

Abstract
Ixodes spp. ticks are known to occasionally harbour spiroplasmas – helical mycoplasmas in the class Mollicutes; a previous study in Slovakia reported an overall prevalence of Spiroplasma ixodetis of 3% in Ixodes ricinus. In the present study, extracts of unfed adult I. ricinus ticks collected from vegetation in south-western Slovakia were added to a panel of cell lines derived from I. ricinus and Ixodes scapularis embryos. The cultures were monitored by preparation and examination of Giemsa-stained cytocentrifuge smears at intervals over the subsequent 16–18 months. Spiroplasma-like microorganisms were detected in cultures of both tick species after 2–3 months and subcultured onto fresh, uninfected cells of the appropriate cell line up to seven times. Molecular analysis using PCR assays targeting fragments of the 16S rRNA, ITS and rpoB genes confirmed the identity of the microorganisms as a Spiroplasma sp., with between 98.9% and 99.5% similarity to S. ixodetis. The sequences of the spiroplasmas isolated from three different pools of ticks collected on two different occasions were identical for all three genes tested.
Keywords: Spiroplasma, Tick, Ixodes ricinus, Tick cell line
Introduction
Spiroplasmas are helical mycoplasmas that infect plants and/or arthropods and may be pathogenic or commensal. Several species of ixodid ticks have been reported to harbour spiroplasmas, including Haemaphysalis leporispalustris (Clark, 1964; Tully et al., 1977), Ixodes pacificus (Tully et al., 1981, 1995), Dermacentor marginatus and Ixodes ricinus (Hornok et al., 2010). Using a PCR assay targeting the Spiroplasma rpoB gene, Subramanian et al. (2012) detected a Spiroplasma that they identified as Spiroplasma ixodetis in 1/52 nymphal and 1/28 adult questing I. ricinus ticks collected in the Podunajske Biskupice area of south-western Slovakia. Tick-borne spiroplasmas can be cultivated in vitro in axenic culture (Tully et al., 1977), and tick cell lines were used successfully to isolate S. ixodetis from I. pacificus ticks (Yunker et al., 1987). Henning et al. (2006) reported isolation in mammalian cells (African green monkey kidney cell line BGM) of a Spiroplasma sp. from a pool of Ixodes ticks collected from a roe deer in Germany; the Spiroplasma was cultured for 10 weeks and sequencing of a fragment of the 16S rRNA gene revealed that it was closely related to S. ixodetis.
In the present study, unfed adult I. ricinus, a widespread tick species that parasitises a wide range of vertebrate hosts and frequently attacks humans, were collected on two different occasions from vegetation in the campus of the Slovak Academy of Sciences (SAS) in Bratislava, Slovakia. Tick tissues were inoculated into a panel of Ixodes spp. tick cell lines with the aim of isolating and propagating microorganisms present in the ticks. Here we report isolation, prolonged in vitro cultivation and partial characterisation of a Spiroplasma sp. from the Slovakian ticks.
Materials and methods
Ticks
Unfed adult male and female I. ricinus ticks were collected by flagging from vegetation in the campus of the SAS, Bratislava, Slovakia, 48.17° N, 17.07°E, altitude circa 190 m above sea level, in April and June 2013. The SAS campus is a fenced area of 32 ha located on the south-western foothills of the Small Carpathians. Patches of the original oak-hornbeam forest with admixture of beech, ash, black locust, maple, limetree, elm, alder, common hazel and elder are fragmented by roads, pavements and built-up areas. Twenty-one male and 19 female ticks were collected in April 2013, and 19 male and 26 female ticks were collected in June 2013. Following microscopic examination to confirm species identity, the ticks were transferred to The Pirbright Institute where they were incubated at 15 °C, 100% relative humidity and processed within 9 days of receipt. Batches of male or female ticks were surface-sterilised by immersion in a 0.1% aqueous solution of benzalkonium chloride for 5 min and 70% ethanol for 1 min, followed by 2 rinses in sterile deionised water. The ticks were allowed to dry on sterile filter paper in a petri dish and then immersed as pools of 4–11 ticks in 1–2 ml Hanks balanced salt solution (HBSS). Using a sterile scalpel blade and watchmakers forceps, the ticks were cut into several pieces and as much of their internal organs separated from the exoskeleton as possible using the forceps and pressure from the flattened end of a glass rod. The tissue suspension was collected by pipetting, leaving as much of the exoskeleton behind as possible, and inoculated into tick cell lines as described below.
