

Abstract
Streptococcus mutans is the primary causative agent of dental caries, one of the most prevalent diseases in the United States. Previously published studies have shown that Pluronic-based tooth-binding micelles carrying hydrophobic antimicrobials are extremely effective at inhibiting S. mutans biofilm growth on hydroxyapatite (HA). Interestingly, these studies also demonstrated that non-binding micelles (NBM) carrying antimicrobial also had an inhibitory effect, leading to the hypothesis that the Pluronic micelles themselves may interact with the biofilm. To explore this potential interaction, three different S. mutans strains were each grown as biofilm in tissue culture plates, either untreated or supplemented with NBM alone (P85), NBM containing farnesol (P85F), or farnesol alone (F). In each tested S. mutans strain, biomass was significantly decreased (SNK test, p < 0.05) in the P85F and F biofilms relative to untreated biofilms. Furthermore, the P85F biofilms formed large towers containing dead cells that were not observed in the other treatment conditions. Tower formation appeared to be specific to formulated farnesol, as this phenomenon was not observed in S. mutans biofilms grown with NBM containing triclosan. Parallel CFU/ml determinations revealed that biofilm growth in the presence of P85F resulted in a 3-log reduction in viability, whereas F decreased viability by less than 1-log. Wild-type biofilms grown in the absence of sucrose or gtfBC mutant biofilms grown in the presence of sucrose did not form towers. However, increased cell killing with P85F was still observed, suggesting that cell killing is independent of tower formation. Finally, repeated treatment of pre-formed biofilms with P85F was able to elicit a 2-log reduction in viability, whereas parallel treatment with F alone only reduced viability by 0.5-log. Collectively, these results suggest that Pluronics-formulated farnesol induces alterations in biofilm architecture, presumably via interaction with the sucrose-dependent biofilm matrix, and may be a viable treatment option in the prevention and treatment of pathogenic plaque biofilms.
Introduction
Dental caries is well established as a biofilm-mediated disease, which if left untreated, can lead to tooth decay and severe patient suffering (pain, tooth loss, infection) [1, 2]. Streptococcus mutans is highly associated with the development of dental caries in humans [3, 4]. Formation of dental caries occurs through the fermentation of carbohydrates by acidogenic microorganisms, leading to the dissolution of mineral in enamel and dentin [5]. S. mutans is particularly cariogenic due to its highly acidogenic properties, ability to ferment sucrose, and production of insoluble extracellular polysaccharides (EPS), which aid in both adhesion and biofilm formation [6, 7]. One such group of exopolysaccharides are the sucrose-derived glucan polymers, a main component of the biofilm matrix, and synthesized by one of three glucosyltransferase enzymes (GtfB, GtfC, GtfD) [8]. Production of these extracellular polymers promotes adherence to the tooth surface [9–11], resistance to detachment by mechanical forces [12–14], and plays a primary role in the formation of dental caries [14, 15].
Currently established prevention and treatment of dental caries relies heavily on tooth-brushing and the strengthening of tooth enamel by fluoride. Antibiotics used in treatment of dental caries have also been pursued, although with somewhat limited effect. Chlorhexidine (CHX) gels and mouthwashes have demonstrated the most effectiveness in limiting dental plaque, but examination of CHX treatment across various subject groups showed inconclusive results [16, 17]. Testing of common antibiotics (erythromycin, penicillin, methicillin, ampicillin, and others) on a panel of 82 S. mutans strains (human, animal, and lab isolates) displayed mixed results, with many strains being resistant [18]. Additionally, most of these antibiotics are much less effective against biofilm cells and therefore cannot be confidently utilized as a primary treatment of dental caries [19–22]. Concerns regarding the emergence of antibiotic resistance among resident microbiota and the inherent recalcitrance of biofilms to antibiotics have therefore led to the exploration of alternative antimicrobial strategies for caries treatment.
Pluronics are surfactant copolymers comprised of both polyethylene oxide (PEO) and polypropylene oxide (PPO) blocks, with the balance of specific hydrophilic and lipophilic components dictating their amphiphilicity and surface activity [23]. Naturally present as unimers, Pluronics self-assemble into micelles when mixed in solutions above their critical micelle concentration (CMC) [24]. Consisting of a hydrophobic core and hydrophilic outer shell, considerable amounts (20–30% wt/vol) of water insoluble compounds can be packaged within these micelles [25]. For the purpose of inhibiting oral biofilm, our previous studies demonstrated that Pluronics could be chemically-conjugated to biomineral-binding moieties such as alendronate [26, 27], diphosphoserine peptides [28], and pyrophosphate [28], each of which displayed high binding affinity to hydroxyapatite (HA). These modified Pluronics were also successfully used to create tooth-binding micelles (TBMs), which when formulated with hydrophobic antimicrobials (triclosan, farnesol), were extremely effective at both inhibiting S. mutans biofilm formation on HA, as well as reducing viability of pre-formed biofilms [26–28]. Additionally, these studies demonstrated that unmodified Pluronic micelles carrying hydrophobic antimicrobials also had a modest inhibitory effect, suggesting that these “non-binding micelles” (NBMs) may also interact with S. mutans cells and/or biofilm. Therefore, this current study was undertaken to better understand the morphological changes and degree of cell death in S. mutans biofilms when co-cultured with NBMs containing hydrophobic antimicrobial compounds.
Materials and Methods
Bacterial strains and culturing conditions
The prototype S. mutans lab strain UA159 [29], UA159 gtfBC mutant (SAB109; this study), S. mutans BM71 [30] and S. mutans P1 [31] were maintained at -80°C in 25%-40% (vol/vol) glycerol. For each experiment, strains were freshly-streaked from frozen stocks onto Todd-Hewitt-Yeast Extract (THYE; Todd-Hewitt broth + 0.3% Yeast Extract) agar (supplemented with 1000 μg/ml kanamycin when culturing the gtfBC mutant), and incubated at 37°C, 5% CO2 for 48 hours.
Creation of a UA159 gtfBC mutant
The gtfBC-deficient strain of S. mutans UA159, SAB109, was created using a PCR ligation mutagenesis approach [32] to replace nearly all of the open reading frame (7 nucleotides downstream of the gtfB start codon through 33 nucleotides upstream of the gtfC stop codon) with a polar kanamycin marker (ΩKm). Briefly, the upstream and downstream regions flanking gtfBC were amplified with primer pairs gftB-A (5’-CTGGGCTTGTTGCTGGAATC-3’)/ gftB-BamHI-B (5’-GTTTATAACGGATCCTCT TGTCCAT-3’) and gftC-BamHI-C (5-CGTTTTAGAGGATCCCGTAATGGAT/ gftC-D (5’-CTCCTAACAGCGCTGCCATA-3’), respectively. These PCR fragments were then digested with BamHI and ligated to the kanamycin resistance cassette, and the mixture was transformed in the presence of competence stimulating peptide into S. mutans cells as previously-described [32]. Transformants were selected on BHI agar with kanamycin. Double-crossover recombination into each gene was confirmed by PCR and sequencing to ensure that no mutations were introduced into flanking genes.
Pluronic micelle preparation and antimicrobial formulation
Pluronic P85 (molecular weight = 4600) or P123 (molecular weight = 5750) were used to prepare all non-binding micelle (NBM) formulations, as indicated. P85 and P123 unimers spontaneously form into micelles when mixed in solution at concentrations exceeding their specific CMCs (6.5x10-5 M and 4.4x10-6 M, respectively) [25]. In this study, all Pluronic solutions were prepared by first mixing either P85 or P123 in deionized water to a concentration of 2% (wt/vol) (4.3x10-3 M and 3.5 x 10−3 M, respectively), and dissolved overnight at 4°C. To prepare NBMs containing farnesol, each P85 or P123 solution was then warmed in a 37°C water bath for 30 minutes before adding farnesol to a final concentration of 1% (vol/vol). These solutions were then allowed to incubate overnight at 37°C with shaking at 105 RPM to allow for complete packaging of farnesol into the P85 or P123 micelles. Each solution was then filter sterilized using a 0.2 μm filter. Solutions of 1% (wt/vol) farnesol alone (F) were prepared in 100% (vol/vol) ethanol.
