

Abstract
The cranial trigeminal and epibranchial ganglia are components of the peripheral nervous system that possess an important somatosensory role. These ganglia arise from the intermixing and coalescence of two different migratory cell types, neural crest cells and neurogenic placodes cells, and thus typify the phenomena of cell migration and intercellular interactions for their creation. The underlying molecular mechanisms of ganglia formation, however, are still poorly understood. To address this, we have analyzed the spatio-temporal expression profile of Annexin A6 during chick gangliogenesis, as Annexin proteins play important, conserved roles in ganglia development and physiology. We observe Annexin A6 protein in cranial neural crest cells prior to, during and after their emergence from the neural tube. Fully migratory cranial neural crest cells, however, are devoid of Annexin A6. Interestingly, we note Annexin A6 protein in trigeminal and epibranchial placode cells as these cells ingress from the ectoderm to initiate ganglia formation. This expression is also maintained in the sensory placodes later on when they coalesce with neural crest cells to assemble the cranial ganglia. These results suggest that the dynamic expression of Annexin A6 in various embryonic cell types may allow Annexin A6 to serve distinct functions throughout embryonic development.
Keywords: cranial ganglia, neural crest cells, placode cells, Annexin A6
1. Introduction
The migration of, and interaction among, multiple cell types underscore the creation of organismal tissues and organs throughout evolution. The formation of the cranial sensory ganglia, which consist of the trigeminal (V) and epibranchial (geniculate (facial VII), petrosal (glossopharyngeal IX), nodose (vagal X)) ganglia of the peripheral nervous system, exemplifies these different cellular behaviors. The trigeminal ganglia are responsible for relaying somatosensory information from the external environment to the central nervous system [1], while the epibranchial ganglia provide gustatory and viscerosensory information from the oropharyngeal cavity to the hindbrain [2]. These sensory ganglia are formed from two migratory embryonic cell populations, placode cells and neural crest cells. Derived from the head ectoderm, placode cells are focal thickenings that arise in a specific spatio-temporal fashion [3–5]. These cells later migrate away from the epithelium as mesenchymal cells and form different sense organs and ganglia, each with a distinct set of cell types, including sensory receptor cells, neurons, endocrine cells, and supporting cells [6]. Trigeminal placode cells develop closer to the central nervous system and are composed of the ophthalmic (opV) and maxillomandibular (mmV) placodes [1], while epibranchial placode cells (geniculate, petrosal and nodose placodes) form at the dorsoanterior margin of the branchial clefts [2]. Conversely, neural crest cells originate in the dorsal region of the embryonic neural tube as multipotent cells that then migrate extensively and give rise to diverse derivatives, including components of the craniofacial cartilage and skeleton, melanocytes, neurons and glia of the peripheral nervous system, and portions of the heart [7]. Neural crest cells in the midbrain and hindbrain, together with the cranial neurogenic placodes, create the cranial sensory ganglia. Because complex molecular pathways underlie the dual embryonic origin of the cranial sensory ganglia [1–6], it is important to understand the signals and effectors that drive cranial gangliogenesis. Studying the coordinated movement and interaction of neural crest and placode cells is critical, as defects during neural crest and placode cell development can lead to cranial dysmorphologies such as cleft lip and/or palate, sensory defects underlying multiple sclerosis, facial paralysis during trauma, and trigeminal neuralgia [8–10]. These conditions are observed in one in one thousand live births for cleft lip and/or palate [8], and one in ten thousand people for trigeminal neuralgia [9].
Members of the Annexin protein superfamily, which interact with both the phospholipid bilayer and form membrane-cytoskeletal protein complexes [11], have played a role in the development and physiology of the peripheral nervous system throughout evolution. Studies have revealed the presence of Annexin-like proteins in invertebrate gastropod snails, where Annexin 1- and Annexin 5-like proteins localize to neurons of various ganglia within the central nervous system [12]. Neurons in the dorsal root ganglia and spinal cord of adult rats differentially express Annexins A1–6 [13] (the “A” is added to signify the vertebrate nomenclature). Specifically, Annexin A6 is observed in dorsal root ganglion neurons of the rat [13]. In addition, loss of Annexin A2, which is expressed in mouse primary sensory neurons, causes neuropathic pain [14], while Annexin 5 possesses a role in the pathogenesis of autoimmune sensory neuropathy in humans [15]. However, the expression of Annexins during the assembly of the cranial ganglia of the peripheral nervous system has not been documented. The Annexin N and C terminal domains interact with cytosolic proteins and the inner plasma membrane leaflet, respectively [16], in both a calcium-dependent and –independent manner. Furthermore, the N terminal region provides regulation over Annexin function, while the C terminus (also called the core domain) possesses a calcium-binding domain [16]. In particular, Annexin A6 is the only dimer of the family consisting of two cores that can orient themselves differently depending on the required function and tissue type [17].