Tick cell lines
The I. ricinus embryo-derived cell lines IRE/CTVM19, IRE/CTVM20 and IRE11, and the Ixodes scapularis embryo-derived cell lines IDE2, IDE8, ISE6 and ISE18 were grown at 28 °C or 32 °C in sealed, flat-sided culture tubes (Nunc) in 2.2 ml L-15 (Leibovitz)-based media supplemented as shown in Table 1. Cultures were inoculated with 0.2–0.3 ml of I. ricinus tissue suspension, incubated at the appropriate temperature for the respective cell line. Medium was changed weekly by removal and replacement of 1.5 ml medium; cultures were monitored weekly by inverted microscope for presence of contamination, and at 2–8 week intervals from day 14–22 post inoculation (p.i.) by preparation and examination of Giemsa-stained cytocentrifuge smears. When presence of putative tick-borne microorganisms was detected by microscopy, supernatant medium was passaged onto fresh cultures of the same cell line and monitoring continued as above.
Table 1.
Ixodes spp. embryo-derived cell lines used in this study.
| Tick species | Cell line | Incubation temperature (°C) | Culture medium | Reference |
|---|---|---|---|---|
| Ixodes ricinus | IRE/CTVM19 | 28 | L-15a | Bell-Sakyi et al. (2007) |
| IRE/CTVM20 | 28 | L-15/L-15Bb,d | Bell-Sakyi et al. (2007) | |
| IRE11 | 32 | L-15B300c | Simser et al. (2002) | |
| Ixodes scapularis | IDE2 | 32 | L-15B300 | Munderloh et al. (1994) |
| IDE8 | 32 | L-15B | Munderloh et al. (1994) | |
| ISE6 | 32 | L-15B300 | Kurtti et al. (1996) | |
| ISE18 | 32 | L-15B300 | Munderloh et al. (1994) | |
L-15: L-15 (Leibovitz) medium supplemented with 10% tryptose phosphate broth (TPB), 20% foetal bovine serum (FBS), 2 mM l-glutamine (l-glut), 100 units/ml penicillin and 100 μg/ml streptomycin (pen/strep).
L-15B: L-15B medium (Munderloh and Kurtti, 1989) supplemented with 10% TPB, 5% FBS, 0.1% bovine lipoprotein (MP Biomedicals), l-glut and pen/strep.
L-15B300: L-15B medium diluted 3:1 with deionised water, supplemented with 10% TPB, 5% FBS, 0.1% bovine lipoprotein (MP Biomedicals), l-glut and pen/strep.
A 1:1 mixture of the two complete media.
Molecular detection and identification of Spiroplasma species
Aliquots of 1 ml of culture suspension were processed for DNA extraction using a DNeasy Blood and Tissue Kit (Qiagen) following the manufacturer's instructions for cultured cells. Presence and initial identification of bacteria was assayed using a pan-bacterial PCR targeting a 528 bp fragment of the 16S rRNA gene (Benson et al., 2004) as described previously (Alberdi et al., 2012).
Specific PCRs targeting the 16S-23S rRNA intergenic transcribed spacer (ITS; 16S-F-MYC & 23S-R1-MYC primers; 600–1000 bp) (Volokhov et al., 2006) and a fragment of the RNA polymerase beta subunit gene (rpoB; RpoBF3 & RpoBR2 primers; 1443 bp) (Haselkorn et al., 2009) of Spiroplasma were carried out as described by the respective authors. In addition, a pan-bacterial PCR targeting a longer fragment of the Spiroplasma 16S rRNA gene (fD1 & rP2 primers; 1500 bp) (Weisburg et al., 1991), incorporating the sequence detected by the PCR of Benson et al. (2004), was performed on the samples that were positive in the Spiroplasma-specific PCRs. A negative control containing water instead of template DNA was included in all PCRs. Positive PCR products of, or close to, the expected size were purified using a High Pure PCR Product Purification kit (Roche Life Science) following the manufacturer's instructions. Amplification products were sequenced in the forward and reverse directions, and homology searches were performed in the NCBI database using the BLAST search programme (http://blast.ncbi.nlm.nih.gov/Blast.cgi). Sequences were aligned using the European Bioinformatics Institute multisequence software ClustalW2 (http://www.ebi.ac.uk/Tools/msa/clustalw2) for multiple sequence alignment. Phylogenetic analyses were conducted using MEGA version 6 (www.megasoftware.net). A phylogenetic tree was constructed by the neighbour-joining method. Confidence values for individual branches of the resulting tree were determined by bootstrap analysis with 1000 replicates. The evolutionary distances were computed using the maximum composite likelihood method.
