

Abstract
Background
Serine proteinase inhibitors (serpins) finely regulate serine proteinase activity via a suicide substrate-like inhibitory mechanism. In parasitic nematodes, some serpins interact with host physiological processes; however, little is known about these essential molecules in Anisakis. This article reports the gene sequencing, cloning, expression and preliminary biochemical and bioinformatically-based structural characterization of a new Anisakis serpin (ANISERP).
Methods
The full AniSerp gene was cloned by specific RACE-PCR after screening an Anisakis simplex (L3) cDNA library. For biochemical assays, the AniSerp gene was subcloned into both prokaryotic and eukaryotic vectors, and the recombinant proteins were purified. The inhibitory properties of the proteins were tested in classical biochemical assays using human serine peptidases and AMC substrates. Immunolocalization of ANISERP, theoretical structural analysis and bioinformatically-based structural modelling of the ANISERP protein were also conducted.
Results
The AniSerp gene was found to have 1194 nucleotides, coding for a protein of 397 amino acid residues plus a putative N-terminal signal peptide. It showed significant similarity to other nematode, arthropod and mammalian serpins. The recombinant ANISERP expressed in the prokaryotic and eukaryotic systems inhibited the human serine proteases thrombin, trypsin and cathepsin G in a concentration-dependent manner. No inhibitory activity against Factor Xa, Factor XIa, Factor XIIa, elastase, plasmin or chymotrypsin was observed. ANISERP also acted on the cysteine protease cathepsin L. ANISERP was mainly localized in the nematode pseudocoelomic fluid, somatic muscle cell bodies and intestinal cells. The findings of molecular dynamics studies suggest that ANISERP inhibits thrombin via a suicide substrate-like inhibitory mechanism, similar to the mechanism of action of mammalian coagulation inhibitors. In contrast to findings concerning human antithrombin III, heparin had no effect on ANISERP anticoagulant inhibitory activity.
Conclusions
Our findings suggest that ANISERP is an internal Anisakis regulatory serpin and that the inhibitory activity against thrombin depends on a suicide substrate-like inhibitory mechanism, similar to that described for human antithrombin (AT)-III. The fact that heparin does not modulate the anticoagulant activity of ANISERP might be explained by the absence in the latter of five of the six positively charged residues usually seen at the AT-III-heparin binding site.
Keywords: Serpin, Proteinase, Anisakis, Trypsin, Thrombin, Cathepsin L, Anticoagulant properties, Modelling analysis, Heparin
Background
Anisakis spp. are parasitic nematodes that infect the gastrointestinal tract of sea mammals [1], producing gastric ulcers and haemorrhagic exudates; they can also penetrate the abdominal cavity by crossing the gastrointestinal wall [2]. Humans are accidental hosts that become affected via the ingestion of Anisakis larvae present in raw or undercooked fish and seafood. Human Anisakis infection causes gastrointestinal disease with mild to severe clinical symptoms such as nausea, vomiting, diarrhoea, and abdominal and epigastric pain [3]. Occult blood is often present in the gastric juice and faeces [4]. Gastrointestinal symptoms are often associated with allergic reactions involving specific IgE responses to parasite allergens [5–7].
The serpins belong to the superfamily of serine peptidase inhibitors and are expressed by many organisms ranging from plants to vertebrates. They help to control proteolysis in molecular pathways associated with tissue homeostasis/cell survival, development, and host defence [8]. Some have cross-class activity and can inhibit cysteine proteinases [9, 10]; others may even behave as non-inhibitory chaperones, tumour suppressors or transport molecules [8].
Serpin expression in parasites, especially helminths, is the subject of intense study [11–13]. In nematodes, serpins interact with endogenous parasite proteinases and some are also believed to play important roles in defence against digestion by host proteinases, inhibition of the host immune response, and even as immunomodulators [12, 14].
In Anisakis, only four serine proteinase inhibitors have been characterized to date: Anisakis ASP1, ASP2 and Ani s 6, which belong to a unique class of nematode inhibitors (smapins), and Ani s 1, an allergen belonging to the Kunitz-type family [15–18]. The present article reports the molecular and biochemical characterization of ANISERP, a new Anisakis serpin [GenBank™: FR694897].
Methods
AniSerp gene cloning
An Anisakis simplex truncated cDNA that showed notable similarity to serpin genes (NCBI) was obtained by screening a cDNA library obtained from the L3 stage larvae. Amplification of the 5′ end was performed by RACE-PCR and using a parasite cDNA collection prepared with the Marathon cDNA amplification kit (Clontech) and the forward primer AP1 (5′ CTAATACGACTCACTATAGGGC 3′), which corresponds to the AP1 adaptor sequence of the A.simplex cDNA collection, and reverse primer SR1 (5′ ACCCGCAGTAGTTTTATCCATTTGTTCG 3′), the design of which was based on the truncated cDNA sequence. On the basis of the information obtained, two new primers were used to amplify the full gene by PCR: the forward primer SER5′ (5′ ATGATGACAGCATTACCGTTTTTAAC 3′) and the reverse primer SER3′ (5′ TCAGTGGAAACGACCAATAAACAGAATGCG 3′). The gene was named AniSerp. Standard amplification protocols were used in all cases [19].
Bioinformatic analysis
The theoretical molecular weight and isoelectric point of the full ANISERP protein were obtained using the Expasy Compute pI/Mw tool (http://web.expasy.org/compute_pi/). The protein sequence was also compared with sequences included in the GenBank database, by using the BLASTp program [20], and its motifs were characterised using ScanProsite software (ExPASY Bioinformatics Resource Server; http://www.expasy.org/proteomics). The putative N-terminal signal peptide in the protein was predicted using the SignalP 4.1 Server program [21].
