

Abstract
Purpose: Hypoxia is one of the important factors in formation of reactive oxygen species (ROS). Ghrelin is a peptide hormone that reduces oxidative stress. However, antioxidant effect of ghrelin on blood and brain in normobaric hypoxia condition has not yet been investigated.
Methods: thirty-two animals were randomly divided into four (n=8) experimental groups: Control (C), ghrelin (Gh), hypoxia (H), hypoxic animals that received ghrelin (H+Gh). Normobaric systemic hypoxia (11% O2) was induced in rats for 48 hours. Effect of ghrelin (80 μg/kg, i.p) on serum TAC and MDA and brain SOD, CAT, GPx and MDA were assessed.
Results: Hypoxia significantly (p<0.001) increased both blood and brain MDA Levels. Ghrelin treatment significantly (p<0.001) decreased blood MDA levels both in control and hypoxia, and brain MDA levels in hypoxia conditions. Brain SOD, CAT and GPx variations were not significant in two days of hypoxia. Ghrelin treatment also could not significantly increase activity of SOD, CAT and GPx in brain. Total antioxidant capacity of serum increased in ghrelin treatment both in control and hypoxic conditions, although it was only significant (p<0.01) in control conditions.
Conclusion: Our findings showed that administration of ghrelin may be useful in reducing blood and brain oxidative stress in normobaric hypoxia condition.
Keywords: Normobaric Hypoxia, Ghrelin, Oxidative stress, Blood, Brain
Introduction
Hypoxia is a decrease in tissue oxygen concentration, normally caused by a reduction in the partial pressure of atmospheric oxygen.1 Hypoxia is an important pathogenic factor in different injuries of CNS such as cerebral ischemia, tumorigenesis, head injury, acute mountain sickness, and in pulmonary disease such as COPD.2-5 Due to the exposition to hypoxia, reactive oxygen species are continuously generated.6 The rate of ROS production from mitochondria is also increased in a variety of pathologic conditions including ischemia, reperfusion, aging and chemical inhibition of mitochondrial respiration.7 Reactive oxygen species (ROS) such as superoxide anion, hydrogen peroxide, and hydroxyl radical are generated continuously in cells grown aerobically.8 These free radicals can interact with bimolecules such as DNA, carbohydrates, protein, and lipids and damage various cellular components.9 An increase in free radicals causes overproduction of malonyl dialaldehyde (MDA) in oxidative stress conditions.10
The brain has a high oxygen consumption, which comprises 20-25% of total body O2 consumption.11,12 In addition, neurons comprise about 5% of the brain volume, whereas they consume one-quarter of total cerebral O2.12 Oxygen, due to its radical nature together with its usefulness as an electron acceptor, is also a source of free radicals and consequently a potent toxin for the cells.1 Therefore, the central nervous system is vulnerable to oxidative damage.13
Ghrelin, a 28-amino acid peptide, is a brain-gut peptide.14 Ghrelin passes into the BBB following systemic administration.15 Its receptors are expressed in various regions of the brain, such as hypothalamus, pituitary, hippocampus, and cerebral cortex.16 Studies have indicated that ghrelin has multiple functions in the CNS such as learning and memory, anxiety and depression, reward and motivation, neuroprotection, and neurogenesis.17 Ghrelin also decreases brain edema induced by hypoxia.18 In addition, ghrelin has several anti-inflammatory properties and decreases neuronal apoptosis.19-22 Antioxidant properties of ghrelin have also been demonstrated in recent studies.23-25
Therefore, the aim of this study was to evaluate the effects of ghrelin on the blood levels of MDA and TAC and brain levels of MDA, SOD, GPx and CAT in the normobaric systemic hypoxia situation in rats.
Materials and Methods
Animals
Adults male Wistar rats (220-250g) were housed in standard cages in a temperature controlled room (22-24ºC), humidity (40-60%), and light period (12h dark-12h light). Food and water were available ad libitum. All experiments were performed in agreement with guidelines of the Tabriz University of Medical Sciences for care and use of laboratory animals.