Annexins possess a wide variety of roles in different cell types that collectively make them intriguing molecules to investigate during processes such as gangliogenesis that involve cell motility and cell-cell interactions. For example, many Annexins have been implicated in tumor progression and metastasis, either as tumor suppressors, oncogenes, and regulators of cell signaling, division and apoptosis [18]. Moreover, Annexins A2 and A7 are involved in vesicular trafficking and cell growth, respectively, which are basic cellular processes that are also important for the formation of new tissues from multiple cell types [19]. In addition, Annexin proteins can organize cellular membranes through actin remodeling, function during endocytosis, and may be externalized by cells through exoctyosis, thereby allowing them to serve as extracellular signaling molecules (reviewed in [16]). Finally, Annexins play a role in the attachment (but not fusion) of cell membranes, indicating that these proteins could be critical to bring different cell types into close proximity [16].
In keeping with these varied roles for Annexin proteins, Annexin A6 possesses different functions depending upon the cellular context (see Table 1). In chick, Annexin A6 impacts chick cranial neural crest cell migration through effects on the premigratory neural crest cell cadherins Cadherin-6B and N-cadherin during the neural crest cell epithelial-to mesenchymal transition (EMT) [20]. These results are consistent with a known function for Annexin A6 in vitro in cytoskeletal remodeling and cell migration via interactions with the actin cytoskeleton [21, 22] and the actin cross-linking protein α-actinin [23]. Given the documented roles for Annexin A6 in early embryonic events such as neural crest cell EMT, and later processes including ganglia formation and physiology, we aimed to establish a detailed spatio-temporal expression profile for Annexin A6 in the developing chick embryo from neural crest cell EMT through the formation of the cranial trigeminal and epibranchial ganglia. Our results show the presence of Annexin A6 protein in premigratory neural crest cells, and those undergoing EMT, in keeping with our prior work examining Annexin A6 transcripts [20], but strikingly reveal a loss of Annexin A6 protein from fully migratory neural crest cells. Intriguingly, trigeminal and epibranchial placodal precursors begin expressing Annexin A6 at the onset of their ingression from the ectoderm into the adjacent mesenchyme, with expression maintained throughout all stages of trigeminal and epibranchial ganglia formation. Collectively, our results highlight the dynamic spatio-temporal expression of Annexin A6 during chick cranial gangliogenesis.
Table 1.
Diverse cellular functions for Annexin A6.
| Annexin A6 function | Cell typet |
|---|---|
| Modulation of EMT and migration20 | Chick cranial neural crest cells |
| Reduction in store-operated Ca2+ entry24 | HEK293 cells |
| Interaction with α-actinin 23, 25 | Rat cardiomyocytes |
| Cholesterol homeostasis and export of caveolin from the Golgi26 | CHO and A431 cells |
| Regulation of mitochondrial morphogenesis27 | Fibroblasts from Annexin A6 mutant mice, A431 cells |
| Cell migration; inhibition of motility in breast cancer carcinoma through focal adhesions26, 28 | BT-549 breast cancer cells |
| Association with endosomes and shuttling proteins to the lysosome for degradation16 | SV40-transformed human fibroblasts |
2. Results
2.1: Annexin A6 protein localizes to neural crest cells prior to, during and after EMT but then declines in migratory neural crest cells
We established the protein expression profile for Annexin A6 in the chick head from stages prior to EMT up to the formation of the cranial ganglia. Immunohistochemistry on chick cranial sections prior to (HH8), during (HH8–9) and post (HH9) EMT stages (Fig. 1) shows Annexin A6 expression throughout the neural tube (NT, red, Fig. 1A, C, E). Premigratory neural crest cells residing in the dorsal neural folds express Annexin A6 (Fig. 1A, A′, caret) (identified by immunohistochemistry for Snail2 on adjacent sections, Fig. 1B, B′, green, arrow, HH8). Annexin A6 expression is maintained (Fig. 1C, C′, caret, HH8+; Fig. 1E, E′, caret, HH9) in Snail2-positive premigratory neural crest cells as these cells round up during EMT (Fig. 1D, D′, arrow) as well as after these cells delaminate from the dorsal neural tube (Fig. 1F, F′, arrow), respectively. Furthermore, we observe localization of Annexin A6 to the chick heart (data not shown), corroborating prior studies showing that Annexin A6 is a major protein in atrial myocytes [29]. Interestingly, Annexin A6 expression (Fig. 2A) is absent in HNK-1-positive migratory neural crest cells by HH10 (Fig. 2B, C, C′, green, arrowheads). Neural tube expression of Annexin A6, however, still persists at this stage (Fig. 2A, caret).
