

Abstract
Background
Almond witches’-broom (AlmWB), a devastating disease of almond, peach and nectarine in Lebanon, is associated with ‘Candidatus Phytoplasma phoenicium’. In the present study, we generated a draft genome sequence of ‘Ca. P. phoenicium’ strain SA213, representative of phytoplasma strain populations from different host plants, and determined the genetic diversity among phytoplasma strain populations by phylogenetic analyses of 16S rRNA, groEL, tufB and inmp gene sequences.
Results
Sequence-based typing and phylogenetic analysis of the gene inmp, coding an integral membrane protein, distinguished AlmWB-associated phytoplasma strains originating from diverse host plants, whereas their 16S rRNA, tufB and groEL genes shared 100 % sequence identity. Moreover, dN/dS analysis indicated positive selection acting on inmp gene. Additionally, the analysis of ‘Ca. P. phoenicium’ draft genome revealed the presence of integral membrane proteins and effector-like proteins and potential candidates for interaction with hosts. One of the integral membrane proteins was predicted as BI-1, an inhibitor of apoptosis-promoting Bax factor. Bioinformatics analyses revealed the presence of putative BI-1 in draft and complete genomes of other ‘Ca. Phytoplasma’ species.
Conclusion
The genetic diversity within ‘Ca. P. phoenicium’ strain populations in Lebanon suggested that AlmWB disease could be associated with phytoplasma strains derived from the adaptation of an original strain to diverse hosts. Moreover, the identification of a putative inhibitor of apoptosis-promoting Bax factor (BI-1) in ‘Ca. P. phoenicium’ draft genome and within genomes of other ‘Ca. Phytoplasma’ species suggested its potential role as a phytoplasma fitness-increasing factor by modification of the host-defense response.
Electronic supplementary material
The online version of this article (doi:10.1186/s12866-015-0487-4) contains supplementary material, which is available to authorized users.
Keywords: Phytoplasma, Parasitism, Vector, Integral membrane protein, BI-1
Background
Phytoplasmas are bacterial plant pathogens that cause economically relevant yield losses in several low- and high- value annual and perennial crops worldwide, including fruit and woody trees [1, 2]. They are transmitted plant-to-plant by phloem feeders of the order Hemiptera, mostly leafhoppers (Cicadellidae), planthoppers (Fulgoroidea) and psyllids (Psyllidae) [3]. Phytoplasmas are classified into the class Mollicutes, which includes bacteria with single membrane that have diverged from a Gram-positive ancestor [4, 5]. Based on molecular and other biological features phytoplasma strains have been classified into 37 species within the provisional genus ‘Candidatus Phytoplasma’ [6, 7] and taxonomic groupings have also been delimited according to the DNA sequence coding for their 16S ribosomal RNA [8].
Phytoplasmas of taxonomic group 16SrIX (pigeon pea witches’-broom group) are associated with diseases affecting crop and wild plants in different geographic areas worldwide [9–12]. ‘Candidatus Phytoplasma phoenicium’, taxonomic subgroup 16SrIX-B [13, 14], also designated as 16SrIX-D [15, 16], and its genetic variants [16] are the etiological agents of a lethal devastating disease of almond trees (almond witches’-broom, AlmWB) in Lebanon and in Iran [10, 13, 17, 18]. A similar disease, inducing almond broomings, was reported in Iran [10] in association with phytoplasmas close to those responsible for Knautia arvensis phyllody (KAP), subgroup 16SrIX-C [18]. Furthermore, almond trees showing different symptoms, such as small and yellow leaves, were found infected by ‘Ca. P. aurantifolia’, ‘Ca. P. solani’ and ‘Ca. P. trifolii’ in Iran [19]. AlmWB was also identified on peach (P. persica) and nectarine (P. persica var. nucipersica) in southern Lebanon [20] and on GF-677 (P. amygdalus x P. persica) in Iran [21].
The most characteristic symptoms caused by AlmWB on almond trees are (i) shoot proliferation on the main trunk with the appearance of a witches’-broom, (ii) the perpendicular development of many axillary buds on the branches, with smaller and yellowish leaves, (iii) the general decline of the tree with final dieback. A total loss of production happens 1–2 years after the initial appearance of the symptoms [13]. In the case of peach and nectarine trees, the first symptom observed is early flowering (15 to 20 days earlier than normal), followed by the earlier development of all the buds of the infected branches. In addition, phyllody at the flowering period and serrate, slim, light green leaves and witches’-brooms developing from the trunk and the crown of the trees several months after are observed [20].
Due to complex ecology of ‘Ca. P. phoenicium’, associated with diverse crops, in which it induces variations in symptoms, it is necessary to evaluate the genetic diversity among AlmWB phytoplasma populations in order to determine phytoplasma strain-specific molecular markers. In previous studies, genetic heterogeneity among phytoplasma strain populations was investigated by multilocus sequence typing analyses [22–24]. Recently, Lee and colleagues [14] found that ‘Ca. P. phoenicium’ strains from Lebanon shared identical nucleotide sequences of the genes rplV-rpsC and secY, distinct from phytoplasmas belonging to other 16SrIX taxonomic subgroups. Such findings highlighted that more variable genes should be investigated to determine the diversity among ‘Ca. P. phoenicium’ strains. In previous studies, draft or full length genome sequencing allowed not only the acquisition of key information on phytoplasma metabolism and mechanisms of interaction with hosts [25–33], but also the identification of hyper-variable genes suitable for distinguishing closely related strains of ‘Ca. P. solani’ (genes vmp and stamp) [34–36] and ‘Ca. P. mali’ (gene hflB) [27, 37].
In the present study, we obtained the draft genome sequence of a ‘Ca. P. phoenicium’ strain SA213 identified in almond from Lebanon. Different house-keeping and variable genes were selected in order to determine genetic variability among ‘Ca. P. phoenicium’ strain populations by sequencing and phylogenetic analyses. The results obtained reveal useful information about the complexity of ‘Ca. P. phoenicium’ population structure, and highlight intriguing preliminary insights into the possible interactions with hosts of this phytoplasma.
Methods
Sample collection and DNA extraction
During the spring season of 2012, leaves and shoots from plants showing typical AlmWB symptoms were collected. Specifically, samples were obtained from almond, peach, and nectarine in North (Lat 34.207880, Long 35.677113) and South Lebanon (Lat 33.278748, Long 35.554823), while almond shoots only were sampled in the Bekaa valley (Lat 34.015622, Long 36.176262) (Table 1). After surface disinfection, the phloem shavings or the leaf veins (about 100 mg per sample) were placed in 1.5 ml tubes, immersed immediately in liquid nitrogen and ground using a pestle attached to an electrical drill. The small scale CTAB protocol was used to extract total nucleic acids as previously described [13]. The nucleic acid pellet was washed with 75 % ethanol, air-dried, suspended in 100 μl of sterile water, and maintained at −80 °C until use.
Table 1.
Host, sampling location and accession numbers of 16S rRNA, tufB, groEL and inmp gene nucleotide sequences of analyzed ‘Ca. P. phoenicium’ strains
| Strain | Host | Locality | Accession No. | |||
|---|---|---|---|---|---|---|
| 16S rRNA a | tufB b | groEL c | inmp d | |||
| A48 | Prunus dulcis (Mill.) D.A.Webb | Bekaa | ||||
| A49 | Prunus dulcis (Mill.) D.A.Webb | |||||
| A50 | Prunus dulcis (Mill.) D.A.Webb | |||||
| A51 | Prunus dulcis (Mill.) D.A.Webb | |||||
| A60 | Prunus dulcis (Mill.) D.A.Webb | |||||
| A65 | Prunus dulcis (Mill.) D.A.Webb | |||||
| NA202-1 | Prunus dulcis (Mill.) D.A.Webb | North | ||||
| NA206-1 | Prunus dulcis (Mill.) D.A.Webb | |||||
| NA223-1 | Prunus dulcis (Mill.) D.A.Webb | KM275495 | ||||
| NN204-1 | Prunus persica var. nucipersica | |||||
| NN209-2 | Prunus persica var. nucipersica | KM275499 | ||||
| NP207-1 | Prunus persica (L.) Batsch | KM275496 | ||||
| SA202 | Prunus dulcis (Mill.) D.A.Webb | South | ||||
| SA213 | Prunus dulcis (Mill.) D.A.Webb | KM275491 | KM275492 | KM275493 | KM275494 | |
| SA214 | Prunus dulcis (Mill.) D.A.Webb | |||||
| SA216 | Prunus dulcis (Mill.) D.A.Webb | |||||
| SN205 | Prunus persica var. nucipersica | |||||
| SN209 | Prunus persica var. nucipersica | |||||
| SP202 | Prunus persica (L.) Batsch | KM275498 | ||||
| SP232 | Prunus persica (L.) Batsch | KM275497 | ||||
a 16S rRNA gene nucleotide sequences of other AlmWB phytoplasma strains are identical to the sequence of the strain SA213 (KM275491)
b tufB gene nucleotide sequences of other AlmWB phytoplasma strains are identical to the sequence of the strain SA213 (KM275492) c groEL gene nucleotide sequences of other AlmWB phytoplasma strains are identical to the sequence of the strain SA213 (KM275493)
d inmp gene nucleotide sequences of other AlmWB phytoplasma strains are identical to the sequence of the strain SA213 (KM275494)
Phytoplasma identification by 16S rDNA amplification and sequence analyses
Total nucleic acids from the plants under investigation were employed as templates in PCR reactions amplifying the phytoplasma 16S rRNA gene. Reactions were carried out using the universal primer pair P1/P7 [38, 39] followed by nested PCR using primer pair R16F2n/R16R2 [40], able to amplify partial 16S rDNA sequences of the known species inside the genus ‘Ca. Phytoplasma’. DNAs extracted from Madagascar periwinkle [Catharanthus roseus (L.) G. Don] plants infected by phytoplasma strains EY1 (‘Ca. P. ulmi’, subgroup 16SrV-A), STOL (‘Ca. P. solani’, subgroup 16SrXII-A), and AY1 (‘Ca. P. asteris’, subgroup 16SrI-B) served as reference controls. DNA from healthy periwinkle and reaction mixture devoid of DNA template were used as negative controls. PCRs were performed in an automated thermal cycler (Mastercycler gradient, Eppendorf, Hamburg, Germany). The presence of PCR amplicons was verified by electrophoresis through 1 % agarose gel.