Subcloning, expression in Escherichia coli and purification of recombinant ANISERP protein
The AniSerp gene without the sequence coding for the predicted putative N-terminal signal peptide was subcloned into the pGEX4T2 expression vector (GE Healthcare Life Sciences) and further transformed into the BL21 strain of E. coli (F-, ompT, hsdS [rb-, mb-], gal) (GE Healthcare Life Sciences). Recombinant protein expression was induced by different protocols to obtain a soluble fusion protein similar to the native protein, which was subsequently purified. Expression protocols including different combinations of incubation temperatures (16 and 37 °C), concentrations of isopropyl β-D-1-thiogalactopyranoside (ITPG) (0.01, 0.1, 0.5 and 1 mM) and incubation times (4-16 h) were used. Once the recombinant ANISERP protein was obtained in its soluble form, it was purified by glutathione-Sepharose 4B bead affinity chromatography and eluted with 15 mM glutathione, 100 mMTris-HCl pH 8, 0.1 % Triton X-100 elution buffer. The purified recombinant protein was then dialyzed. The protein concentration was determined using the BCA Protein Assay Kit (Thermo Scientific), with bovine serum albumin (BSA) as a standard [22]. The expressed protein was then subjected to SDS-PAGE analysis and Coomassie staining, plus Western blot analysis with an anti-GST antibody (GE Healthcare Life Sciences).
Subcloning, expression in Sf9 insect cells, and purification of recombinant His-tagged ANISERP protein. Mass spectrometry analysis
The AniSerp gene was amplified using the oligonucleotides AniSerp_Fw 5′ GGGGACAAGTTTGTACAAAAAAGCAGGCTTCATGCAGCAGACAATCGATGATGCCCAAGC 3′ and AniSerp_Rv 5′ GGGGACCACTTTGTACAAGAAAGCTGGGTTCAGTGGAAACGACCAATAAACAGAATGCG 3′ (Sigma Genosys). A recombinant bacmid carrying the AniSerp gene was obtained using the Gateway Cloning System (Life Technologies), following the manufacturer’s instructions. The bacmid DNA (1–2 μg) was transfected into Sf9 insect cells with Cellfectin® II Reagent (Invitrogen), following the manufacturer’s instructions, yielding infectious recombinant baculovirus particles. The transfected Sf9 insect cells were cultured in TC100-Insect Medium (Sigma-Aldrich), containing 10 % FBS (Gibco), to produce large amounts of the recombinant ANISERP protein.
Soluble His-tagged ANISERP protein was then purified by affinity chromatography, with a 1 ml HiTrapTM FF crude column (GE Healthcare) and an ÅKTA FPLC system (GE Healthcare), according to standard procedures. Fractions were analyzed in SDS-PAGE gels stained with Coomassie Brilliant Blue (Bio-Rad). The ANISERP protein from different fractions was pooled, dialyzed against PBS, quantified by the BCA method, and stored at−80 °C until use. Finally, the identity of the recombinant protein was confirmed by MALDI-TOF mass spectrometry, according to standard procedures [19].
Inhibitory assays
The inhibitory activity of ANISERP against 10 proteinases (trypsin, chymotrypsin, plasmin, elastase, FXa, FXIa, FXIIa, thrombin, cathepsin G and cathepsin L) was examined by measuring the residual proteolytic activity on specific proteinase substrates after the incubation of each enzyme with purified recombinant ANISERP. Table 1 includes the reaction conditions for each proteinase tested. In addition, and taking into account the report by Morris and Sakanari [15], ANISERP inhibition of serine proteinase activity in A. simplex crude extract was determined using Z-Gly-Pro-Arg-AMC in 50 mMTris/HCl pH 7.5 plus 20 mMNaCl as a substrate. All substrates were purchased from Sigma Aldrich. Assays were performed after incubation of ANISERP with each enzyme for 10 min at 37 °C in the appropriate reaction buffer. In all cases, the enzyme concentrations used in the assays were linearly related to the reaction time (30 min). The inhibition controls used were AEBSF (1 mM) (serine-proteinases inhibitor) and E-64 (10 μM; a cysteine proteinase inhibitor), both obtained from Sigma-Aldrich. After addition of the substrate (final concentration 250 μM) to the reaction mixture, the residual enzyme activity was measured by continuous monitoring for AMC substrates at excitation and emission wavelengths of 380 and 460 nm respectively, in a Victor 3 1420 Perkin Elmer Fluorescence microplate reader (Perkin Elmer España S.L.). Reactions with the pNA substrate were monitored, at 405 nm, in an ELX 800TM Bioteck Absorbance microplate reader (Bioteck). Purified GST was also used as a negative control for proteinase inhibition when ANISERP was produced in pGEX4T2/E. coli.
Table 1.
Enzymes, substrates and reaction conditions for each proteinase tested
| Enzyme | [Enzyme] | [Substrate] 250 μM | Activity buffer |
|---|---|---|---|
| Thrombin | 74 nM | Boc-Val-Arg -AMC | 50 mM Tris–HCl pH 8, 100 mM NaCl |
| Trypsin | 54 nM | Boc-Gln-Ala-Arg-AMC | 50 mM Tris–HCl pH 8, 1 mM CaCl2, 0.15 M NaCl |
| Cathepsin G | 0.33 nM | N-Succinyl-Ala-Ala-Phe-AMC. | 50 mM HEPES/NaOH pH 7.5 |
| Cathepsin L | 0.83 nM | Z-Phe-Arg-AMC | Sodium acetate 100 mM pH 5.5, 1 mM EDTA, 4 mM DTT, 0.001% BSA |
| Plasmine | 333 nM | Nt-Boc-Val-Leu-Lys-AMC | 50 mM Tris–HCl pH 8, 50 mM NaCl |
| Elastase | 1.2 nM | N-Methoxysuccinyl-Ala-Ala-Pro-Val-7 amido AMC | 25 mM Tris–HCl pH 8, 100 mM NaCl, 1 mM CaCl2 |
| Chymotrypsin | 0.012 nM | N-succinyl-Lelu-Leu-Val-Tyr-7 amido AMC | 100 mM HEPES/ NaOH pH 7.5 |
| Factor Xa | 0.054 nM | Boc-Ile-Glu-Gly-Arg-7 amido AMC | 50 mM Tris–HCl pH 8.3, 5 mM CaCl2, 0.2 mM NaCl |
| Factor XIa | 14,000 nM | Boc-Phe-Ser-Arg-7 amido AMC | 50 mM Tris–HCl pH 8, 100 mM NaCl, 1 mM CaCl2 |
| Factor XIIa | 0.0017 nM | Boc-Val-Arg-AMC | 4 mM Sodium Acetate-HCl/0.15 M NaCl/pH 5.3 |
Anti-ANISERP hyperimmune sera
A female New Zealand rabbit was immunized, via subcutaneous injection, with ANISERP recombinant protein fused to GST (glutathione transferase), by using the following protocol: 75 μg/ ml of ANISERP recombinant protein was equally emulsified with Freund Adjuvant and administered in three doses, a single dose with Freund Complete Adjuvant (ACF) and the remaining two with Freund Incomplete Adjuvant (FIA) 20 and 40 days after the first injection. Blood samples were collected prior to the first injection and two months after the last injection. Serum samples were collected after centrifugation of clotted blood and stored at−20 °C until use.