Experimental groups
Thirty two animals were randomly divided into four (n=8) experimental groups:
Control (C): animals that were placed in room air (21% O2)
Ghrelin (Gh): animals that received ghrelin (80 μg/kg/day, i.p) for two days
Hypoxia (H): animals exposed to 11% O2 for two days
Hypoxia with ghrelin (H + Gh): animals exposed to 11% O2 and received ghrelin (80 μg/kg/day, i.p) for two days.
Control and hypoxia groups received saline (1ml/kg/day, i.p.) and ghrelin (Innovagen, Sweden) was administered intraperitoneally at a dose of 80μg/kg once daily for 2 consecutive days.18
Hypoxia induction
Environmental chamber :
Control and ghrelin groups were placed in room air (21% O2), hypoxic groups were placed into the hypoxia chamber (GO2 Altitude-Biomedical Australia). Systemic normobaric hypoxia (10.5%-11% O2) was induced for 48 h. Animals were daily emitted for 10-15 min to clean the cages and receive injections.18
Tissue processing and homogenate preparation :
At the end of experiment, rats were deeply anesthetized with ketamin (60 mg/kg) and xylasin (6 mg/kg), decapitated and blood samples were collected from trunk vessels. Immediately, brains were excised, frozen in liquid nitrogen and stored at deep freeze (-70 °C) for later measurements. For antioxidant activities measurement the brain samples were homogenized in 1.15% KCl solution. The homogenates were centrifuged at 1000 rpm for 1 min at 4°C. Then, tissue homogenate were used for determination of lipid peroxides (MDA) and activities of SOD, GPx and catalase.26
Total Antioxidant Capacity, Catalase, Malonyl dealdehyde Assessment :
The total antioxidant capacity (TAC) in homogenates was measured using a commercially kit, according to the manufacturer's instruments and with reference to the Miller method.27 Catalase activity in blood and tissue homogenates was determined based on its ability to decompose H2O2 and measured at 240 nm.28 Malonyl dealdehyde (MDA) levels were measured using the thiobarbituric acid reactive substances (TBARS) method.29
Glutathione Peroxidase and Superoxide Dismutase Assessment in brain :
To measure cytosolic enzyme activity, the brain samples were homogenized in 1.15% KCl solution. Glutathione peroxidase (GPx) activity was measured according to Paglia and Valentine using Randox (United Kingdom). Tissue superoxide dismutase (SOD) was assayed by a spectrophotometric method based on the inhibition of a superoxide-induced reduced nicotinamide adenine dinucleotide (NADH) oxidation according to Paoletti et al.26
Statistical analysis :
Data were statistically analyzed using One way analysis of variance (ANOVA) followed by Tukey’s test. The significant level was set at p<0.05. Results are expressed as Means± SEM.
Results
Effect of ghrelin on blood MDA level in normobaric hypoxia
The effect of two days of ghrelin (80 µg/kg/day) treatment on blood MDA level in animals showed that administration of ghrelin significantly (p<0.001) decreased MDA level in (Gh) group compared to control group. Induction of normobaric hypoxia (11% O2) significantly increased blood MDA level in (H) group. Administration of ghrelin also significantly (p<0.001) decreased MDA level in (H+Gh) group compared to (H) group. Interestingly, administration of ghrelin significantly (p<0.001) decreased MDA level in (H+Gh) group compared to both control and (H) group (Figure 1).
Figure 1.
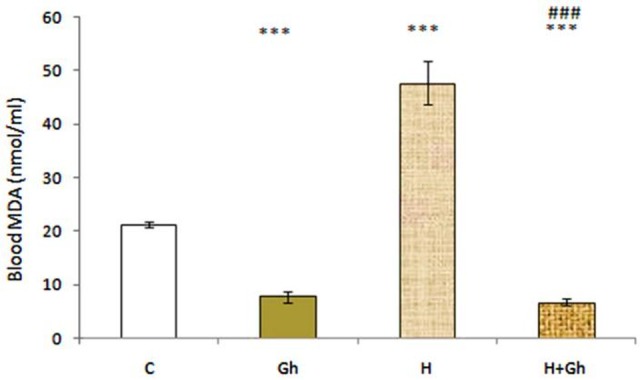
Depicts the effect of two days of ghrelin (80 µg/kg/day) treatment on blood MDA level. Groups are abbreviated as following: Control (C), animals that received ghrelin (Gh), hypoxia (H), hypoxic animals that received ghrelin (H+GH). Data are shown as Mean±SEM. *** p<0.001 when compared with control group and ### p<0.001 when compared to hypoxia group.
