

Abstract
Background
Due to its cost-effectiveness and rich sugar composition, sugarcane molasses is considered to be a promising carbon source for biorefinery. However, the sugar mixture in sugarcane molasses is not consumed as efficiently as glucose in microbial fermentation due to complex interactions among their utilizing pathways, such as carbon catabolite repression (CCR). In this study, 2,3-butanediol-producing Enterobacter aerogenes was engineered to alleviate CCR and improve sugar utilization by modulating its carbon preference.
Results
The gene encoding catabolite repressor/activator (Cra) was deleted in the genome of E. aerogenes to increase the fructose consumption rate. However, the deletion mutation repressed sucrose utilization, resulting in the accumulation of sucrose in the fermentation medium. Cra regulation on expression of the scrAB operon involved in sucrose catabolism was verified by reverse transcription and real-time PCR, and the efficiency of sucrose utilization was restored by disrupting the scrR gene and overexpressing the scrAB operon. In addition, overexpression of the ptsG gene involved in glucose utilization enhanced the glucose preference among mixed sugars, which relieved glucose accumulation in fed-batch fermentation. In fed-batch fermentation using sugarcane molasses, the maximum titer of 2,3-butanediol production by the mutant reached 140.0 g/L at 54 h, which was by far the highest titer of 2,3-butanediol with E. aerogenes achieved through genetic engineering.
Conclusions
We have developed genetically engineered E. aerogenes as a 2,3-butanediol producer that efficiently utilizes sugarcane molasses. The fermentation efficiency was dramatically improved by the alleviation of CCR and modulation of carbon preference. These results offer a metabolic engineering approach for achieving highly efficient utilization of mixed sugars for the biorefinery industry.
Electronic supplementary material
The online version of this article (doi:10.1186/s13068-015-0290-3) contains supplementary material, which is available to authorized users.
Keywords: 2,3-Butanediol; Enterobacter aerogenes; Sugarcane molasses; Fed-batch fermentation; Catabolite repressor/activator; Carbon catabolite repression
Background
The development of biorefineries has attracted a great deal of interest due to increasing energy costs and environmental concerns resulting from fossil fuel utilization [1, 2]. As an example, microbial production of 2,3-butanediol has been intensively studied in the past few years due to its multiple industrial applications, including the production of synthetic rubber, plasticizers, fuel additives, and fumigants [3, 4]. In microbial fermentation for 2,3-butanediol production, the carbon source is one of the major drivers of cost. Therefore, much effort has been made to find inexpensive feedstocks, such as corncob [5], jatropha hulls [6], Jerusalem artichoke tubers [7], and molasses [8]. In general, most biomass derived from lignocellulose and waste materials contains a few mixed sugars. For example, the hydrolysis of lignocellulosic biomass produces a mixture of sugars containing glucose, xylose, and arabinose [9]. The main carbohydrate of Jerusalem artichoke tuber is inulin, which can be hydrolyzed by inulinase to fructose and glucose [10].
When provided with a mixture of different carbon sources, most microorganisms prefer to use one carbon source for the fastest growth. The presence of preferred carbon sources often prevents the utilization of secondary substrates by the regulatory mechanisms, including transcription activation or repression of certain genes concerned with the use of alternative carbon sources, which is called carbon catabolite repression (CCR) [11, 12]. In enteric bacteria, two dominant CCR mechanisms involve transcriptional regulation by cyclic AMP receptor protein (Crp) and by catabolite repressor/activator (Cra) [13]. Crp is known to regulate the genes involved in carbon metabolism, such as lactose, arabinose, mannose, glucosamine, and amino sugars operons, in response to the depletion of preferred carbon source [14, 15]. On the other hand, Cra generally regulates carbon flux through by repression of genes encoding glycolytic pathway enzymes, or by activating key genes involved in the Krebs cycle, glyoxylate shunt, gluconeogenic pathways and electron transfer [16]. After the preferred carbon source is depleted, an intermediate lag-phase is caused by a shift in the catabolic system to utilize a secondary sugar. Because the catabolic characteristic of microbial fermentation with mixed sugars results in a delayed and complicated fermentation process, alleviation of CCR of a host microorganism is very helpful for improving its fermentation efficiency [17].
Sugarcane molasses contains several mixed sugars, a dominant amount of sucrose, and similar amounts of glucose and fructose. It is considered to be a promising feedstock for biorefinery due to its rich sugar content and cost-effectiveness [18]. In our previous study, disruption of the scrR gene, a transcriptional repressor of the scr regulon for sucrose catabolism, was conducted in Enterobacter aerogenes for efficient utilization of sucrose in sugarcane molasses for 2,3-butanediol fermentation [19]. The scrR mutation increased the sucrose consumption rate significantly, resulting in 2,3-butanediol production from sugarcane molasses that was enhanced by 56.8% in batch fermentation over its parent strain. In addition, 98.7 g/L of 2,3-butanediol production was achieved at 36 h of fed-batch fermentation with molasses feeding. However, despite these advances, several obstacles remain for more efficient utilization of sugarcane molasses in fed-batch fermentation. First, the efficiency of fructose utilization in the scrR mutant was relatively reduced, which resulted in a significant amount (~30 g/L) of fructose accumulation in the medium after the fermentation period. Second, the fermentation duration of the scrR mutant with sugarcane molasses for 2,3-butanediol production was shorter by 33.3% than that with glucose, which might be caused by cell stress from the repeated catabolic shift between consumed sugars (glucose and sucrose), according to the feeding of sugarcane molasses.
Therefore, the purpose of this study was to develop a 2,3-butanediol-producing mutant that can utilize all three sugars contained in sugarcane molasses efficiently and simultaneously. As shown in Fig. 1, all sugars in molasses (glucose, fructose, and sucrose) are transported into cells by the phosphotransferase system (PTS) and then converted to intermediates, such as glucose-6-phosphate (G6P), fructose-6-phosphate (F6P), and fructose-1,6-bisphosphate (F1,6BP), involved in the preparatory phase of glycolysis [20–23]. Transcription of the fruBKA operon, encoding genes for fructose uptake and utilization, was negatively regulated by Cra, previously designated as fruR [24]. When glucose is depleted, Cra activates the metabolic pathways utilizing gluconeogenic carbon sources, such as acetate, pyruvate, glycerol, and glucogenic amino acids [25, 26].
Fig. 1.
