further subtyped using a second nested PCR that amplifies a fragment of the 60 kDa glycoprotein (gp60) gene, as described [13]. The amplified DNA fragments were purified, sequenced, and analyzed as described above.
Histological analysis
The stomach and intestine of the fish were removed, fixed in 10% buffered formalin, and paraffin-embedded specimens were sectioned to a thickness of 5 μm to be processed using standard staining techniques (Hematoxylin & Eosin). Inflammation in digestive sections was scored as follows: 0, no inflammation; +1, moderate inflammation, focally distributed; +2, moderate inflammation, widely distributed; +3, severe inflammation, widely distributed throughout the section. The sections were examined by a pathologist using a Leica DMRB microscope equipped with a Leica digital camera connected to an Imaging Research MCID analysis system (MCID Software, Cambridge, UK).
Nucleotide sequence accession numbers
The 18S rRNA nucleotide sequences obtained in this study were deposited in the GenBank database under the accession numbers KP939333-KP939354.
Results
The molecular analysis of digestive tissues identified the presence of Cryptosporidium spp. in 15 out of 41 fish, representing a frequency of 37%. The fish species Arctic char (Salvelinus alpinus) (4/6), Northern pike (Esox lucius) (2/5), European whitefish (Coregonus lavaretus) (4/11), European perch (Perca fluviatilis) (4/12), and roach (Rutilus rutilus) (1/1) were identified as potential new hosts for Cryptosporidium spp. (Table 2).
Table 2. Cryptosporidium species and subtypes in wild freshwater fish from Lake Geneva identified at the 18S rDNA and GP60 loci.
| Code | Fish species | Fish common name | Organ | Cryptosporidium species 18S | Percentage of identity with reference sequence * | SNP** position | SNP** | GP60 |
|---|---|---|---|---|---|---|---|---|
| 5302 | Salvelinus alpinus | Arctic char | Intestine | C. parvum | 99.8% | 347 | T/C | NA |
| 5303 | Salvelinus alpinus | Arctic char | Intestine | C. parvum | 99.6% | 347 | T/C | IIaA17G2R1 |
| 435 | C/T | |||||||
| 5304 | Salvelinus alpinus | Arctic char | Stomach | C. parvum | 99.8% | 390 | G/A | IIaA15G2R1 |
| Intestine | C. parvum | 99.8% | 145 | A/G | IIaA15G2R1 | |||
| 5305 | Salvelinus alpinus | Arctic char | Intestine | C. parvum | 99.6% | 300 | T/C | IIaA15G2R1 |
| 507 | A/G | |||||||
| 5307 | Esox lucius | Northern pike | Stomach | C. molnari | 98.3% | 314 | A/T | NA |
| 322 | T/A | |||||||
| 324 | T/C | |||||||
| 329 | C/T | |||||||
| 341 | A/G | |||||||
| 370 | A/T | |||||||
| 376 | A/T | |||||||
| 377 | C/T | |||||||
| 506 | G/A | |||||||
| Intestine | C. parvum | 99.4% | 244 | G/A | IIaA17G2R1 | |||
| 347 | T/C | |||||||
| 496 | T/C | |||||||
| 5308 | Esox lucius | Northern pike | Stomach | C. molnari | 98.3% | 314 | A/T | NA |
| 322 | T/A | |||||||
| 324 | T/C | |||||||
| 329 | C/T | |||||||
| 341 | A/G | |||||||
| 370 | A/T | |||||||
| 376 | A/T | |||||||
| 377 | C/T | |||||||
| 506 | G/A | |||||||
| 5311 | Coregonus lavaretus | European whitefish | Stomach | C. parvum | 100% | - | - | IIaA15G2R1 |
| Intestine | C. parvum | 99.8% | 437 | T/C | IIaA17G2R1 | |||
| 5312 | Coregonus lavaretus | European whitefish | Stomach | C. parvum | 99.6% | 324 | T/C | IIaA17G2R1 |