Figure 1. Cranial neural crest cells at pre-EMT, during EMT and post-EMT express Annexin A6.

Representative transverse sections taken through the chick midbrain at various stages followed by immunohistochemistry for Annexin A6 (A, C, E, red). Immunohistochemistry was also performed for the premigratory and migratory neural crest cell marker Snail2 (B, D, F, green) on adjacent sections, as double immunohistochemistry could not be performed due to both antibodies being raised in rabbit. The dorsal neural folds boxed in each image (A–F) are shown at a higher magnification in (A′–F′). Arrows in B′, D′ and F′ indicate neural crest cells at premigratory (D′), EMT (E′), and newly emigrating (F′) stages. Annexin A6 is expressed by premigratory neural crest cells residing in the dorsal neural folds (caret, A′), by neural crest cells undergoing EMT (caret, C′), and by newly emigrating neural crest cells (caret, E′). The neural tube (NT) shows persistent Annexin A6 expression throughout all documented stages. DAPI (blue) labels cell nuclei. Scale bar, 50μm (A–F) and 125μm (A′–F′).
Figure 2. Annexin A6 expression is lost in the fully migratory neural crest cell population at HH10.
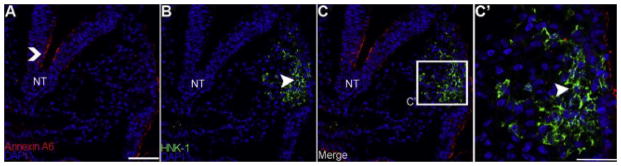
Representative transverse sections taken through the chick midbrain followed by immunohistochemistry for Annexin A6 (A, red) and the migratory neural crest cell marker HNK-1 (B, green), with a corresponding merge image (C). A higher magnification image of the boxed region in (C) is shown in (C′). The caret and arrowhead indicate Annexin A6 and HNK-1 expression, respectively. Annexin A6 expression is lost in migratory neural crest cells (B, C′, arrowhead). The neural tube (NT) still expresses Annexin A (A, caret). DAPI (blue) labels cell nuclei. Scale bar, 50μm (A–C) and 125μm (C′). Ectoderm is denoted by e.
2.2: Annexin A6 expression is absent in migratory neural crest cells but present in the placode cells that form the trigeminal and epibranchial ganglia
Although migratory cranial neural crest cells originating from the chick midbrain and hindbrain are devoid of Annexin A6, placode cells that begin to ingress from the ectoderm at HH13 to form the trigeminal and the epibranchial ganglia express Annexin A6 (Fig. 3). Trigeminal (Fig. 3A–D′), geniculate (Fig. 3E–H′), petrosal (Fig. 3I–L′) and nodose (Fig. 3M–P′) placode cells, identifiable by their Islet-1 immunoreactivity (Fig. 3C, G, K, O, purple, arrow), all express Annexin A6 (Fig. 3A, E, I, M, red, caret). Conversely, neural crest cells forming the proximal portion of the trigeminal (Fig. 3B, HNK-1, green, arrowhead), geniculate (Fig. 3F, HNK-1, green, arrowhead), petrosal (Fig. 3J, HNK-1, green, arrowhead) and nodose (Fig. 3N, HNK-1, green, arrowhead) ganglia at HH13 are not immunoreactive for Annexin A6. Islet-1- and Annexin A6-double-positive trigeminal and epibranchial placode cells are also revealed at a higher magnification (Fig. 3D′, trigeminal; Fig. 3H′, geniculate; Fig. 3L′, petrosal; Fig. 3P′ nodose, #).
Figure 3. Annexin A6 is absent in migratory neural crest cells but present in the placode cells contributing to the developing trigeminal and epibranchial ganglia at HH13.

Representative transverse sections taken through the chick trigeminal (A–D′), geniculate (E–H′), petrosal (I–L′), and nodose (M–P′) ganglia at HH13 followed by immunohistochemistry for Annexin A6 (A, E, I, M, red), HNK-1 (B, F, J, N, green) and Islet-1 (C, G, K, O, purple), with corresponding merge images shown (D, H, L, P). Carets, arrowheads, and arrows indicate Annexin A6-, HNK-1-, and Islet-1-positive cells, respectively. Annexin A6- and Islet-1-double-positive cells are shown by # in the merge images (D, H, L, P; higher magnification shown in (D′, H′, L′, P′). HNK-1-positive migratory neural crest cells (arrowhead) in the trigeminal (B), geniculate (F), petrosal (J), and nodose (N) ganglia are all Annexin A6-negative. DAPI (blue) labels cell nuclei. Scale bar, 50μm (A–L) and 125μm (C′–L′). Rhombomeres 1 and 2 are denoted by r1 and r2, respectively.