Ethical approval
The rabbit was maintained and immunized in accordance with institutional and national guidelines. The protocol was approved by the Ethics Committee for Research and Animal Welfare (CEIyBA) of the ISCIII (CBA N# 09_2014_v2).
Immunohistochemical localization of ANISERP
Live third-stage larvae of A. simplex were collected from the body cavity of blue whiting (Micromesistiuspoutassou) purchased at a local market, washed several times in physiological saline, cut into two portions (of which the anterior one-third includes the ventricle and part of the intestine) and fixed in 10 % buffered formalin for 12 h. After fixation, the samples were washed with PBS, dehydrated, embedded in paraffin and cut into 5 μm-thick sections, as previously described [23]. Once deparaffinized and hydrated, the slides were blocked with PBS containing 0.05 % Tween 20 and 1 % dry skimmed milk (PBS-T-SM) for 2 h at room temperature (RT). As ANISERP was expressed as a GST-fusion protein, and to inhibit any potential cross-reactivity with GST from Anisakis, rabbit hyperimmune serum raised against recombinant ANISERP and preimmune serum (as negative control) were diluted 1/100 and preincubated with or without GST, at 50 μg/ml, in PBS-T-SM for 1 h at RT. The samples were then placed on the slides and incubated for 2 h at RT. The slides were washed three times with PBS-T (for 5 min each time) and then incubated with 0.3 % hydrogen peroxide in PBS for 30 min RT to quench endogenous peroxidase activity. The sections were washed and incubated with peroxidase-conjugated goat anti-rabbit IgG (Bio-Rad) diluted 1/200 in PBS-T-SM for 1 h at RT. The slides were washed again, and bound antibodies were revealed with 0.5 mg/ml 4-chloro-1-naphthol (Sigma-Aldrich) in TBS with 0.005 % hydrogen peroxide. Finally, sections were washed with TBS and mounted with glass coverslips in PBS-glycerol (1:1) for examination and photography. The slides were then stained with Wheatley’s trichrome, as described elsewhere [24].
Effect of heparin on the thrombin inhibition assay
Assays were performed with 200 μl of 50 mMTris/HCl, 100 mMNaCl, pH 8. Inhibition reaction mixtures contained from 0.1 μg/ml to 100 μg/ml (0.1, 5, 15, 25, 45, 70 and 100 μg/ml) heparin and 2.7 nM ANISERP. The reaction was initiated by adding 74 nM thrombin. After incubation of the plates for 5 min, 50 μl of a solution containing 250 μM Boc-Val-Arg-AMC and 0.33 mg/ml Polybrene (Sigma-Aldrich) was added. Residual thrombin activity was determined by measuring the hydrolysis of the fluorescent substrate (Boc-Val-Pro-Arg-AMC) in a Victor 3 1420 Perkin Elmer fluorescent microplate reader. As a positive control, the same experiment was performed with antithrombin (AT) (0.1 mg/ml) instead of ANISERP.
Modelling procedures
Structural three-dimensional models for ANISERP were constructed by homology modelling procedures based on Protein Data Bank structures (selected on the basis of strong sequence similarity, coverage and sequence-to-structure compatibility), using both BLAST [20] and threading (Phyre server [25]) procedures. The template selected was 1JMO (the structure of the human heparin cofactor II [HCII]-thrombin complex) [26]. The model was produced using the SWISS-MODEL server facilities [27–29] at http://swissmodel.expasy.org/SWISS-MODEL.html. The structural quality of the models was checked using the analytical programs provided by the same server (Anolea/Gromos/Verify3D). For geometric optimization, the models were energy minimized using the DeepView GROMOS 43B1 force field routine [30], by applying 500 steps of steepest descent minimization followed by 500 steps of conjugate-gradient minimization. The structure of human thrombin, included in PDB file 1JMO, was also re-modelled to include a serine residue at catalytic position 205, which is an alanine in the crystallized protein. Finally, in order to produce a high quality model for the proposed contact between human thrombin and ANISERP, the structure resulting from homology modelling was subjected to a 2 ns standard molecular dynamics simulation using the PMEMD module of the AMBER9 software package [31] and the parm99 parameter set for the same distribution.
To compare the 3D positions of putative heparin binding sites, structural alignment between the ANISERP model (after molecular dynamics procedures) and the Protein Data Bank structures of human HCII (1JMO) [26] and human AT-III (1 ATH_A) [32]) was performed using the Dali program [33]. Structure plots were generated using the PyMOL program (DeLano Scientific).
Results and discussion
The truncated AniSerp gene was cloned during the screening of an A. simplex L3 cDNA library. The fragment was 1005 nt long (334 aa) and lacked the 5′ end. It showed notable similarity to serine proteinase inhibitors (GenBank). Amplification of the 5′ end was then performed by RACE-PCR with new primers based on the truncated gene, and the full sequence was isolated in the same way. The novel sequence contained 1194 nucleotides and expressed a deduced amino acid backbone of 397 residues (with a predicted molecular weight of 44.6 kDa) plus a putative N-terminal signal peptide of 25 residues. Bioinformatic analysis showed that the gene included a serpin signature (374–384 residues: FIADHPFIFTI) and a potential reactive centre loop (RCL) (p17 [E]-p16 [E/K/R]-p15 [G]-p14 [T/S]-p13 [X]-p12-9 [A/G/S]-p8-1 [X]-p1′-4′). Database comparisons showed similarities between ANISERP and different serine proteinase inhibitors of nematodes and other organisms. The highest similarity scores were obtained for the serpins described in Toxocaracanis (putative serpin-like protein [2e-141, 59 %; KHN 72249.1]), Ascarissuum (putative serpin-like protein [1e-103, 45 %; ERG 78895.1] and serpin b6 [3e-102, 46 %; ERG 82472.1]). ANISERP also showed similarity to the Bostaurus serpin peptidase inhibitor (4e-69, 38 %, NP001193642.1), mouse (Mus musculus) serpin 3b (3e-68, 37 %, NP941373.1) and mouse squamous cell carcinoma antigen 2-related protein 1 (2e-67, 36 %, AAN62872.1). These findings were used together to design ANISERP functional characterization experiments.