| 475 | T/C | |||||||
| Coregonus lavaretus | European whitefish | Intestine | C. parvum | 99.2% | 87 | A/G | - | |
| 151 | A/G | |||||||
| 390 | G/A | |||||||
| 491 | T/C | |||||||
| 5314 | Coregonus lavaretus | European whitefish | Stomach | C. parvum | 99.8% | 235 | A/G | - |
| 5316 | Coregonus lavaretus | European whitefish | Stomach | C. parvum | 99.8% | 390 | G/A | - |
| 5318 | Perca fluviatilis | European perch | Stomach | C. parvum | 99.6% | 300 | T/C | IIaA15G2R1 |
| 507 | A/G | |||||||
| 5320 | Perca fluviatilis | European perch | Stomach | C. parvum | 99.8% | 211 | C/T | - |
| 5322 | Perca fluviatilis | European perch | Stomach | C. parvum | 99.8% | 27 | G/A | IIaA16G2R1 |
| Intestine | C. parvum | 100% | - | - | IIaA16G2R1 | |||
| 5323 | Perca fluviatilis | European perch | Stomach | C. parvum | 100% | - | - | IIaA15G2R1 |
| 5325 | Rutilus rutilus | Roach | Stomach | C. parvum | 100% | - | - | IIaA17G2R1 |
| Intestine | C. parvum | 100% | - | - | - |
The sequence analysis of the 18S rDNA locus identified two species of Cryptosporidium, distributed as follows: 13 C. parvum (87%), 1 C. molnari (7%), and 1 mixed infection (C. molnari and C. parvum) (7%). In 9 of the 15 infected fish, the presence of Cryptosporidium spp. was found either in the stomach or intestine, while in the 6 remaining infected fish, Cryptosporidium spp. were present in both organs. The selective extraction of DNA from these organs, followed by nested 18S PCR and sequencing, confirmed the presence of C. molnari only in the stomach of fish, while C. parvum was found in both stomach and intestine. Among the stomach samples, two were positive for C. molnari, and 10 were positive for C. parvum. Among the intestinal samples, eight were positive for C. parvum only. The 18S rRNA gene sequences of 5 out of 19 isolates of C. parvum found either in the stomach or intestine were 100% identical to that of a previously described species of C. parvum (GenBank: KJ939305 [7]), while 14 isolates exhibited single nucleotide polymorphisms (SNPs). It is common to identify sequence differences and variations such as single nucleotide polymorphisms (SNPs) that can be associated to genetic diversity according to the degree of homology. SNPs were distributed as follows: only 1 SNP for 8 isolates, 2 SNPs for 4 isolates, 3 SNPs for 1 isolate, and 4 SNPs for one isolate (Table 2). All SNPs identified in the C. parvum isolates corresponded to transition mutations. The two isolates identified as C. molnari were identical but showed 9 SNPs in comparison to the C. molnari reference sequence (GenBank: HM243550[7] (Table 2). In particular, 5 SNPs were associated with transition mutations, and 4 SNPs (in positions 322, 330, 378, 384) were associated with transversion mutations between adenine and thymine (A/T). The SNPs could not be associated with a specific sampling site (gastric vs. intestinal site) or with a specific fish species.
In order to identify Cryptosporidium subtypes, sequencing of the highly polymorphic 60-kDa glycoprotein (gp60) was performed. Partial sequences of the gp60 gene were subsequently obtained for 13 isolates identified as C. parvum. Three different subtypes were identified as follows: IIaA15G2R1 (6/13), IIaA17G2R1 (5/13), and IIaA16G2R1 (2/13) (Table 2).
