

Abstract
Background:
Cordyceps militaris is one of well-known medicinal mushrooms with anti-inflammatory, anti-cancer, anti-diabetic, and anti-obesity activities.
Objective:
The objective of the following study is to isolate chemical components from the ethanol extract (Cm-EE) from Cordyceps militaris and to evaluate their anti-inflammatory activities.
Materials and Methods:
Column chromatographic separation was performed and anti-inflammatory roles of these compounds were also examined by using NO production and protein kinase B (AKT) activity assays.
Results:
From Cm-EE, 13 constituents, including trehalose (1), cordycepin (2), 6-hydroxyethyladenosine (3), nicotinic amide (4), butyric acid (5), β-dimorphecolic acid (6), α-dimorphecolic acid (7), palmitic acid (8), linoleic acid (9), cordycepeptide A (10), 4-(2-hydroxy-3-((9E,12E)-octadeca-9,12-dienoyloxy)propoxy)-2-(trimethylammonio)butanoate (11), 4-(2-hydroxy-3-(palmitoyloxy)propoxy)-2-(trimethylammonio)butanoate (12), and linoleic acid methyl ester (13) were isolated. Of these components, compound 2 displayed a significant inhibitory effect on NO production in lipopolysaccharide (LPS)-activated RAW264.7 cells. Furthermore, this compound strongly and directly suppressed the kinase activity of AKT, an essential signalling enzyme in LPS-induced NO production, by interacting with its ATP binding site.
Conclusion:
C. militaris could have anti-inflammatory activity mediated by cordycepin-induced suppression of AKT.
Keywords: Anti-inflammatory activity, cordycepin, Cordyceps militaris (Clavicipitaceae), phytochemical study, protein kinase B
INTRODUCTION
Cordyceps militaris (Clavicipitaceae) is a type of medicinal mushroom with various beneficial activities such as increasing the vitality, stamina, and endurance of the body. More importantly, because of the curative effects of this mushroom on numerous diseases, such as eczema, skin diseases, chronic bronchitis, asthma, and tuberculosis, it has been prescribed for a long time in many different countries such as China, Japan, and Korea. Through systematic research, a large amount of information on its chemical ingredients (which include cordycepin, militarin, 3,6-dihydroxydecanolide, cordycepic acid, and cordyheptapeptide) and its wide range of pharmacological activities (including anti-inflammatory, anti-oxidative, anti-tumourigenic, anti-hyperglycaemic, anti-apoptotic, immunomodulatory, nephroprotective, and hepatoprotective effects) have been revealed.[1,2,3]
In spite of the large number of beneficial roles of this mushroom, development of C. militaris as a nutraceutical remedy was limited due to restrictions in the harvested amount of this mushroom. However, many different culture methods aimed at increasing the cultivation amount of C. militaris have been designed. Our group has also tried to develop new cultivation methods with brown rice to produce artificially cultivated fruit bodies of this mushroom [Figure 1a]. Additionally, we have reported several pharmacological activities of this mushroom, including platelet aggregation and macrophage-mediated inflammatory reaction. Because phytochemical studies with artificially cultivated fruit bodies of C. militaris have not been fully carried out, isolation, structure determination, and anti-inflammatory activities of the compounds from a 70% ethanol extract (EE) of C. militaris fruit bodies are described in this study.
Figure 1.
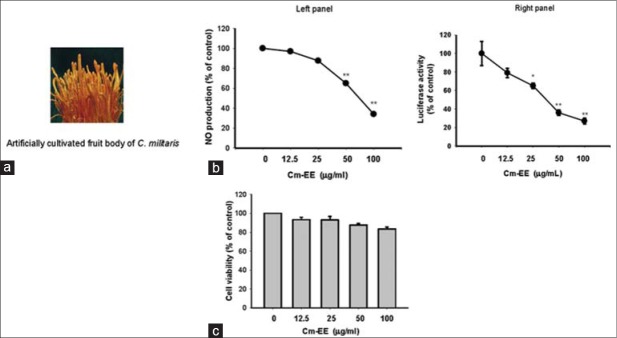
A photo of artificially cultivated fruit bodies of Cordyceps militaris and the anti-inflammatory activity of its ethanol extract (Cm-EE). (a) A photo of artificially cultivated fruit bodies of C. militaris. (b) The level of nitric oxide was determined by the Griess assay from culture supernatants of RAW264.7 cells treated with Cm-EE (0–100 μg/ml) and lipopolysaccharide (1 μg/ml) for 24 h. (c) Cell viability of RAW264.7 cells and HEK293 cells treated with Cm-EE was determined by an 3-4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide, a tetrazole assay (**P < 0.01 compared to control)
MATERIALS AND METHODS
Materials