Effect of ghrelin on brain MDA level in normobaric hypoxia
Induction of normobaric hypoxia (11% O2) significantly (p<0.001) increased brain MDA level in (H) group compared to control group. The effect of two days of ghrelin (80 µg/kg/day) treatment on brain MDA level in (H+GH) groups showed a significant (p<0.001) decrease in brain MDA levels Compared to (H) group (Figure 2).
Figure 2.
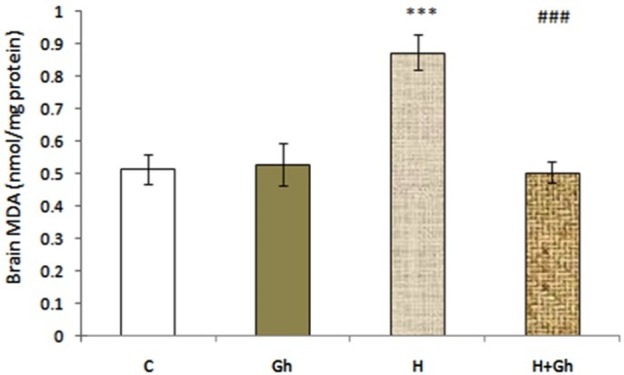
Depicts the effect of two days of ghrelin (80 µg/kg/day) treatment on brain MDA level. Groups are abbreviated as following: Control (C), animals that received ghrelin (Gh), hypoxia (H), animals with hypoxic condition that received ghrelin (H+GH). Data are shown as Mean±SEM. *** p<0.001 when compared with control group and ### p<0.001 when compared to acute hypoxia group.
Effect of ghrelin on serum TAC level in normobaric hypoxia
The effect of two days of ghrelin (80 µg/kg/day) treatment on serum total antioxidant capacity (TAC) level in (Gh) group showed that administration of ghrelin significantly (p<0.01) increased TAC level in (Gh) group compared to control group. Administration of ghrelin although increased serum TAC level in (H+Gh) group compared with (H) group, it was not significant (Figure 3).
Figure 3.
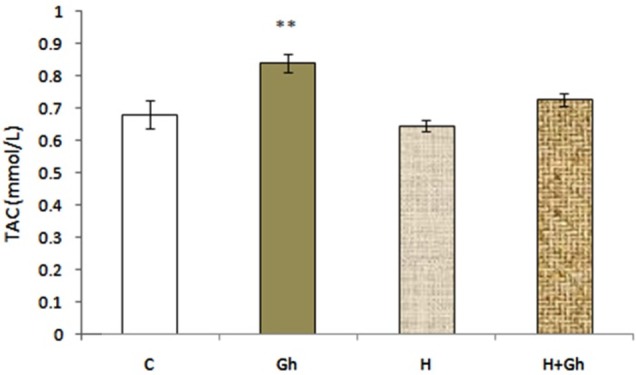
Depicts the effect of two days of ghrelin (80 µg/kg/day) treatment on serum TAC level. Groups are abbreviated as following: Control (C), animals that received ghrelin (GH), hypoxia (H), animal with hypoxia that received ghrelin (H+GH). Data are shown as Mean±SEM. **p<0.01 when compared with control group.
Effect of ghrelin on brain SOD, GPx and CAT level in normobaric hypoxia
The effect of two days of ghrelin (80 µg/kg/day) treatment in hypoxia investigated on brain SOD, GPx and CAT levels. SOD, GPx, and CAT levels in brain homogenates did not significantly change by hypoxia or ghrelin treatment (Table 1).