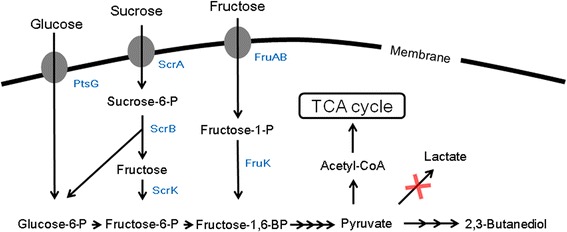
Catabolic pathway of PTS-mediated sugars contained in sugarcane molasses in E. aerogenes. Symbols represent deleted genes (red cross). ScrA EII transport protein for sucrose, ScrB sucrose-6-phosphate hydrolase, ScrK fructokinase, PtsG EIIBCGlc complex, FruA EIIBCfru complex, FruB EIIAfru component, FruK fructose-1-phosphate kinase.
In this study, deletion of the cra gene in E. aerogenes showed a much higher fructose utilization rate, but significantly retarded sucrose utilization. The reduced sucrose consumption in cra-deficient E. aerogenes was examined by reverse transcription real-time PCR. In addition, the carbon preference and efficiency were modulated by overexpression of the key genes involved in each carbon catabolism, which reduced the accumulation of certain carbon sources in fed-batch fermentation. The catabolic regulation of carbohydrates involved in sugarcane molasses and the strategies for constructing 2,3-butanediol production strain are shown in the Additional file 1.
Results and discussion
Effects of cra deletion on sugar utilization and metabolite production
To improve fructose utilization, disruption of cra was performed. An open-reading frame that contained a 95% amino acid sequence identity to Cra of Escherichia coli K-12 MG1655 was identified from the E. aerogenes genome using BLAST (Basic Local Alignment Search Tool). The deletion of cra was performed from EMY-01 (△ldhA) and EMY-68 (△ldhA △scrR), respectively, and was verified by colony PCR (data not shown). The resulting strains were named EMY-69 (△ldhA △cra) and EMY-70 (△ldhA △scrR △cra), respectively (Table 1). In our previous study, the ldhA-deficient E. aerogenes mutant (EMY-01) showed improved carbon flux and NADH availability for 2,3-butanediol production [27]. In addition, acidification of the culture medium was alleviated due to reduced lactate production. Because of these advantages, EMY-01 was used as a control strain in this study.
Table 1.
Strains and plasmids used in this study
| Strains or plasmids | Genotype or relevant characteristics | Source or reference |
|---|---|---|
| Strain | ||
| E. coli DH5α | Invitrogen | |
| E. aerogenes KCTC 2190 | Wild type | Korean Collection for Type Culture |
| EMY-01 | E. aerogenes KCTC 2190 △ldhA | [27] |
| EMY-68 | E. aerogenes KCTC 2190 △ldhA △scrR | [19] |
| EMY-69 | E. aerogenes KCTC 2190 △ldhA △cra | This study |
| EMY-70 | E. aerogenes KCTC 2190 △ldhA △scrR △cra | This study |
| EMY-70S | Plasmid-based scrAB overexpression strain of EMY-70 by introduction of pZS21MCS::scrAB | This study |
| EMY-70SP | Plasmid-based ptsG overexpression strain of EMY-70S by introduction of pZA31MCS::ptsG | This study |
| Plasmid | ||
| pKM208 | lacI, λ Red + Gam-producing vector, tac_promoter, f1_ori, Amp R | Addgene |
| pCP20 | FLP recombinase-producing vector, cI857, pSC101 ori, Amp R, Cm R | [40] |
| pKD4 | FRT flanked resistance cassette involved vector, oriRγ, Km R | [40] |
| pZA31MCS | E. coli—K. pneumoniae shuttle vector, PLtetO-1, p15A ori, CmR | Expressys |
| pZS21MCS | E. coli—K. pneumoniae shuttle vector, PLtetO-1, pSC101 ori, Km R | Expressys |
| pZA31MCS::ptsG | pZA31MCS derivative containing ptsG, PLtetO-1, p15A ori, CmR | This study |
| pZS21MCS::scrAB | pZS21MCS derivative containing scrAB, PLtetO-1, pSC101 ori, Km R | This study |
Flask cultivations of EMY-01, EMY-68, EMY-69, and EMY-70 were conducted with 80 g/L of three different carbon sources (fructose, glucose, and sucrose) for 10 h to confirm the effect of cra deletion on sugar utilization (Fig. 2; Additional file 2). The removal of scrR, a transcriptional repressor of the scr regulon for sucrose catabolism, caused considerable enhancement of sucrose utilization, but did not affect the utilization of fructose and glucose, in accordance with a previous report [19]. The disruption of cra did not affect the consumption of glucose, while significantly increasing fructose utilization by 32.9 and 39.0% in EMY-69 and EMY-70, respectively (Fig. 2a, b). The improvement in fructose utilization increased 2,3-butanediol production by 32.5 and 35.1%, respectively. Interestingly, the removal of cra repressed the utilization of sucrose significantly. The sucrose consumption of EMY-69 and EMY-70 was reduced by 58.2 and 24.6%, respectively, compared to that of their parent strains (Fig. 2c).
Fig. 2.
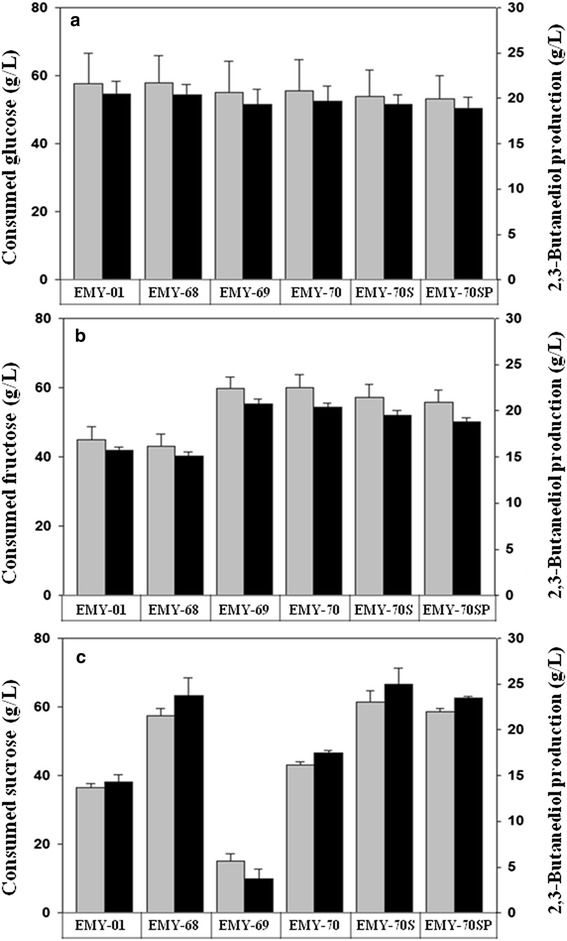
Comparison of consumed concentration of a glucose, b fructose, and c sucrose by E. aerogenes mutants in 10 h of flask cultivation. Gray bars concentration of consumed sugars; black bars 2,3-butanediol production. Error bars represent the standard deviations of three experiments.