To obtain the recombinant ANISERP protein, the gene without the putative N-terminal signal peptide was subcloned into pGEX, and the full sequence was subcloned into the baculovirus vector system. These genes were expressed in prokaryotic and eukaryotic cells respectively, and they were then purified. To collect the proteins in functional form, different expression conditions were tested. For the E. coli system, the conditions were 0.01 mM IPTG and incubation at 16 °C for 16 h. For the baculovirus system, the conditions were a multiplicity of infection of 0.1 (virus titre 5×106pfu/ml), incubation at 27 °C and collection at 72 h.
The effect of the recombinant ANISERP on proteolytic activity was the same in both prokaryotic and eukaryotic systems. ANISERP has two potential N-glycosylation sites, 87-NDSA and 327-NDSL, which do not appear to influence its inhibitory effect. ANISERP inhibited thrombin (IC50 152 nM), trypsin (IC50 173 nM), cathepsin G (IC50 542 nM) and cathepsin L (IC50 217 nM) in a concentration-dependent manner (Fig. 1). No other proteinase, including the parasite peptidase, was affected. This suggests that the Anisakis peptidase described by Morris & Sakanari [15] is not the biological target of ANISERP. Complete inhibition of cysteine and serine proteinases was observed when E-64 (10 μM) and AEBSF (1 mM) were used as positive controls (data not shown). The inhibition of cathepsin L activity confirmed the cross-class nature of ANISERP.
Fig. 1.
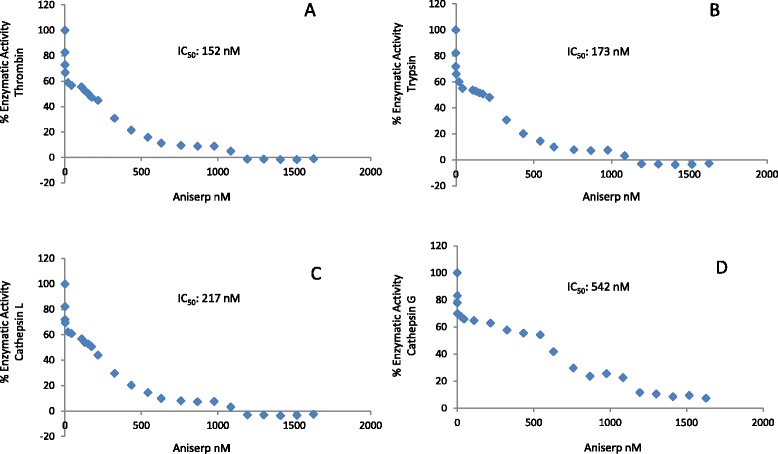
Effect of ANISERP on the proteolytic activity of human thrombin, trypsin, cathepsin L and cathepsin G. Inhibitory effect of different doses of recombinant ANISERP, expressed in a baculovirus system, on the enzymatic activity of a Thrombin (74 nM) on Boc-Val-Arg-AMC, b Trypsin (54 nM) on Boc-Gln-Ala-Arg-AMC, c Cathepsin L (0.83 nM) on Z-Phe-Arg-AMC, and d Cathepsin G (0.33 nM) on N-succinyl-Ala-Ala-Phe-AMC
To locate the possible site of synthesis and final destination of ANISERP, an immunohistochemical study was performed on A. simplex L3 tissues by using a hyperimmune serum raised against recombinant ANISERP-GST. Pre-incubation of the serum with GST did not inhibit the intensity of staining by immunohistochemistry or the signal obtained by indirect ELISA against A. simplex crude extracts (data not shown), indicating that recognition was specific to ANISERP. As shown in Fig. 2a and c, ANISERP was highly concentrated in the pseudocoelomic fluid (haemolymph) and in non-contractile cell bodies (myocytons) of the somatic musculature, and it was absent in the contractile portion of these cells (see Fig. 2d for better identification of the structures). Positive staining was also found to a lesser extent in cells belonging to subventral muscular sectors of the oesophagus and intestine, and weak staining was observed in the lateral hypodermal cords and hypodermis. In addition to the contractile portion of somatic muscle cells, other structures that did not stain included the cuticle, the dorsal oesophageal gland, the excretory cell (including the excretory canal), the intestinal lumen and the ventricle (Fig. 2a and c, and Table 2).
Fig. 2.
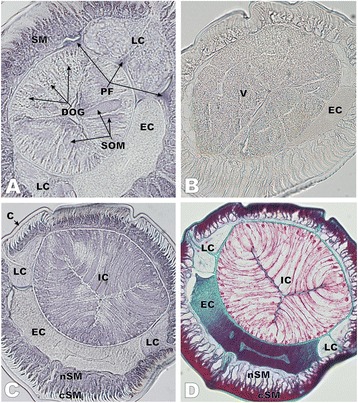
Immunolocalization of ANISERP in sections of Anisakis simplex L3 with a rabbit hyperimmune serum. a Cross section at the level of the oesophagus showing strongly positive staining in SM and PF, moderate staining in SOM and slight staining of LC. No staining was observed in the EC. b The negative control incubated with the preimmune serum did not stain. c Cross section through the intestinal level showing in detail strongly positive immunostaining in the SM, which is restricted to the nSM; no staining was observed in the cSM, the C or EC, including the ca. Less intense staining was also observed in IC. d Section C was decolorized to remove the 4CN stain and subsequently stained with Wheatley’s trichrome. DOG, dorsal oesophageal gland; SOM, subventral muscular sectors of oesophagus; SM, somatic musculature; LC, lateral hypodermal cords; PF, pseudocoelomic fluid; EC, excretory cell; V, ventricle; IC, intestinal cells; nSM, non-contractile portion of somatic muscle cells (myocytons); cSM, contractile portion of somatic muscle cells; C, cuticle; ca, excretory canal; n, nucleus
Table 2.