Table 1. Brain Tissue SOD, GPx, and CAT activity.
| Enzymes | C | Gh | H | H+Gh |
| SOD (U/ mg protein) | 2.37±0.05 | 2.72±0.15 | 2.41±0.10 | 2.54±0.06 |
| GPx (U/ mg protein) | 0.147±0.002 | 0.162±0.006 | 0.147±0.004 | 0.159±0.003 |
| CAT (U/ mg protein) | 0.006±0.003 | 0.022±0.017 | 0.005±0.001 | 0.010±0.002 |
Tab 1. Depicts the effect of two days of hypoxia and ghrelin (80 µg/kg/day) treatment on brain SOD, GPx, and CAT level. Data are shown as Mean±SEM.
Discussion
In the present study, induction of normobaric hypoxia in rats led to an increase in oxidative stress both in blood and brain tissues; and treatment of rats with ghrelin (80 µg/kg/day) decreased blood and brain MDA levels in hypoxia. Ghrelin also increased serum TAC levels in control and hypoxia conditions, although it was only significant in control condition.
Oxygen is an essential element for most organisms and various redox reactions in the cell need oxygen. In addition, oxygen may be a source of the formation of reactive oxygen species.12 The level of oxygen, low (hypoxia) or high (hyperoxia), can induce oxidative injury and increase morbidity and mortality.30 Studies have proved that the intensity of oxidative stress is proportional to the exposure time and the severity of hypoxia.31 The exposure of cells or tissues to moderate hypoxia increases the production of ROS by the mitochondria and the oxidative stress associated with increased generation of ROS triggers apoptosis.32,33
In our study, two days of hypoxia (11% O2) led to an oxidative stress both in blood and brain and ghrelin exhibited some antioxidant properties. Ghrelin has multiple functions in the CNS such as neuroprotection and neurogenesis.17 Intrahippocampal injection of ghrelin has been shown to have inhibitory effect against PTZ-induced seizure that is probably mediated via brain GABAergic or neuropeptide-Y systems.34-36 Previous studies have also demonstrated that ghrelin pretreatment diminishes oxidative stress and prevents the decrease in antioxidant enzyme activities in erythrocytes, liver and brain in a PTZ model of seizure.37 In addition, ghrelin pretreatment reduced the level of lipid peroxidation and enhanced the antioxidant defense against exercise-induced stress oxidative injury in rats’ brain and liver.38 Ghrelin also have some protective effects against the oxidative brain and kidney injuries induced by carbon tetrachloride.39 Kheradmand’s studies showed that ghrelin has antioxidant properties in the rat testis and ovary tissues.40,41 Moreover, one study proposed that ghrelin increases activity of SOD, CAT, GSH-Px in 3T3 L1 preadipocytes.42
In our study activity of brain GPx, SOD and catalase were not influenced significantly by ghrelin administration. In this context, it can be explained that the contents and activities of enzymes of antioxidant system in adult brain are lower than other organs, such as liver, kidneys and heart.12 Studies have also proved that superoxide dismutase content and activity of catalase in the rat brain is low.43,44 Moreover, although GPx is considered to have special importance for the brain antioxidant defense system, in the rat brain GPx activity is two times lower compared to the liver.12,45 According to these explanations, probably, brain tissue utilizes other antioxidant defense against oxidative stress.
Fat-soluble antioxidants, such as α-tocopherols, coenzyme Q, and α-lipolic acid are very important for the brain.44,46 Ascorbate is another vital antioxidant molecule in the brain that can be rapidly accumulated in the brain.47 Low-molecular-weight antioxidants also contribute to brain antioxidant defense. Thyoredoxin system and histidine-containing compounds, such carnosine, homocarnosine, and anserine are very important antioxidants in brain.41,48 The ability of carnosine to protect tissues against oxidative stress has been previously demonstrated in in vivo experiments under hypoxic and ischemic conditions.49,50
Involvement of some other mechanisms of antioxidant effect of ghrelin should also be considered. The NADPH oxidase is a multi-subunit enzyme that catalyzes the reduction of molecular oxygen to form superoxide (O2•−) and associated reactive oxygen species in the CNS.51 Tong et al showed in their study that ghrelin decreased intracellular reactive oxygen species content by inhibiting NADPH oxidases 1.52 Therefore, in our study ghrelin may have played its antioxidant role in the brain through inhibition of the NADPH oxidase or potentiation of the effect of Low-molecular-weight antioxidants.