To prepare triclosan NBMs (P123T), a small magnetic stir bar was placed in a 5 ml P123 solution (prepared as described above), followed by agitation at high-speed on a magnetic stir plate and stepwise addition of 100 mg (2% wt/vol) triclosan over a 1 hour period. The P123 triclosan mixture was then continuously stirred overnight at room temperature, followed by centrifugation at 12,000 rpm for 30 seconds to remove any undissolved triclosan. The supernatant (containing P123 triclosan) was removed and filter sterilized with a 0.2 micron filter prior to experimental use.
Confocal laser scanning microscopy (CLSM) of S. mutans biofilms grown in the presence of Pluronic micelles
Confocal microscopy was used to assess the effect of Pluronic micelles on S. mutans biofilm structure. In brief, overnight THYE broth cultures of each S. mutans strain were diluted to an OD600 of 0.01 in biofilm media (BM) [33] containing either 0.25% (wt/vol) glucose and 0.25% (wt/vol) sucrose (“sucrose-dependent” biofilms) or 0.5% (wt/vol) glucose (“sucrose-independent” biofilms). A 5 μl volume of each treatment stock solution (P85, P85F, P123, P123F, P123T or F alone) was then added to 1 ml of diluted culture, which was then vortexed for fifteen seconds and 200 μl transferred in duplicate to wells of a 96-well optically-clear tissue culture plate (Costar 3720). The final concentration of farnesol or triclosan in each treatment was 50 μg/ml or 36 μg/ml, respectively (each approximately 3.5× MIC). After 48 hours of static incubation at 37°C, 5% CO2, the media was removed from each well, and 200 μl BacLight LIVE/DEAD stain [Life Technologies; 0.5 μl/ml Syto-9 and 1.5 μl/mL propidium iodide (PI) in 0.85% (wt/vol) NaCl] was applied to each well and allowed to incubate for 30 minutes at room temperature. Stain was then removed and replaced with 200 μl 0.85% NaCl. CLSM was performed on an inverted Zeiss LSM Pascal equipped with an argon laser (488 nm excitation, 505–530 nm bandpass emission for Syto-9, 560 nm longpass emission for PI) at 400× magnification. Confocal z-stacks were acquired at 1 μm intervals from random fields of view located in the middle of each well. Biofilm parameters (biomass, roughness co-efficient) were quantified using COMSTAT [34].
CLSM of pre-formed S. mutans biofilms treated with P85 micelles
Overnight cultures of S. mutans UA159 were diluted to an OD600 of 0.01 in BM containing 0.25% glucose, 0.25% sucrose and transferred to an optically-clear tissue-culture plate as described above. After 24-hours of static growth at 37°C, 5% CO2, media was carefully removed from each well and then fresh BM media containing 5 μl of P85, P85F, or F solution (described above) was added to each well. BM containing P85, P85F, or F was again replaced at 48 and 72 hours growth. At 96 hours growth, biofilms were stained with LIVE/DEAD stain and imaged by CLSM as described above.
Quantification of biofilm viability
To quantify biofilm viability, a parallel series of experiments was performed as described above (treatment of sucrose-dependent or sucrose-independent biofilms at the time of inoculation, and treatment of preformed biofilms). For time-of-inoculation biofilms, the culture supernatant of each well was removed at 48 hours growth, and the adherent biofilms were scraped and resuspended in 200 μl sterile THYE, and then transferred into a sterile microcentrifuge tube. Each tube was then vortexed at high-speed for 15 seconds, followed by 1:10 serial dilution in sterile THYE, and track plating on square THYE plates [35]. After 48 hours growth in 5% CO2 and 37°C, the corresponding CFU/biofilm was calculated. CFU determination of preformed biofilm experiments was conducted as described above, except that the biofilms were harvested at 96 hours growth.
Statistical analysis
For all COMSTAT measurements and CFU/ml determinations, Sigmaplot software (version 12.5, build 12.5.0.38) was used to perform statistical analysis. Data was subjected to normality and equal variance tests, followed a one-way ANOVA and appropriate parametric or non-parametric pairwise test to detect significant differences between treatment groups.
Results
S. mutans biofilms display altered structure and increased cell death when cultured in the presence of farnesol-containing NBMs
Although previous studies have demonstrated the effectiveness of antimicrobial-formulated Pluronic micelles in inhibiting growth of S. mutans biofilms [26–28], their contribution to biofilm architecture and physiology has not been investigated. Therefore, CLSM was employed in this study to observe changes in biofilm morphology associated with growth in the presence of farnesol-containing Pluronic micelles. Altered biofilm structure was observed in all three tested S. mutans strains when grown in media containing P85F (Fig 1B, 1F and 1J) or F alone (Fig 1C, 1G and 1K), whereas biofilms grown in the presence of P85 alone (Fig 1A, 1E and 1I) were similar to untreated biofilms (Fig 1D, 1H and 1L). Specifically, growth in the presence of P85F resulted in the formation of large towers containing a centralized area of dead or damaged cells surrounded by live viable bacteria, whereas growth with F alone resulted in a less densely-packed biofilm lacking tower formation, and contained a mixture of live and dead cells throughout the biofilm. S. mutans cells also tended to form longer chains in the F alone-treated biofilms. These qualitative biofilm characteristics were quantified using COMSTAT software, with both P85F and F biofilms of all three S. mutans strains displaying significant decreases in biomass (Fig 2A) and increases in roughness co-efficient (Fig 2B) compared to untreated and NBM biofilms. To determine if tower formation was dependent on the specific Pluronic (P85) and/or antimicrobial (farnesol) used, CLSM experiments were repeated on UA159 biofilms grown in the presence of P123 formulated with farnesol (P123F) as well as P123 formulated with triclosan (P123T). These results showed that tower formation occurred in biofilms grown with P123F (Fig 3B) but not in biofilms grown with P123T (Fig 3C) or P123 alone (Fig 3A), suggesting that tower formation is specific to farnesol-containing Pluronic micelles.
Fig 1. Morphology of S. mutans biofilms grown with Pluronics-formulated farnesol.

Strains UA159 (A-D), BM71 (E-H), and P1 (I-L) static biofilms were grown in BM containing 0.25% glucose and 0.25% sucrose, either untreated or in the presence of 2% P85 (P85), 2% P85 formulated with 50 μg/ml farnesol (P85F), or 50 μg/ml farnesol alone (F). After 48 hours of growth at 37°C in 5% CO2, wells containing adherent biofilm were stained with LIVE/DEAD stain (Life Technologies), whereby live cells fluoresce green and dead/damaged cells fluoresce red. Biofilm z-stacks were acquired at 400× magnification by CLSM. Orthogonal images of each biofilm are shown, representing 12 random fields of view from 3 biological replicates acquired over 2 independent experiments. Scale bars = 20 μm.
Fig 2. Quantification of P85-formulated farnesol effects on S. mutans biofilm architecture and viability.