Following histological examination of sections either from the stomach or intestine, the presence of Cryptosporidium-like bodies within the cells of the digestive epithelium was confirmed in samples from 10 C. parvum-positive fish (Fig 2A, 2B and 2C; Table 3). An inflammatory reaction with leukocyte infiltration was observed occasionally. The presence of other intestinal parasites, identified as nematodes, was confirmed in histological sections of two fish (Fig 2D, Table 2). The histological analysis of the remaining fish was not possible due to autolysis of tissues.
Fig 2. Stained sections of the digestive tract of fish.
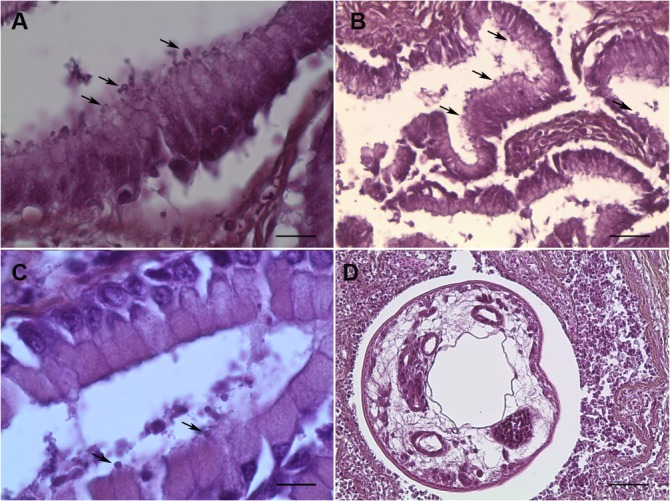
A. Presence of round bodies suggestive of the developmental stages of C. parvum was observed in the apical position (arrows) within the epithelial cells of gastric glands. Bar = 25 μm. B. Presence of round bodies suggestive of the developmental stages of C. parvum observed in the apical position (arrows) within the intestinal epithelial cells. Bar = 75 μm. C. Presence of round bodies suggestive of the developmental stages of C. molnari observed in the apical position (arrows) within the epithelial cells of gastric glands. Bar = 25 μm. D. Section of a nematode in the intestinal mucosa, surrounded by a severe inflammatory reaction. Bar = 200 μm. Hematoxylin & Eosin staining.
Table 3. Histological examination of digestive organs of different fish species from Lake Geneva infected by Cryptosporidium spp.
| Fish code | Fish species | Fish common name | Organ | Histological examination* | Cryptosporidium species (18S rDNA) |
|---|---|---|---|---|---|
| 5302 | Salvelinus alpinus | Arctic char | Intestine | Inflammation: +1 | C. parvum |
| Intracellular Cryptosporidium-like bodies | |||||
| 5303 | Salvelinus alpinus | Arctic char | Intestine | ND | C. parvum |
| 5304 | Salvelinus alpinus | Arctic char | Stomach | Inflammation: 0 Intracellular | C. parvum |
| Cryptosporidium-like bodies | |||||
| Intestine | ND | C. parvum | |||
| 5305 | Salvelinus alpinus | Arctic char | Intestine | Inflammation: +1 | C. parvum |
| Intracellular Cryptosporidium-like bodies | |||||
| 5307 | Esox lucius | Northern pike | Stomach | ND | C. molnari |
| Intestine | ND | C. parvum | |||
| 5308 | Esox lucius | Northern pike | Stomach | Zones of autolysis | C. molnari |
| Inflammation: 0 Intracellular Cryptosporidium-like bodies | |||||
| 5311 | Coregonus lavaretus | European whitefish | Stomach | ND | C. parvum |
| Intestine | Zones of autolysis | C. parvum | |||
| Inflammation: 0 Intracellular Cryptosporidium-like bodies | |||||
| 5312 | Coregonus lavaretus | European whitefish | Stomach | Zones of autolysis | C. parvum |
| Inflammation: 0 Intracellular Cryptosporidium-like bodies | |||||
| Intestine | Zones of autolysis | C. parvum | |||
| Inflammation: 0 Intracellular Cryptosporidium-like bodies | |||||
| 5314 | Coregonus lavaretus | European whitefish | Stomach | Inflammation: 0 Intracellular Cryptosporidium-like bodies | C. parvum |
| 5316 | Coregonus lavaretus | European whitefish | Stomach | Inflammation: 0 Intracellular Cryptosporidium-like bodies | C. parvum |
| 5318 | Perca fluviatilis | European perch | Stomach | Inflammation: +3 | C. parvum |
