

Abstract
Complete sex chromosome dosage compensation has more often been observed in XY than ZW species. In this study, using a population genetic model and the chicken transcriptome, we assess whether sexual conflict can account for this difference. Sexual conflict over expression is inevitable when mutation effects are correlated across the sexes, as compensatory mutations in the heterogametic sex lead to hyperexpression in the homogametic sex. Coupled with stronger selection and greater reproductive variance in males, this results in slower and less complete evolution of Z compared with X dosage compensation. Using expression variance as a measure of selection strength, we find that, as predicted by the model, dosage compensation in the chicken is most pronounced in genes that are under strong selection biased towards females. Our study explains the pattern of weak dosage compensation in ZW systems, and suggests that sexual selection plays a major role in shaping sex chromosome dosage compensation.
 Complete sex chromosome dosage compensation is largely limited to male heterogametic species, with the majority of female heterogametic species displaying incomplete dosage compensation. Here, the authors show that sexual conflict over gene expression combined with sexual selection in males can explain this pattern.
Complete sex chromosome dosage compensation is largely limited to male heterogametic species, with the majority of female heterogametic species displaying incomplete dosage compensation. Here, the authors show that sexual conflict over gene expression combined with sexual selection in males can explain this pattern.
Sex chromosomes diverge following the suppression of recombination, which results in the decay of the non-recombining Y or W sex chromosomes1. The degradation of Y- and W-linked genes causes a reduction in expression levels in the heterogametic sex2,3, which retains only one copy of X- or Z-linked loci. This decrease in expression can have a negative effect on the fitness of the heterogametic sex (XY in male and ZW in female heterogamety). The deleterious effects of Y and W degradation are thought to drive the evolution of dosage compensation, where selection acts to increase expression in the heterogametic sex and so restore the expression level that existed prior to sex chromosome divergence4,5.
Ohno4 suggested that complete dosage compensation would follow gene loss on the Y or W chromosome, and many assumed that complete dosage compensation was an essential feature of all heteromorphic sex chromosomes. However, expression data shows that dosage compensation varies widely among taxa. Some species, such as Drosophila melanogaster6,7 and Caenorhabditis elegans8,9, achieve near complete dosage compensation across the entire X chromosome. This has been observed in several other male heterogametic (XY) species (Table 1). In contrast, complete dosage parity has only been observed in one female heterogametic (ZW) species10. All other female heterogametic species thus far studied display incomplete dosage compensation, with some individual genes compensated, but the average expression of female Z-linked genes below that of the two copies in males, and below the average expression of autosomal copies in females (Table 1, Supplementary Table 1). Age of sex chromosome system does not explain the distribution of incomplete versus complete dosage compensation, as both birds and Lepidoptera possess relatively old, highly conserved sex chromosome systems that are largely uncompensated11. The term incomplete dosage compensation in Table 1 includes several different possible regulatory states, ranging from dosage compensation for a subset of genes to partial compensation for a large proportion of genes. Although it is not possible to differentiate these various forms of incomplete dosage compensation, the striking difference between male and female heterogamety suggests that the evolution of dosage compensation is influenced by different forces in X and Z chromosomes.
Table 1. Current status of dosage compensation.
| Species or clade | Karyotype (female:male) | Average sex chromosome dosage compensation |
|---|---|---|
| Caenorhabditis elegans | XX:XO | Complete |
| Drosophila melanogaster | XX:XY | Complete |
| Teleopsis dalmani | XX:XY | Complete |
| Anopheles gambiae | XX:XY | Complete |
| Tribolium castaneum* | XX:XY | Complete in males |
| Xenos vesparum | XX:XY | Incomplete in new region, complete in old region shared with Tribolium |
| Gasterosteus aculeatus | XX:XY | Incomplete |
| Ornithorhynchus anatinus† | XX:XY | Incomplete |
| Monodelphis domestica | XX:XY | Complete |
| Eutherian mammals‡ | XX:XY | Complete for dosage sensitive genes |
| Silene latifolia | XX:XY | Unclear |
| Rumex hastatulus | XX:XY | Incomplete |
| Schistosoma mansoni | ZW:ZZ | Incomplete |
| Lepidoptera | ZW:ZZ or ZO:ZZ | Ranges from incomplete to complete |
| Cynoglossus semilaevis | ZW:ZZ | Incomplete |
| Serpentes | ZW:ZZ | Incomplete |
| Aves | ZW:ZZ | Incomplete |
Recent studies (see Supplementary Table 1 for references) have assessed the presence or absence of complete dosage compensation by comparing the average X or Z expression to the average autosomal expression in the heterogametic (Xmale:AAmale or Zfemale:AAfemale) and homogametic (XXfemale:AAfemale or ZZmale:AAmale) sex.