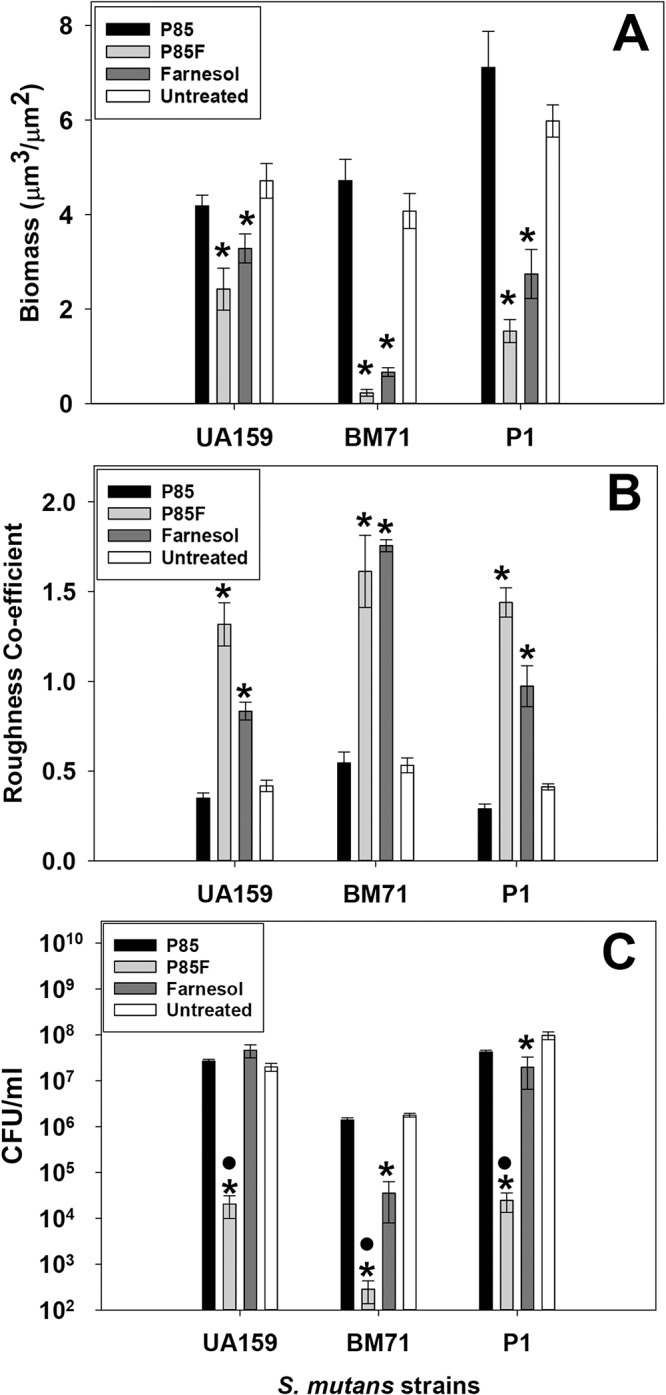
Biofilms described in Fig 1 were analyzed with COMSTAT software [34] to quantify CLSM z-stack images for biomass (A) and roughness co-efficient (B). Viability (CFU/ml) of disrupted biofilms was determined by serial dilution plating (C). For A and B, data = 12 random fields of view from 3 biological replicates acquired over 2 independent experiments. The average CFU/ml of biofilms grown in the presence of each condition (Panel C) was averaged from 3 biological replicates acquired over 2 independent experiments. Error bars = standard error of the mean (SEM). *Denotes statistically-significant difference relative to untreated condition (p < 0.05); •Denotes statistically-significant difference compared to farnesol-treated condition (p < 0.05). A: Holm-Sidak test (UA159) or Dunnett’s test (BM71 and P1); B: Dunnett's test; C: Student-Newman-Keuls (SNK) test.
Fig 3. Farnesol-specific tower formation in S. mutans static biofilm.
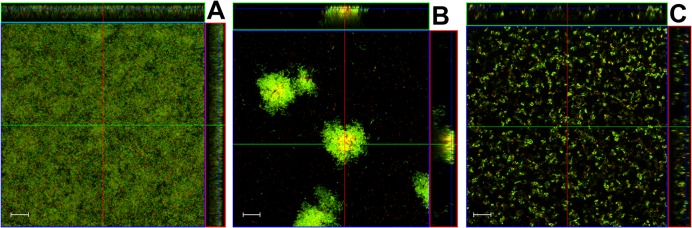
S. mutans UA159 static biofilms were grown in BM media containing 0.25% sucrose and 0.25% glucose, either with 2% P123 alone (A) or in the presence of 2% P123 formulated with farnesol (B) or triclosan (C). After 48 hours of growth, wells containing adherent biofilm were stained with LIVE/DEAD stain, and biofilm z-stacks were acquired at 400× magnification by CLSM. Representative orthogonal images of each biofilm are shown, representing 8–12 random fields of view. Scale bars = 20 μm.
Cell viability in all biofilms that were untreated or grown with P85, P85F or F alone was also measured in parallel experiments by serial dilution and CFU plating of disrupted biofilms (Fig 2C). This analysis revealed that biofilms grown in the presence of P85F underwent a 3-log reduction in viability in strains UA159 and P1, and a 4-log reduction in viability in strain BM71. By comparison, biofilms grown in F alone underwent a 1.5-log decrease in viability in BM71, 0.5-log decrease in P1, and no change in viability in UA159 (Fig 2C), indicating that the degree of biofilm killing by farnesol alone may be somewhat strain-dependent. These results demonstrate that in addition to being significantly more effective at killing S. mutans biofilm relative to farnesol alone, Pluronics-formulated farnesol induces dramatic alterations in biofilm architecture.
NBMs containing farnesol can alter biofilm structure and promote cell death in pre-formed biofilms
In addition to inhibition of biofilm formation, farnesol-formulated Pluronic micelles were tested for their effect on viability and architecture of pre-formed biofilms (Figs 4 and 5). Repeated treatment of pre-formed UA159 biofilms with either P85F or F alone (Fig 4B and 4C) over a three day period resulted in biofilms with decreased biomass (Fig 5A) and increased roughness (Fig 5B) relative to biofilms treated with P85 alone or untreated biofilms (Fig 4A and 4D). Furthermore, formation of cell death towers was also sporadically observed in pre-formed biofilms treated with P85F (Fig 4B), whereas pre-formed biofilms treated with F alone contained an evenly-distributed mixture of live and dead cells throughout the biofilm (Fig 4C). In parallel experiments, pre-formed biofilms treated with P85F experienced a 2-log decrease in cell viability relative to untreated biofilms, and treatment with F alone only reduced biofilm viability by 0.5-log (Fig 5C). These results demonstrate that tower formation and significant cell death are features common to S. mutans biofilms treated with P85F.
Fig 4. Effect of pluronics-formulated farnesol treatment on pre-formed biofilm architecture.

UA159 pre-formed biofilms were treated with P85 (A), P85F (B), F alone (C), or left untreated (D) at 24, 48, and 96 hours growth in 5% CO2 at 37°C. After 96 hours, the supernatant was removed and adherent biofilms were stained with LIVE/DEAD stain. Biofilm z-stacks of 1 μm cross-sectional images were acquired at 400× magnification by confocal laser scanning microscopy (CLSM). Orthogonal images of each biofilm are shown, representing 8 fields of view acquired from 2 biological replicates. Scale bars = 20 μm.
Fig 5. Quantification of P85-formulated farnesol effects on pre-formed S. mutans biofilm architecture.