The effect of cra mutation was also confirmed in flask cultivation with a mixture of the three sugars. As shown in Fig. 3a, b, glucose was the preferred carbon source of EMY-01 and EMY-68. After glucose was depleted, EMY-01 utilized fructose and sucrose at similar rates, while EMY-68 consumed sucrose faster than fructose due to the disruption of scrR. Meanwhile, the most preferred sugar of the cra-deficient mutants was fructose. The fructose consumption rates of EMY-69 and EMY-70 increased by 124.0 and 420.5%, respectively, compared to that of their parent strains in 10 h of flask cultivation with mixed sugars. Furthermore, a significant decrease (81.3%) in the sucrose consumption of EMY-69 was also observed with 12 h of flask cultivation with mixed carbon sources, resulting in a 24.1% reduction of 2,3-butanediol production (Fig. 3; Additional file 3).
Fig. 3.
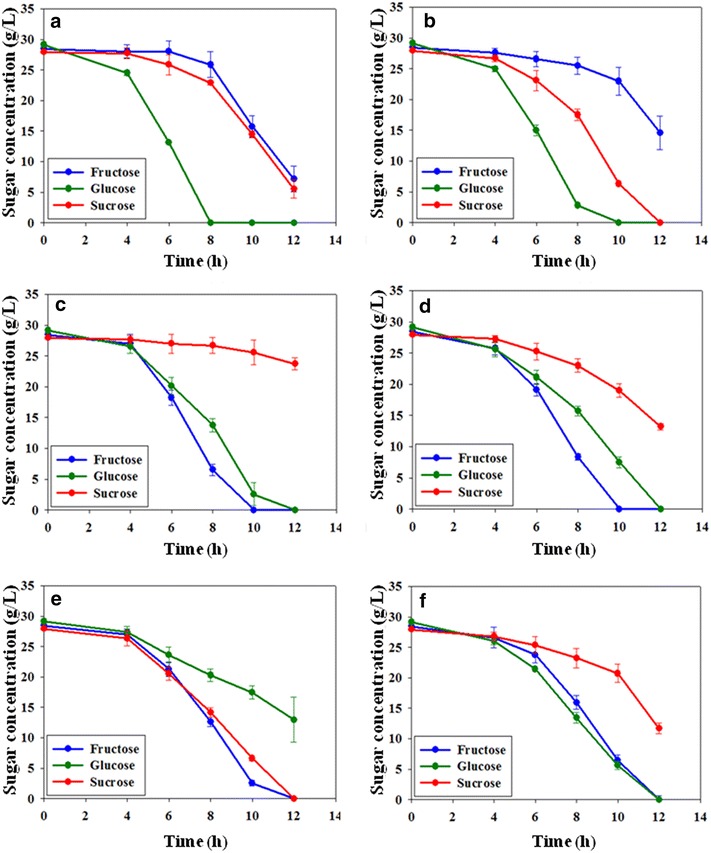
Comparison of flask cultivations of a EMY-01, b EMY-68, c EMY-69, d EMY-70, e EMY-70S, and f EMY-70SP with the consumption of sugars. Error bars represent the standard deviations of three experiments.
Transcription of the fructose operon is repressed when Cra is bound to the operator, which is located on the downstream region of the RNA polymerase binding site [24]. Whereas, derepression of the fructose operon takes place when cytoplasmic glycolytic catabolites, such as fructose-1-phosphate and fructose-1,6-bisphosphate, bind to the Cra protein, causing it to dissociate from the operator. Our results demonstrate that cra deletion improves the consumption rate of fructose significantly and removes CCR with respect to fructose utilization, which increased 2,3-butanediol production. At the same time, a significant decrease in the sucrose uptake rate was observed with deletion of Cra, indicating a significant interaction between the scr regulon and Cra. The efficient utilization of sucrose is essential for maximizing the use of sugarcane molasses, because 70% of the carbon contained in sugarcane molasses is sucrose. However, the regulation of sucrose catabolism by Cra remains unexplored in enteric bacteria.
Regulation of genes involved in sucrose catabolism in cra-deficient E. aerogenes
To observe the mechanism of Cra regulation of sucrose catabolism, reverse transcription and real-time PCR was conducted with a cra-deficient mutant, which was grown in sucrose medium. Six genes related to sucrose utilization (scrA, scrB, scrR, scrK, cra, and crp) were selected for the reverse transcription and real-time PCR assay. The transcription level of genes involved in the sucrose catabolism of EMY-01 and EMY-69 was compared in the mid-exponential growth phase (6 h), using gapA as a reference gene [28]. As shown in Fig. 4, there were no significant differences in the relative expression levels of scrR, scrK, or crp in the strains, whereas the transcription levels of scrA and scrB decreased by 32.0 and 24.9%, respectively, by disruption of the cra gene.
Fig. 4.
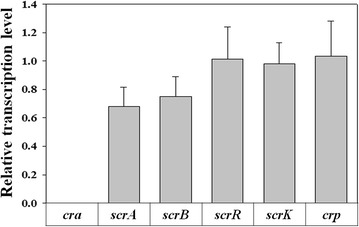
Real-time PCR results of genes involved in sucrose catabolism by the deletion of the cra gene. The transcription level of genes in EMY-01 was used as the control. Error bars represent the standard deviations of three experiments.
In E. aerogenes, the scr regulon for sucrose catabolism consists of four genes, which encode ATP-dependent fructokinase (ScrK), PTS EII transport protein (ScrA), sucrose-6-phosphate hydrolase (ScrB), and sucrose-dependent regulator (ScrR) [29, 30]. The scr regulon contains two independent promoters, one in front of the scrK and a second in front of the scrAB operon, both of which are negatively regulated by ScrR (Additional file 1) [31]. In E. coli, the sequence of the Cra-binding site (Cra box), TGAAACGTTTCA, has been well characterized [16]. Interestingly, a sequence similar to the consensus sequence of the Cra box, TAAAACGTTTTA, was found in the scrAB promoter region of E. aerogenes. In addition, no significant change in the transcription level of crp was observed in our results when cra was disrupted. These results demonstrate that Cra directly binds to the scrAB promoter region to regulate its expression.