Summary of the immunohistochemical staining results indicating the tissue distribution of ANISERP
| Structures | Staininga | Structures | Staininga |
|---|---|---|---|
| Dorsal oesophageal gland | - | Excretory cell | - |
| Subventral muscular sectors of oesophagus | ++ | Excretory canal | - |
| Ventricle | - | Hypodermis | ± |
| Intestine | ++ | Lateral hypodermal cords | ± |
| Intestinal lumen | - | Cuticle | - |
| Pseudocoelom fluid | +++ | Somatic muscle cell bodies | +++ |
a-,no staining; ±, weak staining; +, ++ and +++, increasing intensity of positive staining
When marine mammals or humans are infected by L3 Anisakis larvae, three sources of parasite antigens can be delivered to host tissues while larvae remain alive: i) cuticle antigens, which are in direct contact with host tissues, ii) gut antigens, which can be released through the mouth/anus, and iii) secretory antigens, which are released by the excretory cell through the excretory pore, located at the base of subventral lips; the latter constitutes the main source of known Anisakis allergens. Somatic antigens can also come into contact with host tissues, but only in cases in which larvae die and remain in tissues (typically in chronic infections). The absence of staining in the dorsal oesophageal gland, intestinal lumen, excretory cell and cuticle (Table 2) clearly indicates that ANISERP is not released to the external medium and, consequently, its inhibitory action is probably directed against endogenous proteases. This explains why the recombinant ANISERP was not recognized by sera from patients infected with Anisakis during the acute phase (data not shown).
As the strongest staining was observed in myocytons and pseudocoelomic fluid, we can hypothesize that somatic muscle cells are the major source of ANISERP, which can be then released to the haemolymph. Although the putative secretory functions of somatic muscle cells in Anisakis have not been reported before, this type of activity has been reported in muscular cells of Caenorhabditis elegans transfected with a plasmid encoding a signal peptide fused to green fluorescent protein [34]. The presence of a putative signal peptide in ANISERP is consistent with this hypothesis.
In addition to myocytons and pseudocoelomic fluid, staining for ANISERP was also intense in intestinal cells. As intestinal and muscular cells originate from different germinal layers (i.e. endoderm and mesoderm respectively), we can also hypothesize that more than one Anisakis serpin is recognized by the polyclonal antibody. Although this possibility merits further investigation, the fact that other similar serpins exist in the Anisakis transcriptome (unpublished results) is consistent with the hypothesis.
Finally, an in silico analysis of the ANISERP molecule was performed to predict its tertiary structure and help to determine how it might interact with the serin proteases that it inhibits, based on characteristic and conserved serpin folding. Thrombin protease was chosen as a model as it yielded the lowest IC50. To obtain a reliable representation of the actual interactions between proteins, a homology-based 3D model of ANISERP bound to thrombin was generated. To obtain an even more realistic molecular model for the Anisakis serpin, the Ala205 located at the catalytic centre of human thrombin was substituted by a Ser residue (model 1JMO). On the basis of the initial interaction model, short-step (2 ns) molecular dynamics were applied to the whole structure to help the amino acid side chains and the reactive centre loop (RCL) backbone achieve a stable, low-energy conformation that would allow analysis of specific contacts between the two proteins (Fig. 3a). The ANISERP model showed all the characteristics described for classic mammalian serpins, i.e. mixed parallel and antiparallel six-stranded β-sheets with the central strand containing several residues from the RCL. This observation is consistent with that reported by Zang & Maizels [35], i.e. although overall nematode serpins show less primary sequence similarity to their mammalian homologues, they can acquire a common, highly ordered tertiary structure. Figure 3b shows the thrombin substrate groove involving ANISERP amino acids from Met358 to Phe374. Contacts between residues of the two proteins in this region involve hydrophobic clusters and the junctions at the active centre, including contact between the thrombin catalytic triad His43, Asp99 and Ser205 and the ANISERP Arg361 (P1) and Ser362 (P1′) residues (Fig. 3c).
Fig. 3.
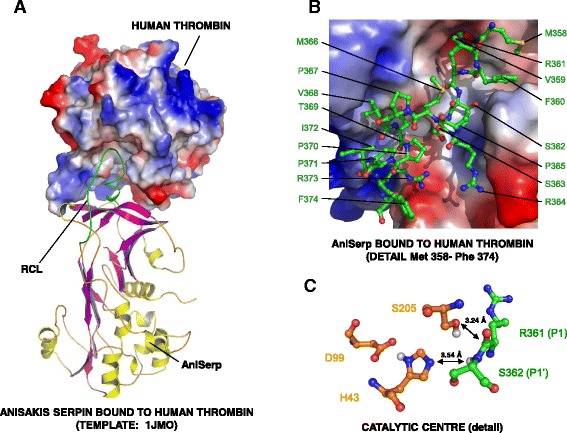
Model for ANISERP-human thrombin binding. a General view of the model for ANISERP binding to human thrombin. The location of the ANISERP reactive centre loop (RCL [green]) in the structural groove of the thrombin is indicated. The thrombin surface is coloured according to its theoretical electrostatic properties (blue = positive, red = negative). b Residues in the heavy chain of human thrombin; binding of amino acids Met358 and Phe374 in ANISERP. c Position of residues in the catalytic triad of human thrombin (His43, Asp99 and Ser205) and their positions relative to ANISERP residues Arg361 and Ser362 (P1 and P1′ respectively). Distances between active residues and atoms in the peptide bond of the substrate, compatible with proteinase activity, are indicated
In the human coagulation cascade, AT III inhibits the serine proteinases trypsin and plasmin along with the coagulation factors thrombin and FXa. The effect of ANISERP on FXa was therefore tested. No inhibition was observed, as indicated above (data not shown). In contrast, HCII inhibits the serine proteases thrombin, chymotrypsin and cathepsin G. The distinct enzyme specificities of ATand HCII appear to be the result of amino acid changes in P1 and the immediately adjacent residues of the RCL-enzyme recognition sequence. AT, which has an Arg at P1, is an inhibitor of the Arg-proteinases thrombin and FXa, while HCII is a specific thrombin inhibitor with an unfavourable Leu residue at P1 [36]. The inhibitory activity of ANISERP, which selectively blocks human thrombin without affecting FXa, may be explained by considering that it has an Arg at P1, as does AT. In addition, the 14 amino acid changes in the adjacent residues of the RCL may be responsible for the different specificities of ANISERP, including the lack of FXa activity. Nevertheless, experiments with ANISERP variants constructed by site-directed mutagenesis should be conducted to assess whether these changes in the RCL are involved in the specific reaction between ANISERP and thrombin.