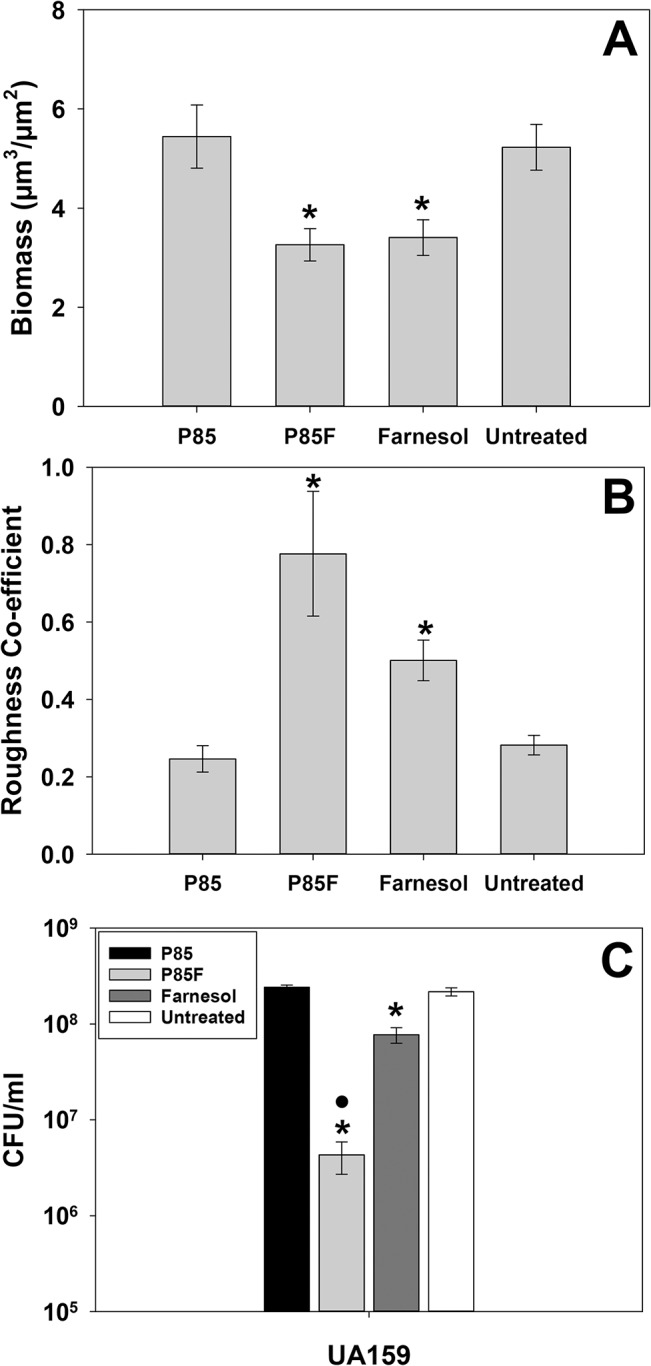
Biofilms described in Fig 4 (8 random fields of view acquired from 2 biological replicates) were analyzed with COMSTAT software [34] to quantify CLSM z-stack images for biomass (A) and roughness co-efficient (B). The average CFU/ml (n = 3 biological replicates) of biofilms grown in the presence of each condition was also determined in a parallel experiment (C). Error bars = SEM. *Indicates statistical significance compared to untreated control •Denotes statistically-significant difference compared to farnesol-treated condition (p < 0.05; SNK test).
Tower formation is dependent on the presence of sucrose-dependent biofilm matrix
S. mutans produces an EPS biofilm matrix primarily composed of a glucan polymer when grown in the presence of sucrose, and production of this sucrose-dependent matrix is dependent on GTF enzymes [36–38]. To determine if formation of towers and/or increased cell death of P85F treated S. mutans biofilms was dependent on production of the sucrose-dependent biofilm matrix, UA159 biofilms were grown in media lacking sucrose in the presence of NBM, P85F, and F. Although formation of “death towers” was not observed in these P85F sucrose-independent biofilms (Fig 6B), they displayed decreased biomass (Fig 7A) and increased roughness (Fig 7B) relative to untreated biofilms (Fig 6D), biofilms treated with P85 (Fig 6A) or farnesol alone (Fig 6C). Additionally, S. mutans cells in both the P85F and F alone biofilms (Fig 6B and 6C) each displayed the characteristic increased cell chain length and patchy biofilm morphology that was observed in F alone biofilms grown in sucrose-containing media (Fig 1C).
Fig 6. Effect of P85-formulated farnesol on S. mutans sucrose-independent biofilm architecture.

UA159 static biofilms were grown in BM lacking sucrose (supplemented with 0.5% glucose) in the presence of P85 (A), P85F (B), F alone (C), or untreated (D). After 48 hours of growth, adherent biofilm was stained with LIVE/DEAD stain. Biofilm z-stacks were acquired at 400× magnification by CLSM, and orthogonal images are representative of 12 random fields of view acquired from 2 independent experiments. Scale bars = 20 μm.
Fig 7. Quantification of P85-formulated farnesol effects on sucrose-independent S. mutans biofilms.
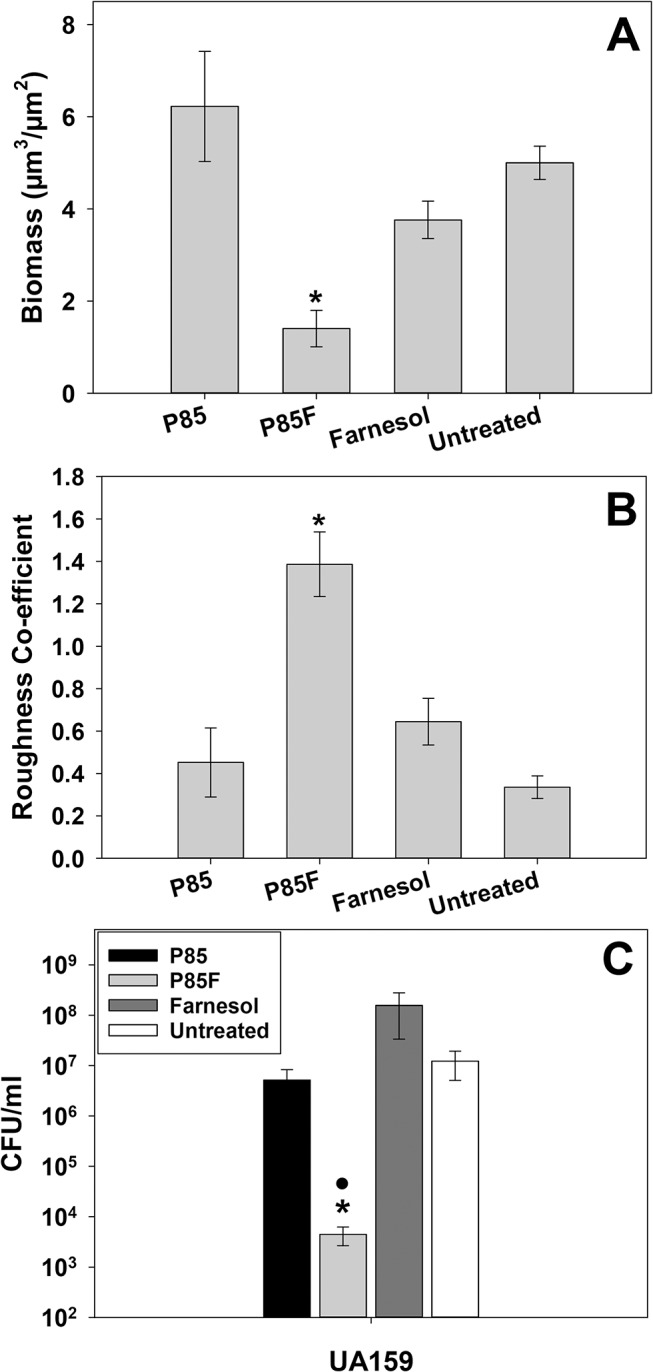
Biofilms described in Fig 6 (12 random fields of view acquired from 2 independent experiments) were analyzed with COMSTAT software [34] to quantify CLSM z-stack images for biomass (A) and roughness co-efficient (B). The average CFU/ml (n = 3 biological replicates, representative of two independent experiments) of biofilms grown in each condition was also determined in a parallel experiment (C). Error bars = SEM. *Indicates statistical significance compared to untreated control •Denotes statistically-significant difference compared to farnesol-treated condition (p < 0.05; SNK test).