Amplicons from nested PCRs were sequenced to achieve at least 4X sequence coverage per base position. DNA sequencing was performed in an ABI PRISM 377 automated DNA sequencer (Applied Biosystems, Carlsbad CA, USA) by a commercial service (Primm, Milan, Italy). Nucleotide sequence data were assembled by employing the CAP3 assembler module of the Bioedit software, version 7.2.5 [41]. Sequences were compared with the GenBank database using the software BlastN (http://www.ncbi.nim.nih.gov/BLAST/). Affiliation of identified phytoplasmas to taxonomic 16Sr group/subgroup was determined by in silico RFLP analyses of F2n/R2 amplicons carried out using the software iPhyClassifier (http://plantpathology.ba.ars.usda.gov/cgi-bin/resource/iphyclassifier.cgi) [8].
Phytoplasma 16S rRNA gene sequences from this study (Table 1) and from GenBank were used to construct phylogenetic trees. Minimum evolution analysis was carried out using the Neighbor-Joining method and bootstrap replicated 1000 times using the software MEGA5 (http://www.megasoft-ware.net/index.html) [42].
Genome sequencing, assembling and annotation
The ‘Ca. P. phoenicium’ strain SA213, identified in almond in South Lebanon, was selected as representative of phytoplasma strain populations examined in the present study for the genome sequencing. Five micrograms of DNA, extracted from phloem tissue of symptomatic shoots as described above, was used for library preparation to Illumina sequencing carried out by a commercial sequencing service (Institute of Applied Genomics, IGA, Udine, Italy). Barcoded libraries were prepared using the NEBnext DNA sample prep Kit (New England Biolabs, Ipswich, MS, USA) according to the manufacturer’s instructions. Libraries were sequenced on a MiSeq Sequencing System (Illumina, San Diego CA, USA) in a 150-base single read multiplex run.
Quality trimming of reads, mapping and de novo assembly was performed in CLC Genomics Workbench 6.0.2 (http://www.clcbio.com/) applying standard parameters. Nucleotide entries for Acholeplasmataceae deposited in GenBank (2013-01-11) were downloaded, imported in CLC Genomics Workbench and used as reference for read mapping. Reads assigned to Acholeplasmataceae by this approach were selected for de novo assembly (positive read selection). The minimal size for contiguous sequences (contigs) was set to 1000 b.
Contigs were compared via BLASTX [43] against NRPROT database (ftp://ftp.ncbi.nlm.nih.gov/blast/db/). Contigs and BLASTX data were uploaded in MEGAN (MEta Genome ANalyzer) [44] handling contigs as reads and applying a minimal support level of one and low complexity filter off. All sequences with an assignment to the phylum Tenericutes were selected for initial analysis but re-evaluated during the annotation with respect to an unambiguous assignment to the phytoplasma clade. The draft genome was analyzed by the automated annotation pipeline RAST [45] and manually curated in Artemis [46]. Functional protein domains of all predicted proteins were identified by InterProScan 4 [47]. Transmembrane topology and signal peptides in protein sequences from annotated genes were predicted by Phobius [48] providing information on cell localizations of proteins.
To estimate the completeness of ‘Ca. P. phoenicium’ draft genome, the percentage of AlmWB phytoplasma proteins included in the core-genome of ‘Ca. Phytoplasma’ [49] was calculated. Therefore, the ‘Ca. Phytoplasma’ core-genome (294 proteins) and the 333 ‘Ca. P. phoenicium’ proteins were used for a PanOCT analysis [50] with the standard parameters of PanOCT (identity of 20 % and e-value cut-off below 1e-05).
To analyze the genetic repertoire of ‘Ca. P. phoenicium’ draft genome, tblastn analyses (low complexity filter off, e-15, minimal identity 25 %) were carried out to compare how many genes (coding sequences), identified in draft genome of AlmWB phytoplasma, are present also in the complete or draft genome of other phytoplasmas available in GenBank.
Multilocus sequence typing (MLST) analyses on ‘Ca. P. phoenicium’ strains
Based on a draft genome sequence of ‘Ca. P. phoenicium’ strain SA213, genes tufB (translational elongation factor Tu, EF-Tu), groEL (chaperonine GroEL), and inmp (integral membrane protein) were selected for investigating the genetic diversity among ‘Ca. P. phoenicium’ strain populations (Table 1) by MLST analyses. For each gene, two primer pairs were designed for carrying out nested PCR reactions (Table 2). Reaction mixture contained 1.5 mM MgCl2, 0.4 μM of each primer, and 0.2 mM of each dNTP. Reaction conditions were 5 min at 94 °C, 35 cycles including 1 min at 94 °C, 1 min at 50 °C (55 °C in nested PCRs), 2 min at 72 °C, and 10 min at 72 °C. DNAs extracted from periwinkle plants infected by phytoplasma strains EY1, STOL and AY1, from almond infected by ‘Ca. P. phoenicium’ strain SA213 (subgroup 16SrIX-B), and from Picris eichioides infected by phytoplasma strain PEY (Picris eichioides yellows phytoplasma, subgroup 16SrIX-C) served as reference controls. DNA from healthy periwinkle and reaction mixture without DNA template were used as negative controls. PCR reactions and electrophoretic analyses were performed as mentioned above.
Table 2.
Primer pairs designed for amplifying tufB, groEL and inmp gene nucleotide sequences in direct (d) and nested (n) PCR reactions
| Gene | Primer | Sequence (5′-3′) | Amplicon size (nt) |
|---|---|---|---|
| tufB | fusAF1 (d) | ATCGTGGTAATGCGATTGTGG | 1431 |
| tufBR1 (d) | ACAGAACCAGCTCCAACAGTACGTCC | ||
| fusAF2 (n) | TGGTTATGCAACCACTTTACGTTC | 1320 | |
| tufBR2 (n) | TAGTGCAATAGGATGAATTAAAGTCAC | ||
| groEL | groELF1 (d) | TGATAATGCAGGCGACGGAACTAC | 1336 |
| groELR1 (d) | TCACAGCCACTACGGCAGCACCAGCTG | ||
| groELF2 (n) | ACTACTACAGCTACTGTATTAGCAC | 1284 | |
| groELR2 (n) | TAGATGCTGCAATAGAAGAAGCATTG | ||
| inmp | inmpF1 (d) | AGTAATTAATTTTCAATATTGGACTG | 668 |
| inmpR1 (d) | TCACATCATCCTCATTCATTTTTGAAGC | ||
| inmpF2 (n) | AGAAATCTTATCAGTGGTATCAGTC | 413 | |
| inmpR2 (n) | TCTTTATCTATTGTTTTATATGCCAC |
Amplicons of tufB, groEL and inmp genes from nested PCRs were sequenced, assembled, compared with the GenBank database, and used to construct phylogenetic trees as described above for 16S rRNA gene. TufB, groEL and inmp gene nucleotide sequences of representative ‘Ca. P. phoenicium’ strains were deposited in the GenBank database (Table 1).
Analysis of synonymous and non-synonymous substitutions per site and codon-based test of positive selection (dN/dS) were carried out using MEGA5 [42] for genes showing nucleotide variability among AlmWB phytoplasma strains examined.
Nucleotide sequences
This Whole Genome Shotgun project has been deposited at DDBJ/EMBL/GenBank under the accession JPSQ00000000. The version described in this paper is version JPSQ01000000. Nucleotide sequences of genes 16S rRNA, tufB, groEL, inmp and BI-1 of representative ‘Ca. P. phoenicium’ strains identified in the present study were deposited in the GenBank database at accession numbers KM275491 to KM275499, and KP640614.
Results and Discussion
16S rDNA-based identification and phylogeny of ‘Ca. P. phoenicium’ in Lebanon
Nested PCRs, carried out with primer pair F2n/R2, revealed the presence of phytoplasmas in all tested plants (almond, peach, and nectarine) showing AlmWB typical symptoms. PCR reliability was proven by DNA amplification from phytoplasma reference strains EY1, STOL, and AY1; no amplification was obtained from DNA of healthy plants and from the reaction mix devoid of DNA. BlastN analysis demonstrated that such phytoplasmas share 100 % sequence identity with the reference strain of the species ‘Ca. P. phoenicium’ (acc. no. AF515636) [10]. Sequence alignment and sequence identity calculation highlighted that ‘Ca. P. phoenicium’ strains, identified in distinct host plants from diverse geographic regions of Lebanon, carried identical 16S rDNA sequences (sequence identity 100 %). In silico RFLP (data not shown) and phylogenetic analysis (Fig. 1) confirmed the affiliation of these phytoplasma strains to the species ‘Ca. P. phoenicium’, taxonomic subgroup 16SrIX-B [14, 51]. This result highlighted the strict association between AlmWB symptoms on almond, peach, and nectarine plants and the infection by ‘Ca. P. phoenicium’ strains belonging to taxonomic subgroup 16SrIX-B and its genetic variants, reported in previous studies [16, 51]. Due to the complete 16S rDNA identity of ‘Ca. P. phoenicium’ strain populations, the strain SA213, identified in almond in South Lebanon, was selected as representative strain of the phytoplasma populations for genome sequencing.