| Presence of a nematode | |||||
| 5320 | Perca fluviatilis | European perch | Stomach | Autolysis | C. parvum |
| 5322 | Perca fluviatilis | European perch | Stomach | ND | C. parvum |
| Intestine | Inflammation: +3 | C. parvum | |||
| Presence of a nematode | |||||
| 5323 | Perca fluviatilis | European perch | Stomach | Inflammation: 0 Intracellular Cryptosporidium-like bodies | C. parvum |
| 5325 | Rutilus rutilus | Roach | Stomach | Inflammation: 0 Intracellular Cryptosporidium-like bodies | C. parvum |
| Intestine | Inflammation: 0 Intracellular Cryptosporidium-like bodies | C. parvum |
ND: Not done
* Inflammation in digestive sections was scored as follows: 0, no inflammation; +1, moderate inflammation, focally distributed; +2, moderate inflammation, widely distributed; +3, severe inflammation, widely distributed throughout the section.
The potential contamination of fish flesh with Cryptosporidium spp. was evaluated. One hundred fish fillets of European perch (Perca fluviatilis) were analyzed by nested PCR and sequencing. The presence of C. molnari was detected in only one fillet. The 18S rRNA gene sequence of this C. molnari isolate was identical to that of the two isolates found in the stomach of two Northern pike (Table 2).
Discussion
This study reports the first epidemiological and molecular data on the presence of Cryptosporidium in fish in France. The overall frequency of Cryptosporidium spp. in fish sampled from Lake Geneva was high, reaching 37%. Previous studies have reported a high prevalence of Cryptosporidium spp. in fish, but mainly in juvenile marine fish. For instance, Sitjà-Bobadilla et al. reported 100% C. scophthalmi prevalence in juvenile turbot in Europe [7]. In contrast, a recent study in Australia found no Cryptosporidium isolates in freshwater fish [9], while a Cryptosporidium prevalence of 0.2% was found in wild freshwater species in Papua New Guinea [10]. Therefore, even if the comparative data is scarce, this is to our knowledge the first time that Cryptosporidium has been detected at a very high prevalence in freshwater fish.
Five new species of fish hosts for Cryptosporidium were identified: Arctic char (Salvelinus alpinus), Northern pike (Esox lucius), European whitefish (Coregonus lavaretus), European perch (Perca fluviatilis) and Roach (Rutilus rutilus). Although it is generally accepted that the prevalence of Cryptosporidium is higher in juvenile fish, all of the fish analyzed in our study were adults, according to size and weight, and according to sexual maturity when this parameter could be determined (Table 1).
Two species of Cryptosporidium were detected in fish hosts: C. molnari and C. parvum. C. molnari was identified in freshwater aquaculture fish [14], but this is the first time that this parasite species has been found in wild freshwater fish. The 18S rRNA gene sequences of the 3 C. molnari isolates identified in our study were 98% identical to those of the C. molnari reference sequences collected from the databases. Interestingly, these three sequences amplified from different individuals presented the same points of mutation, suggesting the circulation of the same parasite isolates in the lake environment.