To independently verify that P85F-induced tower formation does not occur in the absence of sucrose-dependent biofilm matrix, this experiment was repeated on S. mutans gtfBC mutant biofilms grown in the presence of sucrose (Fig 8). As predicted, tower formation was not observed in gtfBC mutant biofilms grown in the presence of sucrose and P85F (Fig 8B), confirming a role for the glucan biofilm matrix in promoting P85F induced tower formation. Growth of either sucrose-independent UA159 biofilms (Fig 6A) or sucrose-dependent gtfBC mutant biofilms (Fig 8A) in the presence of P85 alone did not alter their biofilm structure relative to corresponding untreated biofilms (Figs 6D and 8D).
Fig 8. Effect of gtfBC mutation on S. mutans biofilm architecture and viability in response to P85-formulated farnesol.

S. mutans gtfBC mutant was grown for 48 hours in the presence of P85 (A), P85F (B), F alone (C) or untreated (D) in BM media containing 0.25% sucrose and 0.25% glucose. Wells containing adherent biofilm were stained with LIVE/DEAD stain. Biofilm z-stacks of 1 μm cross-sectional images were acquired at 400× magnification by CLSM. Representative orthogonal images of each biofilm are shown. Scale bars = 20 μm. In a parallel experiment (n = 3 biological replicates), the average CFU/ml of each biofilm was also determined (E). All data are representative of 2 independent experiments. Error bars = SEM. *Denotes statistically-significant difference relative to untreated condition (p < 0.05), SNK Test.
Surprisingly, P85F was still able to induce a 3-log reduction in UA159 biofilm viability in media lacking sucrose, whereas treatment with F or P85 alone had no effect (Fig 7C). These results are nearly-identical to the viability patterns observed for UA159 biofilms grown in media containing sucrose (Fig 2C). Similar patterns of viability were also observed in S. mutans gtfBC mutant biofilms grown in media containing sucrose (Fig 8E), where P85F and F alone biofilms experienced 3-log and 1-log reductions in viability, respectively, compared to untreated biofilms. Collectively, these results suggest that although tower formation is dependent on S. mutans production of a sucrose-dependent biofilm matrix, the formation of towers is not required for increased killing of S. mutans biofilms by P85F.
Discussion
Pluronic copolymers are FDA-approved and have been used to efficiently formulate various drugs for their delivery to the stomach [39], brain [40, 41], liver [42], and various tumor cells [43–47]. Additionally, Pluronics conjugated to alendronate, pyrophosphate, and phosphoserine have shown great promise as tooth-targeting systems when used in the micellar formulation of hydrophobic antimicrobial compounds [26–28]. These seminal in vitro studies demonstrated that tooth-binding Pluronics micelles loaded with either triclosan or farnesol were much more effective at inhibiting S. mutans biofilm growth as well as killing pre-formed S. mutans biofilms grown on HA discs, compared to treatment with each respective free antimicrobial. More recently, this drug delivery principle has been elaborated by others to the use of nanoparticles consisting of a cationic corona and a hydrophobic and pH-responsive core, which exhibited high-affinity targeted binding to the tooth surface, pellicle, and biofilm EPS [48]. When applied topically to the tooth surface twice-daily, these farnesol-loaded nanoparticles were shown to significantly reduce the number and severity of carious lesions in a rat model of S. mutans-initiated dental caries, relative to treatment with farnesol alone [48]. Up to this point, the anti-caries properties of unformulated farnesol had demonstrated very limited effectiveness against in vitro and in vivo S. mutans biofilms [49, 50], likely due to the poor solubility of farnesol in aqueous solution and limited retention on the tooth surface.
The above-mentioned studies clearly illustrate the powerful translational potential of targeted antimicrobial delivery as an effective treatment strategy against dental caries. However, a challenge to eventual clinical approval of these delivery systems is validation of the safety profiles of newly developed formulation excipients. For example, although alendronate is used clinically in the treatment of osteoporosis [51, 52], there is concern regarding the potential long-term impact of bisphosphonates on bone resorptive activity and its rare association with osteonecrosis of the jaw [53, 54]. Therefore, use of GRAS (Generally Regarded as Safe) excipients that have already been approved for use in humans (including unmodified Pluronic copolymers) as antimicrobial delivery vehicles may represent a relatively streamlined approach for caries treatment. In this respect, the results reported in this manuscript, along with previously-published observations by our group [26–28] suggest that unmodified Pluronics also show great potential as alternative excipients. P85-formulated farnesol was found to both significantly inhibit biofilm growth when added to culture at time of inoculation, as well as efficiently kill established S. mutans biofilms (Figs 2 and 5). Furthermore, P85-formulated farnesol was able to effectively inhibit biofilm growth of three different S. mutans strains, suggesting that use of this treatment should be effective against the wide variety of genetically-diverse S. mutans isolates encountered in the human oral cavity.
Farnesol was chosen as the antimicrobial focus in these studies because (1) it has already been approved for use as a fragrance ingredient in topical cosmetics, shampoos, soaps, and perfumes [55]; (2) it is a naturally-occurring isoprenyl alcohol product found in propolis (a beehive product), fruits and vegetables [56]; (3) it has been shown to exhibit low toxicity in mice and rats [55, 57, 58]; and (4) farnesol has been previously shown to act as a membrane-active antimicrobial with significant inhibitory activity against S. mutans planktonic cultures [59, 60]. Farnesol has been shown to inhibit growth of multiple bacterial species including Staphylococcus aureus [61, 62], S. epidermidis [63], and S. mutans [49, 60], by targeting the membrane and increasing hydrogen permeability into the cell [59]. As well, Candida albicans synthesizes farnesol as a quorum sensing molecule [64–66] that can trigger apoptosis of this organism when present at high levels [67–69]. Interestingly, our results demonstrate that treatment of S. mutans biofilms with Pluronics-formulated farnesol results in “death” tower formation. This phenomenon was independent of the type of Pluronic used, but was specific to formulated farnesol (i.e. Pluronics-formulated triclosan did not induce tower formation). This suggests that Pluronics-formulated farnesol interacts with the S. mutans biofilm in a manner distinct from farnesol alone. Although the specific molecular mechanism behind this interaction was not identified in this current study, our data suggest that the sucrose-dependent biofilm EPS is likely involved (Figs 6and 8).
Farnesol alone also induced alteration of S. mutans biofilm structure in this study, whereby cells formed longer chains relative to untreated biofilms. Farnesol treatment of other microbial biofilms has been shown to induce changes in biofilm development and/or EPS production. For example, exposure of pre-formed S. epidermidis biofilms to high levels (300 μM) of farnesol resulted in significantly increased EPS production, reduced biofilm thickness, and altered biofilm structure with increased numbers of dead/damaged cells [70, 71]. A similar result was reported for S. aureus, where growth in the presence of farnesol had an inhibitory effect on biofilm formation and viability [72, 73]. At high concentrations (300 μM), farnesol has also been shown to prevent germination of adherent C. albicans cells, whereas at lower farnesol concentrations, these biofilms displayed increased density and were comprised of a mixture of yeast cells, pseudohyphae, and true hyphae [74].