Fig. 1.
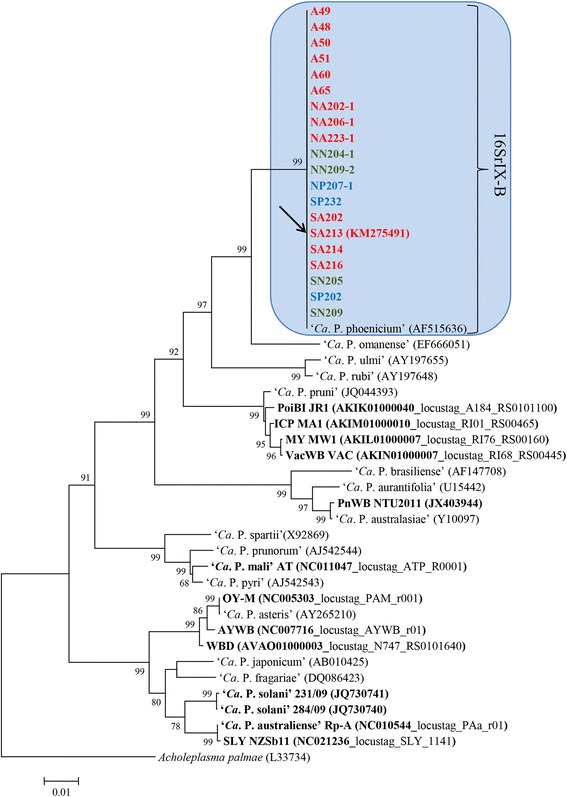
Phylogenetic tree inferred from ‘Ca. P. phoenicium’ strain nucleotide sequences of gene 16S rRNA. Minimum evolution analysis was carried out using the neighbor-joining method and bootstrap replicated 1,000 times. List of ‘Ca. P. phoenicium’ strains is reported in Table 1. ‘Ca. P. phoenicium’ strains identified in almond, peach and nectarine are written in bold red, blue and green, respectively. Names of other phytoplasmas included in the phylogenetic analysis are presented on the tree image. The GenBank accession number of each sequence is given in parenthesis. Phytoplasmas, among those whose genome was partially or fully sequenced, are indicated in bold characters: PoiBI JR1, Poinsettia branch-inducing phytoplasma strain JR1; ICP MA1, Italian clover phyllody phytoplasma strain MA1; MY MW1, Milkweed yellows phytoplasma strain MW1; VacWB VAC, Vaccinium witches’-broom phytoplasma strain VAC; PnWB NTU2011, Peanut witches’-broom phytoplasma strain NTU2011; OY-M, Onion yellows phytoplasma strain OY-M; AYWB, Aster yellows witches’-broom phytoplasma strain AYWB; WBD, Wheat blue dwarf phytoplasma; SLY NZSb11, Strawberry lethal yellows phytoplasma strain NZSb11
Draft genome overview
Illumina sequencing was performed for libraries resulting in 12,966,415 quality-passed short-reads with an average length of 145 bases (1,875 Tb read data in total). Assembly of 307,478 positive-selected reads resulted in 198 contigs with an average length of 2,054 bp comprising a total contig length of 406,850 bp. 78 contigs (39 %) with a 38-fold sequencing coverage could be assigned to the taxon ‘Ca. Phytoplasma’ using the MEGAN approach [44]. Phytoplasma assigned contigs (354 kbp, GC percentage 26 %) included 333 protein coding sequences (average length 871 bp per deduced protein), 3 tRNA and one 16SrRNA gene (Additional file 1: Table S1). An estimation of the covered gene content of ‘Ca. P. phoenicium’ draft genome was performed by comparison to the phytoplasmas protein coding gene core set [49]. Even obtained data evidenced that ‘Ca. P. phoenicium’ draft genome is far away from being complete [49], it can be shown that 81 % (238 protein coding genes) of the core set protein coding genes are also included in the ‘Ca. P. phoenicium’ draft sequence (Additional file 2: Table S2). Moreover, tblastn analyses evidenced the presence of 34 proteins (32 described as ‘hypothetical protein’) predicted to be unique for ‘Ca. P. phoenicium’ (Additional file 3: Tabel S3). Based on the presence of transmembrane domain and/or secretion signal peptide, identified by Phobius analysis, 19 and 14 AlmWB phytoplasma-unique proteins were predicted to be membrane and cytosolic proteins, respectively. One protein was predicted to be secreted (Additional file 3: Table S3). Due to the phytoplasma lifestyle (obligate intracellular parasitism), it should be interesting to carry out additional studies to determine the role and activity of predicted membrane and secreted proteins identified only in AlmWB phytoplasma. Obtained data should be useful to improve the knowledge of mechanisms of interaction between ‘Ca. P. phoenicium’ and its host(s).
Tblastn analysis results evidenced the best number of shared proteins between AlmWB phytoplasma and strains of the species ‘Ca. P. pruni’ (taxonomic group 16SrIII) underlying their close relationship, also evidenced by phylogenetic analysis (Fig. 1, Additional file 3: Table S3).
Multiple gene typing of AlmWB phytoplasma strain populations
Based on draft genome sequence, genes tufB, groEL, and inmp were selected for investigating the genetic diversity among ‘Ca. P. phoenicium’ strain populations by MLST analyses. In previous studies, nucleotide sequence analyses of tufB and groEL housekeeping genes allowed to distinguish closely related phytoplasma strains of the species ‘Ca. P. solani’ and ‘Ca. P. asteris’, showing also the relationship between strain-specific molecular markers and biological features of the phytoplasma strain(s) [52, 53]. As distinction resolution power among closely related phytoplasma strains was improved by the molecular characterization of hyper-variable genes coding membrane proteins [34–36], inmp gene, coding an integral membrane protein predicted to be unique for AlmWB phytoplasma, was chosen to determine the genetic diversity among ‘Ca. P. phoenicium’ strain populations in Lebanon. Amplicons of tufB, groEL, and inmp genes were obtained by nested PCRs from the 20 AlmWB phytoplasma-infected plants, examined in the present study, including the almond plant infected by ‘Ca. P. phoenicium’ strain SA213 (subgroup 16SrIX-B). No amplification could be generated from plants infected by other phytoplasmas of 16Sr groups I, V, XII and of subgroup IX-C, and from reaction mixtures devoid of DNA. This result demonstrated the 16SrIX-B subgroup-specificity of the primer pairs designed in the present study.
Multiple sequence analyses and sequence identity determination showed that all AlmWB phytoplasma strains shared 100 % identity of tufB and groEL gene sequence fragments. Alternatively, 15 AlmWB phytoplasma strains (including the strain SA213, selected for the draft genome sequencing) shared identical inmp gene nucleotide sequences, while 5 AlmWB phytoplasma strains exhibited a nucleotide sequence identity from 93.5 to 99.7 % with respect to the SA213 strain sequence. Interestingly, three strains exhibiting lower identities (from 93.5 to 98.3 %) were isolated from peach plants (Additional file 4: Table S4).
Phylogenetic analyses confirmed the presence of one cluster based on tufB and groEL genes, and of one main cluster including a subcluster (peach infecting strains) based on the inmp gene (Fig. 2a, b, c, respectively). Nucleotide sequence analyses revealed that AlmWB phytoplasma strain SA213 is representative of the ‘Ca. P. phoenicium’ strain populations in Lebanon. Based on phylogenetic trees, PnWB phytoplasma, ‘Ca. P. mali’ and four phytoplasmas of group 16SrIII (Poinsettia branch-inducing phytoplasma, Milkweed yellows phytoplasma, Italian clover phyllody phytoplasma, and Vaccinium witches broom phytoplasma) show the closest relationship to ‘Ca. P. phoenicium’ among the phytoplasmas whose draft or complete genome has already been published. Evidence for a close relationship with PnWB phytoplasma and group 16SrIII phytoplasmas is also supported by common genome features, as evidenced by tblastn analysis (Additional file 3: Table S3).
Fig. 2.