2.3 Annexin A6 expression is absent in migratory neural crest cells but present in the placode cells during later stages of trigeminal and epibranchial ganglia assembly
The placode cells that coalese and condense with neural crest cells at HH17 to form the trigeminal and the epibranchial ganglia express Annexin A6 (Fig. 4). Trigeminal (Fig. 4A–D′), geniculate (Fig. 4E–H′), petrosal (Fig. 4I–L′) and nodose (Fig. 4M–P′) placode cells, identifiable by the Islet-1 placodal marker (Fig. 4C, G, K, O, purple, arrow), all express Annexin A6 (Fig. 4A, E, I, M, red, caret). As observed at HH13, neural crest cells forming the trigeminal (Fig. 4B, HNK-1, green, arrowhead), geniculate (Fig. 4F, HNK-1, green, arrowhead), petrosal (Fig. 4J, HNK-1, green, arrowhead) and nodose (Fig. 4N, HNK-1, green, arrowhead) ganglia at HH17 continue to be devoid of Annexin A6 protein. Islet-1- and Annexin A6-double-positive trigeminal and epibranchial placode cells are also revealed at a higher magnification (Fig. 4D′, trigeminal; Fig. 4H′, geniculate; Fig. 4L′, petrosal; Fig. 4P′ nodose, #). Importantly, the expression of Annexin A6 in placode cells, and its absence in neural crest cells, is observed throughout cranial gangliogenesis (HH13–17; HH14, 15 and 16 not shown).
Figure 4. Annexin A6 is absent in migratory neural crest cells but present in the placode cells contributing to the developing trigeminal and epibranchial ganglia at HH17.

Representative transverse sections taken through the chick trigeminal (A–D′), geniculate (E–H′), petrosal (I–L′) and nodose (M–P′) ganglia at HH17 followed by immunohistochemistry for Annexin A6 (A, E, I, M, red), HNK-1 (B, F, J, N, green) and Islet-1 (C, G, K, O, purple), with corresponding merge images shown (D, H, L, P). Carets, arrowheads, and arrows indicate Annexin A6-, HNK-1-, and Islet-1-positive cells, respectively. Annexin A6- and Islet-1-double-positive cells are shown by # in the merge images (D, H, L, P; higher magnification shown in (D′, H′, L′, P′). HNK-1-positive migratory neural crest cells (arrowhead) in the trigeminal (B), geniculate (F), petrosal (J), and nodose (N) ganglia are all Annexin A6-negative. DAPI (blue) labels cell nuclei. Scale bar, 50μm (A–L) and 125μm (C′–L′). Rhombomeres 1 and 2 are denoted by r1 and r2, respectively.
2.4: Opthalmic and maxillomandibular placode cells express Annexin A6 during early stages of trigeminal ganglia formation
Although migratory neural crest cells are devoid of Annexin A6, ophthalmic (opV) and maxillomandibular (mmV) placode cells that will intermingle with them to form the different lobes of the trigeminal ganglia both express Annexin A6 (Fig. 5, HH13). These placode cells were distinguished by immunostaining for Pax3 (opV marker only) and Islet-1 (differentiating opV and mmV marker [30]). Annexin A6 (Fig. 5A, caret; Fig. 5D, #) is noted in a few Pax3- (Fig. 5B, green, arrowhead) and Islet-1- (Fig. 5C, purple, arrow) double-positive opV placode cells ingressed into the mesenchyme. Likewise, Annexin A6 (Fig. 5E, caret; Fig. 5H, #) is observed in Pax-3-negative (Fig. 5F, *) and Islet-1-positive (Fig. 5G, purple, arrow) mmV placode cells ingressing from the ectoderm and coalescing with neural crest cells.
Figure 5. Annexin A6 is expressed in the trigeminal placode cells during early trigeminal gangliogenesis.