The results reported in this study have also demonstrated that the killing efficacy of Pluronics-formulated farnesol is unrelated to the tower formation that occurs in S. mutans sucrose-dependent biofilms. Specifically, sucrose-independent wild-type biofilms grown in the presence of P85F did not form towers but still displayed a 3-log reduction in viability compared to the untreated control (Figs 6 and 7). Likewise, gtfBC mutant biofilms grown in the presence of sucrose and P85F also did not form towers but displayed a 3-log reduction in viability (Fig 8). Dissection of the cellular changes that precede tower formation and cell death in these biofilms will be the subject of future study in our lab, in addition to determining if tower formation affects the interaction of S. mutans with other bacterial members of dental plaque. This information will be critical in pursuing the use of unmodified Pluronics formulated with farnesol as a viable strategy in the treatment and prevention of dental caries.
Acknowledgments
The authors thank Dr. Dennis Cvitkovitch (University of Toronto) for providing S. mutans BM71, and Dr. Kazuhiko Nakano (Osaka University) for providing S. mutans strain P1.
Data Availability
All relevant data are within the paper.
Funding Statement
This work was supported by University of Florida-University Scholars undergraduate research fellowship (http://scholars.ufl.edu/) to ABM, and in part by National Institutes of Health grants R03 DE019179 (KCR), R01 DE13239 (RAB), and a “Proof of Concept” grant award from University of Nebraska Foundation (https://nufoundation.org/unmc) (DW).
References
- 1. Tanzer JM. Dental caries is a transmissible infectious disease: the Keyes and Fitzgerald revolution. J Dent Res. 1995;74(9):1536–42. . [DOI] [PubMed] [Google Scholar]
- 2. Petersen PE. The World Oral Health Report 2003: continuous improvement of oral health in the 21st century—the approach of the WHO Global Oral Health Programme. Community Dent Oral Epidemiol. 2003;31 Suppl 1:3–23. . [DOI] [PubMed] [Google Scholar]
- 3. Fitzgerald RJ, Keyes PH. Demonstration of the etiologic role of streptococci in experimental caries in the hamster. J Am Dent Assoc. 1960;61:9–19. . [DOI] [PubMed] [Google Scholar]
- 4. Loesche WJ. Role of Streptococcus mutans in human dental decay. Microbiological Reviews. 1986;50(4):353–80. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Arends J, Christoffersen J. The nature of early caries lesions in enamel. J Dent Res. 1986;65(1):2–11. . [DOI] [PubMed] [Google Scholar]
- 6. Shen S, Samaranayake LP, Yip HK. In vitro growth, acidogenicity and cariogenicity of predominant human root caries flora. J Dent. 2004;32(8):667–78. 10.1016/j.jdent.2004.07.002 . [DOI] [PubMed] [Google Scholar]
- 7. Shaw JH. Causes and control of dental caries. N Engl J Med. 1987;317(16):996–1004. 10.1056/NEJM198710153171605 . [DOI] [PubMed] [Google Scholar]
- 8. Monchois V, Willemot RM, Monsan P. Glucansucrases: mechanism of action and structure-function relationships. FEMS Microbiol Rev. 1999;23(2):131–51. . [DOI] [PubMed] [Google Scholar]
- 9. Vacca-Smith AM, Venkitaraman AR, Quivey RG Jr., Bowen WH. Interactions of streptococcal glucosyltransferases with alpha-amylase and starch on the surface of saliva-coated hydroxyapatite. Arch Oral Biol. 41 England1996. p. 291–8. [DOI] [PubMed] [Google Scholar]
- 10. Vacca-Smith AM, Bowen WH. Binding properties of streptococcal glucosyltransferases for hydroxyapatite, saliva-coated hydroxyapatite, and bacterial surfaces. Arch Oral Biol. 43 England1998. p. 103–10. [DOI] [PubMed] [Google Scholar]
- 11. Schilling KM, Bowen WH. Glucans synthesized in situ in experimental salivary pellicle function as specific binding sites for Streptococcus mutans . Infect Immun. 1992;60(1):284–95. Epub 1992/01/01. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Cross SE, Kreth J, Zhu L, Sullivan R, Shi W, Qi F, et al. Nanomechanical properties of glucans and associated cell-surface adhesion of Streptococcus mutans probed by atomic force microscopy under in situ conditions. Microbiology. 153 England 2007. p. 3124–32. [DOI] [PubMed] [Google Scholar]
- 13. Klapper I, Rupp CJ, Cargo R, Purvedorj B, Stoodley P. Viscoelastic fluid description of bacterial biofilm material properties. Biotechnol Bioeng. 2002;80(3):289–96. Epub 2002/09/13. 10.1002/bit.10376 . [DOI] [PubMed] [Google Scholar]
- 14. Korstgens V, Flemming HC, Wingender J, Borchard W. Uniaxial compression measurement device for investigation of the mechanical stability of biofilms. Journal of microbiological methods. 46 Netherlands2001. p. 9–17. [DOI] [PubMed] [Google Scholar]
- 15. Hamada S, Koga T, Ooshima T. Virulence factors of Streptococcus mutans and dental caries prevention. J Dent Res. 1984;63(3):407–11. Epub 1984/03/01. . [DOI] [PubMed] [Google Scholar]
- 16. Twetman S. Antimicrobials in future caries control? A review with special reference to chlorhexidine treatment. Caries Res. 2004;38(3):223–9. 10.1159/000077758 . [DOI] [PubMed] [Google Scholar]
- 17. Emilson CG. Potential efficacy of chlorhexidine against mutans streptococci and human dental caries. J Dent Res. 1994;73(3):682–91. . [DOI] [PubMed] [Google Scholar]
- 18. Little WA, Thomson LA, Bowen WH. Antibiotic susceptibility of Streptococcus mutans: comparison of serotype profiles. Antimicrob Agents Chemother. 1979;15(3):440–3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Liu J, Ling JQ, Zhang K, Wu CD. Physiological properties of Streptococcus mutans UA159 biofilm-detached cells. FEMS Microbiol Lett. 2013;340(1):11–8. 10.1111/1574-6968.12066 . [DOI] [PubMed] [Google Scholar]
- 20. Nguyen PT, Baldeck JD, Olsson J, Marquis RE. Antimicrobial actions of benzimidazoles against oral streptococci. Oral Microbiol Immunol. 2005;20(2):93–100. 10.1111/j.1399-302X.2004.00197.x . [DOI] [PubMed] [Google Scholar]
- 21. Tang ZS, Zhu M, Liu Z. [Susceptibility of Streptococcus mutans biofilm to antimicrobial agents]. Zhonghua Kou Qiang Yi Xue Za Zhi. 2006;41(5):266–8. Epub 2006/06/21. . [PubMed] [Google Scholar]
- 22. Olson ME, Ceri H, Morck DW, Buret AG, Read RR. Biofilm bacteria: formation and comparative susceptibility to antibiotics. Can J Vet Res. 2002;66(2):86–92. [PMC free article] [PubMed] [Google Scholar]
- 23. Housley L, Anderson T, Sontag N, Han SH, Britt DW, Anderson AJ. Pluronics' influence on pseudomonad biofilm and phenazine production. FEMS Microbiol Lett. 2009;293(1):148–53. Epub 2009/02/26. FML1528 [pii] 10.1111/j.1574-6968.2009.01528.x . [DOI] [PubMed] [Google Scholar]