*Tribolium exhibits X chromosome dosage compensation in males, although this appears to have resulted in hyperexpression of X-linked genes in females47.
†The Platypus has five separate X chromosomes (X1, X2, X3, X4, X5), which are diploid in females and matched by five Y chromosomes (Y1, Y2, Y3, Y4, Y5) in males.
‡Recent work in the therian mammals indicates that dosage-sensitive genes are compensated in both sexes48, through the evolution of female X chromosome inactivation and upregulation of the single active X in both sexes. However, X:AA and XX:AA ratios are <1 for a small number of dosage-insensitive genes and genes with active Y homologues.
Here we build a population genetic model in which dosage compensation arises through the accumulation of cis-regulatory mutations that alter the expression of X- or Z-linked genes12. To be consistent with our subsequent data analysis, we define dosage compensation as the evolution of equal expression in males and females following the loss of the Y or W allele in the heterogametic sex. As correlations in expression between the sexes are typically strongly positive13,14, we allow mutations altering expression in one sex to cause correlated change in the other sex. Therefore, mutations that increase expression in the heterogametic sex to compensate for reduced unbalanced selection between the sexes may lead to hyperexpression in the homogametic sex, resulting in sexually antagonistic fitness effects. When there is sexual conflict, the adaptive response is dominated by the sex under stronger selection15,16. In addition, in the case of sex chromosomes, selection is greater in the homogametic sex relative to that in the heterogametic sex17. As the Z chromosome resides two-thirds of the time in males while the X chromosomes resides two-thirds of the time in females, selection on males has a disproportionate effect on the Z and selection on females has a disproportionate effect on the X. Finally, the efficacy of selection on a mutation can differ according to its location on the Z or the X chromosome. Reproductive variance may vary between the sexes, and is often higher in males18,19. This affects Z and X chromosomes differently, as the Z chromosome is more sensitive to increased male reproductive variance than the X16,20,21,22, lowering the rate of adaptation on the Z relative to that on the X. We examine how the combination of these forces results in differences between XY and ZW systems in the evolutionary rates of dosage compensation, and find that slower and less complete Z dosage compensation evolution is associated with a positive correlation in mutational effects across the sexes, stronger selection on males and greater male reproductive variance. Our model also predicts that dosage compensation should be most pronounced in genes that are under strong selection, biased towards the heterogametic sex. Using expression variance as a measure of selection strength on expression, we find support for these predictions from the chicken transcriptome.
Results
Rates of dosage compensation evolution
In the model (see Methods for details), expression evolves via the recurrent fixation of cis-regulatory mutations of small effect, consistent with previous studies of gene expression evolution23. We assume that mutations are sufficiently rare for each one to be fixed or lost before another arises. Mutations cause small changes δm and δf in male and female expression relative to the resident allele (Table 2). We assume that the variance of mutational effects δm and δf is fixed to a constant value (Var[δm]=Var[δf]=0.1). The scope for sexually antagonistic effects is determined by the correlation ρ between the effects of mutation in the two sexes, which can be independent (ρ=0), similar (ρ>0) or opposed (ρ<0). The model assumes that gene expression in the homogametic sex is additive and taken to be the sum of the effects of the alleles carried by an individual (Table 2). Expression is assumed to be under stabilizing selection, with the optimum expression arbitrarily set to zero and symmetric decline following a Gaussian function (Table 2). Although stabilizing selection operates in both sexes, its intensity in males and females, denoted by Sm and Sf, may differ (Table 2).
Table 2. Sex-specific expression in male and female heterogametic systems.
|
Male |
Female |
|||||
|---|---|---|---|---|---|---|
| Genotype | Expression | Fitness | Genotype | Expression | Fitness | |
| Male heterogamety (XY) | R | zm |  |
RR | zf | 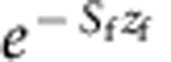 |
| RM | zf+δf | 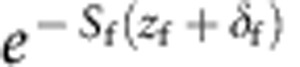 |
||||
| M | zm+δm | 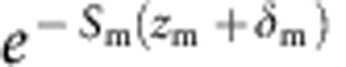 |
MM | zf+2δf | 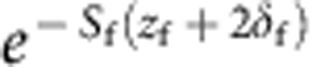 |
|
| Female heterogamety (ZW) | RR | zm | 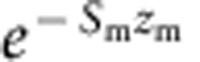 |
R | zf | 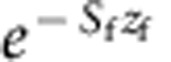 |
| RM | zm+δm | 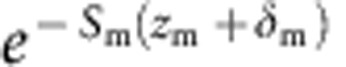 |
||||
| MM | zm+2δm | 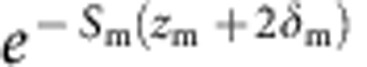 |
M | zf+δf | 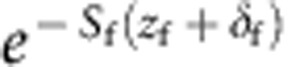 |
|
The resident allele (R) has expression level zm in males and zf in females. The mutant allele (M) causes a small quantitative shift of δm and δf in males and females, respectively.