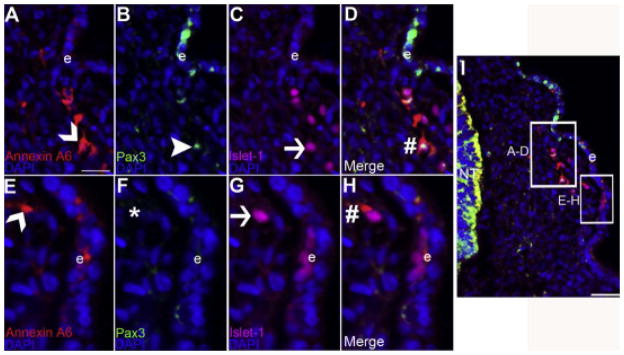
(A–H) High magnification images of the boxed regions shown in (I), which is a representative transverse section taken through the forming trigeminal ganglia at HH13 followed by immunohistochemistry for Annexin A6 (A, E, red, carets), the opV placode cell marker Pax3 (B, green, arrowhead), and the placode cell differentiation marker Islet-1 (C, G, purple, arrow). The opV and mmV placode cell populations are shown in (A–D) and (E–H), respectively. Pax3- (B) and Islet-1- (C) double-positive opV cells are Annexin A6-positive (A, caret), also visible in the merge image (D, #). Likewise, Pax3-negative (F, *) and Islet-1-positive (G) mmV cells are Annexin A6-positive (E, caret), also apparent in the merge image (H, #). DAPI (blue) labels cell nuclei. Scale bar, 125μm (A–H) and 50μm (I). Ectoderm is denoted by e.
2.5: Opthalmic and maxillomandibular placode cells express Annexin A6 at later stages of trigeminal ganglia formation
In the trigeminal ganglia at HH16, opV (Fig. 6A–D) and mmV (Fig. 6E–H) placode cells condensing with neural crest cells are immunoreactive for Annexin A6. As such, Annexin A6 (Fig. 6A, red, caret; Fig 6D, #) is observed in Pax3- (Fig. 6B, green, arrowhead) and Islet-1- (Fig. 6C, purple, arrow) double-positive opV placode cells in the mesenchyme. Similarly, Annexin A6 (Fig. 6E, red, caret; Fig. 6H, #) is noted in Pax-3-negative (Fig. 6F, *), Islet-1-positive (Fig. 6G, purple, arrow) mmV placode cells coalescing with neural crest cells.
Figure 6. Annexin A6 is expressed in the trigeminal placode cells during later trigeminal gangliogenesis.
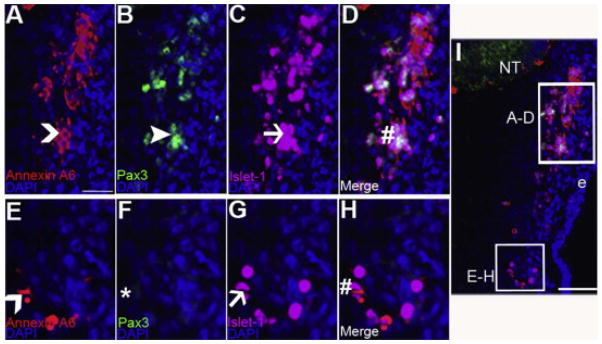
(A–H) High magnification images of the boxed regions shown in (I), which is a representative transverse section taken through the forming trigeminal ganglia at HH16 followed by immunohistochemistry for Annexin A6 (A, E, red, carets), the opV placode cell marker Pax3 (B, green, arrowhead), and the placode cell differentiation marker Islet-1 (C, G, purple, arrow). The opV and mmV placode cell populations are shown in (A–D) and (E–H), respectively. Pax3- (B) and Islet-1- (C) double-positive opV cells are Annexin A6-positive (A, caret), also visible in the merge image (D, #). Likewise, Pax3-negative (F, *) and Islet-1-positive (G) mmV cells are Annexin A6-positive (E, caret), also apparent in the merge image (H, #). DAPI (blue) labels cell nuclei. Scale bar, 125μm (A–H) and 50μm (I). Ectoderm is denoted by e.
3. Discussion
Given the role of Annexin A6 during chick cranial neural crest cell EMT and migration [20], we set out to further investigate Annexin A6 function at later stages of neural crest cell development. To this end, we employed immunohistochemistry for Annexin A6 to document Annexin A6 protein distribution in the developing chick head. In agreement with our previous data for Annexin A6 transcripts [20], we observe Annexin A6 protein throughout the neural tube including in the dorsal neural folds, within which premigratory neural crest cells reside. Throughout all stages examined, we note persistent expression of Annexin A6 in the developing neural tube (uniformly in the neural tube at early stages and more ventrally at later stages), and we speculate that this may be due to a general function for Annexin A6 in neuronal development (see below).
As premigratory neural crest cells undergo EMT and migrate out of the neural tube, they show persistent expression of Annexin A6 (HH8). Strikingly, ninety minutes later in development (during which time neural crest cells are actively migrating (HH9+)), we observe a loss of Annexin A6. This absence of Annexin A6 in neural crest cells persists throughout the formation of the cranial trigeminal and epibranchial ganglia. Since Annexin A6 has various documented roles in cell migration [16,17,19], cytoskeletal remodeling [21,22,24], and cell regulatory processes [23], we hypothesize that Annexin A6 is required for the initiation of neural crest cell EMT and early migration, but not for later neural crest cell migration, particularly as the cranial ganglia form.