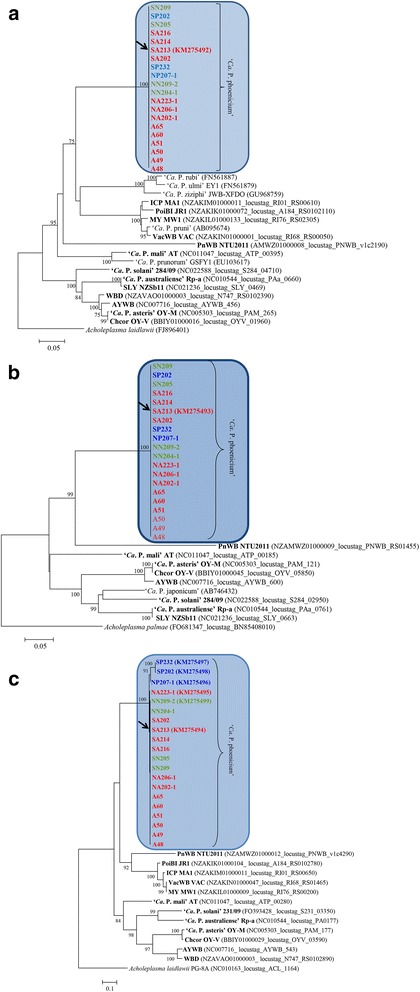
Phylogenetic trees inferred from ‘Ca. P. phoenicium’ strain nucleotide sequences of genes tufB (a), groEL (b), and inmp (c). Minimum evolution analysis was carried out using the neighbor-joining method and bootstrap replicated 1,000 times. List of ‘Ca. P. phoenicium’ strains is reported in Table 1. ‘Ca. P. phoenicium’ strains identified in almond, peach and nectarine are written in bold red, blue and green color, respectively. Names of other phytoplasmas included in phylogenetic analysis are written on the tree image. Acronyms of phytoplasma names are in the legend of Fig. 1. GenBank accession number of each sequence is given in parenthesis
Due to the role of phytoplasma membrane proteins in determining the vectoring activity of insects [36, 54] and the interaction with plant hosts [55], it is notable that within an extremely homogeneous population of AlmWB phytoplasma strains, the sole differences were revealed on integral membrane protein gene sequences. Alignments of nucleotide (358 nt) and amino acid (119 aa) sequences of inmp gene evidenced the presence of 21 nucleotide substitutions and all of them were non-synonymous (Fig. 3). Codon-based test of positive selection (dN/dS) carried out using different methods rejected the null hypothesis of strict-neutrality (dN = dS) in favor of the alternative hypothesis (dN > dS) (Table 3), showing the presence of a positive selection. Intriguingly, these preliminary data seem to indicate that AlmWB phytoplasma strains identified in peach plants are distinct from strains infecting almond and nectarine based on molecular markers within the inmp nucleotide sequences. Recently, insect species of the families Cicadellidae and Cixiidae have been reported to be capable of transmitting AlmWB phytoplasma [56, 57]. Considering this evidence, it is reasonable to hypothesize the possible implication of inmp diversity on multiple vector-specific epidemiological cycles of AlmWB phytoplasma in the diverse plant hosts. As for other phytoplasmas associated with important diseases (i.e., grapevine Flavescence dorée) [58, 59], the high genetic homogeneity within ‘Ca. P. phoenicium’ strains suggests that originally a unique strain (or a few strains) entered Lebanon, where variety in ecological niches lead to the clonal replication of this strain and its further adaptation to diverse environments, i.e. by relatively frequent mutation of the nucleotide and amino acid sequences of the membrane proteins interacting with hosts, as reported for ‘Ca. P. asteris’ strain OY-M and ‘Ca. P. solani’ [36, 60]. Molecular markers identified on the inmp gene in the present study will be employed for multiple gene sequence analyses of AlmWB phytoplasma strains identified in plant crops, insect vectors and weeds hosting the vectors, in order to increase knowledge of AlmWB disease spread and to improve possibilities for the development of sustainable strategies for its management.
Fig. 3.
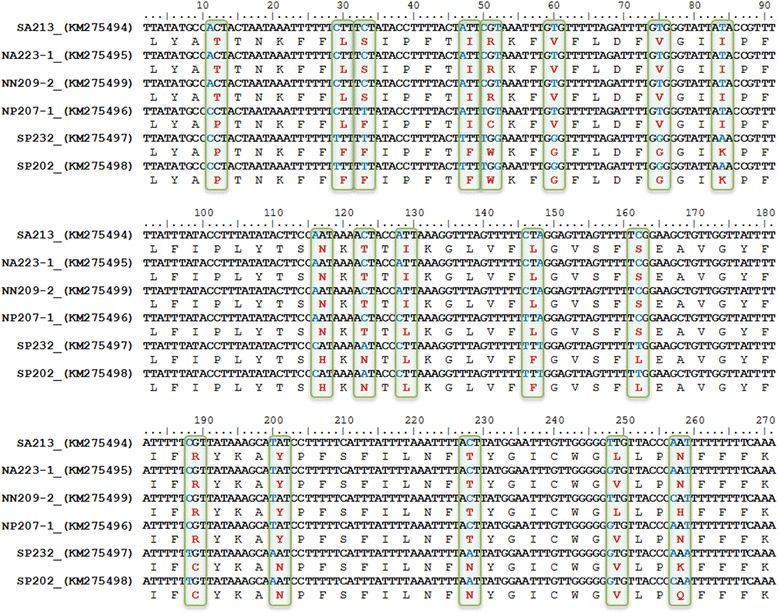
Single nucleotide polymorphisms (SNPs) and amino acid variations identified in inmp gene sequence of ‘Ca. P. phoenicium’ strains. Codons including SNPs are evidenced by green boxes. SNPs are shown in blue blod characters; amino acid variations are shown in red bold characters
Table 3.
Codon-based test of positive selection for analysis between inmp sequences of ‘Ca. P. phoenicium’ strains
| Model | Proba | Statb |
|---|---|---|
| Modified Nei-Gojobori Method (Proportion) | 0.026 | 1.961 |
| Modified Nei-Gojobori Method (Jukes-Cantor) | 0.034 | 1.836 |
| Pamilo-Bianchi-Li Method (Kimura 2-para) | 0.015 | 2.191 |
| Kumar Method (Kimura 2-para) | 0.015 | 2.195 |
a The probability of rejecting the null hypothesis of strict-neutrality (dN = dS) in favor of the alternative hypothesis (dN > dS). Values of P less than 0.05 are considered significant at the 5 % level
b The test statistic (dN - dS) is shown. dS and dN are the numbers of synonymous and nonsynonymous substitutions per site, respectively. The variance of the difference was computed using the bootstrap method (1000 replicates)
Identification of phytoplasma-host interaction key genes within ‘Ca. P. phoenicium’ draft genome
Phobius prediction analyses of transmembrane (TM) domains and signal peptides (SPs) within annotated protein sequences of ‘Ca. P. phoenicium’ draft genome resulted in the assignment of 247 cytosolic proteins (not exhibiting a TM domain or a SP), 69 membrane proteins (characterized by at least one TM domain), three cell surface proteins (characterized by SP and TM domain(s)), and 14 secreted proteins (characterized by a SP and no TM domain) (Additional file 1: Table S1). Recent studies demonstrated that phytoplasma membrane proteins, exposing extracellular domain(s)/loop(s) within the host cell cytoplasm, and proteins secreted to host cell cytoplasm play a crucial role in the interactions between phytoplasmas and hosts, determining respectively the insect vectoring activity and specificity and the re-programming of gene expression in plant hosts [54, 61–65].
One of those integral membrane proteins (AlmWB_00860) was predicted as Bax inhibitor-1 (BI-1), an inhibitor of apoptosis-promoting Bax factor (sequence homology score: 25-31 % in comparison with phytoplasmas and prokaryotes; < 10 % in comparison with eukaryotic organisms). This anti-apoptotic gene has usually been identified within the genomes of plants and animals, but has also been reported for prokaryotes [66, 67], and is known to induce a reduction of programmed cell death (PCD) [68]. It remains remarkable that the overexpression of Bax inhibitor-1 in plants decreases resistance to pathogens [69, 70]. BI-1 nucleotide sequences of AlmWB phytoplasma strains analyzed in the present study were identical to the sequence of the strain SA213 (data not shown), deposited in NCBI GenBank at accession number KP640614. BlastP and InterProScan 4 analyses revealed the presence of putative genes coding for inhibitors of Bax factor, originally assigned hypothetical proteins, in the draft and complete phytoplasma genome sequences. Alignment of BI-1 amino acid sequences of phytoplasmas, acholeplasmas, prokaryotes, plants and animals evidenced the presence of conserved trans-kingdom amino acid patterns (Fig. 4) determining the architecture of the protein [68] (Table 4). Moreover, BI-1 based phylogenetic clustering (Fig. 5) is consistent with that obtained by analyzing housekeeping genes, supporting the idea that BI-1 is an evolutionary conserved gene and might be an ancient PCD regulator of general importance for cellular homeostasis. It is intriguing to discuss the possible role of BI-1, positioned in membrane protein, in phytoplasma interaction with hosts. In plants, BI-1 is an endoplasmic reticulum (ER)-resident transmembrane protein that can interact with multiple partners to alter intracellular Ca2+ flux control and lipid dynamics. Functionally, the level of BI-1 protein has been hypothesized to have the role of a rheostat to regulate the threshold of ER-stress inducible cell death [71]. It is largely reported that phytoplasmas can colonize not only phloem sieve elements, but also companion cells [72]. Further studies should be carried out to investigate the activity of phytoplasma BI-1 inside host(s). In particular, it should be interesting to verify if, inside companion cells, phytoplasma BI-1 could act in synergy with plant ER-resident BI-1 influencing the Ca2+ homeostasis and determining the signals related with PCD. In this way, BI-1 may act as a phytoplasma fitness-increasing factor by modifying the defense response of host plants. Alternatively, BI-1 could have a internal function in homeostasis and lipid dynamics inside phytoplasma cells.
Fig. 4.