Conclusion
In summary, our findings showed that administration of ghrelin may be useful in reducing blood and brain oxidative stress in normobaric hypoxia condition. Ghrelin shows its antioxidant effect in blood by increasing the total antioxidant capacity.
Acknowledgments
This study was financially supported by the Drug Applied Research Centre of Tabriz University of Medical Sciences. The article is derived from the MSc dissertation of Mr Hasan Omrani entitled "Effect of ghrelin on acute and chronic systemic hypoxia-induced oxidative stress in brain of adult male rats".
Ethical Issues
Not applicable.
Conflict of Interest
The authors report no conflicts of interest.
References
- 1.Carbonell T, Mara M. Respiratoty hypoxia and oxidative stress in brain. Is the endogenous erythropoietin an antioxidant? Curr Chem Biol. 2009;3(3):238–52. doi: 10.2174/2212796810903030238. [DOI] [Google Scholar]
- 2.Dong Z, Wang JZ, Yu F, Venkatachalam MA. Apoptosis-resistance of hypoxic cells: multiple factors involved and a role for IAP-2. Am J Pathol. 2003;163(2):663–71. doi: 10.1016/s0002-9440(10)63693-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Pilitsis JG, Rengachary SS. Complications of head injury. Neurol Res. 2001;23(2-3):227–36. doi: 10.1179/016164101101198389. [DOI] [PubMed] [Google Scholar]
- 4.Hackett PH. High altitude cerebral edema and acute mountain sickness. A pathophysiology update. Adv Exp Med Biol. 1999;474:23–45. doi: 10.1007/978-1-4615-4711-2_2. [DOI] [PubMed] [Google Scholar]
- 5.Pierson DJ. Pathophysiology and clinical effects of chronic hypoxia. Respir Care. 2000;45(1):39–51; discussion 51-3. [PubMed] [Google Scholar]
- 6.Turrens JF. Mitochondrial formation of reactive oxygen species. J Physiol. 2003;552(Pt 2):335–44. doi: 10.1113/jphysiol.2003.049478. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Chen Q, Vazquez EJ, Moghaddas S, Hoppel CL, Lesnefsky EJ. Production of reactive oxygen species by mitochondria: central role of complex III. J Biol Chem. 2003;278(38):36027–31. doi: 10.1074/jbc.m304854200. [DOI] [PubMed] [Google Scholar]
- 8.Cabiscol E, Tamarit J, Ros J. Oxidative stress in bacteria and protein damage by reactive oxygen species. Int Microbiol. 2000;3(1):3–8. [PubMed] [Google Scholar]
- 9.Devasagayam TP, Tilak JC, Boloor KK, Sane KS, Ghaskadbi SS, Lele RD. Free radicals and antioxidants in human health: current status and future prospects. J Assoc Physicians India. 2004;52:794–804. [PubMed] [Google Scholar]
- 10.Gawel S, Wardas M, Niedworok E, Wardas P. Malondialdehyde (MDA) as a lipid peroxidation marker. Wiad Lek. 2004;57(9-10):453–5. [PubMed] [Google Scholar]
- 11.Halliwell B, Gutteridge MC. Oxygen radicals and the nervous system. Trends Neurosci. 1985;8:22–6. doi: 10.1016/0166-2236(85)90010-4. [DOI] [Google Scholar]
- 12.Galkina OV. The specific features of free-radical processes and the antioxidant defense in the adult brain. Neruochemical J. 2013;30(2):89–97. doi: 10.1134/s1819712413020025. [DOI] [Google Scholar]