Artificially cultivated C. militaris [Figure 1a] was purchased from Mush-Tech (Gangwon-do, Korea). C. militaris was identified by Prof. Jae Mo Sung (Kangwon National University, Chuncheon, Korea). A voucher specimen (Registration number: RDA-NHJ-312-Cm-EE) was deposited in the herbarium at National Institute of Horticultural and Herbal Science (RDA, Eumsung, Korea). Lipopolysaccharide (LPS, Escherichia coli 0111:B4), staurosporin (Stau: Purity >95%), and (3-4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide, a tetrazole (MTT), were purchased from Sigma Chemical Co. (St. Louis, MO). LY294002 (LY: Purity >95%) and Wortmannin (Wort: Purity >95%), PI3K inhibitors, were obtained from Calbiochem (La Jolla, CA). The luciferase construct containing binding promoters for NF-κB was gift from Prof. Chung, Hae Young (Pusan National University, Pusan, Korea). Antibodies against inducible nitric oxide synthase (iNOS), cyclooxygenase (COX)-2, and β-actin were purchased from Santa Cruz Biotechnology (Santa Cruz, CA). Fetal bovine serum (FBS) and RPMI 1640 were obtained from GIBCO (Grand Island, NY). RAW264.7 and HEK293 cells were purchased from ATCC (Rockville, MD). All of the other chemicals were purchased from Sigma. GFP-fused wild-type (WT) protein kinase B construct (GFP-Akt-WT) and its mutant form (GFP-Akt-C310A) were used as reported previously.[4]
General experimental procedures
Nuclear magnetic resonance (NMR) spectra were measured on a Varian UNITY INOVA 500 spectrometer or on a Bruker AMX 500 spectrometer operating at 500 MHz for 1H-NMR and 125 MHz for 13C-NMR with chemical shifts given in ppm (δ). Electrospray ionization-tandem mass spectrometry (ESI-MS) was measured on an Agilent 1100 liquid chromatography/mass spectrometry (LC/MS) spectrometer with a Phenomenex Luna C18 analytical column (5 μm, 4.6 × 100 mm). The high performance liquid chromatography (HPLC) apparatus (Waters Alliance series 2795 system) was composed of a vacuum degasser, a quaternary pump, a photodiode array detector, an auto-injector, and a column compartment with a thermostat. Flash and open column chromatography was carried out on Merck silica gel 60 (70–230 mesh). Reversed-phase HPLC was performed on a Gilson 321 pump system using Phenomenex Luna C18 column (10 μm, 10 × 250 mm) with a flow rate of 4 ml/min or 2 ml/min.
Extraction and isolation
Dried and powdered fresh fruiting bodies of C. militaris (1.0 kg) were extracted with 70% EtOH to prepare the Cm-EE. The powder was then treated with 100% MeOH (three times, each with 10 l) using a reflux apparatus that yielded a crude extracts (402 g) upon removal of the solvent in vacuo. The ethyl acetate layer (600 mg) was subjected to a C18 sepak chromatography eluting with step gradients of 20% (220 mg), 40% (130 mg), 60% (100 mg), 80% (60 mg), and 100% MeOH (40 mg) in H2O, successively. The 20% MeOH fraction (220 mg) was separated by reversed-phase HPLC (Phenomenex Luna 10 μm C18 (2), [10 × 250 mm, 4 ml/min, UV 210 nm detection]) using a gradient solvent system from 10% to 100% acetonitrile over 60 min to afford compound 1 (tR = 3 min, 5 mg) and compound 2 (tR = 6.5 min, 3 mg). The 40% MeOH fraction (130 mg), contained the compound 3 and 4, was separated by reversed-phase HPLC (Phenomenex Luna 10 μm C18 (2), [10 × 250 mm, 4 ml/min, UV 210 nm detection]) using a gradient solvent system from 10% to 100% acetonitrile over 60 min. Compound 3 and 4 were finally purified by reversed-phase HPLC (Phenomenex Luna 10 μm C18 (2) column [10 × 250 mm, 2 ml/min, UV 254 nm detection]) with 10% methanol isocratic condition to yield 1.5 mg (tR = 8 min) and 1 mg (tR = 6 min), respectively. The methylene chloride crude extract (1.5 g) was fractionated on a silica gel (230–400 mesh) column and eluted with CH2Cl2-MeOH (100:1 → 1:1, gradient system) to yield 10 fractions (MF1-MF10). The MF1 sub-fraction (400 mg) was purified by preparative C18 HPLC (10–100% acetonitrile in water for 60 min, 4 ml/min, 205 nm) to yield 13 (tR = 55 min, 1.5 mg). The sub-fraction MF4 (250 mg) was purified by separation with preparative C18 HPLC (10–100% acetonitrile in water for 60 min, 4 ml/min, 210 nm) to yield 5 (2 mg), 8 (2.5 mg), 9 (5 mg), and 10 (2 mg) (tR = 20 min, 57 min, 60 min, and 45 min, respectively). The compounds 6 and 7, which were isolated as cis-trans mixture (3 mg), were also isolated from sub-fraction M7 by preparative C18 HPLC (10–100% acetonitrile in water for 60 min, 4 ml/min, 240 nm). The MF10 sub-fraction (150 mg) was chromatographed on C18 sepak (230-400 mesh) column, and eluted with water and methanol (60%→100% methanol) to yield three fractions (MF1060, MF1080, and MF10100). The fraction MF10100 was purified by preparative C18 HPLC (10–100% acetonitrile in water for 60 min, 4 ml/min, 205 nm) to yield 11 (tR = 45 min, 1.5 mg) and 12 (tR = 47 min, 1.0 mg). The tested compounds were demonstrated to be pure as evidenced by NMR and HPLC analyses (purity ≥95%).
Spectral and physicochemical data
Trehalose (1)
1H NMR (500 MHz, D2O) δ 5.02 (1H, d, J = 3.8 Hz, H-1), 3.47 (1H, dd, J = 10.0, 3.8 Hz, H-2), 3.64 (1H, m, H-3), 3.27 (1H, t, J = 9.5 Hz, H-4), 3.67 (1H, m, H-5), 3.67 (1H, m, H-6a), 3.58 (1H, dd, J = 12.0, 5.0 Hz, H-6b); 13C NMR (125 MHz, D2O) d 93.1 (C-1), 70.8 (C-2), 72.3 (C-3), 69.5 (C-4), 71.9 (C-5), 60.3 (C-6); ESIMS m/z 365 [M + Na]+.