Assuming that gene expression is at the optimum before degradation of the Y or W allele occurs, the heterogametic sex initially suffers a decrease of −z0 in expression, where z0 is the expression contribution of the Y/W chromosome prior to degradation. Expression in the homogametic sex is unaffected and remains at the optimum level. The evolutionary trajectory from this starting point depends on the fixation probability of newly arising mutations. This depends on the sex-specific selection pressure acting on a mutant, given by Sm and Sf, and the effective population size NeX or NeZ, which scale with the level of sex-specific variance in reproductive success24. Iterating this mutation-fixation process and integrating across the distribution of mutational effects δm and δf, the expected rates of change in male (zm) and female (zf) expression are given by
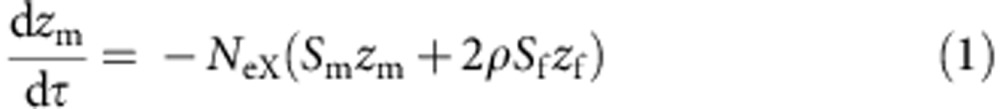 |
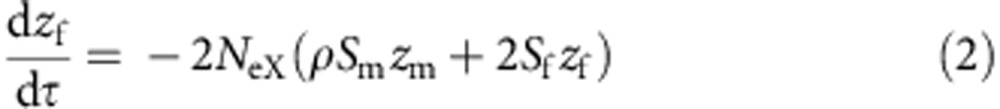 |
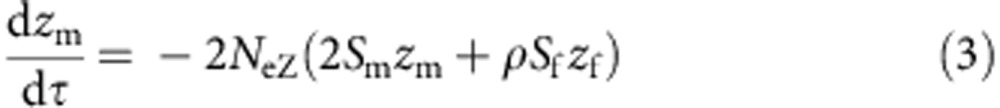 |
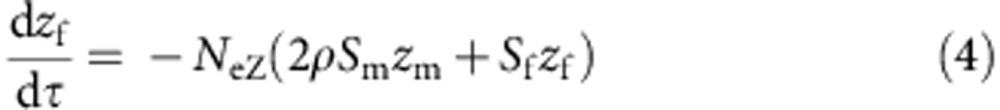 |
for XY (equations (1) and (2)) and ZW (equations (3) and (4)) species. Evolutionary time τ in equations (1)–(4), , , is scaled with respect to the mutation rate. This means that a unit of τ corresponds to fewer generations when the mutation rate is higher. Equations (1)–(4), , , highlight how the evolution of X- and Z-linked gene expression depends on the asymmetry in sex chromosome dose (two in the homogametic and one in the heterogametic sex), the inter-sexual correlation in expression (ρ), the balance between selection in males and females (Sm and Sf) and the effective population size (NeX and NeZ).
Given sufficient time, the model predicts that both XY and ZW systems will evolve complete dosage compensation for genes whose expression levels depend on gene dose, where expression in the heterogametic sex converges to the ancestral level and males and females reach their respective fitness optima (zm=0 and zf=0).
In the absence of any inter-sexual correlation in mutational effects (ρ=0), the evolutionary dynamics of dosage compensation are governed by the strength of selection on the heterogametic sex alone. The heterogametic sex returns to the optimal level at an exponential rate 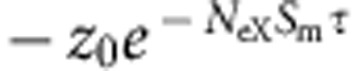 in XY species, and
in XY species, and 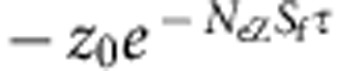 in ZW species. If the effective population sizes and the mutation rates are the same for the X and Z chromosomes and selection is equivalent in males and females (Sm=Sf), complete dosage compensation will be attained at the same rate in both XY and ZW systems.
in ZW species. If the effective population sizes and the mutation rates are the same for the X and Z chromosomes and selection is equivalent in males and females (Sm=Sf), complete dosage compensation will be attained at the same rate in both XY and ZW systems.