The CCR mechanisms by Crp and Cra are complex and interconnected. Indeed, Cra regulates the utilization of several carbon sources, such as xylose, glycerol, lactose, and sorbitol, through activating expression of the crp gene, even though Cra does not regulate the transcription of relevant catabolic genes directly [13]. In this experiment, we demonstrated the sugar consumption genes can be modulated by cra deletion, and that regulation is independent with Crp. This result is another example of controlling CCR. Previously Ji et al. tried it by overexpression of crp(in) gene, which made Klebsiella oxytoca utilize glucose and xylose simultaneously and resulted in higher growth and 2,3-butanediol productivity [32].
Effects of scrAB and ptsG overexpression on the sugar utilization preference in sugarcane molasses
The decreased sucrose utilization by deletion of cra in EMY-69 was partially relieved by scrR mutation, shown in EMY-70 (Fig. 2c). However, sucrose utilization in that strain was not as efficient as glucose or fructose. Therefore, the scrAB operon was cloned in the expression vector pZS21MCS and overexpressed in EMY-70 to increase sucrose utilization. The resulting strain, EMY-70S, showed considerably enhanced sucrose utilization, as shown in Fig. 2c. EMY-70S consumed 61.5 g/L sucrose for 10 h of flask culture using sucrose as a sole carbon source, representing an increase of 42.2% compared to that of EMY-70. In flask cultivation of EMY-70S with mixed sugars, the consumption rate of sucrose improved to a similar rate as that of fructose consumption (Fig. 3e). Interestingly, glucose was the carbon source with the lowest priority among the three carbon sources in EMY-70S. The consumption of glucose was reduced by 46.0% compared to that of its parent strain (Figs. 3d, e).
Thus, we attempted to resolve the reduced efficiency of glucose utilization in EMY-70S at the genetic level. In glucose catabolism, the EIIBCGlc complex encoded by ptsG is known to be a major controller of glucose flux in E. coli [33, 34]. Therefore, a strain overexpressing the ptsG gene was constructed from EMY-70S and named EMY-70SP. Although the overexpression of ptsG did not affect the carbon source utilization of a sole carbohydrate, such as fructose, glucose, or sucrose, the efficiency of glucose utilization was restored in flask cultivations with a mixture of sugars (Figs. 2, 3f). In EMY-70SP, the consumption rate of sucrose was slightly reduced. Among the carbohydrate consumption pathways (Fig. 1), G6P is converted not only from glucose by the ptsG gene product, but also from sucrose by a series of enzymes involved in the scrAB operon. Therefore, the result suggested that overexpression of the ptsG gene presumably activated glucose transport with G6P formation, which resulted in the relative repression of G6P synthesis derived from sucrose.
As shown in the Additional file 3, there were no significant differences in the total concentration of the carbon source consumed by E. aerogenes mutants when the flask cultivations were performed with mixed sugars. These results indicate that the maximum carbon source consumption through glycolysis might be reached, but the sugar preference was modulated by genetic engineering. Still, predictable control of sugar preference would be very beneficial for efficient utilization of biomass that contains a variety of carbon sources. This is because the concentration and content ratio of sugars contained in biomass vary depending on the type of biomass resources and local climate and soil conditions even the same type of biomass [35–37].
Fed-batch fermentation with E. aerogenes mutants using sugarcane molasses
To improve 2,3-butanediol production from sugarcane molasses, a fed-batch fermentation was performed with EMY-70S. As shown in Fig. 5c and Additional file 4, 2,3-butanediol production reached 140.0 g/L at 54 h of cultivation. To the best of our knowledge, this was by far the highest titer of 2,3-butanediol with E. aerogenes achieved by genetic engineering, and was 18.6% higher than that with glucose as a carbon source [27]. A total of 361.7 g/L of sugars in sugarcane molasses was consumed. Among the major byproducts, ethanol production reached 13.7 g/L at 14 h of fed-batch fermentation, after which its concentration was gradually decreased to 1.6 g/L at 54 h, in accordance with a previous report [19]. The production of acetoin, which is a precursor of 2,3-butanediol, was under 2.0 g/L until 40 h of fermentation, but its titer steadily increased to 8.6 g/L at 54 h. The acetoin accumulation is due to the reduced activity of 2,3-butanediol dehydrogenase (BudC) by a high concentration of 2,3-butanediol in the fermentation broth. The production of 2,3-butanediol by EMY-70S was prolonged until 54 h of cultivation, which was 50% longer than the fed-batch with both EMY-01 and EMY-68 (Figs. 5, 6). Even at 36 h of fed-batch fermentation, 2,3-butanediol productivity (g/L/h) of EMY-70S increased by 23.8% compared to that of EMY-68. These results indicate that disruption of both scrR and cra genes presumably relieved the cell stress due to repeated catabolic shift according to the feeding of sugarcane molasses in fed-batch fermentation. However, there was significant accumulation (30 g/L) of glucose in the medium after the fermentation period (Fig. 6c).
Fig. 5.
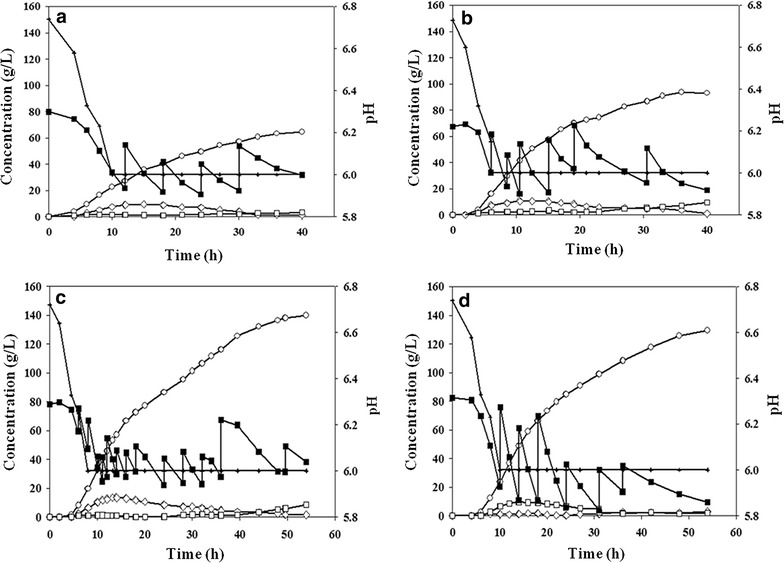
Results of fed-batch fermentations using sugarcane molasses with a EMY-01, b EMY-68, c EMY-70S and d EMY-70SP. Symbols represent 2,3-butanediol (white circles), total sugars (closed squares), pH (pluses), ethanol (open diamonds), and acetoin (open squares).
Fig. 6.