Results
Extracts of 8 pools of 4–7 male ticks and 8 pools of 4–11 female ticks were inoculated into a total of 44 cultures from one or more tick cell lines (Table 2). Of these, 37 were lost to gross bacterial and/or fungal contamination between 6 and 200 days p.i. Subculture series were initiated from surviving cultures between 69 and 116 days p.i.; between two and seven subcultures were carried out (Table 2). Between 478 and 533 days p.i., all seven surviving culture series were harvested, parent cultures (if surviving) and their subcultures were pooled, aliquots were cryopreserved in liquid nitrogen with 10% DMSO, and DNA was extracted from 1 ml of pooled culture suspension.
Table 2.
Inoculation of tick cell lines with pooled Ixodes ricinus extracts. Only those pools from which at least one surviving culture was obtained are shown. The results of PCR analysis of the seven surviving cultures are shown with the day of final sampling and highest passage level (p) achieved.
| Tick origin | Pool composition | Cell lines inoculated | Result |
|---|---|---|---|
| Collected 16th–17th April 2013, processed 6th May 2013 | 4 males | IRE/CTVM19 | PCR negative, day 533, p4 |
| 4 males | ISE6 | PCR negative, day 533, p4 | |
| 5 males | IDE8 | PCR negative, day 533, p2 | |
| Collected 16th–17th April 2013, processed 12th May 2013 | 4 males | IRE/CTVM19 | Spiroplasma positive, day 527, p5 |
| ISE6 | Contaminated | ||
| ISE18 | PCR negative, day 527, p2 | ||
| IDE8 | Contaminated | ||
| 8 females | IRE/CTVM19 | Contaminated | |
| IRE/CTVM20 | Contaminated | ||
| IRE11 | Contaminated | ||
| ISE6 | Contaminated | ||
| IDE2 | Spiroplasma positive, day 527, p5 | ||
| IDE8 | Contaminated | ||
| Collected 19th–20th June 2013, processed 30th June 2013 | 6 males | IRE/CTVM19 | Contaminated |
| IRE/CTVM20 | Contaminated | ||
| IRE11 | Spiroplasma positive, day 478, p7 | ||
| ISE6 | Contaminated | ||
Spiroplasmas were first identified in Giemsa-stained smears (Fig. 1) from three of the parent cultures in the cell lines IRE/CTVM19, IRE11 and IDE2 after 2–3 months in vitro (Table 2). Most of the spiroplasmas were located within intracellular vacuoles, forming more or less loosely-packed (Fig. 1A and B) or densely-packed (Fig. 1B) accumulations of thread-like and coccal organisms within cytoplasmic vacuoles. Occasionally free spiroplasmas with typical helical morphology were seen (Fig. 1C), possibly released from vacuoles during the cytocentrifugation process. Spiroplasma infection and growth was maintained through five subcultures in IRE/CTVM19 and IDE2 cells and seven subcultures in IRE11 cells. Presence of the Spiroplasma resulted in complete destruction of IDE2 cells, while some IRE11 cells and most IRE/CTVM19 cells survived the infection; the latter phenomenon appeared to result from development of tolerance by the whole culture, rather than the presence of a subpopulation of cells refractory to Spiroplasma infection within the cell lines. Spiroplasmas were not seen in any of the remaining cultures, including ISE18 cells originally inoculated with the same pooled material as the Spiroplasma-positive IRE/CTVM19 cells.
Fig. 1.