As there is some evidence [37, 38] that the activity of the AT and HCII inhibitors is greatly potentiated by heparin, the effect of heparin on ANISERP inhibitory activity on thrombin was also studied. Heparin (0.1 to 100 μg/ml) did not affect the ANISERP inhibitory activity, although it did increase the inhibitory activity of the AT control in a concentration-dependent manner (Fig. 4).
Fig. 4.
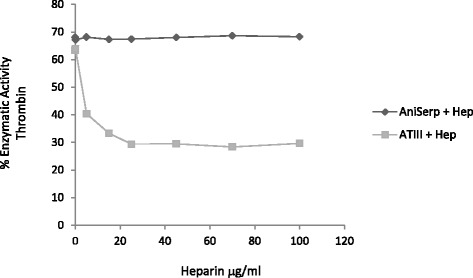
Effect of heparin on ANISERP inhibition of human thrombin proteolytic activity. Residual activity of thrombin after incubation with recombinant ANISERP (circles) or AT (squares) with increasing concentrations of heparin (0.1-100 μg/ ml). ANISERP was used at a fixed concentration of 2.17 nM (IC50: 152 nM)
In an attempt to understand the lack of effect of heparin on the ANISERP anticoagulant action, the hypothetical heparin binding site residues were aligned with the AT and HCII heparin binding site residues described elsewhere [26]. Figure 5 shows the sequence and structural alignments of the three proteins. Five residues (Lys173, Arg184, Lys185, Arg189, and Arg192 blue spheres in Fig. 5) out of six critical for HCII/heparin binding appear to be absent/altered in ANISERP.
Fig. 5.
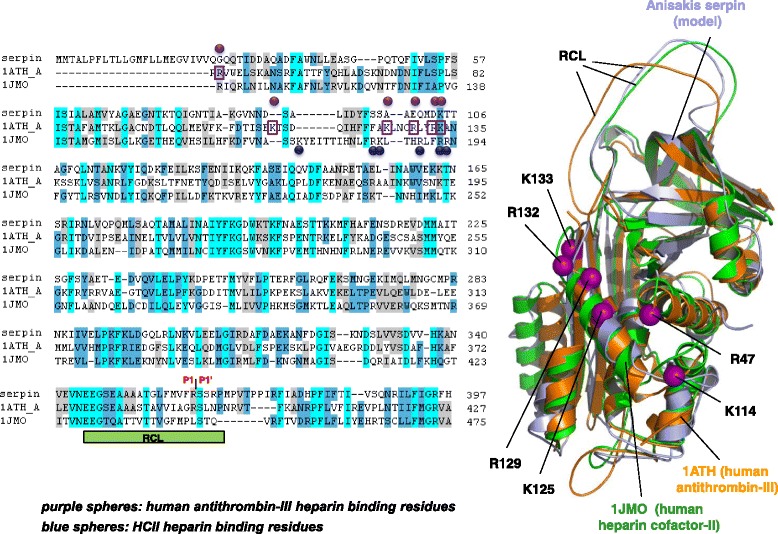
Sequence and structural alignment of ANISERP, human antithrombin III (1 ATH_A), and human heparin cofactor-II (1JMO). The key residues involved in heparin binding described for human antithrombin III (Arg 47, Lys114, Lys125, Arg 129, Arg132 and Lys133) are shown as purple boxes and spheres. The key residues involved in heparin binding described for heparin cofactor-II (Lys173, Arg184, Lys185, Arg189, Arg192 and Arg193) are shown as blue spheres. The positions of the P1-P1’residues in the ANISERP sequence are indicated. The residues in the 1JMO and 1ATH_A sequences are numbered according to Baglin et al. [26]
Conclusions
ANISERP is an internally-secreted Anisakis serpin that displays in vitro inhibitory activity. The inhibitory action of ANISERP against thrombin appears to depend on a suicide substrate-like inhibitory mechanism, similar to that displayed by human AT-III. The fact that heparin does not modulate the anticoagulant activity of ANISERP might be explained by the absence in its structure of five of the six positively charged residues usually seen in the AT-III-heparin binding site. The findings of the immunolocalization study indicate that ANISERP exerts its function as an internal regulatory serpin and rule out a role at the parasite-host interface.
Acknowledgements
We thank Biomol-Informatics SL (http://www.biomol-informatics.com/) for bioinformatic consultation. We also thank Juan Francisco Alcaide and Julia Medrano for their invaluable help in the ANISERP patenting process. We thank Dr Raúl Iglesias (Laboratorio de Parasitología, Facultad de Biología, Universidad de Vigo) for his helpful comments on the interpretation of the IHQ studies.
Financial disclosure
This work was funded by the Spanish Ministry of Science and Innovation (SAF2002-04057-CO2-01) and the Instituto de Salud Carlos III (ISCIII) as part of project MPY 1404/09, and ISCIII-AESI project MPY 1279/15. Carolina Hurtado and Pamela Campioli were supported by grants from the ISCIII (Fondo de InvestigacionesSanitarias), through VI NP of I + D + I (2008–2011), ISCIII- General Sub-Direction of Networks and Centers for Collaborative Research (RETIC-RICET, RD06/0021/0019, RD12/0018/011). Victoria Martínez Sernández holds a predoctoral fellowship from the Spanish Ministerio de Educación, Cultura y Deporte (Programa de Formación del Profesorado Universitario). The funders did not have any role in the design of the study, data collection, analysis, decision to publish, or preparation of the manuscript.