Cordycepin (2)
1H NMR (500 MHz, D2O) δ 8.09 (1H, s, H-8), 7.97 (1H, s, H-2), 5.84 (1H, d, J = 2.2 Hz, H-1’), 4.62 (1H, m, H-2’), 4.44 (1H, m, H-4’), 3.74 (1H, dd, J = 12.7, 2.7 Hz, H-5’), 3.53 (1H, dd, J = 12.7, 4.5 Hz, H-5’), 2.12 (1H, ddd, J = 14.7, 8.9, 5.9 Hz, H-3’), 2.01 (1H, ddd, J = 13.8, 6.5, 3.2 Hz, H-3’); 13C NMR (125 MHz, D2O) d 155.3 (C-4), 152.3 (C-2), 149.0 (C-6), 139.6 (C-8), 119.6 (C-5), 90.7 (C-1’), 81.1 (C-4’), 74.8 (C-2’), 62.5 (C-5’), 33.0 (C-3’); ESIMS m/z 252 [M + H]+.
N6-(2-Hydroxyethyl) adenosine (3)
Colourless gum; 1H NMR (500 MHz, D2O) δ 8.09 (1H, s, H-8), 8.04 (1H, s, H-2), 5.87 (1H, d, J = 5.6 Hz, H-1’), 4.61 (1H, t, J = 5.6 Hz, H-2’), 4.24 (1H, m, H 4’), 4.11 (1H, m, H-3’), 3.73 (1H, dd, J = 12.9, 3.6 Hz, H-5’), 3.67-3.65 (5H, m, H-5’, H-9, H-10); ESIMS m/z 312 [M + H]+.
Nicotinic amide (4)
White powder; 1H NMR (500 MHz, D2O) δ 8.70 (1H, s, H-6), 8.49 (1H, d, J = 5.0 Hz, H-2), 8.01 (1H, d, J = 8.0 Hz, H-4), 7.37 (1H, m, H-3); 13C NMR (125 MHz, D2O) δ 170.6 (C-7), 151.6 (C-6), 147.4 (C-2), 136.3 (C-4), 129.1 (C-5), 124.0 (C-3); ESIMS m/z 123 [M + H]+.
Butyric acid (5)
1H NMR (500 MHz, CD3 OD) δ 2.27 (2H, t, J = 15.0 Hz, H-2), 1.63 (2H, dq, J = 15.0, 8.0 Hz, H-3), 0.96 (3H, t, J = 8.0 Hz, H-4); 13C NMR (125 MHz, CD3 OD) δ 176.1 (C-1), 35.4 (C-2), 18.0 (C-3), 12.6 (C-4).
β-Dimorphecolic acid (6)
1H NMR (500 MHz, CDCl3) δ 0.89 (3H, t, J = 7.0, H-18), 1.32 (14H, m, H-4, H-5, H-6, H-7, H-15, H-16, H-17), 1.52 (2H, m, H-8), 1.62 (2H, m, H-3), 2.16 (2H, m, H-14), 2.36 (2H, t, J = 7.5 Hz, H-2), 4.11 (1H, dd, J = 13.0, 6.5 Hz, H-9), 5.58 (1H, dd, J = 15.0, 7.0 Hz, H-13), 5.71 (1H, dd, J = 15.0, 7.0 Hz, H-10), 6.03 (1H, dd, J = 15.0, 10.0 Hz, H-12), 6.18 (1H, dd, J = 15.0, 10.0 Hz, H-11); ESIMS m/z 319 [M + Na]+.
α-Dimorphecolic acid (7)
Cis and trans form mixture, ESIMS m/z 319 [M + Na]+.
Palmitic acid (8)
1H NMR (500 MHz, CDCl3) δ 2.36 (2H, t, J = 7.5 Hz, H-2), 1.64 (2H, dt, J = 15.0, 7.5 Hz, H-15), 1.38-1.22 (24H, m, H-3,4,5,6,7,8,9,10,11,12,13,14), 0.89 (3H, t, J = 7.0 Hz, H-16); 13C NMR (125 MHz, CDCl3) δ 180.0 (C-1), 34.0 (C-2), 31.9 (C-14), 29.6 (C- 6, 7, 8, 9, 10, 11, 12), 29.3 (C-5), 29.2 (C-13), 29.0 (C-4), 24.7 (C-3), 22.7 (C-15), 14.1 (C-16).
Linoleic acid (9)
Colourless oil; 1H NMR (500 MHz, CDCl3) δ 5.44-5.25 (4H, m, H-9, 10, 12, 13), 2.78 (2H, m, H-11), 2.35 (2H, m, H-2), 2.06 (4H, m, H-8, 14), 1.64 (2H, m, H-17), 1.44-1.22 (14H, m, H-3,4,5,6,7,15,16), 0.90 (3H, m, H-18); 13C NMR (125 MHz, CDCl3) δ 180.6 (C-1), 130.1 (C-9), 129.9 (C-13), 128.0 (C-10), 127.8 (C-12), 34.1 (C-2), 31.5 (C-16), 29.5 (C-6), 29.4 (C-15), 29.1 (C-7), 29.0 (C-5), 29.0 (C-4), 27.2 (C-14), 27.1 (C-8), 25.6 (C-11), 24.6 (C-3), 22.6 (C-17), 14.1 (C-18).