Although Annexin A6 is lost in migratory neural crest cells by HH9+, we note Annexin A6 expression in the cranial sensory placode cells at HH13. Trigeminal placodal precursors in the ectoderm lack Annexin A6, but those precursors preparing to ingress from the ectoderm, or have already ingressed into the mesenchyme (opV and mmV neuronal precursors), express Annexin A6 and will later coalesce with neural crest cells to form the trigeminal ganglia. At the peak of trigeminal ganglia condensation (HH16), Annexin A6 expression persists in opV and mmV placodal cells within the mesenchyme.
As Annexin A6, and its vertebrate and invertebrate homologues, possesses a general function in ganglia development and physiology [11–13], we examined the pattern of Annexin A6 expression in the chick epibranchial sensory ganglia. Our results reveal the presence of Annexin A6 in geniculate, petrosal and nodose placode cells from early stages of ingression (HH13) to later stages of epibranchial ganglia formation (HH17). Interestingly, when we scanned the vestibulocochlear VIIIg ganglia (derived from just otic placode cells), we observed no Annexin A6 expression (data not shown), implying specific functions for Annexin A6 in sensory ganglia formed at different axial levels. Intriguingly, though, a subset of TuJ1-positive, post-mitotic neurons at the core of the forming neural crest cell-derived dorsal root ganglia express Annexin A6 (Fig. 7, HH20). Taken together with our cranial ganglia data, these observations suggest that one potential role of Annexin A6 could be in the development of embryonic cell types that possess neuronal characteristics and attributes. Our results also fall in line with and add to the known roles of Annexin A6 throughout evolution in the formation of the ganglia.
Figure 7. Annexin A6 is expressed in neurons found in the core of developing dorsal root ganglia.
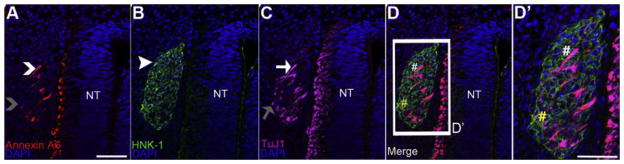
A representative transverse section taken through an HH20 chick trunk followed by immunohistochemistry for Annexin A6 (A, red, caret), the migratory neural crest cell marker HNK-1 (B, green, arrowhead), and the differentiating neuronal marker TuJ1 (C, purple, arrow), with a corresponding merge image (D). A higher magnification image of the boxed region in (D) is shown in (D′). Annexin A6 expression is present in a subset of post-mitotic neurons at the core of the developing dorsal root ganglia, indicated by TuJ1- and Annexin A6-double-positive cells (D, D′, white, #). A few of the TuJ1-positive neurons (C, gray arrow) are Annexin A6-negative (A, gray caret; D, D′, yellow, #). The neural tube (NT) still expresses Annexin A6, but the expression is confined to the lateral zone of differentiating neurons (A–D′). DAPI (blue) labels cell nuclei. Scale bar, 50μm (A–D) and 125μm (D′). Ectoderm is denoted by e.
Conclusion
The dynamic spatio-temporal expression of Annexin A6 during chick cranial gangliogenesis sheds light on how embryos re-deploy the same protein at different developmental times and in diverse embryonic cell types. Our results demonstrate a conserved role for Annexin A6 in cranial sensory placodes and may have implications for general tissue formation during vertebrate development.
4. Experimental Procedures
Materials and Methods
Chick embryos
Fertilized chicken eggs were obtained from B & E Farms (York, PA, USA) and incubated at 38°C in humidified incubators (EggCartons.com, Manchaug, MA, USA). Embryos were staged according to Hamburger Hamilton in later development [31].
Immunohistochemistry
Embryos were collected at the documented stages and fixed in 4% paraformaldehyde (PFA) overnight at room temperature. The embryos were permeabilized in fresh Phosphate-buffered saline (PBS) containing 0.5% TX-100 (0.5% PBSTX), gelatin embedded and transverse sectioned at 12μm. Antigen retrieval (required for use of the Annexin A6 antibody) was performed by incubating de-gelatinized sections at 94° C for 20 minutes in 0.1 M sodium citrate. The following primary antibodies and concentrations were diluted in 0.1% TX-100 + 1X PBS (0.1% PBSTX) + 5% sheep serum: Annexin A6 (1:100, Abnova, PAB18085), Pax3 (1:200, Developmental Studies Hybridoma Bank (DSHB)), Islet-1 (1:250, DSHB, 40.2D6), Snail2 (1:200, Cell Signaling Technologies, C19G7), and TuJ1 (1:500, Covance, MMS-435P). The following secondary antibodies from Life Technologies were obtained and used at a 1:200 dilution (with the exception of TuJ1, which was used at 1:500) in 0.1 % PBSTX+ 5% sheep serum: Goat anti-rabbit IgG (Annexin A6), goat anti-mouse IgG2a (Pax3 and TuJ1), goat anti-mouse IgG1 (Islet-1), and goat anti-rabbit IgG (Snail2). Sections were viewed at room temperature using a Zeiss AxioObserver.Z1 inverted microscope, and images were acquired using the Zeiss Axiovision Rel 4.6 software with the Zeiss Axiocam HRC camera. All exported images were processed in Adobe Photoshop 9.0 (Adobe Systems).