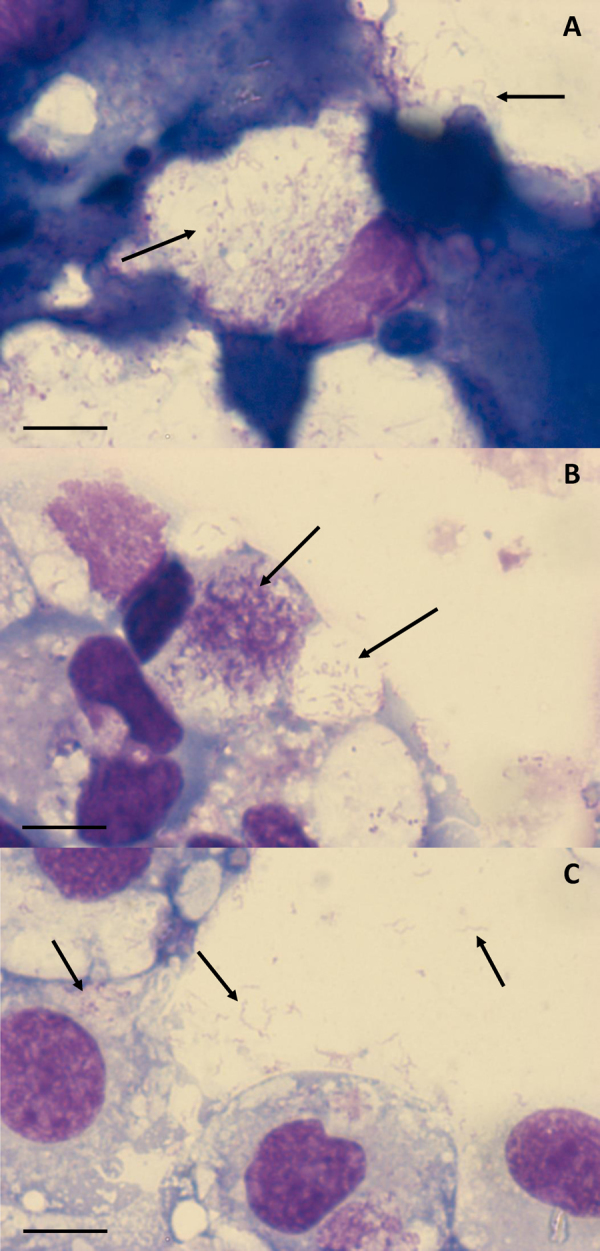
Giemsa-stained cytocentrifuge smears of Ixodes-derived cell lines inoculated with Ixodes ricinus tissues. (A) I. ricinus IRE/CTVM19 cells 533 days p.i.; (B) Ixodes scapularis IDE2 cells 527 days p.i.; (C) I. ricinus IRE11 cells 478 days p.i. Arrows indicate spiroplasmas. Scale bars = 10 μm.
Molecular analysis of DNA extracted from the seven surviving uncontaminated cultures confirmed the presence of a Spiroplasma sp. in the three cultures that showed spiroplasmas in Giemsa-stained smears; sequencing of PCR products from all three cultures gave identical results for each target gene. No PCR products were amplified from any of the other four cultures by any of the PCR assays. Initial screening with the pan-bacterial 16S rRNA PCR of Benson et al. (2004) yielded 483 bp products that showed high levels of identity with published sequences of Spiroplasma spp. and uncultured bacteria obtained from a variety of arthropods (Table 3). The highest similarity (100%) was with an uncultured Spiroplasma sp. of the little housefly Fannia manicata, and highest similarity to a valid Spiroplasma species was 99.4% with S. ixodetis strain Y32 isolated from the tick I. pacificus (GenBank accession number NR104852). Similar results were obtained with the pan-bacterial PCR of Weisburg et al. (1991) targeting the longer 16S rRNA gene fragment, that yielded 1391 bp products which again showed high levels of identity with arthropod-derived Spiroplasma spp. and uncultured bacteria (Table 3), including 99.5% similarity to S. ixodetis strain Y32.
Table 3.
Levels of identity between the 16SrRNA sequences obtained in the present study and published sequences from Spiroplasma spp. and uncultured bacteria detected in ticks and other arthropods.
| Bacterium | Arthropod host | GenBank accession no. | % Identity (bp) |
|
|---|---|---|---|---|
| Short fragmenta | Long fragmentb | |||
| Spiroplasma sp. | Fannia manicata | AY569829 | 100 (483/483) | 100 (1391/1391) |
| Uncultured bacterium | Ctenocephalides felis | EF121346 | 100 (483/483) | 99.8 (1388/1391) |
| Spiroplasma sp. | Anisosticta novemdecimpunctata | AM087471 | 99.8 (482/483) | 99.9 (1389/1391) |
| Spiroplasma sp. | Anopheles funestus | AY837731 | 99.8 (482/483) | 99.8 (1388/1391) |
| Spiroplasma sp. | A. funestus | AY837732 | 99.8 (482/483) | 99.7 (1387/1391) |
| Uncultured bacterium | Meimuna mongolica | KC424774 | 99.8 (482/483) | 99.6 (1386/1391) |
| Uncultured bacterium | M. mongolica | KC424775 | 99.8 (482/483) | 99.6 (1385/1391) |
| Spiroplasma sp. | Antonina crawii | AB030022 | 99.6 (481/483) | 99.8 (1388/1391) |
| Spiroplasma sp. | Harmonia axyridis | AJ132412 | 99.6 (481/483) | 99.6 (1385/1391) |
| Spiroplasma ixodetis | Ixodes pacificus | NR_104852 | 99.4 (481/484) | 99.5 (1386/1393) |
| Spiroplasma sp. | Acyrthosiphon pisum | AB048263 | 99.4 (480/483) | 99.7 (1387/1391) |
| Spiroplasma sp. | Laodelphax striatellus | AB553862 | 99.4 (480/483) | 99.6 (1383/1389) |
| Spiroplasma sp. | Agathemera claraziana | JF266577 | 99.4 (480/483) | 99.6 (1386/1391) |
483 bp sequence obtained using the PCR of Benson et al. (2004).