Cordycepeptide (10)
ESIMS m/z 488 [M + H]+.
Glycerfatty acid (11)
ESIMS m/z 498 [M + H]+.
Glycerfatty acid (12)
ESIMS m/z 474 [M + H]+.
Linoleic acid methyl ester (13)
1H NMR (500 MHz, CDCl3) δ 5.44-5.30 (4H, m, H-9, 10, 12, 13), 3.67 (3H, s,-OCH3), 2.78 (2H, m, H-11), 2.35 (2H, m, H-2), 2.05 (4H, m, H-8, 14), 1.62 (2H, m, H-3), 1.41-1.26 (14H, m, H-4,5,6,7,15,16,17), 0.89 (3H, m, H-18); 13C NMR (125 MHz, CDCl3) d 174.4 (C-1), 130.2 (C-9), 130.3 (C-13), 128.0 (C-10), 127.8 (C-12), 51.4 (OCH3), 34.1 (C-2), 31.5 (C-16), 29.6 (C-6), 29.4 (C-15), 29.1 (C-7), 29.1 (C-5), 29.1 (C-4), 27.2 (C-14), 27.2 (C-8), 25.6 (C-11), 24.9 (C-3), 22.6 (C-17), 14.1 (C-18).
Cell culture
RAW264.7 and HEK293 cells were cultured with RPMI 1640 medium supplemented with 10% heat-inactivated FBS, glutamine, and antibiotics (penicillin and streptomycin) at 37°C in a 5% CO2 atmosphere. For each experiment, the cells were detached with a scraper. Examination of cell densities of 2 × 106 cells/ml revealed that the proportion of dead cells was consistently < 1% using Trypan blue dye exclusion as the criteria for viability.
Nitric oxide and PGE2 production
After pre-incubation of RAW264.7 cells (1 × 106 cells/ml) for 18 h, the cells were pre-treated with Cm-EE and its chemical ingredients for 30 min. The cells were then further incubated with LPS (1 μg/ml) for 24 h. The inhibitory effects of Cm-EE and its chemical ingredients on NO and PGE2 production were determined by analyzing NO and PGE2 levels using the Griess reagent and an EIA kit, as described previously.[5,6]
mRNA analysis by semi-quantitative reverse transcriptase-polymerase chain reaction
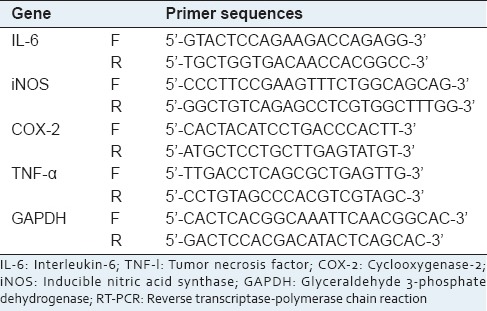
To evaluate cytokine mRNA expression levels, total RNA was isolated from the LPS-treated RAW264.7 cells with TRIzol Reagent (Gibco BRL), according to the manufacturer's instructions. The total RNA was stored at −70°C until use. Semi-quantitative reverse transcriptase (RT) reactions were conducted as reported previously.[7,8] The primers (Bioneer, Seoul, Korea) used are indicated in Table 1.
Table 1.
Primer sequences used in the RT-PCR analysis
Cell viability test
After pre-incubation of RAW264.7 cells (1 × 106 cells/ml) for 18 h, Cm-EE and its chemical ingredients were added to the cells and incubated for 24 h. The cytotoxic effects of Cm-EE and its chemical ingredients were then evaluated by a conventional MTT assay, as reported previously.[9] Three hours prior to culture termination, 10 μL of a MTT solution (10 mg/ml in phosphate buffered saline, pH 7.4; PBS) was added, and the cells were continuously cultured until termination of the experiment. The incubation was halted by the addition of 15% sodium dodecyl sulfate into each well to solubilize the produced formazan.[10] The absorbance at 570 nm (OD570) was measured using a SpectraMax 250 microplate reader (BioTex, Bad Friedrichshall, Germany).
Immunoblotting analysis
HEK293 cells (5 × 106 cells/ml) were lysed in lysis buffer (20 mM Tris-HCl, pH 7.4, 2 mM EDTA, 2 mM EGTA, 50 mM β-glycerophosphate, 1 mM sodium orthovanadate, 1 mM dithiothreitol, 1% Triton X-100, 10% glycerol, 10 μg/ml leupeptin, 10 μg/ml aprotinin, 10 μg/ml pepstatin, 1 mM benzimidine, and 2 mM hydrogen peroxide) for 30 min rotating at 4°C. The lysates were clarified by centrifugation at 16,000 × g for 10 min at 4°C. The soluble cell lysates were immunoblotted, and the total level of AKT was visualised as previously reported.[11]
Immunoprecipitation and in vitro kinase assay for protein kinase B
HEK 293 cells were placed on ice and extracted with lysis buffer as reported previously.[4] Lysates prepared from GFP-AKT-WT or GFP-AKT-C310A overexpressing cells was immunoprecipitated with anti-GFP monoclonal antibody immobilized on protein G-Sepharose (Amersham Biosciences). The kinase activity of the immunoprecipitated beads was measured with peptide substrate, Suntide (RRKDGATMKTFCGTPE), as reported previously.[4] The kinase activity of purified AKT1 enzyme was evaluated by the kinase profiler service from Millipore (Billerica MA) as reported previously.[4]
Statistical analysis
All of the data except Figures 1a and 2 are presented as the means ± standard mean of the error of three different experiments performed with three samples. For the statistical comparisons, these results were analyzed using an ANOVA/Scheffe's post hoc test and a Kruskal–Wallis/Mann–Whitney test. A P < 0.05 was considered to be statistically significant. All of the statistical tests were performed using SPSS (SPSS Inc., Chicago, IL, USA).