A matter of importance to public health was the high rate of detection of C. parvum among fish hosts, as this species is the most common source of zoonotic infections [4]. Previous studies in Papua New Guinea and Australia also reported consistent detection of C. parvum in fish [9,10]. We speculate that the presence of C. parvum, and in particular the IIa subtype, in fish samples from Lake Geneva could be due to waterborne contamination with human and animal waste. In fact, the zoonotic C. parvum IIa subtype family has predominantly been found in calves and humans in North America, Europe, and Australia [15,16]. In addition, even if we did not search for the presence of Cryptosporidium in the lake water, it is well known that Cryptosporidium oocysts are found in groundwater, lakes, rivers, estuaries, and oceans, as a consequence of the great amount of feces from humans, pets, and domesticated or wild animals that is discharged, dumped, or carried in runoff into these waters [17]. In particular, in Lake Geneva, an increase in fecal bacteria of human and animal origin was described in sediment contaminated with wastewater treatment plant effluent, suggesting the presence of both human and animal sources of fecal pollution in the lake environment [11]. In parallel, it has been suggested that when fecal bacteria is present in water, Cryptosporidium could be present as well, and even though water quality monitoring and water treatment can reduce the presence of pathogens, they do not ensure absolute safety, due to the fact that Cryptosporidium oocysts are highly resistant [18].
Partial sequences of the gp60 gene subsequently amplified from C. parvum isolates allowed the identification of three different subtypes belonging to the IIa family, as follows: IIaA15G2R1, IIaA16G2R1, and IIaA17G2R1. The IIaA15G2R1 subtype has also been identified consistently in Papua New Guinea in mackerel scad, Decapterus maracellus [10], a wild marine fish. The subtypes IIaA15G2R1 and IIaA17G2R1 have been identified in cattle [19], the first of which is the most dominant zoonotic Cryptosporidium subtype infecting dairy cattle and humans in industrialized countries [4]. Indeed, the IIaA15G2R1 subtype represents up to 75% of the identified Cryptosporidium subpopulation in French calves [16]. On the other hand, the IIaA16G2R1 subtype has been identified in diarrheic calves [20–22] and also in wild boars (Sus scrofa) [23]. In rural areas, it is well known that animals can cohabit with livestock, often by sharing grazing and water sources. Other zoonotic Cryptosporidium species already identified in marine fish such as C. hominis C. xiaoi and C. scrofarum were not found in this study [4].
Since C. parvum is a zoonotic species, fish potentially contaminated by the same subtypes infecting terrestrial mammals would be an additional source of infection for humans and other animals, and may also contribute to the contamination of the environment with this parasite. However, it is not clear if fish are only carriers of C. parvum, or if C. parvum can develop its life cycle and multiply in this fish host.
In order to clarify this question, histological analysis of digestive tissues from C. parvum-positive fish was performed, and round bodies suggestive of C. parvum developmental stages were observed in an apical position within the cells, either in the stomach or intestine. These observations suggest that C. parvum is actually infecting fish, rather than being passively carried. Fluorescent-antibody staining assay using an anti-Cryptosporidium antibody (Crypto Cel immunofluorescence test, Cellabs, Brookvale, New South Wales, Australia) was tried to confirm the detection of Cryptosporidium oocysts from fish tissues but unfortunately, no signal was detected. This failure was probably due to the vulnerability of the oocyst antigens to formalin as was already described, particularly after more than one month of formalin fixation which was the case in our study [14]. Mild to moderate inflammation was occasionally found in gastrointestinal tissues, but we could not determine whether it was Cryptosporidium that was causing this reaction, since co-infection with other parasites was present. In some cases, the histological analysis of fish was not possible due to autolysis of tissues.
Furthermore, to evaluate a potential contamination of fish fillets with Cryptosporidium spp., 100 fish fillets of European perch (Perca fluviatilis) were analyzed by nested 18S PCR and sequencing, and the presence of C. molnari was detected in fillets from one individual. Fillet contamination with C. molnari could occur as a consequence of evisceration of the infected fish during the cleaning and preparation process. Although previous studies have shown no conclusive evidence of transmission of fish-hosted Cryptosporidium to mammals [24], the presence of the parasite also in fillets clearly highlights the risk of Cryptosporidium infection to humans, either when handling fish or consuming raw or undercooked fish carrying zoonotic species of Cryptosporidium. In our study, only C. molnari, apparently a non-pathogenic species for humans, was isolated from perch fillets. However, C. parvum isolated from the fish digestive tract could certainly be present in fish fillets. Further studies should be done to clarify this aspect.