1391 bp sequence obtained using the PCR of Weisburg et al. (1991).
The sequences obtained with the Spiroplasma-specific PCRs targeting the ITS (815 bp) and rpoB (1398 bp) gene fragments showed the highest identities (98.9% and 99.2% respectively) amongst valid Spiroplasma species with those from S. ixodetis strain Y30 (GenBank accession numbers DQ004912 and DQ313833 respectively). There are no published sequences for the ITS or rpoB genes of most of the uncultured Spiroplasma spp. for which 16S rRNA sequences are available (Table 3); an exception is the Spiroplasma sp. detected in the planthopper Laodelphax striatellus for which the rpoB sequence (GenBank accession number AB797266) showed 99.5% similarity to the rpoB sequence obtained in the present study.
The sequences obtained for the 16S rRNA (1391 bp), ITS and rpoB genes, from the spiroplasmas isolated in the IRE/CTVM19 cell line and designated Spiroplasma sp. strain Bratislava 1, were submitted to GenBank with accession numbers KP967685, KP967686 and KP967687 respectively. Table 4 shows the similarities between the partial 16S rRNA gene sequence amplified from Spiroplasma sp. strain Bratislava 1 in the present study and other sequences amplified from Ixodes spp. ticks available in GenBank. A phylogenetic tree based on the three concatenated genes (16SrRNA, ITS, rpoB) of Spiroplasma sp. strain Bratislava 1 presents the relationship between them and those of other Spiroplasma spp. (Fig. 2), and shows that the Spiroplasma sp. isolated in the present study belongs to the same clade as S. ixodetis.
Table 4.
Similarity of the partial 16S rRNA gene sequence of Spiroplasma sp. strain Bratislava 1 isolated from Ixodes ricinus ticks in the present study with Spiroplasma spp. sequences amplified from Ixodes spp. ticks and deposited in GenBank.
| Spiroplasma (reference) | Tick species | GenBank accession no. | Length (bp) | No. | 1 | 2 | 3 | 4 | 5 | 6 |
|---|---|---|---|---|---|---|---|---|---|---|
| Spiroplasma sp. Bratislava 1 (this study) | I. ricinus | KP967685 | 1391 | 1 | 100 | |||||
| S. ixodetis (ATCC:33835) (Tully et al., 1995) | I. pacificus | NR_104852 | 1451 | 2 | 99.5 | 100 | ||||
| Spiroplasma sp. (Henning et al., 2006) | Ixodes sp. | DQ012506 | 219 | 3 | 98.6 | 98.2 | 100 | |||
| Spiroplasma sp. (Hornok et al., 2010) | I. ricinus |
EU170606 EU170609 EU1706014 EU1706015 |
195 | 4 | 98.5 | 97.5 | 97.7 | 100 | ||
| Spiroplasma sp. (Hornok et al., 2010) | I. ricinus |
EU170607 EU170608 |
195 | 5 | 96.4 | 95.4 | 96.6 | 98.0 | 100 | |
| Spiroplasma sp. (Halos et al., unpublished, 2005) | I. ricinus | DQ065809 | 171 | 6 | 76.8 | 81.2 | 88.4 | 93.2 | 91.4 | 100 |
Fig. 2.

Unrooted dendrogram showing the phylogenetic position of Spiroplasma sp. strain Bratislava 1 (♦), isolated in the present study, among valid Spiroplasma species. Phylogeny is inferred from comparison of 16S rRNA (1391 bp), ITS (815 bp) and rpoB (1398 bp) amino-acid sequences by the neighbour-joining method (1000 replicates). Mycoplasma hominis is used as outgroup. GenBank accession numbers of the genes used in the comparison are shown in brackets following each Spiroplasma species, with multiple accession numbers separated by dashes.
Discussion
Several ixodid tick genera and species have previously been reported to harbour spiroplasmas, but actual microorganisms have only been isolated from H. leporispalustris (Spiroplasma mirum, Clark, 1964), I. pacificus (S. ixodetis, Tully et al., 1977; Yunker et al., 1987) and an unspecified Ixodes species in Germany (Spiroplasma sp., Henning et al., 2006). All other reports depended on molecular detection of Spiroplasma DNA sequences in tick extracts, for example Taroura et al. (2005), Hornok et al. (2010), Tveten and Sjastad (2011), Subramanian et al. (2012) and Qiu et al. (2014). In the present study, a Spiroplasma sp. with highest similarity to S. ixodetis was isolated from three separate pools of unfed adult I. ricinus ticks; the microorganism was present in both male and female ticks, grew well in cell lines derived from both I. ricinus and I. scapularis at 28 °C or 32 °C and could be maintained in vitro with serial passage in tick cells for over 500 days.