- 13.Wang X, Michaelis EK. Selective neuronal vulnerability to oxidative stress in the brain. Front Aging Neurosci. 2010;2:12. doi: 10.3389/fnagi.2010.00012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Korbonits M, Goldstone AP, Gueorguiev M, Grossman AB. Ghrelin--a hormone with multiple functions. Front Neuroendocrinol. 2004;25(1):27–68. doi: 10.1016/j.yfrne.2004.03.002. [DOI] [PubMed] [Google Scholar]
- 15.Banks WA, Tschop M, Robinson SM, Heiman ML. Extent and direction of ghrelin transport across the blood-brain barrier is determined by its unique primary structure. J Pharmacol Exp Ther. 2002;302(2):822–7. doi: 10.1124/jpet.102.034827. [DOI] [PubMed] [Google Scholar]
- 16.Zigman JM, Jones JE, Lee CE, Saper CB, Elmquist JK. Expression of ghrelin receptor mRNA in the rat and the mouse brain. J Comp Neurol. 2006;494(3):528–48. doi: 10.1002/cne.20823. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Andrews ZB. The extra-hypothalamic actions of ghrelin on neuronal function. Trends Neurosci. 2011;34(1):31–40. doi: 10.1016/j.tins.2010.10.001. [DOI] [PubMed] [Google Scholar]
- 18.Hossienzadeh F, Babri S, Alipour MR, Ebrahimi H, Mohaddes G. Effect of ghrelin on brain edema induced by acute and chronic systemic hypoxia. Neurosci Lett. 2013;534:47–51. doi: 10.1016/j.neulet.2012.11.062. [DOI] [PubMed] [Google Scholar]
- 19.Granado M, Priego T, Martin AI, Villanua MA, Lopez-Calderon A. Anti-inflammatory effect of the ghrelin agonist growth hormone-releasing peptide-2 (GHRP-2) in arthritic rats. Am J Physiol Endocrinol Metab. 2005;288(3):E486–92. doi: 10.1152/ajpendo.00196.2004. [DOI] [PubMed] [Google Scholar]
- 20.Pamukcu O, Kumral ZN, Ercan F, Yegen BC, Ertem D. Anti-inflammatory effect of obestatin and ghrelin in dextran sulfate sodium-induced colitis in rats. J Pediatr Gastroenterol Nutr. 2013;57(2):211–8. doi: 10.1097/mpg.0b013e318294711e. [DOI] [PubMed] [Google Scholar]
- 21.Chung H, Kim E, Lee DH, Seo S, Ju S, Lee D. et al. Ghrelin inhibits apoptosis in hypothalamic neuronal cells during oxygen-glucose deprivation. Endocrinology. 2007;148(1):148–59. doi: 10.1210/en.2006-0991. [DOI] [PubMed] [Google Scholar]
- 22.Lee J, Lim E, Kim Y, Li E, Park S. Ghrelin attenuates kainic acid-induced neuronal cell death in the mouse hippocampus. J Endocrinol. 2010;205(3):263–70. doi: 10.1677/joe-10-0040. [DOI] [PubMed] [Google Scholar]
- 23.El Eter E, Al Tuwaijiri A, Hagar H, Arafa M. In vivo and in vitro antioxidant activity of ghrelin: Attenuation of gastric ischemic injury in the rat. J Gastroenterol Hepatol. 2007;22(11):1791–9. doi: 10.1111/j.1440-1746.2006.04696.x. [DOI] [PubMed] [Google Scholar]
- 24.Liu L, Xu H, Jiang H, Wang J, Song N, Xie J. Ghrelin prevents 1-methyl-4-phenylpyridinium ion-induced cytotoxicity through antioxidation and NF-kappaB modulation in MES23.5 cells. Exp Neurol. 2010;222(1):25–9. doi: 10.1016/j.expneurol.2009.11.009. [DOI] [PubMed] [Google Scholar]
- 25.Alantary AK, Rezk MY, Soliman Kh EA. Protective effect of ghrelin on paracetamol induced acute hepatotoxicity in rats. J Physiol Pathophysiol. 2014;5(2):7–14. doi: 10.5897/JPAP2014.0091. [DOI] [Google Scholar]
- 26.Somi MH, Hajipour B, Asl NA, Estakhri R, Azar AN, Zade MN. et al. Pioglitazone attenuates ischemia/reperfusion-induced liver injury in rats. Transplant Proc. 2009;41(10):4105–9. doi: 10.1016/j.transproceed.2009.09.075. [DOI] [PubMed] [Google Scholar]