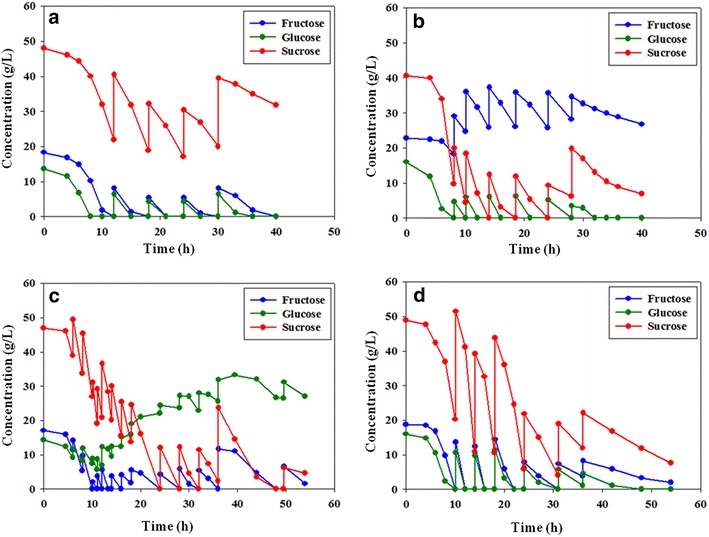
Comparison of fed-batch fermentations of a EMY-01, b EMY-68, c EMY-70S and d EMY-70SP with the consumption of sugars.
To observe the effect of ptsG overexpression on glucose consumption, fed-batch fermentation was performed with EMY-70SP. As shown in Fig. 5d and Additional file 4, 335.58 g/L of sugars in sugarcane molasses was consumed, and 2,3-butanediol production reached 129.36 g/L at 54 h of cultivation. Although 2,3-butanediol production was decreased by 7.6% compared to that of EMY-70S, all three carbons in sugarcane molasses were used efficiently (Fig. 6d). The metabolic burden according to overexpression of the scrAB operon and ptsG gene may be the main reason for the slight reduction observed in fed-batch fermentation with EMY-70SP. These results demonstrate the advantage of genetic engineering in utilizing mixtures of sugars contained in biomass under the fermentation process [38].
Conclusions
The efficient utilization of biomass is necessary in order to develop an economic biorefinery industry. In this study, sugarcane molasses was used as a cheap feedstock for 2,3-butanediol production with metabolically engineered E. aerogenes. For efficient utilization of sugarcane molasses, the removal of transcriptional repressors enabled the mutant strain to metabolize all sugars in sugarcane molasses simultaneously, which increased fermentation duration and 2,3-butanediol productivity. In addition, the carbon preference was modulated by overexpression of key genes involved in each carbon catabolism, which relieved the accumulation of certain carbon sources in fed-batch fermentation. The metabolic engineering approach provided much higher 2,3-butanediol production and efficient utilization of carbon sources involved in sugarcane molasses. The strategy developed in this study is a promising approach for the biorefinery industry, through employing sugar mixtures derived from inexpensive biomass.
Methods
Construction of the gene-deficient mutants
All E. aerogenes strains were derived from the wild-type strain KCTC 2190 (Korean Collection for Type Culture), for which the genome has been sequenced [39]. Previously, the lactate dehydrogenase (LdhA, accession number YP_004594301.1) and sucrose regulator (ScrR, accession number YP_004593287.1) were disrupted from the KCTC 2190 genome by using a λ red recombination method [40], generating EMY-01 (△ldhA) [27] and EMY-68 (△ldhA△scrR) [19], respectively. In this study, the gene encoding catabolite repressor/activator, Cra (previously designated fruR, accession number YP_004592435.1), was deleted from the genome of EMY-01 and EMY-68 in a similar manner, and the resulting strains were named EMY-69 (△ldhA△cra) and EMY-70 (△ldhA△scrR△cra), respectively. The cra_FKF_fw and cra_FKF_rv primers were used for cra disruption, which was confirmed by colony PCR with the primers cra_con_A and cra_con_B. All resulting strains, along with primers and plasmids, used in this study are listed in Tables 1 and 2.
Table 2.
Primers used in this study
| Primer name | Primer sequence (5′ → 3′)a |
|---|---|
| cra_FKF_fw | GTATGTCTATTTAATGGTTGTTTTTTGTACTTCTTACCCAAGGGGCAATTGTGTAGGCTGGAGCTGCTTC |
| cra_FKF_rv | CCATCTGGCGAATAACCTACGAAGAATCTTAACCTTTTTTCGCAAATGAACCTCCTTAGTTCCTATTCC |
| cra_con_A | ACGTAAAAACAGCCCGACAC |
| cra_con_B | CGCTTTTTCTTGCACCATTT |
| pZS21_scrAB_fw | TTTAAGCTTATGGATTTTCAACAGATTTCTCG |
| pZS21_scrAB_rv | AAACCCGGGGCCTGAAAGCAAAACGCTTA |
| pZA31_ptsG_fw | TTTAAGCTTACTCAGGAGCACTCTCAATTATGTT |
| pZA31_ptsG_rv | AAAGGATCCTTAGCTATTGCGGATGTACTCA |
| RT_scrA_fw | TCGGCGGTAACCCTTATCTT |
| RT_scrA_rv | ATTAGCCATCGCCCAGATAG |
| RT_scrB_fw | TGCTACACCGGTAATGTGAAAT |
| RT_scrB_rv | TAGAATTCGAAGCCGCTATCC |
| RT_scrR_fw | CCGGCGTACAGCTGCTTAT |
| RT_scrR_rv | ACGTAGTACCCCTTCCAGCAA |
| RT_scrK_fw | TCAGCCATCTTTCTTTAGATCC |
| RT_scrK_rv | CGGGAAGTGAATATGCTGTTG |
| RT_cra_fw | CCGTATTGCGAACTATCTGGA |
| RT_cra_rv | TAAACAGCAGTTGCGGCATT |
| RT_crp_fw | ATCAAAGAGCACGCTGATTC |
| RT_crp_rv | CCAGCATCTTCAGAATACGG |
| RT_gapA_fw | TTGGTGTTGACGTTGTTGCT |
| RT_gapA_rv | TTCGTAGGACGCTGCTTTTT |
aUnderlined sequences are the homologous sequences with the target genes of E. aerogenes. Restriction sites highlighted in bold.