- 24. Batrakova EV, Kabanov AV. Pluronic block copolymers: evolution of drug delivery concept from inert nanocarriers to biological response modifiers. J Control Release. 2008;130(2):98–106. Epub 2008/06/07. S0168-3659(08)00221-6 [pii] 10.1016/j.jconrel.2008.04.013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Kozlov MY, Melik-Nubarov NS, Batrakova EV, Kabanov AV. Relationship between Pluronic Block Copolymer Structure, Critical Micellization Concentration and Partitioning Coefficients of Low Molecular Mass Solutes. Macromolecules. 2000;33(9):3305–13. 10.1021/ma991634x [DOI] [Google Scholar]
- 26. Chen F, Liu XM, Rice KC, Li X, Yu F, Reinhardt RA, et al. Tooth-binding micelles for dental caries prevention. Antimicrob Agents Chemother. 2009;53(11):4898–902. Epub 2009/08/26. AAC.00387-09 [pii] 10.1128/AAC.00387-09 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Chen F, Rice KC, Liu XM, Reinhardt RA, Bayles KW, Wang D. Triclosan-loaded tooth-binding micelles for prevention and treatment of dental biofilm. Pharm Res. 2010;27(11):2356–64. Epub 2010/04/14. 10.1007/s11095-010-0119-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Chen F, Jia Z, Rice KC, Reinhardt RA, Bayles KW, Wang D. The development of dentotropic micelles with biodegradable tooth-binding moieties. Pharm Res. 2013;30(11):2808–17. 10.1007/s11095-013-1105-5 . [DOI] [PubMed] [Google Scholar]
- 29. Murchison HH, Barrett JF, Cardineau GA, Curtiss R 3rd. Transformation of Streptococcus mutans with chromosomal and shuttle plasmid (pYA629) DNAs. Infect Immun. 1986;54(2):273–82. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Li YH, Bowden GH. The effect of environmental pH and fluoride from the substratum on the development of biofilms of selected oral bacteria. J Dent Res. 1994;73(10):1615–26. . [DOI] [PubMed] [Google Scholar]
- 31. Nomura R, Nakano K, Nemoto H, Fujita K, Inagaki S, Takahashi T, et al. Isolation and characterization of Streptococcus mutans in heart valve and dental plaque specimens from a patient with infective endocarditis. J Med Microbiol. 2006;55(Pt 8):1135–40. 10.1099/jmm.0.46609-0 . [DOI] [PubMed] [Google Scholar]
- 32. Lau PC, Sung CK, Lee JH, Morrison DA, Cvitkovitch DG. PCR ligation mutagenesis in transformable streptococci: application and efficiency. J Microbiol Methods. 2002;49(2):193–205. . [DOI] [PubMed] [Google Scholar]
- 33. Biswas I, Drake L, Biswas S. Regulation of gbpC expression in Streptococcus mutans . J Bacteriol. 2007;189(18):6521–31. 10.1128/JB.00825-07 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Heydorn A, Nielsen AT, Hentzer M, Sternberg C, Givskov M, Ersboll BK, et al. Quantification of biofilm structures by the novel computer program COMSTAT. Microbiology. 2000;146 (Pt 10):2395–407. . [DOI] [PubMed] [Google Scholar]
- 35. Jett BD, Hatter KL, Huycke MM, Gilmore MS. Simplified agar plate method for quantifying viable bacteria. BioTechniques. 1997;23(4):648–50. . [DOI] [PubMed] [Google Scholar]
- 36. Aoki H, Shiroza T, Hayakawa M, Sato S, Kuramitsu HK. Cloning of a Streptococcus mutans glucosyltransferase gene coding for insoluble glucan synthesis. Infect Immun. 1986;53(3):587–94. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Pucci MJ, Jones KR, Kuramitsu HK, Macrina FL. Molecular cloning and characterization of the glucosyltransferase C gene (gtfC) from Streptococcus mutans LM7. Infect Immun. 1987;55(9):2176–82. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Yamashita Y, Bowen WH, Burne RA, Kuramitsu HK. Role of the Streptococcus mutans gtf genes in caries induction in the specific-pathogen-free rat model. Infect Immun. 1993;61(9):3811–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Jagdale SC, Kamble SB, Kuchekar BS, Chabukswar AR. Design and evaluation of polyox and pluronic controlled gastroretentive delivery of troxipide. Journal of drug delivery. 2014;2014:804616 10.1155/2014/804616 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Liu JS, Wang JH, Zhou J, Tang XH, Xu L, Shen T, et al. Enhanced brain delivery of lamotrigine with Pluronic((R)) P123-based nanocarrier. Int J Nanomedicine. 2014;9:3923–35. 10.2147/IJN.S62263 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Kim JY, Choi WI, Kim YH, Tae G. Brain-targeted delivery of protein using chitosan- and RVG peptide-conjugated, pluronic-based nano-carrier. Biomaterials. 2013;34(4):1170–8. 10.1016/j.biomaterials.2012.09.047 . [DOI] [PubMed] [Google Scholar]
- 42. Li X, Huang Y, Chen X, Zhou Y, Zhang Y, Li P, et al. Self-assembly and characterization of Pluronic P105 micelles for liver-targeted delivery of silybin. J Drug Target. 2009;17(10):739–50. 10.3109/10611860903062062 . [DOI] [PubMed] [Google Scholar]
- 43. Patra CR, Bhattacharya R, Wang E, Katarya A, Lau JS, Dutta S, et al. Targeted delivery of gemcitabine to pancreatic adenocarcinoma using cetuximab as a targeting agent. Cancer Res. 2008;68(6):1970–8. 10.1158/0008-5472.CAN-07-6102 . [DOI] [PubMed] [Google Scholar]
- 44. Wang Y, Yu L, Han L, Sha X, Fang X. Difunctional Pluronic copolymer micelles for paclitaxel delivery: synergistic effect of folate-mediated targeting and Pluronic-mediated overcoming multidrug resistance in tumor cell lines. Int J Pharm. 2007;337(1–2):63–73. Epub 2007/02/10. S0378-5173(06)01101-X [pii] 10.1016/j.ijpharm.2006.12.033 . [DOI] [PubMed] [Google Scholar]
- 45. Zhou Q, Guo X, Chen T, Zhang Z, Shao S, Luo C, et al. Target-specific cellular uptake of folate-decorated biodegradable polymer micelles. J Phys Chem B. 2011;115(43):12662–70. 10.1021/jp207951e . [DOI] [PubMed] [Google Scholar]
- 46. Tavano L, Muzzalupo R, Mauro L, Pellegrino M, Ando S, Picci N. Transferrin-conjugated pluronic niosomes as a new drug delivery system for anticancer therapy. Langmuir. 2013;29(41):12638–46. 10.1021/la4021383 . [DOI] [PubMed] [Google Scholar]
- 47. Hu Z, Luo F, Pan Y, Hou C, Ren L, Chen J, et al. Arg-Gly-Asp (RGD) peptide conjugated poly(lactic acid)-poly(ethylene oxide) micelle for targeted drug delivery. J Biomed Mater Res A. 2008;85(3):797–807. 10.1002/jbm.a.31615 . [DOI] [PubMed] [Google Scholar]
- 48. Horev B, Klein MI, Hwang G, Li Y, Kim D, Koo H, et al. pH-Activated Nanoparticles for Controlled Topical Delivery of Farnesol To Disrupt Oral Biofilm Virulence. ACS nano. 2015;9(3):2390–404. 10.1021/nn507170s . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Koo H, Pearson SK, Scott-Anne K, Abranches J, Cury JA, Rosalen PL, et al. Effects of apigenin and tt-farnesol on glucosyltransferase activity, biofilm viability and caries development in rats. Oral Microbiol Immunol. 2002;17(6):337–43. Epub 2002/12/18. omi170602 [pii]. . [DOI] [PubMed] [Google Scholar]
- 50. Koo H, Schobel B, Scott-Anne K, Watson G, Bowen WH, Cury JA, et al. Apigenin and tt-farnesol with fluoride effects on S. mutans biofilms and dental caries. J Dent Res. 2005;84(11):1016–20. Epub 2005/10/26. 84/11/1016 [pii]. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. Russell RG, Xia Z, Dunford JE, Oppermann U, Kwaasi A, Hulley PA, et al. Bisphosphonates: an update on mechanisms of action and how these relate to clinical efficacy. Annals of the New York Academy of Sciences. 2007;1117:209–57. 10.1196/annals.1402.089 . [DOI] [PubMed] [Google Scholar]