Highlights.
Annexin A6 is dynamically expressed in distinct cell types during development.
Neural crest cells prior to, during and after their emigration from the neural tube express Annexin A6, but fully migratory neural crest cells are devoid of the protein.
Trigeminal and epibranchial placodes possess Annexin A6 protein at the beginning of ganglia formation, and this expression is persistent throughout stages of ganglia condensation.
Acknowledgments
We thank Ms. Lizbeth Hu for excellent technical assistance. This work was supported by grants to L.A.T. (NSF IOS-0948525, NIH R01DE024217A).
Abbreviations
- EMT
epithelial-to-mesenchymal transition
- opV
ophthalmic
- mmV
maxillomandibular
- HH
Hamburger Hamilton
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Hamburger V. Experimental analysis of the dual origin of the trigeminal ganglion in the chick embryo. J Exp Zool. 1961;148:91–124. doi: 10.1002/jez.1401480202. [DOI] [PubMed] [Google Scholar]
- 2.Begbie J, Graham A. Integration between the epibranchial placodes and the hindbrain. Science. 2001;294(5542):595–598. doi: 10.1126/science.1062028. [DOI] [PubMed] [Google Scholar]
- 3.Saint-Jeannet J, Moody S. Establishing the pre-placodal region and breaking it into placodes with distinct identities. Dev Biol. 2014;389:13–27. doi: 10.1016/j.ydbio.2014.02.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Park B, Saint-Jeannet J. Induction and segregation of the vertebrate cranial placodes. San Rafael, CA: Morgan & Claypool Life Sciences; 2010. [PubMed] [Google Scholar]
- 5.Schlosser G. Induction and specification of cranial placodes. Developmental Biology. 2005;294:303–351. doi: 10.1016/j.ydbio.2006.03.009. [DOI] [PubMed] [Google Scholar]
- 6.Jidigam VK, Gunhaga L. Development of cranial placodes: Insights from studies in chick. Development, Growth and Differentiation. 2013;55:79–95. doi: 10.1111/dgd.12027. [DOI] [PubMed] [Google Scholar]
- 7.Bronner ME, LeDouarin NM. Development and evolution of the neural crest: An overview. Developmental Biology. 2012;366(1):2–9. doi: 10.1016/j.ydbio.2011.12.042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.CDC. 2014 [Google Scholar]
- 9.NIH/NINDS fact sheet. 2014 [Google Scholar]
- 10.Donkelaar HJ, Lammens M, Hori A. Clinical Neurobiology: Development and developmental disorders of the human central nervous system. Springer; 2006. [Google Scholar]
- 11.Lizarbe MA, Barrasa JI, Olmo N, Gavilanes F, Turnay J. Annexin-Phospholipid interactions. Functional Implications. International Journal of Molecular Sceinces. 2013;14:2652–2683. doi: 10.3390/ijms14022652. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Kerschbaum H, Donato R, Hermann A. Annexin-immunoreactive proteins in the nervous system and eye of the gastropods, Aplysia and Helix. Brain Research. 1997;746(1–2):133–140. doi: 10.1016/s0006-8993(96)01217-6. [DOI] [PubMed] [Google Scholar]
- 13.Naciff J, Kaetzel M, Behbehani M, Dedman J. Differential expression of annexins IVI in the rat dorsal root ganglia and spinal cord. Journal of Comparative Neurology. 1998;368(3):356–370. doi: 10.1002/(SICI)1096-9861(19960506)368:3<356::AID-CNE3>3.0.CO;2-4. [DOI] [PubMed] [Google Scholar]
- 14.Foulkes T, Nassar M, Lane T, Mathews E, Baker M, Gerke V, Okuse K, Dickenson A, Wood J. Deletion of Annexin 2 light chain p11 in nociceptors causes deficits in somatosensory coding and pain behavior. The Journal of Neuroscience. 2006;26(41):10499–10507. doi: 10.1523/JNEUROSCI.1997-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Li Q, Gaun H, Pu X. Identification of a 35kDa protein in rat spinal ganglia and sensory fibres. Neuroreport. 2004;15(13):2083–7. doi: 10.1097/00001756-200409150-00017. [DOI] [PubMed] [Google Scholar]