Construction of plasmids
Enterobacter aerogenes KCTC 2190 was cultured in Luria–Bertani medium overnight, and then total genomic DNA was extracted using the Wizard Genomic DNA Purification Kit (Promega, WI, USA). For scrAB overexpression, the scrAB fragment (a 2,771-bp segment of truncated scrAB gene, accession number YP_004593289.1 and YP_004593288.1) was amplified by PCR using the genomic DNA as a template and the primers pZS21_scrAB_fw and _rv at an annealing temperature (Tm) of 62.4°C. The PCR mixture consisted of 100 ng of genomic DNA, 200 μmol of dNTPs, 0.5 pmol each primer, 10 μL of 5× Phusion GC buffer, 1.5 μL of DMSO, and 1.0 unit of Phusion DNA polymerase (NEB, MA, USA) in a total volume of 50 μL. The amplified DNA fragments were cloned into the pZS21MCS plasmid joined by the HindIII and XmaI restriction sites using the DNA Ligation Kit Mighty Mix (TaKaRa Bio Inc., Shiga, Japan). The E. coli strain DH5α was used for amplification and confirmation of the constructed plasmid. The resulting plasmid was verified by sequencing. The ptsG fragment (a 1,434-bp segment of truncated ptsG gene, accession number YP_004593473.1) was amplified by PCR using the corresponding primers. The PCR mixture composition and procedure were the same as that of scrAB, but the annealing temperature was 64°C. The amplified ptsG fragments were cloned into the pZA31MCS plasmid after digestion with HindIII and BamHI. The plasmids and primers used in this study are listed in Tables 1 and 2.
Media and cultivation conditions
The fermentation medium to produce 2,3-butanediol contained (per L): 3 g KH2PO4, 6.8 g Na2HPO4, 0.75 g KCl, 5.35 g (NH4)2SO4, 0.28 g Na2SO4, 0.26 g MgSO4·7H2O, 0.42 g citric acid, 5 g yeast extract, 10 g casamino acid, and 0.3 mL microelement solution (1 L) containing 34.2 g ZnCl2, 2.7 g FeCl3·6H2O, 10 g MnCl2·4H2O, 0.85 g CuCl2·2H2O, and 0.31 g H3BO3, as described previously [19]. In a flask culture, 80 g/L of the individual sugars (glucose, fructose, or sucrose) was added to the medium as the sole carbon source, and the total 90 g/L mixture of sugars, containing 30 g/L each of glucose, fructose, and sucrose, was added to the medium for the sugar mixture cultivation. For pH neutralization of the flask cultivation, 5% calcium carbonate (CaCO3) was added to the medium before cultivation. The flask, sealed with a silicon stopper, was incubated at 250 rpm and 37°C in a 250-mL flask containing 50 mL medium with appropriate antibiotics or inducers, when needed, at the following concentrations: kanamycin (50 μg/mL), chloramphenicol (50 μg/mL), and anhydrotetracycline (50 ng/mL). The fed-batch fermentation was carried out in a 5-L stirring bioreactor (Bio Control and System, Daejeon, Korea) with a working volume of 3 L. The seed culture prepared previously was inoculated (5%, v/v) into the fermentation medium with an initial pH of 6.8. The pH value of the fermentation medium was decreased gradually to 6.0, and then maintained by the automatic addition of 5 M NaOH. During the fed-batch fermentation process, the operating temperature, airflow, and agitation speed were maintained at 37°C, 1.5 vvm, and 280 rpm, respectively. Antifoam 204 (Sigma, MO, USA) was added when needed. Brazilian sugarcane molasses was used, and its sugar content was 87 g/L fructose, 81 g/L glucose, and 387 g/L sucrose, as described previously [19]. The sterilized sugarcane molasses was fed before the sugar was depleted, and the sugar concentration was maintained under 80 g/L during fed-batch fermentation.
Quantification of the transcription level of genes involved in sucrose catabolism by real-time PCR
Total RNA isolation was performed as previously described [41]. Briefly, cells (1 × 109) were harvested at the mid-exponential phase (6 h) and, then, the total RNA in the cell was stabilized using the bacterial reagent RNAprotect (Qiagen, Hilden, Germany). Cell lysis was conducted with TE buffer containing lysozyme (400 μg/mL), and total RNA isolation was carried out using an RNeasy Mini Kit (Qiagen) following the manufacturer’s protocol. To synthesize the cDNA from total RNA, SuperScript® Reverse Transcriptase (Invitrogen, CA, USA) was used. Real-time PCR was performed using the synthesized cDNA as a template (Model: MJ Mini thermocycler, Software: Opticon Monitor 3, Bio-Rad, CA, USA) with six sets of PCR primers and a SYBR Green mix (Prime Q-Master mix, Genet Bio, Daejon, Korea). The primers used in real-time PCR are listed in Table 2. The transcription levels of the five genes involved in sucrose catabolism were normalized to the housekeeping gene glyceraldehyde 3-phosphate dehydrogenase (gapA). The experiment was repeated three times independently.
Analysis methods
Cell density was monitored by measurement of the optical density at 600 nm (OD600) with a UV/Visible spectrophotometer (DU730, Beckman Coulter, CA, USA). The concentrations of metabolites, obtained from cultivation, were measured by high-performance liquid chromatography (Waters HPLC 1500 series, MA, USA), equipped with a refractive index (RI) detector at 45°C. The amounts of fructose, glucose, and sucrose were measured using a High Performance Carbohydrate Column (Waters) at 35°C, and 80% acetonitrile was used as the mobile phase. Organic acids, 2,3-butanediol, acetoin, and ethanol, were measured using a Sugar SH1011 column (Shodex, Tokyo, Japan) at 75°C, and 10 mM sulfuric acid was used as the mobile phase. The flow rate of both mobile phases was maintained at 0.5 mL/min.
Authors’ contributions
MY Jung designed the study, performed the experiments, analyzed the data, and drafted the manuscript. HM Jung executed the experimental work. JW Lee provided some materials and reagents and helped design the experiments. MK Oh designed the experiments, revised the manuscript, and supervised the project. All authors have read and approved the final manuscript.
Acknowledgements
This study was supported by the R&D program of MKE/KEIT (No. 10035578, Development of 2,3-butanediol and derivate production technology for C-Zero bio-platform industry) and the New & Renewable Energy Core Technology Program of the Korea Institute of Energy Technology Evaluation and Planning (KETEP), granted financial resource from the Ministry of Trade, Industry & Energy, Republic of Korea. (No. 20143030091040).
Compliance with ethical guidelines
Competing interests The authors declare that they have no competing interests.
Abbreviations
- Cra
catabolite repressor/activator
- CCR
carbon catabolite repression
- PTS
phosphotransferase system
- G6P
glucose-6-phosphate
- F6P
fructose-6-phosphate
- F1,6BP
fructose-1,6-bisphosphate
- PCR
polymerase chain reaction
- Ldh
lactate dehydrogenase
- OD
optical density
- ScrA
EII transport protein for sucrose
- ScrB
sucrose-6-phosphate hydrolase
- ScrK
fructokinase
- ScrR
transcriptional repressor of scr regulon
- PtsG
EIIBCGlc complex
- FruA
EIIBCfru complex
- FruB
EIIAfru component
- FruK
fructose-1-phosphate kinase
- Crp
cyclic AMP receptor protein
Additional files
Figure S1. Regulation mechanism related to the utilization of sugars involved in sugarcane molasses and the strategies for constructing the 2,3-butanediol-producing strain in this study. Symbols represent deleted genes (red cross) and overexpressed genes (blue box).