Figure 2.

Phytochemical analysis of cultivated fruit bodies of Cordyceps militaris. (a) Isolation of ingredients from the ethyl acetate layer of Cm-EE. (b) Isolation of ingredients from the methylene chloride layer of Cm-EE. (c) Chemical structures of purified compounds (1–13)
RESULTS AND DISCUSSION
After establishing a cultivation method to greatly increase production of C. militaris [Figure 1a], we sought to determine the pharmacological activities of the mushroom for later application to various purposes, such as development of functional food products, cosmetics, and new druggable compounds.[12] A 70% EE of C. militaris (Cm-EE) was able to strongly inhibit the production of nitric oxide (NO) from LPS-treated RAW264.7 cells up to 100 mg/mL [Figure 1b, left panel] without altering cell viability [Figure 1c]. Similarly, Cm-EE dose-dependently suppressed luciferase activity mediated by nuclear factor (NF)- κB [Figure 1b, right panel], a representative inflammation-regulatory transcription factor.[13] These results led us to employ column chromatography and HPLC to further investigate what types of compounds from the EE of C. militaris can attenuate inflammation [Figure 2a and b]. As we expected, we isolated 13 different compounds from 6 fractions [Figure 2a and b]. Through spectroscopic analysis, these compounds [Figure 2c] were identified as trehalose (1), cordycepin (2), 6-hydroxyethyladenosine (3), nicotinic amide (4), butyric acid (5), β-dimorphecolic acid (6), α-dimorphecolic acid (7), palmitic acid (8), linoleic acid (9), cordycepeptide A (10), 4-(2-hydroxy-3-((9E,12E)-octadeca-9,12-dienoyloxy) propoxy)-2-(trimethylammonio) butanoate (11), 4- (2-hydroxy-3-(palmitoyloxy) propoxy)-2-(trimethylammonio) butanoate (12), and linoleic acid methyl ester (13) by comparison with their previously published spectroscopic data.[14,15,16,17,18,19,20] Of these compounds, cordycepeptide A (10), 4-(2-hydroxy-3-((9E,12E)-octadeca-9,12-dienoyloxy) propoxy)-2-(trimethylammonio) butanoate (11), and 4-(2-hydroxy-3-(palmitoyloxy) propoxy)-2-(trimethylammonio) butanoate (12) were isolated for the first time from the C. militaris, indicating that these compounds could potentially serve as useful chemotaxonomic markers for this mushroom.
To confirm which of these components possess anti-inflammatory activity, the suppressive activities of the 13 compounds on the production of NO and PGE2 and the expression of inflammatory genes such as iNOS, COX-2, tumor necrosis factor (TNF)-α, and interleukin (IL)-6 were examined in LPS-treated RAW264.7 cells.[21,22,23,24] We also evaluated the molecular target of the main active component using immunoprecipitated and purified enzyme.
Of 13 compounds, compound 2 showed the highest NO inhibitory activity at 5 and 10 mM, while other compounds (7 and 9) also significantly decreased NO production [Figure 3a and b]. Furthermore, compound 2 dose-dependently suppressed the release of NO and PGE2 [Figure 4a, left panel] and the expression of various inflammatory genes such as TNF-α, iNOS, COX-2, and IL-6 [Figure 4b], without altering the viability of RAW264.7 cells [Figure 4c], implying that the inhibitory activity of compound 2 is not due to its nonspecific pharmacological action. Meanwhile, standard compounds (L-NAME and indomethacine) diminished the release of NO and PGE2 [Figure 4a, right panel], indicating that our experimental conditions were well established.
Figure 3.
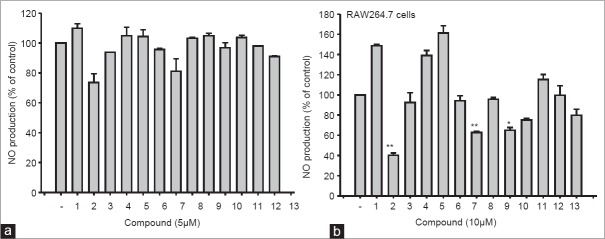
Inhibitory effects of compounds (1–13) on the production of nitric oxide (NO) in lipopolysaccharide (LPS)-treated RAW264.7 cells. (a and b) Level of NO was determined by the Griess assay from culture supernatants of RAW264.7 cells treated with compounds (1–13) and LPS (1 ⊠g/ml) for 24 h (a: Right panel)
Figure 4.