As in the present study, all previously-described tick-borne spiroplasmas were isolated either from nymphal or adult ticks (Clark, 1964; Tully et al., 1977, 1995; Yunker et al., 1987) or from ticks of unspecified developmental stage but feeding on a roe deer (Henning et al., 2006). Therefore it was not possible to determine whether these spiroplasmas originated from a current or previous bloodmeal, or were transovarially-transmitted tick endosymbionts. However, spiroplasmas have also been isolated from primary cell cultures of embryonic tissues of I. ricinus and Dermacentor reticulatus ticks originating from The Netherlands (author's unpublished results), indicating that in these cases the spiroplasmas were transovarially transmitted.
Considering the ease and frequency with which spiroplasmas can be isolated from field ticks, it is surprising that there are not more records of their presence in field tick samples. The low PCR-based prevalence of 3% reported for Slovakian I. ricinus ticks by Subramanian et al. (2012), combined with the time taken for spiroplasmas to appear in Giemsa-stained smears in the present study, suggests that they may be present at very low densities in intact ticks. In the present study, spiroplasmas were detected in surviving cultures derived from three out of six tick pools; the positive pools comprised 4, 6 and 8 ticks each, giving a minimum prevalence in positive pools of 3/18 or 16.7% and a minimum prevalence overall of 3/31 or 9.7%. It is possible that tick cell culture isolation is a more sensitive, albeit much slower, test for Spiroplasma infection in ticks than molecular detection by PCR amplification. The 16S amplicon pyrosequencing approach of Qiu et al. (2014) indicated a much higher prevalence of Spiroplasma in Japanese Ixodes ovatus, but the difference between the results for this species and for I. ricinus could be due to different methodology, different infection rates or a combination of both.
The Spiroplasma sp. isolated in the present study appears to be closely related to S. ixodetis isolated from North American I. pacificus ticks (Tully et al., 1995). Similarly closely-related spiroplasmas have been detected in other tick species including European I. ricinus from Slovakia (Subramanian et al., 2012) and Ixodes sp. from Germany (Henning et al., 2006). Furthermore, Spiroplasma spp. closely related to S. ixodetis have been reported from several insect groups, including planthoppers (L. striatellus) (Sanada-Morimura et al., 2013), mosquitoes (Anopheles funestus) (Lindh et al., 2005), ladybirds (Anisosticta novemdecimpunctata) (Tinsley & Majerus, 2006) and mealybugs (Antonina crawii) (Fukatsu & Nikoh, 2000). However, those reports were based on the homology of a few gene fragments, usually a single fragment of the 16S rRNA gene. Sequence comparison of additional genes will be required to clarify the phylogenetic relationships between tick- and insect-borne spiroplasmas.
Spiroplasma spp. are increasingly being recognised as playing a role in human and animal disease. The first tick-derived spiroplasma to be discovered, S. mirum, has long been known to induce cataract formation in experimentally-infected suckling mice (Clark, 1964). More recently, a Spiroplasma closely related to mosquito-borne Spiroplasma taiwanense was found in the conjunctiva of a premature baby that acquired unilateral cataract with anterior uveitis (Lorenz et al., 2002), and Spiroplasma turonicum was implicated as the causal agent of a systemic human infection in Spain (Aquilino et al., 2015). Furthermore, spiroplasmas have been linked with transmissible spongiform encephalopathy in ruminants and humans (Bastian, 2014). While the aforementioned presence of spiroplasmas in tick eggs indicates that these microorganisms are natural endosymbionts of at least two European tick species, the putative role of ticks in transmission of these spiroplasmas to vertebrate hosts, and the potential infectivity and pathogenicity of tick-borne spiroplasmas other than S. mirum for vertebrates, require further investigation. Such studies will be greatly facilitated by the ability to isolate spiroplasmas from all developmental stages of field ticks into tick cell lines and to cultivate the spiroplasmas therein over prolonged periods and to high numbers, as demonstrated by the present study.
Acknowledgements
The authors would like to thank Dr Ulrike Munderloh of the University of Minnesota for provision of the I. scapularis cell lines and the IRE11 cell line. All cell lines used in this study are maintained in the Tick Cell Biobank at The Pirbright Institute. LBS is supported by the United Kingdom Biotechnology and Biological Sciences Research Council's National Capability Grant to the Pirbright Institute. AMP is supported by Fundación Rioja Salud (grant: FRS/PIF-01/10). The study was partially funded by EU grant FP7-261504 EDENext and is catalogued by the EDENext Steering Committee as EDENext341 (http://www.edenext.eu). The contents of this publication are the sole responsibility of the authors and do not necessarily reflect the views of the European Commission.