- 27.Miller NJ, Rice-Evans C, Davies MJ, Gopinathan V, Milner A. A novel method for measuring antioxidant capacity and its application to monitoring the antioxidant status in premature neonates. Clin Sci (Lond) 1993;84(4):407–12. doi: 10.1042/cs0840407. [DOI] [PubMed] [Google Scholar]
- 28.Luck H. Catalase. In: Bergmeyer HU, editor. Methods of enzymatic analysis. New York: Academic Press;1965. [Google Scholar]
- 29.Kaya H, Sezik M, Ozkaya O, Dittrich R, Siebzehnrubl E, Wildt L. Lipid peroxidation at various estradiol concentrations in human circulation during ovarian stimulation with exogenous gonadotropins. Horm Metab Res. 2004;36(10):693–5. doi: 10.1055/s-2004-826018. [DOI] [PubMed] [Google Scholar]
- 30.Zhao HW, Haddad GG. Review: Hypoxic and oxidative stress resistance in Drosophila melanogaster. Placenta. 2011;32 Suppl 2:S104–8. doi: 10.1016/j.placenta.2010.11.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Maiti P, Singh SB, Sharma AK, Muthuraju S, Banerjee PK, Ilavazhagan G. Hypobaric hypoxia induces oxidative stress in rat brain. Neurochem Int. 2006;49(8):709–16. doi: 10.1016/j.neuint.2006.06.002. [DOI] [PubMed] [Google Scholar]
- 32.Guzy RD, Hoyos B, Robin E, Chen H, Liu L, Mansfield KD. et al. Mitochondrial complex III is required for hypoxia-induced ROS production and cellular oxygen sensing. Cell Metab. 2005;1(6):401–8. doi: 10.1016/j.cmet.2005.05.001. [DOI] [PubMed] [Google Scholar]
- 33.Satoh T, Enokido Y, Aoshima H, Uchiyama Y, Hatanaka H. Changes in mitochondrial membrane potential during oxidative stress-induced apoptosis in PC12 cells. J Neurosci Res. 1997;50(3):413–20. doi: 10.1002/(SICI)1097-4547(19971101)50:3<413::AID-JNR7>3.0.CO;2-L. [DOI] [PubMed] [Google Scholar]
- 34.Babri S, Ghahramanian Golzar M, Ataie Z, Ebrahimi H, Mirzaie F, Mohaddes G. Effect of acute and chronic intrahippocampal microinjection of ghrelin on pentylentetrazole-induced seizures in rats. Pharm Sci. 2011;17(1):11–8. [Google Scholar]
- 35.Ataie Z, Babri S, Ghahramanian Golzar M, Ebrahimi H, Mirzaie F, Mohaddes G. GABAB Receptor Blockade Prevents Antiepileptic Action of Ghrelin in the Rat Hippocampus. Adv Pharm Bull. 2013;3(2):353–8. doi: 10.5681/apb.2013.057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Ghahramanian Golzar M, Babri S, Ataie Z, Ebrahimi H, Mirzaie F, Mohaddes G. NPY Receptors Blockade Prevents Anticonvulsant Action of Ghrelin in the Hippocampus of Rat. Adv Pharm Bull. 2013;3(2):265–71. doi: 10.5681/apb.2013.043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Obay BD, Tasdemir E, Tumer C, Bilgin H, Atmaca M. Dose dependent effects of ghrelin on pentylenetetrazole-induced oxidative stress in a rat seizure model. Peptides. 2008;29(3):448–55. doi: 10.1016/j.peptides.2007.11.020. [DOI] [PubMed] [Google Scholar]
- 38.Samir S, Mostafa A. Role of ghrelin in exhaustive exercise-induced oxidative stress in rat brain and liver. Int J App Exerc Phys. 2013;2(1):25–36. [Google Scholar]
- 39.Çetin E, Çetin N. Protective effect of ghrelin against the oxidative brain and kidney injuries induced by carbon tetrachloride in rats. Atatürk Üniversitesi Vet Bil Derg. 2011;6(3):195–200. [Google Scholar]