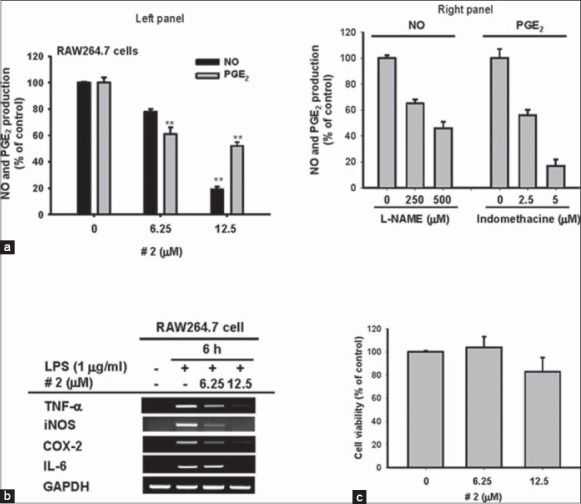
Inhibitory effects of compound 2 on the production of nitric oxide (NO) and PGE2, the expression of inflammatory genes, and the viability of RAW264.7 cells. (a: Left panel and right panel) Levels of NO and PGE2 were determined by the Griess assay, ELISA, and EIA from the culture supernatants of RAW264.7 cells treated with compound 2 (0–12.5 μM), standard compounds (L-NAME and indomethacine), and lipopolysaccharide (LPS) (1 μg/ml) for 24 h. (b) The mRNA levels of inducible nitric oxide synthase, cyclooxygenase-2, tumor necrosis factor-α, and interleukin-6 were determined by reverse transcriptase-polymerase chain reaction using RAW264.7 cells treated with compound 2 (0–12.5 μM) and LPS (1 μg/ml) for 24 h. (c) Cell viability of RAW264.7 cells treated with compound 2 (0–12.5 μM) was determined by 3-4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide, a tetrazole assay. All data are presented as the mean ± standard error of the mean of three different experiments performed with three samples (*P < 0.05 and **P < 0.01 compared to control or normal group)
It has been reported that compound 2 (cordycepin) is capable of suppressing the phosphorylation of AKT,[25] an essential step for inducing inflammatory signaling for NF-κB activation,[4] in LPS-treated RAW264.7 cells. Therefore, we examined whether this compound can directly suppress the enzyme activity of AKT using an in vitro kinase assay. As we expected, compound 2 blocked immunoprecipitated AKT enzyme activity up to 85%, similar to the effect of hydroquinone [Figure 5a], a powerful thiolation-inducing benzene metabolite with chemical reactivity.[4] In agreement with a previous report,[25] compound 2 also diminished p-AKT levels [Figure 5b]. Unlike hydroquinone, this compound still maintained its suppressive activity in the kinase assay performed with mutant Akt (Akt-C310A) with alanine at the cysteine-310 residue [Figure 5a], which is one of the allosteric sites affecting its kinase activity.[4] This indicates that compound 2 is not able to bind to this site. Instead, co-treatment of compound 2 with staurosporin, a competitive inhibitor to the ATP binding site, indicated the possibility that cordycepin can also bind to the ATP binding site, since no additive or synergistic inhibitory activity was exhibited toward purified AKT even with co-treatment of these two compounds [Figure 5c]. The importance of AKT in the NO production process in LPS-treated RAW264.7 cells was also confirmed by treatment with PI3K/AKT inhibitors LY294002 (LY) and wortmannin (Wort). These compounds clearly diminished the level of NO produced by LPS exposure in RAW264.7 cells [Figure 5d]. Therefore, these results strong suggest that compound 2 is an active component in Cm-EE with anti-inflammatory effects associated with direct suppression of AKT.
Figure 5.
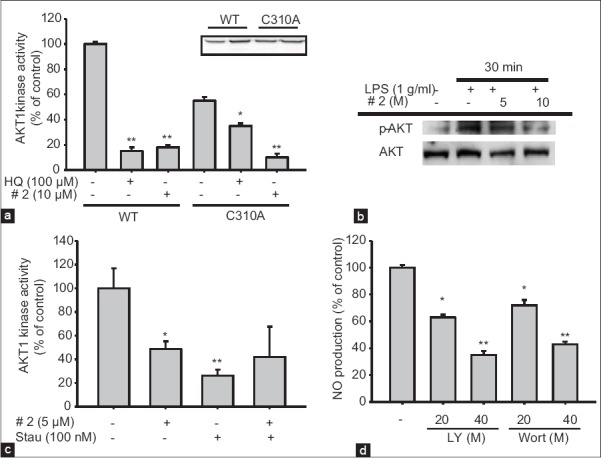
Inhibitory effect of compound 2 on protein kinase B (AKT) kinase activity. (a and c) Direct inhibition of AKT kinase activity by compound 2 or staurosporin (Stau) was measured using immunoprecipitated or purified AKT (WT or mutant [C310A]). The level of AKT transfected was measured by immunoblotting analysis. (b) After immunoblotting, levels of phospho-or total AKT in whole lysates from lipopolysaccharide (LPS)-activated RAW264.7 cells pretreated with compound 2 (5 and 10 μM) were identified using specific antibodies. (d) Level of nitric oxide was determined by the Griess assay from the culture supernatants of RAW264.7 cells treated with standard compounds (LY294002 [LY] and wortmannin [Wort]), and LPS (1 μg/ml) for 24 h. All of the data are presented as the mean ± standard error of the mean of three different experiments performed with three samples *P < 0.05 and **P < 0.01 compared to control or normal
Cordycepin (compound 2) is well-known major component identified in C. militaris, which has been received much attention because of its broad-spectrum biological activities, such as anti-cancer, antioxidant and anti-inflammatory activities.[26,27,28] In fact, in our study, compound 2 was also revealed to inhibit inflammatory responses through the blockade of AKT [Figure 5]. Even though the possibility that cordyceptin can suppress AKT pathway by measuring its simple phosphorylation level,[25,29] there was no direct evidence to prove this possibility. However, we apparently verified in detail that cordycepin can directly suppress AKT activity [Figure 5a]. Thus, cordyceptin blocked the AKT kinase activity by competing with ATP to ATP binding domain of AKT, indicating direct AKT regulatory role of cordyceptin. Because AKT is a critical molecule in numerous inflammatory responses,[4] our data seem to open further application of cordyceptin for the development of various functional products, such as, cosmetics, functional foods and new drugs to ameliorate various inflammation-based symptoms.