References
- Alberdi M.P., Dalby M.J., Rodriguez-Andres J., Fazakerley J.K., Kohl A., Bell-Sakyi L. Detection and identification of putative bacterial endosymbionts and endogenous viruses in tick cell lines. Ticks Tick-borne Dis. 2012;3:137–146. doi: 10.1016/j.ttbdis.2012.05.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aquilino A., Masiá M., López P., Galiana A.J., Tovar J., Andrés M., Gutiérrez F. First human systemic infection caused by Spiroplasma. J. Clin. Microbiol. 2015;53:719–721. doi: 10.1128/JCM.02841-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bastian F.O. The case for involvement of spiroplasma in the pathogenesis of transmissible spongiform encephalopathies. J. Neuropathol. Exp. Neurol. 2014;73:104–114. doi: 10.1097/NEN.0000000000000033. [DOI] [PubMed] [Google Scholar]
- Bell-Sakyi L., Zweygarth E., Blouin E.F., Gould E.A., Jongejan F. Tick cell lines: tools for tick and tick-borne disease research. Trends Parasitol. 2007;23:450–457. doi: 10.1016/j.pt.2007.07.009. [DOI] [PubMed] [Google Scholar]
- Benson M.J., Gawronski J.D., Eveleigh D.E., Benson D.R. Intracellular symbionts and other bacteria associated with deer ticks (Ixodes scapularis) from Nantucket and Wellfleet, Cape Cod, Massachusetts. Appl. Environ. Microbiol. 2004;70:616–620. doi: 10.1128/AEM.70.1.616-620.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clark H.F. Suckling mouse cataract agent. J. Infect. Dis. 1964;114:476–487. doi: 10.1093/infdis/114.5.476. [DOI] [PubMed] [Google Scholar]
- Fukatsu T., Nikoh N. Endosymbiotic microbiota of the bamboo pseudococcid Antonina crawii (Insecta, Homoptera) Appl. Environ. Microbiol. 2000;66:643–650. doi: 10.1128/aem.66.2.643-650.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haselkorn T.S., Markow T.A., Moran N.A. Multiple introductions of the Spiroplasma bacterial endosymbiont into Drosophila. Mol. Ecol. 2009;18:1294–1305. doi: 10.1111/j.1365-294X.2009.04085.x. [DOI] [PubMed] [Google Scholar]
- Henning K., Greiner-Fischer S., Hotzel H., Ebsen M., Theegarten D. Isolation of a Spiroplasma sp. from an Ixodes tick. Int. J. Med. Microbiol. 2006;296(S1):157–161. doi: 10.1016/j.ijmm.2006.01.012. [DOI] [PubMed] [Google Scholar]
- Hornok S., Meli M.L., Perreten A., Farkas R., Willi B., Beugnet F., Lutz H., Hofmann-Lehmann R. Molecular investigation of hard ticks (Acari: Ixodidae) and fleas (Siphonaptera: Pulicidae) as potential vectors of rickettsial and mycoplasmal agents. Vet. Microbiol. 2010;140:98–104. doi: 10.1016/j.vetmic.2009.07.013. [DOI] [PubMed] [Google Scholar]
- Kurtti T.J., Munderloh U.G., Andreadis T.G., Magnarelli L.A., Mather T.N. Tick cell culture isolation of an intracellular prokaryote from the tick Ixodes scapularis. J. Invert. Pathol. 1996;67:318–321. doi: 10.1006/jipa.1996.0050. [DOI] [PubMed] [Google Scholar]
- Lindh J.M., Terenius O., Faye I. 16S rRNA gene-based identification of midgut bacteria from field-caught Anopheles gambiae sensu lato and A. funestus mosquitoes reveals new species related to known insect symbionts. Appl. Environ. Microbiol. 2005;71:7217–7223. doi: 10.1128/AEM.71.11.7217-7223.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lorenz B., Schroeder J., Reisch U. First evidence of an endogenous Spiroplasma sp. infection in humans manifesting as unilateral cataract associated with anterior uveitis in a premature baby. Graefe's Arch. Clin. Exp. Ophthalmol. 2002;240:348–353. doi: 10.1007/s00417-002-0453-3. [DOI] [PubMed] [Google Scholar]
- Munderloh U.G., Kurtti T.J. Formulation of medium for tick cell culture. Exp. Appl. Acarol. 1989;7:219–229. doi: 10.1007/BF01194061. [DOI] [PubMed] [Google Scholar]