One study in Maryland consistently reported that urban anglers are at a risk of contracting cryptosporidiosis from exposure received while fishing and consuming caught fish with a mean probability of infection of almost one [25]. Another study showed that blue crabs can transfer C. parvum oocysts to people who handle the crustaceans [26]. In addition, it has been reported that immunosuppressed patients are at risk of contracting cryptosporidiosis, either by contact with fish during preparation and handling, or by consumption of undercooked fish [27].
It was not unexpected to find Cryptosporidium in fish from Lake Geneva, as this parasite has already been found to be responsible for a human outbreak occurring in 2003 due to the contamination of the water supply network, in the nearby city of Divonne-les-Bains, affecting more than 700 individuals [28]. In addition, in Switzerland, a study reported the presence of C. parvum in samples collected from the drinking water distribution system in alpine rural regions, and it was suspected that the drinking water was contaminated by grazing cattle [29]. However, future studies should be conducted to detect the presence of the parasite in the lake environment.
In conclusion, these findings suggest that the transmission of Cryptosporidium could potentially occur in the interfaces between human, livestock, and fish populations. In fact, the wide host range of Cryptosporidium spp., together with the high output of oocyst shedding, allows a high level of contamination of the environment [23]. In particular, for fish hosts, the dispersion and transmission of zoonotic parasites would be facilitated by the aquatic habitat of the host that could potentially release fully sporulated oocysts contributing to the perpetuation of Cryptosporidium circulation. Finally, fish may be a good sentinel for the detection of water contamination caused by sewage or agricultural runoff.
Acknowledgments
We wish to thank Dr. Slapeta for kindly providing us with the DNA from Cryptosporidium molnari, used as a positive control in our PCR, the CIPEL (Commission Internationale pour la Protection des Eaux du Léman) for its assistance in the design of Fig 1 and the Fish-Parasites Network.
Data Availability
All relevant data are within the paper.
Funding Statement
This work was supported by the French National Research Agency (grant no. ANR-10-ALIA-004) and the regional competitiveness center, AQUIMER (Boulogne s/mer, France).
References
- 1. Ryan U, Power M. Cryptosporidium species in Australian wildlife and domestic animals. Parasitology. 2012;139: 1673–1688. 10.1017/S0031182012001151 [DOI] [PubMed] [Google Scholar]
- 2. Alvarez-Pellitero P, Sitjà-Bobadilla A. Cryptosporidium molnari n. sp. (Apicomplexa: Cryptosporidiidae) infecting two marine fish species, Sparus aurata L. and Dicentrarchus labrax L . Int J Parasitol. 2002;32: 1007–1021. [DOI] [PubMed] [Google Scholar]
- 3. Alvarez-Pellitero P, Quiroga MI, Sitjà-Bobadilla A, Redondo MJ, Palenzuela O, Padrós F, et al. Cryptosporidium scophthalmi n. sp. (Apicomplexa: Cryptosporidiidae) from cultured turbot Scophthalmus maximus. Light and electron microscope description and histopathological study. Dis Aquat Organ. 2004;62: 133–145. [DOI] [PubMed] [Google Scholar]