Footnotes
Elizabeth Valdivieso and Maria J. Perteguer contributed equally to this work.
Competing interests
The authors have no interests that conflict with the publication of this work.
Authors’ contributions
Conception and design of experiments: EV, MJP, CH, ER, AS, VMS, PGP, FMU, TG. Experimental work: EV, MJP, CH, PC, ER, VMS, PGP. Data analysis: EV, MJP, CH, AS, VMS, PGP, FMU, TG. Contribution of reagents/materials/analysis tools: EV, MJP, ER, PGP, FMU, TG. Manuscript preparation: EV, MJP, CH, VMS, PGP, FMU, TG. All authors read and approved the final version of the manuscript.
Contributor Information
Elizabeth Valdivieso, Email: elizabeth.valdivieso@ciens.ucv.ve.
Maria J. Perteguer, Phone: (+34) 91 822 3670, Email: chus.perteguer@isciii.es
Carolina Hurtado, Email: carolina.hurtadomarcos@ceu.es.
Pamela Campioli, Email: plcampioli@isciii.es.
Esperanza Rodríguez, Email: erodrgez@isciii.es.
Ana Saborido, Email: asaborido@quim.ucm.es.
Victoria Martínez-Sernández, Email: victoria.sernandez@gmail.com.
Paulino Gómez-Puertas, Email: pagomez@cbm.csic.es.
Florencio M. Ubeira, Email: fm.ubeira@usc.es
Teresa Gárate, Email: tgarate@isciii.es.
References
- 1.Audícana MT, del Pozo MD, Iglesias R, Ubeira FM. Anisakis simplex and Pseudoterranova decipiens. In: Miliotis M, Bier J, editors. International handbook on foodborne pathogens. New York: Marcell Dekker; 2003. pp. 613–36. [Google Scholar]
- 2.Geraci JR, St Aubin DJ. Effects of parasites on marine mammals. Int J Parasitol. 1987;17:407–14. doi: 10.1016/0020-7519(87)90116-0. [DOI] [PubMed] [Google Scholar]
- 3.Ishikura H, Kikuchi K, Nagasawa K, Ooiwa T, Takamiya H, Sato N, et al. Anisakidae and anisakidosis. Prog Clin Parasitol. 1993;3:43–102. doi: 10.1007/978-1-4612-2732-8_3. [DOI] [PubMed] [Google Scholar]
- 4.Bier JW. Anisakiasis. Laboratory diagnosis of infectious diseases. In: Balows A, Hausler WJ Jr, Ohashi M, Turano A, editors. Bacterial, mycotic and parasitic diseases. New York: Springer; 1988. pp. 768–74. [Google Scholar]
- 5.Valiñas B, Lorenzo S, Eiras A, Figueiras A, Sanmartín ML, Ubeira FM. Prevalence of and risk factors for IgE sensitization to Anisakis simplex in a Spanish population. Allergy. 2001;56:667–71. doi: 10.1034/j.1398-9995.2001.00987.x. [DOI] [PubMed] [Google Scholar]
- 6.Audicana MT, Kennedy MW. Anisakis simplex: from obscure infectious worm to inducer of immune hypersensitivity. Clin Microbiol Rev. 2008;21:360–79. doi: 10.1128/CMR.00012-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Anadón AM, Romarís F, Escalante M, Rodríguez E, Gárate T, Cuéllar C, et al. The Anisakis simplex Ani s 7 major allergen as an indicator of true Anisakis infections. Clin Exp Immunol. 2009;156:471–8. doi: 10.1111/j.1365-2249.2009.03919.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Silverman GA, Whisstock JC, Bottomley SP, Huntington JA, Kaiserman D, Luke CJ, et al. Serpins flex their muscle: I. Putting the clamps on proteolysis in diverse biological systems. J Biol Chem. 2010;285:24299–305. doi: 10.1074/jbc.R110.112771. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Sakata Y, Arima K, Takai T, Sakurai W, Masumoto K, Yuyama N, et al. The squamous cell carcinoma antigen 2 inhibits the cysteine proteinase activity of a major mite allergen, Der p 1. J Biol Chem. 2004;279:5081–7. doi: 10.1074/jbc.M311585200. [DOI] [PubMed] [Google Scholar]
- 10.Law RH, Zhang Q, McGowan S, Buckle AM, Silverman GA, Wong W, et al. An overview of the serpin superfamily. Genome Biol. 2006;7:216. doi: 10.1186/gb-2006-7-5-216. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Yi D, Xu L, Yan R, Li X. Haemonchuscontortus: cloning and characterization of serpin. Exp Parasitol. 2010;125:363–70. doi: 10.1016/j.exppara.2010.03.002. [DOI] [PubMed] [Google Scholar]
- 12.Molehin AJ, Gobert GN, Driguez P, McManus DP. Characterisation of a secretory serine protease inhibitor (SjB6) from Schistosomajaponicum. Parasit Vectors. 2014;14:7–330. doi: 10.1186/1756-3305-7-330. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Molehin AJ, Gobert GN, McManus DP. Serine protease inhibitors of parasitic helminths. Parasitology. 2012;139:681–95. doi: 10.1017/S0031182011002435. [DOI] [PubMed] [Google Scholar]
- 14.Zang X, Yazdanbakhsh M, Jiang H, Kanost MR, Maizels RM. A novel serpin expressed by blood-borne microfilariae of the parasitic nematode Brugiamalayi inhibits human neutrophil serine proteinases. Blood. 1999;94:1418–28. [PubMed] [Google Scholar]
- 15.Morris SR, Sakanari JA. Characterization of the serine protease and serine protease inhibitor from the tissue-penetrating nematode Anisakis simplex. J Biol Chem. 1994;269:27650–6. [PubMed] [Google Scholar]
- 16.Nguyen TT, Qasim MA, Morris S, Lu CC, Hill D, Laskowski M, Jr, et al. Expression and characterization of elastase inhibitors from the ascarid nematodes Anisakis simplex and Ascaris suum. Mol Biochem Parasitol. 1999;30(102):79–89. doi: 10.1016/S0166-6851(99)00088-2. [DOI] [PubMed] [Google Scholar]