CONCLUSION
Thirteen compounds were identified from the EE of C. militaris. Using phytochemical and pharmacological studies, compound 2 was found to be a major anti-inflammatory compound in C. militaris with direct AKT inhibitory property. Therefore, further investigation will be conducted to evaluate its potential application using various in vivo inflammatory disease models such as gastritis, colitis, arthritis, and hepatitis, mediated by AKT.
ACKNOWLEDGMENTS
This work was supported by the Rural Development Administration Grant by the Korean Government-PJ008477, PJ008321.
Footnotes
Source of Support: Nil
Conflict of Interest: None declared.
REFERENCES
- 1.Yue K, Ye M, Zhou Z, Sun W, Lin X. The genus Cordyceps: A chemical and pharmacological review. J Pharm Pharmacol. 2013;65:474–93. doi: 10.1111/j.2042-7158.2012.01601.x. [DOI] [PubMed] [Google Scholar]
- 2.Killen JC, Axford LC, Newberry SE, Simpson TJ, Willis CL. Convergent syntheses of 3,6-dihydroxydec-4-enolides. Org Lett. 2012;14:4194–7. doi: 10.1021/ol3018566. [DOI] [PubMed] [Google Scholar]
- 3.Das SK, Masuda M, Sakurai A, Sakakibara M. Medicinal uses of the mushroom Cordyceps militaris: Current state and prospects. Fitoterapia. 2010;81:961–8. doi: 10.1016/j.fitote.2010.07.010. [DOI] [PubMed] [Google Scholar]
- 4.Lee JY, Lee YG, Lee J, Yang KJ, Kim AR, Kim JY, et al. Akt Cys-310-targeted inhibition by hydroxylated benzene derivatives is tightly linked to their immunosuppressive effects. J Biol Chem. 2010;285:9932–48. doi: 10.1074/jbc.M109.074872. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Cho JY, Baik KU, Jung JH, Park MH. In vitro anti-inflammatory effects of cynaropicrin, a sesquiterpene lactone, from Saussurea lappa. Eur J Pharmacol. 2000;398:399–407. doi: 10.1016/s0014-2999(00)00337-x. [DOI] [PubMed] [Google Scholar]
- 6.Kim KS, Choi HM, Yang HI, Yoo MC. WIN-34B may have analgesic and anti-inflammatory effects by reducing the production of pro-inflammatory mediators in cells via inhibition of IkB signaling pathways. Biomol Ther (Seoul) 2012;20:50–6. doi: 10.4062/biomolther.2012.20.1.050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Lee YG, Chain BM, Cho JY. Distinct role of spleen tyrosine kinase in the early phosphorylation of inhibitor of kappaB alpha via activation of the phosphoinositide-3-kinase and Akt pathways. Int J Biochem Cell Biol. 2009;41:811–21. doi: 10.1016/j.biocel.2008.08.011. [DOI] [PubMed] [Google Scholar]
- 8.Park JM, Ho DH, Yun HJ, Kim HJ, Lee CH, Park SW, et al. Dexamethasone induces the expression of LRRK2 and a-synuclein, two genes that when mutated cause Parkinson's disease in an autosomal dominant manner. BMB Rep. 2013;46:454–9. doi: 10.5483/BMBRep.2013.46.9.234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Kim MY, Cho JY. 20S-dihydroprotopanaxatriol modulates functional activation of monocytes and macrophages. J Ginseng Res. 2013;37:300–7. doi: 10.5142/jgr.2013.37.300. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Eo SH, Cho H, Kim SJ. Resveratrol inhibits nitric oxide-induced apoptosis via the NF-Kappa B pathway in rabbit articular chondrocytes. Biomol Ther. 2013;21:364–70. doi: 10.4062/biomolther.2013.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Lee DH, Cho HJ, Kim HH, Rhee MH, Ryu JH, Park HJ. Inhibitory effects of total saponin from Korean red ginseng via vasodilator-stimulated phosphoprotein-Ser (157) phosphorylation on thrombin-induced platelet aggregation. J Ginseng Res. 2013;37:176–86. doi: 10.5142/jgr.2013.37.176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Wu G, Li L, Sung GH, Kim TW, Byeon SE, Cho JY, et al. Inhibition of 2,4-dinitrofluorobenzene-induced atopic dermatitis by topical application of the butanol extract of Cordyceps bassiana in NC/Nga mice. J Ethnopharmacol. 2011;134:504–9. doi: 10.1016/j.jep.2010.12.012. [DOI] [PubMed] [Google Scholar]
- 13.Lee YG, Lee J, Byeon SE, Yoo DS, Kim MH, Lee SY, et al. Functional role of Akt in macrophage-mediated innate immunity. Front Biosci (Landmark Ed) 2011;16:517–30. doi: 10.2741/3702. [DOI] [PubMed] [Google Scholar]
- 14.Rukachaisirikul V, Pramjit S, Pakawatchai C, Isaka M, Supothina S. 10-membered macrolides from the insect pathogenic fungus Cordyceps militaris BCC 2816. J Nat Prod. 2004;67:1953–5. doi: 10.1021/np0401415. [DOI] [PubMed] [Google Scholar]