BI-1 amino acid patterns. BI-1 protein sequences of phytoplasmas, acholeplasmas, bacteria, plants and animals were retrieved from NCBI GenBank. Acronyms of phytoplasmas are in bold characters. Black arrows indicate the amino acid positions in the conserved BI-1 trans-kingdom pattern. Amino acid pattern starts in the third of seven predicted transmembrane domains and ends in the fourth transmembrane domain. Pattern positions are indicated in the column named AA position
Table 4.
Phobius prediction of BI-1 protein domains in organisms of diverse kingdoms
| BI-1 amino acid position | Phobius prediction | ||
|---|---|---|---|
| Arabidopsis thaliana | Escherichia coli | 'Ca. P. phoenicium' SA213 | |
| BAA89541 | CAA25218 | KP640614 | |
| 1–38 | 1–19 | 1–51 | Cytoplasmic |
| 39–56 | 20–41 | 52–72 | Transmembrane |
| 57–61 | 42–46 | 73–77 | Non cytoplasmic |
| 62–80 | 47–67 | 78–98 | Transmembrane |
| 81–91 | 68–73 | 99–109 | Cytoplasmic |
| 92–112 | 74–94 | 110–131 | Transmembrane |
| 113–117 | 95–105 | 132–142 | Non cytoplasmic |
| 118–139 | 106–126 | 143–165 | Transmembrane |
| 140–145 | 127–132 | 166–176 | Cytoplasmic |
| 146–169 | 133–154 | 177–199 | Transmembrane |
| 170–174 | 155–159 | 200–204 | Non cytoplasmic |
| 175–193 | 160–184 | 205–223 | Transmembrane |
| 194–213 | 185–195 | 224–243 | Cytoplasmic |
| 214–232 | 196–216 | 244–263 | Transmembrane |
| 233–247 | 217–219 | 264–275 | Non cytoplasmic |
Fig. 5.
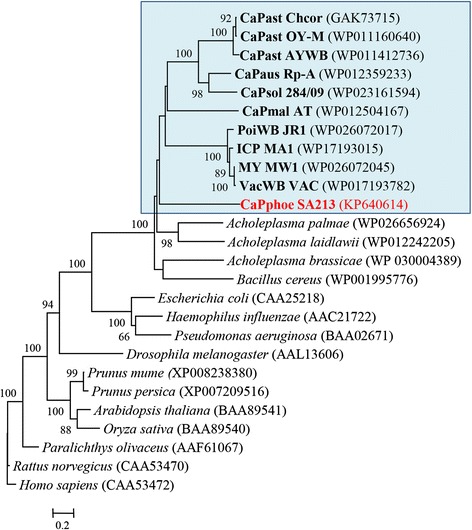
Phylogenetic tree inferred from BI-1 amino acid sequences. Minimum evolution analysis was carried out using the neighbor-joining method and bootstrap replicated 1,000 times. BI-1 protein sequences of phytoplasmas, acholeplasmas, bacteria, plants and animals were retrieved from NCBI GenBank. The GenBank accession number of each sequence is given in parenthesis. Phytoplasma strains (acronyms are in the legend of Fig. 1) are in bold characters; AlmWB phytoplasma strain SA213, identified in the present study, is in red bold characters
Two putative secreted effector proteins, containing sequence-variable mosaic (SVM) sequence (SAP05- and SAP11-like), and one predicted hemolysin, containing a cystathionine-beta-synthase (CBS) domain, were identified within the predicted secreted proteins. Effectors have been described as small proteins, positioned within potential mobile units and/or phage-related SVM genomic regions of phytoplasma genomes, able to exit the phloem (where phytoplasmas are confined), spread to plant tissues, and interact directly or indirectly with plant nuclear transcription factors inducing or repressing the expression of specific plant genes [61–63]. Effector searching among phytoplasma genomes revealed the presence of SAP11-like proteins in AlmWB phytoplasma draft genome and in the genomes of ‘Ca. P. asteris’, ‘Ca. P. mali’, ‘Ca. P. australiense’, and PnWB phytoplasma (data not shown). Furthermore, genes coding for other phage-related elements were identified within the AlmWB phytoplasma draft genome, such as phage integrase (AlmWB_00650, AlmWB_02430, AlmWB_02490), and YqaJ-like viral recombinase (AlmWB_00050, AlmWB_00770).
Further features from ‘Ca. P. phoenicium’ draft genome
Common gene sets for phytoplasmas encoding proteins for replication, DNA modification and structure, and DNA repair [73] were identified. The AlmWB phytoplasma draft genome included the entire S10-spc-alpha superoperon not encoding the gene adk, showing a spc operon with a gene order rplO-secY-map. Such gene organization within S10-spc-alpha superoperon is present also in ‘Ca. P. mali’ and PnWB phytoplasma group (‘Ca. P. australasia’), and diverges from that reported in ‘Ca. P. asteris’, ‘Ca. P. australiense’, ‘Ca. P. solani’ and Acholeplasma sp. [74]. Also the streptomycin (str) operon, carrying the genes rpsL, rpsG, fusA, and tufB, was completely identified in the AlmWB phytoplasma draft genome.
Common ABC-transporters for the ATP-dependent putative import of manganese/zinc, cobalt, spermidine/putrescine, methionine, oligopeptide and glycerol-3-phosphate were encoded in the draft genome sequence analyzed here. Furthermore, an ABC-transporter for the ATP-dependent multidrug resistance with permease function was identified in the AlmWB phytoplasma draft genome. A gene set necessary for building the Sec-dependent pathway was identified (ffh, ftsH, secA, secY, yidC) except the gene secE. As already shown for the ‘Ca. P. mali’ genome, the genes groEL and groES, coding for heat shock protein 60 family chaperones, were not in the sequence synteny groEL-amp-nadE common to other phytoplasmas [36].
With regard to the carbohydrate metabolism, the suggested conserved ATP-providing pathway of phytoplasmas has been identified encoded in all complete five phytoplasma genomes and in the draft sequences of AlmWB and PnWB phytoplasmas (this study, [73]). This pathway depends on the uptake of carboxylic acids (such as malate or oxaloacetate) mediated by the symporter MleP in phytoplasmas [65]. The draft sequence of PnWB phytoplasma also highlights the genetic repertoire for the utilization of citrate, which can be also imported by MleP (syn. CitS). The citrate lyase complex (CytEF) mediates the formation of oxaloacetate and acetate from citrate. Oxaloacetate may be decarboxylated by the common malate dehydrogenases encoded in phytoplasmas [65]. This central enzyme and MleP indicate a peculiarity of the phytoplasmas separating them from the genus Acholeplasma [75] and supporting their phylogenetic Gram-positive origin [65]. The utilization of oxaloacetate, beside malate, requires a malate dehydrogenase decarboxylating both substrates to produce pyruvate [65]. This step might be performed by NAD(P)H dependent malate dehydrogenase. In silico analysis of phytoplasmal malate dehydrogenases enabled the identification of the NADP+ binding motif (IPR016040) indicating a MaeB-like malate dehydrogenase (EC 1.1.1.38 and EC 1.1.1.40). The generated pyruvate is the entry molecule of the encoded pyruvate dehydrogenase multienzyme complex (PdhABCD) enabling the formation of acetyl-CoA, which is subsequently converted to acetyl-phosphate by the PduL-like phospotransacetylase and metabolized to acetate (AckA) releasing ATP [27, 73]. Experimental verification of the functions fulfilled by the key enzymes malate dehydrogenase (SfcA/MaeB) and phosphotransacetylase (PduL) was provided recently for ‘Ca. P. asteris’ [76].
Conclusion
In the present study, we first determined the draft genome sequence of ‘Ca. P. phoenicium’ associated with AlmWB disease in Lebanon. The results obtained revealed important insights into the genetic diversity among ‘Ca. P. phoenicium’ strains associated with the disease. Multiple gene typing analyses of ‘Ca. P. phoenicium’ strains infecting almond, peach and nectarine in Lebanon (i) revealed a substantial genetic homogeneity within the analyzed phytoplasma populations based on house-keeping gene sequence analyses, and (ii) allowed the identification of distinct AlmWB-associated phytoplasma strains from diverse host plants based on inmp (integral membrane protein) gene sequence analysis. This evidence, along with prior reports of multiple insect vectors of AlmWB phytoplasma [56, 57], suggests that AlmWB could be associated with phytoplasma strains derived from the adaptation of an original strain to diverse hosts.
Analyses of the available genome features allowed the identification of candidate determinants of pathogenicity and highlighted the coding of the conserved ATP-providing pathway of phytoplasmas, based on MleP/CitS mediated malate uptake and subsequent formation of acetate. Two putative secreted effector proteins, containing SVM sequence (SAP05- and SAP11-like), and one predicted hemolysin, containing a CBS domain, were identified within the predicted secreted proteins and can be proposed as pathogenicity determinants. Intriguingly, the identification of a putative inhibitor of apoptosis-promoting Bax factor in AlmWB phytoplasma draft genome and within genomes of other ‘Ca. Phytoplasma’ species suggested its potential role as a phytoplasma fitness-increasing factor by modification of the host-defense response. Reports of reduction of leaf cell death in phytoplasma-infected plants [77], and increased fitness of phytoplasma-infected insect vectors [78, 79] could support this hypothesis.
Acknowledgments
We thank Simon Pierce (University of Milan) for critically reading and editing the manuscript; Giulia Morlotti (University of Milan) for helpful technical assistance in phytoplasma identification and characterization; all the AVSI technicians for useful help during field trials. A sincere thank goes to Marco Perini, representative of AVSI Foundation in Lebanon, and H. E. the Minister of Agriculture of Lebanon Dr. Hussein Hajj Hassan. The authors are greatly indebted to the Ministry Officers, Mrs. Lama Haidar and Mrs. Rola Al-Achi for their constant and fruitful collaboration and to the Advisor Ministry of Agriculture Dr. Salah Hajj Hassan for his coordination.