- 17.Kobayashi Y, Ishizaki S, Shimakura K, Nagashima Y, Shiomi K. Molecular cloning and expression of two new allergens from Anisakis simplex. Parasitol Res. 2007;100:1233–41. doi: 10.1007/s00436-006-0396-4. [DOI] [PubMed] [Google Scholar]
- 18.Kobayashi Y, Ishizaki S, Nagashima Y, Shiomi K. Ani s 1, the major allergen of Anisakis simplex: purification by affinity chromatography and functional expression in Escherichia coli. Parasitol Int. 2008;57:314–9. doi: 10.1016/j.parint.2008.01.005. [DOI] [PubMed] [Google Scholar]
- 19.Perteguer MJ, Gómez-Puertas P, Cañavate C, Dagger F, Gárate T, Valdivieso E. Ddi1-like protein from Leishmania major is an active aspartyl proteinase. Cell Stress Chaperones. 2013;18:171–81. doi: 10.1007/s12192-012-0368-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Altschul SF, Madden TL, Schäffer AA, Zhang J, Zhang Z, Miller W, et al. Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res. 1997;25:3389–402. doi: 10.1093/nar/25.17.3389. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Petersen TN, Brunak S, von Heijne G, Nielsen H. Locating proteins in the cell using TargetP, SignalP, and related tools. Nat Methods. 2011;8:785–6. doi: 10.1038/nmeth.1701. [DOI] [PubMed] [Google Scholar]
- 22.Smith PK, Krohn RI, Hermanson GT, Mallia AK, Gartner FH, Provenzano MD, et al. Measurement of protein using bicinchoninic acid. Anal Biochem. 1985;150:76–85. doi: 10.1016/0003-2697(85)90442-7. [DOI] [PubMed] [Google Scholar]
- 23.Martínez-Sernández V, Mezo M, González-Warleta M, Perteguer MJ, Muiño L, Guitián E, et al. The MF6p/FhHDM-1 major antigen secreted by the trematode parasite Fasciola hepatica is a heme-binding protein. J Biol Chem. 2014;289:1441–56. doi: 10.1074/jbc.M113.499517. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Sanmartín ML, Iglesias R, Santamarina MT, Leiro J, Ubeira FM. Anatomical location of phosphorylcholine and other antigens on encysted Trichinella using immunohistochemistry followed by Wheatley’s trichrome stain. Parasitol Res. 1991;77:301–6. doi: 10.1007/BF00930905. [DOI] [PubMed] [Google Scholar]
- 25.Bennett-Lovsey RM, Herbert AD, Sternberg MJ, Kelley LA. Exploring the extremes of sequence/structure space with ensemble fold recognition in the program Phyre. Proteins. 2008;70:611–25. doi: 10.1002/prot.21688. [DOI] [PubMed] [Google Scholar]
- 26.Baglin TP, Carrell RW, Church FC, Esmon CT, Huntington JA. Crystal structures of native and thrombin-complexed heparin cofactor II reveal a multistep allosteric mechanism. Proc Natl Acad Sci USA. 2002;99:11079–84. doi: 10.1073/pnas.162232399. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Guex N, Diemand A, Peitsch MC. Protein modelling for all. Trends Biochem Sci. 1999;24:364–7. doi: 10.1016/S0968-0004(99)01427-9. [DOI] [PubMed] [Google Scholar]
- 28.Peitsch MC. ProMod and Swiss-Model: Internet-based tools for automated comparative protein modelling. Biochem Soc Trans. 1996;24:274–9. doi: 10.1042/bst0240274. [DOI] [PubMed] [Google Scholar]
- 29.Schwede T, Kopp J, Guex N, Peitsch MC. SWISS-MODEL: An automated protein homology-modeling server. Nucleic Acids Res. 2003;31:3381–5. doi: 10.1093/nar/gkg520. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Guex N, Peitsch MC. SWISS-MODEL and the Swiss-PdbViewer: an environment for comparative protein modeling. Electrophoresis. 1997;18:2714–23. doi: 10.1002/elps.1150181505. [DOI] [PubMed] [Google Scholar]
- 31.Case DA, Cheatham TE, 3rd, Darden T, Gohlke H, Luo R, Merz KM, Jr, et al. The Amber biomolecular simulation programs. J Comput Chem. 2005;26:1668–71. doi: 10.1002/jcc.20290. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Schreuder HA, de Boer B, Dijkema R, Mulders J, Theunissen HJ, Grootenhuis PD, et al. The intact and cleaved human antithrombin III complex as a model for serpin-proteinase interactions. Nat Struct Biol. 1994;1:48–54. doi: 10.1038/nsb0194-48. [DOI] [PubMed] [Google Scholar]
- 33.Holm L, Kaariainen S, Rosenstrom P, Schenkel A. Searching protein structure databases with DaliLite v. 3. Bioinformatics. 2008;24:2780–1. doi: 10.1093/bioinformatics/btn507. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Fares H, Greenwald I. Genetic analysis of endocytosis in Caenorhabditis elegans: coelomocyte uptake defective mutants. Genetics. 2001;159:133–45. doi: 10.1093/genetics/159.1.133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Zang X, Maizels RM. Serine proteinase inhibitors from nematodes and the arms race between host and pathogen. Trends Biochem Sci. 2001;26:191–7. doi: 10.1016/S0968-0004(00)01761-8. [DOI] [PubMed] [Google Scholar]
- 36.Gettins PG, Olson ST. Exosite determinants of serpin specificity. J Biol Chem. 2009;284:20441–5. doi: 10.1074/jbc.R800064200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Dementiev A, Petitou M, Herbert JM, Gettins PG. The ternary complex of antithrombin-anhydrothrombin-heparin reveals the basis of inhibitor specificity. Nat Struct Mol Biol. 2004;11:863–7. doi: 10.1038/nsmb810. [DOI] [PubMed] [Google Scholar]
- 38.Rossi EB, Duboscq CL, Kordich LC. Heparin cofactor II, a thrombin inhibitor with a still not clarified physiologic role. Medicina (B Aires) 1999;59:95–104. [PubMed] [Google Scholar]