- 16.Gerke V, Cruetz CE, Moss SE. Annexins: Linking Ca2+ signalling to membrane dynamics. Nature reviews: Molecular Cell Biology. 2005;6:449–576. doi: 10.1038/nrm1661. [DOI] [PubMed] [Google Scholar]
- 17.Rescher U, Gerke V. Annexins—unique membrane binding proteins with diverse functions. J Cell Science. 2004;117:2631–2639. doi: 10.1242/jcs.01245. [DOI] [PubMed] [Google Scholar]
- 18.Mussunoor S, Murray GI. The role of annexins in tumor development and progression. J pathol. 2008;216(2):131–40. doi: 10.1002/path.2400. [DOI] [PubMed] [Google Scholar]
- 19.Moss SE, Morgan RO. The annexins. Genome Biol. 2004;5(4):219. doi: 10.1186/gb-2004-5-4-219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Wu C, Taneyhill L. Annexin a6 modulates chick cranial neural crest cell emigration. PLosOne. 2012;7:e44903. doi: 10.1371/journal.pone.0044903. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Locate S, Coyler J, Gawler D, Walker J. Annexin A6 at the cardiac myocytes sarcolemma-Evidence for self-association and binding to actin. Cell Biology International. 2008;32:1388–1396. doi: 10.1016/j.cellbi.2008.08.009. [DOI] [PubMed] [Google Scholar]
- 22.Monastyrskaya M, Babiychuk B, Hostettler A, Wood P, Grewal T, Draeger A. Plasma membrane associated Annexin A6 reduces Ca2+ entry by stabilizing the cortical actin cytoskeleton. The Journal of Biological Chemistry. 2009;284:17227–17242. doi: 10.1074/jbc.M109.004457. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Mishra S, Chander V, Banerjee P, Oh JG, Lifirsu E, Park WJ, Kim DH, Bandyopadhyay A. Interaction of annexin A6 with alpha actinin in cardiomyocytes. BMC Cell Biology. 2011;12:7. doi: 10.1186/1471-2121-12-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Monastyrskaya M, Babiychuk B, Hostettler A, Wood P, Grewal T, Draeger A. Plasma membrane associated Annexin A6 reduces Ca2+ entry by stabilizing the cortical actin cytoskeleton. The Journal of Biological Chemistry. 2009;284:17227–17242. doi: 10.1074/jbc.M109.004457. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Camors E, Monceau V, Charlemagne D. Annexins and Ca2+ handling in the heart. Cardiovascular research. 2004;65:793–802. doi: 10.1016/j.cardiores.2004.11.010. [DOI] [PubMed] [Google Scholar]
- 26.Cornely R, Rentero C, Enrich C, Grewal T, Gaus K. Annexin A6 is an organizer of membrane microdomains to regulate receptor localization and signalling. IUBMB Life. 2011;63(11):1009–17. doi: 10.1002/iub.540. [DOI] [PubMed] [Google Scholar]
- 27.Chylstun M, Campanella M, Law A, Duchen MR, Fatimathas L, Levine TP, Gerke V, Moss SE. Regulation of mitochondrial morphogenesis by Annexin A6. PLOS ONE. 2013;8:1. doi: 10.1371/journal.pone.0053774. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Sakwe AM, Koumangoye R, Guillory B, Ochieng J. Annexin A6 contributes to the invasiveness of breast carcinoma cells by influencing the organization and localization of functional focal adhesions. Experimental cell research. 2011;317:823–837. doi: 10.1016/j.yexcr.2010.12.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Doubell AF, Bester AJ, Thibault G. Annexins V and VI: major calcium-dependent atrial secretory granule-binding proteins. Hypertension. 1991;5:648–56. doi: 10.1161/01.hyp.18.5.648. [DOI] [PubMed] [Google Scholar]
- 30.Xu H, Dude C, Baker C. Fine-grained fate maps for the opthalmic and maxillomandibular trigeminal placodes in the chick embryo. Develpmental Biology. 2008;317(1):174–186. doi: 10.1016/j.ydbio.2008.02.012. [DOI] [PubMed] [Google Scholar]
- 31.Hamburger V, Hamilton HL. A series of normal stages in the development of the chick embryo. Developmental dynamics : an official publication of the American Association of Anatomists. 1992;1951(195):231–272. doi: 10.1002/aja.1001950404. [DOI] [PubMed] [Google Scholar]