This research was funded by the Italian Cooperation (Ministry of Foreign Affairs, http://www.cooperazioneallosviluppo.esteri.it/pdgcs/) within the project “Lotta integrata al fitoplasma delle drupacee in Libano” (Project No. L09 A0500), Comune di Milano within the project “Milano per la difesa, incremento e valorizzazione della Biodiversita 2009-2010” (http://www.comune.milano.it/portale/wps/portal/CDMHome) and the National Program for the Improvement of Olive Oil’s Quality and Actions against the Diffusion of Stone Fruit Phytoplasma (Project No. AID 9627) implemented by the Lebanese Ministry of Agriculture, Bir Hassan, Beirut, Lebanon (http://www.agriculture.gov.lb/English/Pages/Main.aspx), all coordinated by the Italian NGO AVSI Foundation (http://www.avsi.org/who-we-are/). This work was also supported by the Deutsche Forschungsgemeinschaft (http://www.dfg.de/en/) through projects KU 2679/2-1 and BU 890/21-1. The AUB team was partially supported by a grant from the Lebanese National Council for Scientific Research. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Additional files
Annotated genes within the draft genome of ‘Ca. P. phoenicium’ strain SA213. (XLS 85 kb)
PanOCT analysis of proteins identified within ‘Ca. P. phoenicium’ strain SA213 draft genome and phytoplasma core genome. (XLS 115 kb)
Tblastn comparison of ‘Ca. P. phoenicium’ strain SA213 draft genome with other phytoplasmas. (XLS 103 kb)
Inmp gene nucleotide sequence identity matrix within ‘Ca. P. phoenicium’ strains. (XLS 18 kb)
Footnotes
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
FQ designed the whole study, performed bioinformatics and MLST analyses, and drafted the manuscript; CS and MK performed draft genome assembling and annotation, and contributed to manuscript revision and results discussion; MJ, YAJ, EC, HS and MML carried out surveys on AlmWB disease in Lebanon, collected symptomatic plants and extracted total nucleic acids, and contributed to results discussion; PC and RT carried out molecular analyses on AlmWB phytoplasma strains and contributed to manuscript preparation; AA and PAB designed the whole study, revised critically the article for important improving of the contents. All authors read and approved the final manuscript.
Contributor Information
Fabio Quaglino, Phone: 0039-02-50316789, Email: fabio.quaglino@unimi.it.
Michael Kube, Email: michael.kube@agrar.hu-berlin.de.
Maan Jawhari, Email: maanjawhari@gmail.com.
Yusuf Abou-Jawdah, Email: abujawyf@aub.edu.lb.
Christin Siewert, Email: christin.siewert@agrar.hu-berlin.de.
Elia Choueiri, Email: echoueiri@lari.gov.lb.
Hana Sobh, Email: hs05@aub.edu.lb.
Paola Casati, Email: paola.casati@unimi.it.
Rosemarie Tedeschi, Email: rosemarie.tedeschi@unito.it.
Marina Molino Lova, Email: marina.molinolova@avsi.org.
Alberto Alma, Email: alberto.alma@unito.it.
Piero Attilio Bianco, Email: piero.bianco@unimi.it.
References
- 1.Lee I-M, Davis RE, Gundersen-Rindal DE. Phytoplasma: phytopathogenic mollicutes. Ann Rev Microbiol. 2000;54:221–55. doi: 10.1146/annurev.micro.54.1.221. [DOI] [PubMed] [Google Scholar]
- 2.Bertaccini A, Duduk B. Phytoplasmas and phtoplasma diseases: a review of recent research. Phytopath Mediterr. 2009;48:355–78. [Google Scholar]
- 3.Weintraub PG, Beanland L. Insect vectors of phytoplasmas. Ann Rev Entomol. 2006;51:91–111. doi: 10.1146/annurev.ento.51.110104.151039. [DOI] [PubMed] [Google Scholar]
- 4.Weisburg WG, Tully JG, Rose DL, Petzel JP, Oyaizu H, Yang D, et al. A phylogenetic analysis of the mycoplasmas: basis for their classification. J Bacteriol. 1989;171:6455–67. doi: 10.1128/jb.171.12.6455-6467.1989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Zhao Y, Davis RE, Lee I-M. Phylogenetic positions of ‘Candidatus Phytoplasma asteris’ and Spiroplasma kunkelii as inferred from multiple sets of concatenated core housekeeping proteins. Int J Syst Evol Microbiol. 2005;55:2131–41. doi: 10.1099/ijs.0.63655-0. [DOI] [PubMed] [Google Scholar]
- 6.IRPCM Phytoplasma/Spiroplasma Working Team − Phytoplasma Taxonomy Group ‘Candidatus Phytoplasma’, a taxon for the wall-less, non-helical prokaryotes that colonize plant phloem and insects. Int J Syst Evol Microbiol. 2004;54:1243–55. doi: 10.1099/ijs.0.02854-0. [DOI] [PubMed] [Google Scholar]
- 7.Duduk B, Bertaccini A. Phytoplasma classification: taxonomy based on 16S ribosomal gene, is it enough? Phytopath Mollic. 2011;1:1–13. doi: 10.5958/j.2249-4669.1.1.001. [DOI] [Google Scholar]
- 8.Zhao Y, Wei W, Lee I-M, Shao J, Suo X, Davis RE. Construction of an interactive online phytoplasma classification tool, iPhyClassifier, and its application in analysis of the peach X-disease phytoplasma group (16SrIII) Int J Syst Evol Microbiol. 2009;59:2582–93. doi: 10.1099/ijs.0.010249-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Kenyon L, Harrison NA, Ashburner GR, Boa ER, Richardson PA. Detection of a pigeon pea witches’-broom-related phytoplasma in trees of Gliricidia sepium affected by little-leaf disease in Central America. Plant Pathol. 1998;47:671–80. doi: 10.1046/j.1365-3059.1998.00281.x. [DOI] [Google Scholar]
- 10.Verdin E, Salar P, Danet J, Choueiri E, Jreijiri F, El Zammar S, et al. ‘Candidatus phytoplasma phoenicium’ sp. nov., a novel phytoplasma associated with an emerging lethal disease of almond trees in Lebanon and Iran. Int J Syst Evol Microbiol. 2003;53:833–38. doi: 10.1099/ijs.0.02453-0. [DOI] [PubMed] [Google Scholar]
- 11.Khan AJ, Al-Subhi AM, Calari A, Al-Saady NA, Bertaccini A. A new phytoplasma associated with witches’ broom of Cassia italica in Oman. B Insectol. 2007;60:269–70. [Google Scholar]
- 12.Davis RE, Dally EL, Zhao Y, Lee I-M, Jomantiene R, Detweiler AJ, et al. First report of a new subgroup 16SrIX-E (‘Candidatus Phytoplasma phoenicium’-related) phytoplasma associated with juniper witches’ broom disease in Oregon. USA Plant Pathol. 2010;59:1161. doi: 10.1111/j.1365-3059.2010.02294.x. [DOI] [Google Scholar]
- 13.Abou-Jawdah Y, Karakashian A, Sobh H, Martini M, Lee I-M. An epidemic of almond witches’-broom in Lebanon: classification and phylogenetic relationship of the associated phytoplasma. Plant Dis. 2002;86:477–84. doi: 10.1094/PDIS.2002.86.5.477. [DOI] [PubMed] [Google Scholar]
- 14.Lee I-M, Bottner-Parker KD, Zhao Y, Bertaccini A, Davis RE. Differentiation and classification of phytoplasmas in the pigeon pea witches’-broom group (16SrIX): an update based on multiple gene sequence analysis. Int J Syst Evol Microbiol. 2012;62:2279–85. doi: 10.1099/ijs.0.038273-0. [DOI] [PubMed] [Google Scholar]
- 15.Wei W, Davis RE, Lee I-M, Zhao Y. Computer-simulated RFLP analysis of 16S rRNA genes: identification of ten new phytoplasma groups. Int J Syst Evol Microbiol. 2007;57:1855–67. doi: 10.1099/ijs.0.65000-0. [DOI] [PubMed] [Google Scholar]
- 16.Molino Lova M, Quaglino F, Abou-Jawdah Y, Choueiri E, Sobh H, Casati P, et al. Identification of new 16SrIX subgroups, −F and -G, among ‘Candidatus Phytoplasma phoenicium’ strains infecting almond, peach and nectarine in Lebanon. Phytopathol Mediterr. 2011;50:273–82. [Google Scholar]
- 17.Choueiri E, Jreijiri F, Issa S, Verdin E, Bové J, Garnier M. First report of a phytoplasma disease of almond (Prunus amygdalus) in Lebanon. Plant Dis. 2001;85:802. doi: 10.1094/PDIS.2001.85.7.802C. [DOI] [PubMed] [Google Scholar]
- 18.Salehi M, Izadpanah K, Heydarnejad J. Characterization of a new almond witches’-broom phytoplasma in Iran. J Phytopathol. 2006;154:386–91. doi: 10.1111/j.1439-0434.2006.01109.x. [DOI] [Google Scholar]
- 19.Zirak L, Bahar M, Ahoonmanesh A. Characterization of phytoplasmas associated with almond diseases in Iran. J Phytopathol. 2009;157:736–41. doi: 10.1111/j.1439-0434.2009.01567.x. [DOI] [Google Scholar]
- 20.Abou-Jawdah Y, Sobh H, Akkary M. First report of Almond witches’-broom phytoplasma (‘Candidatus Phytoplasma phoenicium’) causing a severe disease on nectarine and peach trees in Lebanon. B OEPP/EPPO. 2009;39:94–8. doi: 10.1111/j.1365-2338.2009.02223.x. [DOI] [Google Scholar]