- Munderloh U.G., Liu Y., Wang M., Chen C., Kurtti T.J. Establishment, maintenance and description of cell lines from the tick Ixodes scapularis. J. Parasitol. 1994;80:533–543. [PubMed] [Google Scholar]
- Qiu Y., Nakao R., Ohnuma A., Kawamori F., Sugimoto C. Microbial population analysis of the salivary glands of ticks; a possible strategy for the surveillance of bacterial pathogens. PLOS ONE. 2014;9(8):e103961. doi: 10.1371/journal.pone.0103961. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sanada-Morimura S., Matsumura M., Noda H. Male killing caused by a Spiroplasma symbiont in the small brown planthopper, Laodelphax striatellus. J. Hered. 2013;104:821–829. doi: 10.1093/jhered/est052. [DOI] [PubMed] [Google Scholar]
- Simser J.A., Palmer A.T., Fingerle V., Wilske B., Kurtti T.J., Munderloh U.G. Rickettsia monacensis sp. nov., a spotted fever group rickettsia, from ticks (Ixodes ricinus) collected in a European city park. Appl. Env. Microbiol. 2002;68:4559–4566. doi: 10.1128/AEM.68.9.4559-4566.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Subramanian G., Sekeyova Z., Raoult D., Mediannikov O. Multiple tick-associated bacteria in Ixodes ricinus from Slovakia. Ticks Tick-borne Dis. 2012;3:405–409. doi: 10.1016/j.ttbdis.2012.10.001. [DOI] [PubMed] [Google Scholar]
- Taroura S., Shimada Y., Sakata Y., Miyama T., Hiraoka H., Watanabe M., Itamoto K., okuda M., Inokuma H. Detection of DNA of “Candidatus Mycoplasma haemominutum” and Spiroplasma sp. in unfed ticks collected from vegetation in Japan. J. Vet. Med. Sci. 2005;67:1277–1279. doi: 10.1292/jvms.67.1277. [DOI] [PubMed] [Google Scholar]
- Tinsley M.C., Majerus M.E.N. A new male-killing parasitism: Spiroplasma bacteria infect the ladybird beetle Anisosticta novemdecimpunctata (Coleoptera: Coccinellidae) Parasitology. 2006;132:757–765. doi: 10.1017/S0031182005009789. [DOI] [PubMed] [Google Scholar]
- Tveten A.K., Sjastad K.K. Identification of bacteria infecting Ixodes ricinus ticks by 16S rDNA amplification and denaturing gradient gel electrophoresis. Vector-borne Zoonot. Dis. 2011;11:1329–1334. doi: 10.1089/vbz.2011.0657. [DOI] [PubMed] [Google Scholar]
- Tully J.G., Whitcomb R.F., Clark H.F., Williamson D.L. Pathogenic mycoplasmas: cultivation and vertebrate pathogenicity of a new Spiroplasma. Science. 1977;195:892–894. doi: 10.1126/science.841314. [DOI] [PubMed] [Google Scholar]
- Tully J.G., Rose D.L., Yunker C.E., Cory J., Whitcomb R.F., Williamson D.L. Helical mycoplasmas (spiroplasmas) from Ixodes ticks. Science. 1981;212:1043–1045. doi: 10.1126/science.7233197. [DOI] [PubMed] [Google Scholar]
- Tully J.G., Rose D.L., Yunker C.E., Carle P., Bové J.M., Williamson D.L., Whitcomb R.F. Spiroplasma ixodetis sp. nov., a new species from Ixodes pacificus ticks collected in Oregon. Int. J. Syst. Bacteriol. 1995;45:23–28. doi: 10.1099/00207713-45-1-23. [DOI] [PubMed] [Google Scholar]
- Volokhov D.V., George J., Liu S.X., Ikonomi P., Anderson C., Chizhikov V. Sequencing of the intergenic 16S-23S rRNA spacer (ITS) region of Mollicutes species and their identification using microarray-based assay and DNA sequencing. Appl. Microbiol. Biotechnol. 2006;71:680–698. doi: 10.1007/s00253-005-0280-7. [DOI] [PubMed] [Google Scholar]
- Weisburg W.G., Barns S.M., Pelletier D.A., Lane D.J. 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 1991;173:697–703. doi: 10.1128/jb.173.2.697-703.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yunker C.E., Tully J.G., Cory J. Arthropod cell lines in the isolation and propagation of tickborne spiroplasmas. Curr. Microbiol. 1987;15:45–50. [Google Scholar]