- 40.Kheradmand A, Alirezaei M, Asadian P, Rafiei Alavi E, Joorabi S. Antioxidant enzyme activity and MDA level in the rat testis following chronic administration of ghrelin. Andrologia. 2009;41(6):335–40. doi: 10.1111/j.1439-0272.2009.00932.x. [DOI] [PubMed] [Google Scholar]
- 41.Kheradmand A, Alirezaei M, Birjandi M. Ghrelin promotes antioxidant enzyme activity and reduces lipid peroxidation in the rat ovary. Regul Pept. 2010;162(1-3):84–9. doi: 10.1016/j.regpep.2010.02.008. [DOI] [PubMed] [Google Scholar]
- 42.Zwirska-Korczala K, Adamczyk-Sowa M, Sowa P, Pilc K, Suchanek R, Pierzchala K. et al. Role of leptin, ghrelin, angiotensin II and orexins in 3T3 L1 preadipocyte cells proliferation and oxidative metabolism. J Physiol Pharmacol. 2007;58(Suppl 1):53–64. [PubMed] [Google Scholar]
- 43.Nakano M. Determination of superoxide radical and singlet oxygen based on chemiluminescence of luciferin analogs. Methods Enzymol. 1990;186:585–91. doi: 10.1016/0076-6879(90)86154-n. [DOI] [PubMed] [Google Scholar]
- 44.Shohami E, Beit-Yannai E, Horowitz M, Kohen R. Oxidative stress in closed-head injury: brain antioxidant capacity as an indicator of functional outcome. J Cereb Blood Flow Metab. 1997;17(10):1007–19. doi: 10.1097/00004647-199710000-00002. [DOI] [PubMed] [Google Scholar]
- 45.Hayes JD, Milner SW, Walker SW. Expression of glyoxalase, glutathione peroxidase and glutathione S-transferase isoenzymes in different bovine tissues. Biochim Biophys Acta. 1989;994(1):21–9. doi: 10.1016/0167-4838(89)90057-5. [DOI] [PubMed] [Google Scholar]
- 46.Halliwell B. Oxidative stress and neurodegeneration: where are we now? J Neurochem. 2006;97(6):1634–58. doi: 10.1111/j.1471-4159.2006.03907.x. [DOI] [PubMed] [Google Scholar]
- 47.Harrison FE, May JM. Vitamin C function in the brain: vital role of the ascorbate transporter SVCT2. Free Radic Biol Med. 2009;46(6):719–30. doi: 10.1016/j.freeradbiomed.2008.12.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Boldyrev AA, Gallant SC, Sukhich GT. Carnosine, the protective, anti-aging peptide. Biosci Rep. 1999;19(6):581–7. doi: 10.1023/a:1020271013277. [DOI] [PubMed] [Google Scholar]
- 49.Boldyrev AA, Stvolinsky SL, Tyulina OV, Koshelev VB, Hori N, Carpenter DO. Biochemical and physiological evidence that carnosine is an endogenous neuroprotector against free radicals. Cell Mol Neurobiol. 1997;17(2):259–71. doi: 10.1023/A:1026374114314. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Alabovsky VV, Boldyrev AA, Vinokurov AA, Shchavratsky V. Effect of histidine-containing dipeptides on isolated heart under ischemia/reperfusion. Biochemistry (Mosc) 1997;62(1):77–87. [PubMed] [Google Scholar]
- 51.Infanger DW, Sharma RV, Davisson RL. NADPH oxidases of the brain: distribution, regulation, and function. Antioxid Redox Signal. 2006;8(9-10):1583–96. doi: 10.1089/ars.2006.8.1583. [DOI] [PubMed] [Google Scholar]
- 52.Tong XX, Wu D, Wang X, Chen HL, Chen JX, Wang XX. et al. Ghrelin protects against cobalt chloride-induced hypoxic injury in cardiac H9c2 cells by inhibiting oxidative stress and inducing autophagy. Peptides. 2012;38(2):217–27. doi: 10.1016/j.peptides.2012.06.020. [DOI] [PubMed] [Google Scholar]