- 21.Salehi M, Haghshenas F, Khanchezar A, Esmailzadeh-Hosseini SA. Association of ‘Candidatus Phytoplasma phoenicium’ with GF-677 witches’ broom in Iran. B Insectol. 2011;64:S113–4. [Google Scholar]
- 22.Casati P, Quaglino F, Stern AR, Tedeschi R, Alma A, Bianco PA. Multiple gene analyses reveal extensive genetic diversity among ‘Candidatus Phytoplasma mali’ populations. Ann Appl Biol. 2011;158:257–66. doi: 10.1111/j.1744-7348.2011.00461.x. [DOI] [Google Scholar]
- 23.Davis RE, Zhao Y, Dally E, Lee I-M, Jomantiene R, Douglas SM. ‘Candidatus Phytoplasma pruni’, a novel taxon associated with X-disease of stone fruits, Prunus spp.: multilocus characterization based on 16S rRNA, secY, and ribosomal protein genes. Int J Syst Evol Microbiol. 2013;63:766–76. doi: 10.1099/ijs.0.041202-0. [DOI] [PubMed] [Google Scholar]
- 24.Quaglino F, Zhao Y, Casati P, Bulgari D, Bianco PA, Wei W, et al. ‘Candidatus Phytoplasma solani’, a novel taxon associated with stolbur- and bois noir-related diseases of plants. Int J Syst Evol Microbiol. 2013;63:2879–94. doi: 10.1099/ijs.0.044750-0. [DOI] [PubMed] [Google Scholar]
- 25.Oshima K, Kakizawa S, Nishigawa H, Jung HY, Wei W, Suzuki S, et al. Reductive evolution suggested from the complete genome sequence of a plant-pathogenic phytoplasma. Nat Genet. 2004;36:27–9. doi: 10.1038/ng1277. [DOI] [PubMed] [Google Scholar]
- 26.Bai X, Zhang J, Ewing A, Miller SA, Jancso Radek A, Shevchenko DV, et al. Living with genome instability: the adaptation of phytoplasmas to diverse environments of their insect and plant hosts. J Bacteriol. 2006;188:3682–96. doi: 10.1128/JB.188.10.3682-3696.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Kube M, Schneider B, Kuhl H, Dandekar T, Heitmann K, Migdoll AM, et al. The linear chromosome of the plant-pathogenic mycoplasma ‘Candidatus Phytoplasma mali’. BMC Genomics. 2008;9:306e. doi: 10.1186/1471-2164-9-306. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Tran-Nguyen LT, Kube M, Schneider B, Reinhardt R, Gibb KS. Comparative genome analysis of ‘Candidatus Phytoplasma australiense’ (subgroup tuf-Australia I; rpA) and ‘Ca. Phytoplasma asteris’ strains OY-M and AYWB. J Bacteriol. 2008;190:3979–91. doi: 10.1128/JB.01301-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Saccardo F, Martini M, Palmano S, Ermacora P, Scortichini M, Loi N, et al. Genome drafts of four phytoplasma strains of the ribosomal group 16SrIII. Microbiology. 2012;158:2805–14. doi: 10.1099/mic.0.061432-0. [DOI] [PubMed] [Google Scholar]
- 30.Andersen MT, Liefting LW, Havukkala I, Beever RE. Comparison of the complete genome sequence of two closely related isolates of ‘Candidatus Phytoplasma australiense’ reveals genome plasticity. BMC Genomics. 2013;14:529. doi: 10.1186/1471-2164-14-529. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Chung WC, Chen LL, Lo WS, Lin CP, Kuo CH. Comparative analysis of the peanut witches’-broom phytoplasma genome reveals horizontal transfer of potential mobile units and effectors. PLoS ONE. 2013;8 doi: 10.1371/journal.pone.0062770. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Mitrović J, Siewert C, Duduk B, Hecht J, Mölling K, Broecker F, et al. Generation and analysis of draft sequences of ‘stolbur’ phytoplasma from multiple displacement amplification templates. J Mol Microbiol Biotechnol. 2013;24:1–11. doi: 10.1159/000353904. [DOI] [PubMed] [Google Scholar]
- 33.Chen W, Li Y, Wang Q, Wang N, Wu Y. Comparative genome analysis of wheat blue dwarf phytoplasma, an obligate pathogen that causes wheat blue dwarf disease in China. PLoS ONE. 2014;9:e96436. doi: 10.1371/journal.pone.0096436. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Cimerman A, Arnaud G, Foissac X. Stolbur phytoplasma genome survey achieved using a suppression subtractive hybridization approach with high specificity. Appl Environ Microbiol. 2006;72:3274–83. doi: 10.1128/AEM.72.5.3274-3283.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Pacifico D, Alma A, Bagnoli B, Foissac X, Pasquini G, Tessitori M, et al. Characterization of Bois noir isolates by restriction fragment length polymorphism of a Stolbur-specific putative membrane protein gene. Phytopathology. 2009;99:711–15. doi: 10.1094/PHYTO-99-6-0711. [DOI] [PubMed] [Google Scholar]
- 36.Fabre A, Danet J-L, Foissac X. The stolbur phytoplasma antigenic membrane protein gene stamp is submitted to diversifying positive selection. Gene. 2011;472:37–41. doi: 10.1016/j.gene.2010.10.012. [DOI] [PubMed] [Google Scholar]
- 37.Seemüller E, Sule S, Kube M, Jelkmann W, Schneider B. The AAA+ ATPases and HflB/FtsH proteases of ‘Candidatus Phytoplasma mali’: phylogenetic diversity, membrane topology, and relationship to strain virulence. Mol Plant Microbe Interact. 2013;26:367–76. doi: 10.1094/MPMI-09-12-0221-R. [DOI] [PubMed] [Google Scholar]
- 38.Deng S, Hiruki C. Genetic relatedness between two nonculturable mycoplasmalike organisms revealed by nucleic acid hybridization and polymerase chain reaction. Phytopathology. 1991;81:1475–9. doi: 10.1094/Phyto-81-1475. [DOI] [Google Scholar]
- 39.Schneider B, Seemüller E, Smart CD, Kirkpatrick BC. Phylogenetic classification of plant pathogenic mycoplasma-like organisms orphytoplasmas. In: Razin S, Tully JG, editors. Molecular and diagnostic procedures in mycoplasmology. San Diego, CA, USA: Academic; 1995. pp. 369–80. [Google Scholar]
- 40.Gundersen DE, Lee I-M. Ultrasensitive detection of phytoplasmas by nested-PCR assays using two universal primer pairs. Phytopathol Mediterr. 1996;35:144–51. [Google Scholar]
- 41.Hall TA. BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl Acids Symp Ser. 1999;41:95–8. [Google Scholar]
- 42.Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S. MEGA5: Molecular Evolutionary Genetics Analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol Biol Evol. 2011;28:2731–9. doi: 10.1093/molbev/msr121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Altschul SF, Madden TL, Schaffer AA, Zhang J, Zhang Z, Miller W, et al. Gapped blast and psi-blast: a new generation of protein database search programs. Nucleic Acids Res. 1997;25:3389–402. doi: 10.1093/nar/25.17.3389. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Huson DH, Mitra S, Ruscheweyh HJ, Weber N, Schuster SC. Integrative analysis of environmental sequences using MEGAN4. Genome Res. 2011;21:1552–60. doi: 10.1101/gr.120618.111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Aziz RK, Bartels D, Best AA, DeJongh M, Disz T, Edwards RA, et al. The RAST Server: rapid annotations using subsystems technology. BMC Genomics. 2008;9:75. doi: 10.1186/1471-2164-9-75. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Rutherford K, Parkhill J, Crook J, Horsnell T, Rice P, Rajandream MA, et al. Artemis: sequence visualization and annotation. Bioinformatics. 2000;16:944–5. doi: 10.1093/bioinformatics/16.10.944. [DOI] [PubMed] [Google Scholar]
- 47.Quevillon E, Silventoinen V, Pillai S, Harte N, Mulder N, Apweiler R, et al. InterProScan: protein domains identifier. Nucleic Acids Res. 2005;33:116–20. doi: 10.1093/nar/gki442. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Käll L, Krogh A, Sonnhammer ELL. Advantages of combined transmembrane topology and signal peptide prediction - the Phobius web server. Nucleic Acids Res. 2007;35:429–32. doi: 10.1093/nar/gkm256. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Siewert C, Hess WR, Duduk B, Huettel B, Reinhardt B, Büttner C, et al. Complete genome determination and analysis of Acholeplasma oculi strain 19 L, highlighting the loss of basic genetic features in the Acholeplasmataceae. BMC Genomics. 2014;15:391. doi: 10.1186/1471-2164-15-931. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Fouts DE, Brinkac L, Beck E, Inman J, Sutton G. PanOCT: automated clustering of orthologs using conserved gene neighborhood for pan-genomic analysis of bacterial strains and closely related species. Nucleic Acids Res. 2012;40 doi: 10.1093/nar/gks757. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Abou-Jawdah Y, Dakhil H, El-Mehtar S, Lee I-M. Almond witches’-broom phytoplasma, a potential threat to almond, peach and nectarine. Can J Plant Pathol. 2003;25:28–32. doi: 10.1080/07060660309507046. [DOI] [Google Scholar]