- 52. Russell RG. Bisphosphonates: mode of action and pharmacology. Pediatrics. 2007;119 Suppl 2:S150–62. 10.1542/peds.2006-2023H . [DOI] [PubMed] [Google Scholar]
- 53. Edwards BJ, Gounder M, McKoy JM, Boyd I, Farrugia M, Migliorati C, et al. Pharmacovigilance and reporting oversight in US FDA fast-track process: bisphosphonates and osteonecrosis of the jaw. The Lancet Oncology. 2008;9(12):1166–72. 10.1016/S1470-2045(08)70305-X . [DOI] [PubMed] [Google Scholar]
- 54. Edwards BJ, Hellstein JW, Jacobsen PL, Kaltman S, Mariotti A, Migliorati CA, et al. Updated recommendations for managing the care of patients receiving oral bisphosphonate therapy: an advisory statement from the American Dental Association Council on Scientific Affairs. J Am Dent Assoc. 2008;139(12):1674–7. . [DOI] [PubMed] [Google Scholar]
- 55. Lapczynski A, Bhatia SP, Letizia CS, Api AM. Fragrance material review on farnesol. Food Chem Toxicol. 2008;46 Suppl 11:S149–56. Epub 2008/07/22. S0278-6915(08)00336-0 [pii] 10.1016/j.fct.2008.06.046 . [DOI] [PubMed] [Google Scholar]
- 56. Tatman D, Mo H. Volatile isoprenoid constituents of fruits, vegetables and herbs cumulatively suppress the proliferation of murine B16 melanoma and human HL-60 leukemia cells. Cancer Lett. 2002;175(2):129–39. . [DOI] [PubMed] [Google Scholar]
- 57. Horn T, Long L, Cwik M, Johnson W, Morrissey R, Kapetanovic I, et al. Subchronic oral toxicity/enzyme modulation study of farnesol in rats. The Toxicologist. 2004;78:15. [Google Scholar]
- 58. Horn TL, Long L, Cwik MJ, Morrissey RL, Kapetanovic IM, McCormick DL. Modulation of hepatic and renal drug metabolizing enzyme activities in rats by subchronic administration of farnesol. Chem Biol Interact. 2005;152(2–3):79–99. Epub 2005/04/21. S0009-2797(05)00059-1 [pii] 10.1016/j.cbi.2005.02.006 . [DOI] [PubMed] [Google Scholar]
- 59. Jeon JG, Pandit S, Xiao J, Gregoire S, Falsetta ML, Klein MI, et al. Influences of trans-trans farnesol, a membrane-targeting sesquiterpenoid, on Streptococcus mutans physiology and survival within mixed-species oral biofilms. Int J Oral Sci. 2011;3(2):98–106. Epub 2011/04/13. 10.4248/IJOS11038 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60. Koo H, Rosalen PL, Cury JA, Park YK, Bowen WH. Effects of compounds found in propolis on Streptococcus mutans growth and on glucosyltransferase activity. Antimicrob Agents Chemother. 2002;46(5):1302–9. Epub 2002/04/18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61. Inoue Y, Shiraishi A, Hada T, Hirose K, Hamashima H, Shimada J. The antibacterial effects of terpene alcohols on Staphylococcus aureus and their mode of action. FEMS Microbiol Lett. 237 Netherlands2004. p. 325–31. [DOI] [PubMed] [Google Scholar]
- 62. Jabra-Rizk MA, Meiller TF, James CE, Shirtliff ME. Effect of farnesol on Staphylococcus aureus biofilm formation and antimicrobial susceptibility. Antimicrob Agents Chemother. 50 United States2006. p. 1463–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63. Gomes F, Leite B, Teixeira P, Cerca N, Azeredo J, Oliveira R. Farnesol as antibiotics adjuvant in Staphylococcus epidermidis control in vitro. Am J Med Sci. 341(3):191–5. Epub 2010/11/26. 10.1097/MAJ.0b013e3181fcf138 . [DOI] [PubMed] [Google Scholar]
- 64. Langford ML, Atkin AL, Nickerson KW. Cellular interactions of farnesol, a quorum-sensing molecule produced by Candida albicans . Future Microbiol. 2009;4(10):1353–62. Epub 2009/12/10. 10.2217/fmb.09.98 . [DOI] [PubMed] [Google Scholar]
- 65. Decanis N, Tazi N, Correia A, Vilanova M, Rouabhia M. Farnesol, a fungal quorum-sensing molecule triggers Candida albicans morphological changes by downregulating the expression of different secreted aspartyl proteinase genes. Open Microbiol J. 5 Netherlands2011. p. 119–26. 10.2174/1874285801105010119 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66. Hall RA, Turner KJ, Chaloupka J, Cottier F, De Sordi L, Sanglard D, et al. The quorum-sensing molecules farnesol/homoserine lactone and dodecanol operate via distinct modes of action in Candida albicans . Eukaryot Cell. 10 United States2011. p. 1034–42. 10.1128/EC.05060-11 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67. Shirtliff ME, Krom BP, Meijering RA, Peters BM, Zhu J, Scheper MA, et al. Farnesol-induced apoptosis in Candida albicans . Antimicrob Agents Chemother. 2009;53(6):2392–401. Epub 2009/04/15. AAC.01551-08 [pii] 10.1128/AAC.01551-08 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68. Zhu J, Krom BP, Sanglard D, Intapa C, Dawson CC, Peters BM, et al. Farnesol-induced apoptosis in Candida albicans is mediated by Cdr1-p extrusion and depletion of intracellular glutathione. PLoS One. 6(12):e28830 Epub 2011/12/30. doi: 10.1371/journal.pone.0028830 PONE-D-11-19084 [pii]. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69. Jabra-Rizk MA, Shirtliff M, James C, Meiller T. Effect of farnesol on Candida dubliniensis biofilm formation and fluconazole resistance. FEMS Yeast Res. 2006;6(7):1063–73. Epub 2006/10/18. FYR121 [pii] 10.1111/j.1567-1364.2006.00121.x . [DOI] [PubMed] [Google Scholar]
- 70. Gomes F, Teixeira P, Cerca N, Azeredo J, Oliveira R. Effect of farnesol on structure and composition of Staphylococcus epidermidis biofilm matrix. Curr Microbiol. 2011;63(4):354–9. 10.1007/s00284-011-9984-3 . [DOI] [PubMed] [Google Scholar]
- 71. Cerca N, Gomes F, Pereira S, Teixeira P, Oliveira R. Confocal laser scanning microscopy analysis of S. epidermidis biofilms exposed to farnesol, vancomycin and rifampicin. BMC Res Notes. 2012;5:244 10.1186/1756-0500-5-244 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72. Jabra-Rizk MA, Meiller TF, James CE, Shirtliff ME. Effect of farnesol on Staphylococcus aureus biofilm formation and antimicrobial susceptibility. Antimicrob Agents Chemother. 2006;50(4):1463–9. Epub 2006/03/30. 50/4/1463 [pii] 10.1128/AAC.50.4.1463-1469.2006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73. Unnanuntana A, Bonsignore L, Shirtliff ME, Greenfield EM. The effects of farnesol on Staphylococcus aureus biofilms and osteoblasts. An in vitro study. J Bone Joint Surg Am. 2009;91(11):2683–92. Epub 2009/11/04. 91/11/2683 [pii] 10.2106/JBJS.H.01699 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74. Ramage G, Saville SP, Wickes BL, Lopez-Ribot JL. Inhibition of Candida albicans biofilm formation by farnesol, a quorum-sensing molecule. Appl Environ Microbiol. 2002;68(11):5459–63. Epub 2002/10/31. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
All relevant data are within the paper.