- 4. Ryan U, Fayer R, Xiao L. Cryptosporidium species in humans and animals: current understanding and research needs. Parasitology. 2014;141: 1667–1685. 10.1017/S0031182014001085 [DOI] [PubMed] [Google Scholar]
- 5. Ryan U, Paparini A, Tong K, Yang R, Gibson-Kueh S, O’Hara A, et al. Cryptosporidium huwi n. sp. (Apicomplexa: Eimeriidae) from the guppy (Poecilia reticulata). Exp Parasitol. 2015;150: 31–35. 10.1016/j.exppara.2015.01.009 [DOI] [PubMed] [Google Scholar]
- 6. Murphy BG, Bradway D, Walsh T, Sanders GE, Snekvik K. Gastric cryptosporidiosis in freshwater angelfish (Pterophyllum scalare). J Vet Diagn Invest. 2009;21: 722–727. [DOI] [PubMed] [Google Scholar]
- 7. Sitjà-Bobadilla A, Padrós F, Aguilera C, Alvarez-Pellitero P. Epidemiology of Cryptosporidium molnari in Spanish gilthead sea bream (Sparus aurata L.) and European sea bass (Dicentrarchus labrax L.) cultures: from hatchery to market size. Appl Environ Microbiol. 2005;71: 131–139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Morine M, Yang R, Ng J, Kueh S, Lymbery AJ, Ryan UM. Additional novel Cryptosporidium genotypes in ornamental fishes. Vet Parasitol. 2012;190: 578–582. 10.1016/j.vetpar.2012.06.036 [DOI] [PubMed] [Google Scholar]
- 9. Reid A, Lymbery A, Ng J, Tweedle S, Ryan U. Identification of novel and zoonotic Cryptosporidium species in marine fish. Vet Parasitol. 2010;168: 190–195. 10.1016/j.vetpar.2009.11.015 [DOI] [PubMed] [Google Scholar]
- 10. Koinari M, Karl S, Ng-Hublin J, Lymbery AJ, Ryan UM. Identification of novel and zoonotic Cryptosporidium species in fish from Papua New Guinea. Vet Parasitol. 2013;198: 1–9. 10.1016/j.vetpar.2013.08.031 [DOI] [PubMed] [Google Scholar]
- 11. Thevenon F, Regier N, Benagli C, Tonolla M, Adatte T, Wildi W, et al. Characterization of fecal indicator bacteria in sediments cores from the largest freshwater lake of Western Europe (Lake Geneva, Switzerland). Ecotoxicol Environ Saf. 2012;78: 50–56. 10.1016/j.ecoenv.2011.11.005 [DOI] [PubMed] [Google Scholar]
- 12. Ryan U, Xiao L, Read C, Zhou L, Lal AA, Pavlasek I. Identification of novel Cryptosporidium genotypes from the Czech Republic. Appl Environ Microbiol. 2003;69: 4302–4307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Gatei W, Das P, Dutta P, Sen A, Cama V, Lal AA, et al. Multilocus sequence typing and genetic structure of Cryptosporidium hominis from children in Kolkata, India. Infect Genet Evol. 2007;7: 197–205. [DOI] [PubMed] [Google Scholar]
- 14. Barugahare R, Dennis MM, Becker JA, Slapeta J. Detection of Cryptosporidium molnari oocysts from fish by fluorescent-antibody staining assays for Cryptosporidium spp. affecting humans. Appl Environ Microbiol. 2011;77: 1878–1880. 10.1128/AEM.02691-10 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Xiao L. Molecular epidemiology of cryptosporidiosis: An update. Exp Parasitol. 2010;124: 80–89. 10.1016/j.exppara.2009.03.018 [DOI] [PubMed] [Google Scholar]
- 16. Follet J, Guyot K, Leruste H, Follet-Dumoulin A, Hammouma-Ghelboun O, Certad G, et al. Cryptosporidium infection in a veal calf cohort in France: Molecular characterization of species in a longitudinal study. Vet Res. 2011;42. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Fayer R, Dubey JP, Lindsay DS. Zoonotic protozoa: from land to sea. Trends Parasitol. 2004;20: 531–536. [DOI] [PubMed] [Google Scholar]