- 52.Langer M, Maixner M. Molecular characterisation of grapevine yellows associated phytoplasmas of the stolbur-group based on RFLP-analysis of non ribosomal DNA. Vitis. 2004;43:191–9. [Google Scholar]
- 53.Mitrović J, Kakizawa S, Duduk B, Oshima K, Namba S, Bertaccini A. The groEL gene as an additional marker for finer differentiation of ‘Candidatus Phytoplasma asteris’-related strains. Ann Appl Biol. 2011;159:41–8. doi: 10.1111/j.1744-7348.2011.00472.x. [DOI] [Google Scholar]
- 54.Suzuki S, Oshima K, Kakizawa S, Arashida R, Jung H-Y, Yamaji Y, et al. Interaction between the membrane protein of a pathogen and insect microfilament complex determines insect-vector specificity. Proc Natl Acad Sci U S A. 2006;103:4252–7. doi: 10.1073/pnas.0508668103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Boonrod K, Munteanu B, Jarausch B, Jarausch W, Krczal G. An immunodominant membrane protein (Imp) of ‘Candidatus Phytoplasma mali’ binds to plant actin. Mol Plant Microbe Interact. 2012;25:889–95. doi: 10.1094/MPMI-11-11-0303. [DOI] [PubMed] [Google Scholar]
- 56.Abou-Jawdah Y, Abdel Sater A, Jawhari M, Sobh H, Abdul-Nour H, Bianco PA, et al. Asymmetrasca decedens (Cicadellidae, Typhlocybinae), a natural vector of ‘Candidatus Phytoplasma phoenicium’. Ann Appl Biol. 2014;165:395–403. doi: 10.1111/aab.12144. [DOI] [Google Scholar]
- 57.Tedeschi R, Picciau L, Quaglino F, Abou-Jawdah Y, Molino Lova M, Jawhari M, et al. A cixiid survey for natural potential vectors of ‘Candidatus Phytoplasma phoenicium’ in Lebanon and preliminary transmission trials. Ann Appl Biol. 2014;166:372–88. doi: 10.1111/aab.12188. [DOI] [Google Scholar]
- 58.Arnaud G, Malembic-Maher S, Salar P, Bonnet P, Maixner M, Marcone C, et al. Multilocus sequence typing confirms the close genetic interrelatedness of three distinct “flavescence dorée” phytoplasma strain clusters and group 16SrV phytoplasmas infecting grapevine and alder in Europe. Appl Environ Microbiol. 2007;73:4001–10. doi: 10.1128/AEM.02323-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Malembic-Maher S, Salar P, Filippin L, Carle P, Angelini E, Foissac X. Genetic diversity of European phytoplasmas of the 16SrV taxonomic group and proposal of ‘Candidatus Phytoplasma rubi’. Int J Syst Evol Microbiol. 2011;61:2129–34. doi: 10.1099/ijs.0.025411-0. [DOI] [PubMed] [Google Scholar]
- 60.Kakizawa S, Oshima K, Jung H-Y, Suzuki S, Nishigawa H, Arashida R, et al. Positive selection acting on a surface membrane protein of the plant-pathogenic phytoplasmas. J Bacteriol. 2006;188:3424–8. doi: 10.1128/JB.188.9.3424-3428.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Bai X, Correa VR, Toruño TY, Ammar D, Kamoun S, Hogenhout SA. AY-WB phytoplasma secretes a protein that targets plant cell nuclei. Mol Plant Microbe Interact. 2009;22:18–30. doi: 10.1094/MPMI-22-1-0018. [DOI] [PubMed] [Google Scholar]
- 62.Hoshi A, Oshima K, Kakizawa S, Ishii Y, Ozeki J, Hashimoto M, et al. A unique virulence factor for proliferation and dwarfism in plants identified from a phytopathogenic bacterium. Proc Natl Acad Sci U S A. 2009;106:6416–21. doi: 10.1073/pnas.0813038106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.MacLean AM, Sugio A, Makarova OV, Findlay KC, Grieve VM, Toth R, et al. Phytoplasma effector SAP54 induces indeterminate leaf-like flower development in Arabidopsis plants. Plant Physiol. 2011;157:831–41. doi: 10.1104/pp.111.181586. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Oshima K, Ishii Y, Kakizawa S, Sugawara K, Neriya Y, Himeno M, et al. Dramatic transcriptional changes in an intracellular parasite enable host switching between plant and insect. PLoS ONE. 2011;6 doi: 10.1371/journal.pone.0023242. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Siewert C, Luge T, Duduk B, Seemüller E, Büttner C, Sauer S, et al. Analysis of expressed genes of the bacterium ‘Candidatus Phytoplasma mali’ highlights key features of virulence and metabolism. PLoS ONE. 2014;9 doi: 10.1371/journal.pone.0094391. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Henke N, Lisak DA, Schneider L, Habicht J, Pergande M, Methner A. The ancient cell death suppressor BAX inhibitor-1. Cell Calcium. 2011;50:251–60. doi: 10.1016/j.ceca.2011.05.005. [DOI] [PubMed] [Google Scholar]
- 67.Wang JH, Bayles KW. Programmed cell death in plants: lessons from bacteria? Trends Plant Sci. 2013;18:133–9. doi: 10.1016/j.tplants.2012.09.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Hückelhoven R. BAX Inhibitor-1, an ancient cell death suppressor in animals and plants with prokaryotic relatives. Apoptosis. 2004;9:299–307. doi: 10.1023/B:APPT.0000025806.71000.1c. [DOI] [PubMed] [Google Scholar]
- 69.Hückelhoven R, Dechert C, Kogel KH. Overexpression of barley BAX inhibitor 1 induces breakdown of mlo-mediated penetration resistance to Blumeria graminis. Proc Natl Acad Sci U S A. 2003;100:5555–60. doi: 10.1073/pnas.0931464100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Eichmann R, Schultheiss H, Kogel KH, Hückelhoven R. The barley apoptosis suppressor homologue BAX inhibitor-1 compromises nonhost penetration resistance of barley to the inappropriate pathogen Blumeria graminis f. sp. tritici. Mol Plant Microbe Interact. 2004;17:484–90. doi: 10.1094/MPMI.2004.17.5.484. [DOI] [PubMed] [Google Scholar]
- 71.Ishikawa T, Watanabe N, Nagano M, Kawai-Yamada M, Lam E. Bax inhibitor-1: a highly conserved endoplasmic reticulum-resident cell death suppressor. Cell Death Diff. 2011;18:1271–8. doi: 10.1038/cdd.2011.59. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Christensen NM, Axelsen KB, Nicolaisen M, Schulz A. Phytoplasmas and their interactions with hosts. Trends Plant Sci. 2005;10:526–35. doi: 10.1016/j.tplants.2005.09.008. [DOI] [PubMed] [Google Scholar]
- 73.Kube M, Mitrovic J, Duduk B, Rabus R, Seemüller E. Current view on phytoplasma genomes and encoded metabolism. Sci World J. 2012;185942. [DOI] [PMC free article] [PubMed]
- 74.Lee I-M, Bottner-Parker KD, Zhao Y, Davis RE, Harrison NA. Phylogenetic analysis and delineation of phytoplasmas based on secY gene sequences. Int J Syst Evol Microbiol. 2010;60:2887–97. doi: 10.1099/ijs.0.019695-0. [DOI] [PubMed] [Google Scholar]
- 75.Kube M, Siewert C, Migdoll AM, Duduk B, Holz S, Rabus R, et al. Analysis of the complete genomes of Acholeplasma brassicae, A. palmae and A. laidlawii and their comparison to the obligate parasites from ‘Candidatus Phytoplasma’. J Mol Microbiol Biotechnol. 2014;24:19–36. doi: 10.1159/000354322. [DOI] [PubMed] [Google Scholar]
- 76.Saigo M, Golic A, Alvarez CE, Andreo CS, Hogenhout SA, Mussi MA, et al. Metabolic regulation of phytoplasma malic enzyme and phosphotransacetylase supports the use of malate as energy source in these plant-pathogens. Microbiology. 2014;doi:10.1099/mic.0.083469-0. [DOI] [PubMed]
- 77.Himeno M, Kitazawa Y, Yoshida T, Maejima K, Yamaji Y, Oshima K, et al. Purple top symptoms are associated with reduction of leaf cell death in phytoplasma-infected plants. Sci Rep. 2014;4:4111. doi: 10.1038/srep04111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Bressan A, Clair D, Sémétey O, Boudon-Padieu E. Effect of two strains of Flavescence dorée phytoplasma on the survival and fecundity of the experimental leafhopper vector Euscelidius variegatus Kirschbaum. J Invertebr Pathol. 2005;89:144–9. doi: 10.1016/j.jip.2005.03.001. [DOI] [PubMed] [Google Scholar]
- 79.Sugio A, MacLean AM, Kingdom HN, Grieve VM, Manimekalai R, Hogenhout SA. Diverse targets of phytoplasma effectors: from plant development to defence against insects. Ann Rev Phytopath. 2011;49:175–95. doi: 10.1146/annurev-phyto-072910-095323. [DOI] [PubMed] [Google Scholar]