Table S1. Comparison of metabolite profiles obtained from E. aerogenes mutants using sole carbon sources in 10 h of flask cultivation.
Table S2. Comparison of metabolite profiles obtained from E. aerogenes mutants using mixed carbon sources in 12 h of flask cultivation.
Table S3. Comparison of fed-batch fermentation with EMY-01, EMY-68, EMY-70S, and EMY-70SP using sugarcane molasses.
Contributor Information
Moo-Young Jung, Email: artsora@korea.ac.kr.
Hwi-Min Jung, Email: jhm1215@korea.ac.kr.
Jinwon Lee, Email: jinwonlee@sogang.ac.kr.
Min-Kyu Oh, Email: mkoh@korea.ac.kr.
References
- 1.Ragauskas AJ, Williams CK, Davison BH, Britovsek G, Cairney J, Eckert CA, et al. The path forward for biofuels and biomaterials. Science. 2006;311:484–489. doi: 10.1126/science.1114736. [DOI] [PubMed] [Google Scholar]
- 2.van Haveren J, Scott EL, Sanders J. Bulk chemicals from biomass. Biofuel Bioprod Biorefin. 2008;2:41–57. doi: 10.1002/bbb.43. [DOI] [Google Scholar]
- 3.Ji XJ, Huang H, Ouyang PK. Microbial 2,3-butanediol production: a state-of-the-art review. Biotechnol Adv. 2011;29:351–364. doi: 10.1016/j.biotechadv.2011.01.007. [DOI] [PubMed] [Google Scholar]
- 4.Celinska E, Grajek W. Biotechnological production of 2,3-butanediol—current state and prospects. Biotechnol Adv. 2009;27:715–725. doi: 10.1016/j.biotechadv.2009.05.002. [DOI] [PubMed] [Google Scholar]
- 5.Wang AL, Wang Y, Jiang TY, Li LX, Ma CQ, Xu P. Production of 2,3-butanediol from corncob molasses, a waste by-product in xylitol production. Appl Microbiol Biotechnol. 2010;87:965–970. doi: 10.1007/s00253-010-2557-8. [DOI] [PubMed] [Google Scholar]
- 6.Jiang LQ, Fang Z, Guo F, Yang LB. Production of 2,3-butanediol from acid hydrolysates of Jatropha hulls with Klebsiella oxytoca. Bioresour Technol. 2012;107:405–410. doi: 10.1016/j.biortech.2011.12.083. [DOI] [PubMed] [Google Scholar]
- 7.Sun LH, Wang XD, Dai JY, Xiu ZL. Microbial production of 2,3-butanediol from Jerusalem artichoke tubers by Klebsiella pneumoniae. Appl Microbiol Biotechnol. 2009;82:847–852. doi: 10.1007/s00253-008-1823-5. [DOI] [PubMed] [Google Scholar]
- 8.Dai JY, Zhao P, Cheng XL, Xiu ZL. Enhanced production of 2,3-butanediol from sugarcane molasses. Appl Biochem Biotechnol. 2015;175:3014–3024. doi: 10.1007/s12010-015-1481-x. [DOI] [PubMed] [Google Scholar]
- 9.Mosier N, Wyman C, Dale B, Elander R, Lee YY, Holtzapple M, et al. Features of promising technologies for pretreatment of lignocellulosic biomass. Bioresour Technol. 2005;96:673–686. doi: 10.1016/j.biortech.2004.06.025. [DOI] [PubMed] [Google Scholar]
- 10.Gao L, Chi Z, Sheng J, Ni X, Wang L. Single-cell protein production from Jerusalem artichoke extract by a recently isolated marine yeast Cryptococcus aureus G7a and its nutritive analysis. Appl Microbiol Biotechnol. 2007;77:825–832. doi: 10.1007/s00253-007-1210-7. [DOI] [PubMed] [Google Scholar]
- 11.Goerke B, Stulke J. Carbon catabolite repression in bacteria: many ways to make the most out of nutrients. Nat Rev Microbiol. 2008;6:613–624. doi: 10.1038/nrmicro1932. [DOI] [PubMed] [Google Scholar]
- 12.Deutscher J. The mechanisms of carbon catabolite repression in bacteria. Curr Opin Microbiol. 2008;11:87–93. doi: 10.1016/j.mib.2008.02.007. [DOI] [PubMed] [Google Scholar]
- 13.Zhang ZG, Aboulwafa M, Saier MH. Regulation of crp gene expression by the catabolite repressor/activator, Cra, in Escherichia coli. J Mol Microbiol Biotechnol. 2014;24:135–141. doi: 10.1159/000362722. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Plumbridge J. Control of the expression of the manXYZ operon in Escherichia coli: Mlc is a negative regulator of the mannose PTS. Mol Microbiol. 1998;27:369–380. doi: 10.1046/j.1365-2958.1998.00685.x. [DOI] [PubMed] [Google Scholar]
- 15.Newman JR, Fuqua C. Broad-host-range expression vectors that carry the l-arabinose-inducible Escherichia coli araBAD promoter and the araC regulator. Gene. 1999;227:197–203. doi: 10.1016/S0378-1119(98)00601-5. [DOI] [PubMed] [Google Scholar]
- 16.Shimada T, Yamamoto K, Ishihama A. Novel members of the Cra regulon involved in carbon metabolism in Escherichia coli. J Bacteriol. 2011;193:649–659. doi: 10.1128/JB.01214-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Gawand P, Hyland P, Ekins A, Martin VJJ, Mahadevan R. Novel approach to engineer strains for simultaneous sugar utilization. Metab Eng. 2013;20:63–72. doi: 10.1016/j.ymben.2013.08.003. [DOI] [PubMed] [Google Scholar]
- 18.Akaraonye E, Moreno C, Knowles JC, Keshavarz T, Roy I. Poly(3-hydroxybutyrate) production by Bacillus cereus SPV using sugarcane molasses as the main carbon source. Biotechnol J. 2012;7:293–303. doi: 10.1002/biot.201100122. [DOI] [PubMed] [Google Scholar]
- 19.Jung MY, Park BS, Lee J, Oh MK. Engineered Enterobacter aerogenes for efficient utilization of sugarcane molasses in 2,3-butanediol production. Bioresour Technol. 2013;139:21–27. doi: 10.1016/j.biortech.2013.04.003. [DOI] [PubMed] [Google Scholar]
- 20.Sprenger GA, Lengeler JW. Analysis of sucrose catabolism in Klebsiella-pneumoniae and in Scr+ derivatives of Escherichia coli K12. J Gen Microbiol. 1988;134:1635–1644. doi: 10.1099/00221287-134-6-1635. [DOI] [PubMed] [Google Scholar]