- 15.Bu T, Liu M, Zheng L, Guo Y, Lin X. a-Glucosidase inhibition and the in vivo hypoglycemic effect of butyl-isobutyl-phthalate derived from the Laminaria japonica rhizoid. Phytother Res. 2010;24:1588–91. doi: 10.1002/ptr.3139. [DOI] [PubMed] [Google Scholar]
- 16.Mojarrab M, Delazar A, Moghadam SB, Nazemiyeh H, Nahar L, Kumarasamy Y, et al. Armenin and isoarmenin – two prenylated coumarins from the aerial parts of Artemisia armeniaca. Chem Biodivers. 2011;8:2097–103. doi: 10.1002/cbdv.201000284. [DOI] [PubMed] [Google Scholar]
- 17.Choi JN, Kim J, Lee MY, Park DK, Hong YS, Lee CH. Metabolomics revealed novel isoflavones and optimal cultivation time of Cordyceps militaris fermentation. J Agric Food Chem. 2010;58:4258–67. doi: 10.1021/jf903822e. [DOI] [PubMed] [Google Scholar]
- 18.Ballas LM, Bell RM. Topography of glycerolipid synthetic enzymes. Synthesis of phosphatidylserine, phosphatidylinositol and glycerolipid intermediates occurs on the cytoplasmic surface of rat liver microsomal vesicles. Biochim Biophys Acta. 1981;665:586–95. doi: 10.1016/0005-2760(81)90274-5. [DOI] [PubMed] [Google Scholar]
- 19.Leung PH, Zhang QX, Wu JY. Mycelium cultivation, chemical composition and antitumour activity of a Tolypocladium sp. fungus isolated from wild Cordyceps sinensis. J Appl Microbiol. 2006;101:275–83. doi: 10.1111/j.1365-2672.2006.02930.x. [DOI] [PubMed] [Google Scholar]
- 20.Jonas J, Xie CL, Jonas A, Grandinetti PJ, Campbell D, Driscoll D. High-resolution 13C NMR study of pressure effects on the main phase transition in L-alpha-dipalmitoyl phosphatidylcholine vesicles. Proc Natl Acad Sci U S A. 1988;85:4115–7. doi: 10.1073/pnas.85.12.4115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Oh J, Yu T, Choi SJ, Yang Y, Baek HS, An SA, et al. Syk/Src pathway-targeted inhibition of skin inflammatory responses by carnosic acid. Mediators Inflamm 2012. 2012 doi: 10.1155/2012/781375. 781375. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Seok Yang W, Lee J, Woong Kim T, Hye Kim J, Lee S, Hee Rhee M, et al. Src/NF-κB-targeted inhibition of LPS-induced macrophage activation and dextran sodium sulphate-induced colitis by Archidendron clypearia methanol extract. J Ethnopharmacol. 2012;142:287–93. doi: 10.1016/j.jep.2012.04.026. [DOI] [PubMed] [Google Scholar]
- 23.Yu T, Shim J, Yang Y, Byeon SE, Kim JH, Rho HS, et al. 3-(4-(tert-Octyl)-phenoxy) propane-1,2-diol suppresses inflammatory responses via inhibition of multiple kinases. Biochem Pharmacol. 2012;83:1540–51. doi: 10.1016/j.bcp.2012.02.018. [DOI] [PubMed] [Google Scholar]
- 24.Yang Y, Yu T, Jang HJ, Byeon SE, Song SY, Lee BH, et al. In vitro and in vivo anti-inflammatory activities of Polygonum hydropiper methanol extract. J Ethnopharmacol. 2012;139:616–25. doi: 10.1016/j.jep.2011.12.003. [DOI] [PubMed] [Google Scholar]
- 25.Kim HG, Shrestha B, Lim SY, Yoon DH, Chang WC, Shin DJ, et al. Cordycepin inhibits lipopolysaccharide-induced inflammation by the suppression of NF-kappaB through Akt and p38 inhibition in RAW 264. 7 macrophage cells. Eur J Pharmacol. 2006;545:192–9. doi: 10.1016/j.ejphar.2006.06.047. [DOI] [PubMed] [Google Scholar]
- 26.Wang XA, Xiang SS, Li HF, Wu XS, Li ML, Shu YJ, et al. Cordycepin induces S phase arrest and apoptosis in human gallbladder cancer cells. Molecules. 2014;19:11350–65. doi: 10.3390/molecules190811350. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Tuli HS, Sharma AK, Sandhu SS, Kashyap D. Cordycepin: A bioactive metabolite with therapeutic potential. Life Sci. 2013;93:863–9. doi: 10.1016/j.lfs.2013.09.030. [DOI] [PubMed] [Google Scholar]
- 28.Tuli HS, Sandhu SS, Sharma AK. Pharmacological and therapeutic potential of Cordyceps with special reference to Cordycepin. 3 Biotech. 2013;4:1–12. doi: 10.1007/s13205-013-0121-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Jeong JW, Jin CY, Park C, Han MH, Kim GY, Moon SK, et al. Inhibition of migration and invasion of LNCaP human prostate carcinoma cells by Cordycepin through inactivation of Akt. Int J Oncol. 2012;40:1697–704. doi: 10.3892/ijo.2012.1332. [DOI] [PubMed] [Google Scholar]