- 18. Poté J, Goldscheider N, Haller L, Zopfi J, Khajehnouri F, Wildi W. Origin and spatial-temporal distribution of faecal bacteria in a bay of Lake Geneva, Switzerland. Environ Monit Assess. 2009;154: 337–348. 10.1007/s10661-008-0401-8 [DOI] [PubMed] [Google Scholar]
- 19. Ng J, Eastwood K, Durrheim D, Massey P, Walker B, Armson A, et al. Evidence supporting zoonotic transmission of Cryptosporidium in rural New South Wales. Exp Parasitol. 2008;119: 192–195. 10.1016/j.exppara.2008.01.010 [DOI] [PubMed] [Google Scholar]
- 20. Geurden T, Berkvens D, Martens C, Casaert S, Vercruysse J, Claerebout E. Molecular epidemiology with subtype analysis of Cryptosporidium in calves in Belgium. Parasitology. 2007;134: 1981–1987. [DOI] [PubMed] [Google Scholar]
- 21. Quilez J, Torres E, Chalmers RM, Robinson G, Del Cacho E, Sanchez-Acedo C. Cryptosporidium species and subtype analysis from dairy calves in Spain. Parasitology. 2008;135: 1613–1620. 10.1017/S0031182008005088 [DOI] [PubMed] [Google Scholar]
- 22. Díaz P, Quílez J, Chalmers RM, Panadero R, López C, Sánchez-Acedo C, et al. Genotype and subtype analysis of Cryptosporidium isolates from calves and lambs in Galicia (NW Spain). Parasitology. 2010;137: 1187–1193. 10.1017/S0031182010000181 [DOI] [PubMed] [Google Scholar]
- 23. García-Presedo I, Pedraza-Díaz S, González-Warleta M, Mezo M, Gómez-Bautista M, Ortega-Mora LM, et al. Presence of Cryptosporidium scrofarum, C. suis and C. parvum subtypes IIaA16G2R1 and IIaA13G1R1 in Eurasian wild boars (Sus scrofa). Vet Parasitol. 2013;196: 497–502. 10.1016/j.vetpar.2013.04.017 [DOI] [PubMed] [Google Scholar]
- 24. Certad G, Creusy C, Guyot K, Mouray A, Chassat T, Delaire B, et al. Fulminant cryptosporidiosis associated with digestive adenocarcinoma in SCID mice infected with Cryptosporidium parvum TUM1 strain. Int J Parasitol. 2010;40: 1469–1475. 10.1016/j.ijpara.2010.07.007 [DOI] [PubMed] [Google Scholar]
- 25. Roberts JD, Silbergeld EK, Graczyk T. A probabilistic risk assessment of Cryptosporidium exposure among Baltimore urban anglers. J Toxicol Environ Health A. 2007;70: 1568–1576. [DOI] [PubMed] [Google Scholar]
- 26. Graczyk TK, McOliver C, Silbergeld EK, Tamang L, Roberts JD. Risk of handling as a route of exposure to infectious waterborne Cryptosporidium parvum oocysts via Atlantic blue crabs (Callinectes sapidus). Appl Environ Microbiol. 2007;73: 4069–4070. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. McOliver CC, Lemerman HB, Silbergeld EK, Moore RD, Graczyk TK. Risks of recreational exposure to waterborne pathogens among persons with HIV/AIDS in Baltimore, Maryland. Am J Public Health. 2009;99: 1116–1122. 10.2105/AJPH.2008.151654 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Gofti-Laroche L et Schmitt M. Outbreak of gastroenteritis related to the pollution of water distribution system in the commune of Divonne-lesBains, Ain. DRASS Rhône Alpes, CIRE Rhône Alpes–Auvergne, Inst Veill Veill Sanit. 2003.
- 29. Füchslin HP, Kotzsch S, Egli T. Cryptosporidium spp. in drinking water: Samples from rural sites in Switzerland. Swiss Med Wkly. 2012;142 10.4414/smw.2012.13683 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
All relevant data are within the paper.