- 21.Kornberg HL. Routes for fructose utilization by Escherichia coli. J Mol Microbiol Biotechechnol. 2001;3:355–359. [PubMed] [Google Scholar]
- 22.Deutscher J, Francke C, Postma PW. How phosphotransferase system-related protein phosphorylation regulates carbohydrate metabolism in bacteria. Microbiol Mol Biol Rev. 2006;70:939–1031. doi: 10.1128/MMBR.00024-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Kelker NE, Hanson TE, Anderson RL. Alternate pathways of d-fructose metabolism in Aerobacter aerogenes—a specific d-fructokinase and its preferential role in metabolism of sucrose. J Biol Chem. 1970;245:2060–2065. [PubMed] [Google Scholar]
- 24.Saier MH, Ramseier TM. The catabolite repressor/activator (Cra) protein of enteric bacteria. J Bacteriol. 1996;178:3411–3417. doi: 10.1128/jb.178.12.3411-3417.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Yimga MT, Leatham MP, Allen JH, Laux DC, Conway T, Cohen PS. Role of gluconeogenesis and the tricarboxylic acid cycle in the virulence of Salmonella enterica serovar Tyhimurium in BALB/c mice. Infect Immun. 2006;74:1130–1140. doi: 10.1128/IAI.74.2.1130-1140.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Ramseier TM, Bledig S, Michotey V, Feghali R, Saier MH. The global regulatory protein FruR modulates the direction of carbon flow in Escherichia coli. Mol Microbiol. 1995;16:1157–1169. doi: 10.1111/j.1365-2958.1995.tb02339.x. [DOI] [PubMed] [Google Scholar]
- 27.Jung MY, Ng CY, Song H, Lee J, Oh MK. Deletion of lactate dehydrogenase in Enterobacter aerogenes to enhance 2,3-butanediol production. Appl Microbiol Biotechnol. 2012;95:461–469. doi: 10.1007/s00253-012-3883-9. [DOI] [PubMed] [Google Scholar]
- 28.Diancourt L, Passet V, Verhoef J, Grimont PAD, Brisse S. Multilocus sequence typing of Klebsiella pneumoniae nosocomial isolates. J Clin Microbiol. 2005;43:4178–4182. doi: 10.1128/JCM.43.8.4178-4182.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Reid SJ, Abratt VR. Sucrose utilisation in bacteria: genetic organisation and regulation. Appl Microbiol Biotechnol. 2005;67:312–321. doi: 10.1007/s00253-004-1885-y. [DOI] [PubMed] [Google Scholar]
- 30.Ebner R, Lengeler JW. DNA-sequence of the gene scrA encoding the sucrose transport protein Enzymellscr of the phosphotransferase system from enteric bacteria—homology of the Enzymellscr and Enzymellbgl Proteins. Mol Microbiol. 1988;2:9–17. doi: 10.1111/j.1365-2958.1988.tb00002.x. [DOI] [PubMed] [Google Scholar]
- 31.Jahreis K, Lengeler JW. Molecular analysis of 2 ScrR repressors and of a ScrR-FruR hybrid repressor for sucrose and d-fructose specific regulons from enteric bacteria. Mol Microbiol. 1993;9:195–209. doi: 10.1111/j.1365-2958.1993.tb01681.x. [DOI] [PubMed] [Google Scholar]
- 32.Ji XJ, Nie ZK, Huang H, Ren LJ, Peng C, Ouyang PK. Elimination of carbon catabolite repression in Klebsiella oxytoca for efficient 2,3-butanediol production from glucose-xylose mixtures. Appl Microbiol Biotechnol. 2011;89:1119–1125. doi: 10.1007/s00253-010-2940-5. [DOI] [PubMed] [Google Scholar]
- 33.Plumbridge J. Expression of ptsG, the gene for the major glucose PTS transporter in Escherichia coli, is repressed by MIc and induced by growth on glucose. Mol Microbiol. 1998;29:1053–1063. doi: 10.1046/j.1365-2958.1998.00991.x. [DOI] [PubMed] [Google Scholar]
- 34.Kimata K, Tanaka Y, Inada T, Aiba H. Expression of the glucose transporter gene, ptsG, is regulated at the mRNA degradation step in response to glycolytic flux in Escherichia coli. EMBO J. 2001;20:3587–3595. doi: 10.1093/emboj/20.13.3587. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Fernando S, Adhikari S, Chandrapal C, Murali N. Biorefineries: current status, challenges, and future direction. Energy Fuel. 2006;20:1727–1737. doi: 10.1021/ef060097w. [DOI] [Google Scholar]
- 36.Himmel ME, Ding SY, Johnson DK, Adney WS, Nimlos MR, Brady JW, et al. Biomass recalcitrance: engineering plants and enzymes for biofuels production. Science. 2007;315:804–807. doi: 10.1126/science.1137016. [DOI] [PubMed] [Google Scholar]
- 37.Cherubini F. The biorefinery concept: using biomass instead of oil for producing energy and chemicals. Energy Convers Manag. 2010;51:1412–1421. doi: 10.1016/j.enconman.2010.01.015. [DOI] [Google Scholar]
- 38.Kim SR, Ha SJ, Wei N, Oh EJ, Jin YS. Simultaneous co-fermentation of mixed sugars: a promising strategy for producing cellulosic ethanol. Trends Biotechnol. 2012;30:274–282. doi: 10.1016/j.tibtech.2012.01.005. [DOI] [PubMed] [Google Scholar]
- 39.Shin SH, Kim S, Kim JY, Lee S, Um Y, Oh MK, et al. Complete genome sequence of Enterobacter aerogenes KCTC 2190. J Bacteriol. 2012;194:2373–2374. doi: 10.1128/JB.00028-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Datsenko KA, Wanner BL. One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc Natl Acad Sci USA. 2000;97:6640–6645. doi: 10.1073/pnas.120163297. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Jung MY, Mazumdar S, Shin SH, Yang KS, Lee J, Oh MK. Improvement of 2,3-butanediol yield in Klebsiella pneumoniae by deletion of the pyruvate formate-lyase gene. Appl Environ Microbiol. 2014;80:6195–6203. doi: 10.1128/AEM.02069-14. [DOI] [PMC free article] [PubMed] [Google Scholar]