The speed with which dosage compensation evolves is dramatically reduced under the more plausible assumption that mutations have correlated effects across the sexes (ρ≠0). When mutational effects are positively correlated (ρ>0), mutations that increase expression and improve dosage compensation in the heterogametic sex are associated with antagonistic fitness effects in the other sex, as they raise expression in the homogametic sex above the fitness optimum. As the homogametic sex carries more chromosomal copies, antagonistic effects impede selection in favour of dosage compensation in the heterogametic sex. If selection is equal between males and females (Sm=Sf), the expectation is that dosage compensation will be slow to evolve both for Z- (Fig. 1a) and X-linked genes (Fig. 1b).
Figure 1. The evolution of dosage compensation on Z and X chromosomes.

Expected gene expression is shown for males (blue) and females (red) under a very low (ρ=0.1, dotted line), intermediate (ρ=0.5, dashed line) and strong inter-sexual correlation (ρ=0.8, solid line)—(see equations (13) and (14) for dynamics). a,b show equal selection in males and females (Sm=Sf=0.5); c,d show stronger selection in males (Sm=1, Sf=0.1). Evolutionary time refers to the number of generations, ignoring the time taken by successive mutations to fix. Expression is scaled according to the initial degradation z0 of expression in the heterogametic sex due to the loss of one gene copy, which here is set at z0=1. Other parameters were also held equal across the two sex chromosome systems (NeX=NeZ=1125, μ=0.0003).
However, selection is typically stronger in males (Sm>Sf) as a result of sexual selection25,26,27,28. This creates markedly different evolutionary trajectories of dosage compensation. For Z-linked genes, the combination of male homogamety with more intense selection in males maintains male expression near its optimum (Fig. 1c). Only mutations that increase female expression while minimally affecting males are favoured (Fig. 1c). With a positive inter-sexual correlation for mutational effects (ρ>0), the frequency with which such suitable mutations arise is low and the rate at which dosage compensation evolves is very slow (Fig. 1c).
The slow and gradual trajectory of Z-linked genes contrasts with more rapid and sexually antagonistic dynamics for X-linked genes (Fig. 1d). Here more intense selection on males (Sm>Sf) compensates for male X chromosome monosomy, and the evolution of dosage compensation proceeds rapidly. But it does so at a pronounced cost to females, which are displaced from their optimal expression (Fig. 1d). This effect is greater the higher the correlation of mutational effects (ρ). Once males approach their optimum expression level, the selective advantage of further change in male expression decreases, and selection on females becomes increasingly important. This causes female expression to slowly evolve back towards the optimum through the fixation of rare mutations that simultaneously relieve the antagonistic effects on female expression while allowing male expression to remain close to its optimum.
Our baseline model assumes additive effects on expression and the instantaneous loss of the W or Y allele. However, we also consider dominance effects on expression and progressive loss of the W or Y allele (see Methods for details). Neither of these variants change our result that dosage compensation evolves faster on the X than on the Z in the presence of more intense selection on males and positive inter-sexual correlation of mutational effects (Supplementary Figs 1 and 2). However, in both cases, the evolutionary trajectory of X expression in females is altered compared with our baseline model. Because selection is more efficient at purging dominant deleterious alleles, the overshoot of X expression in females is mitigated in the presence of dominant mutations (Supplementary Fig. 1). Also, when expression of the Y allele slowly degrades, selection on males to compensate is weaker, and as a result causes weaker detrimental effects to females, whose X expression remains closer to its optimal value (Supplementary Table 2, Supplementary Fig. 2).
The role of effective population size
The evolutionary dynamics at Z- and X-linked genes also depend on the chromosomes' effective population sizes NeZ and NeX in equations (1)–(4), , , . Larger Ne results in faster evolution of dosage compensation, due to the more reliable fixation of positively selected mutations and elimination of negatively selected mutations (Fig. 2). With effective population size inversely proportional to the stochasticity in replicate evolutionary paths, small values of Ne not only slow down the evolution of dosage compensation but also result in more variable expression levels across replicate evolutionary trajectories. This effect is observed both during adaptation (Fig. 2), as well as at the evolutionary equilibrium (equation (15)).
Figure 2. Variance in male reproductive success and the evolution of dosage compensation.

Stochastic evolutionary trajectories of male (blue) and female (red) expression for (a) Z-linked and (b) X-linked genes (see equations (11) and (12) for dynamics). Levels of expression are shown for low (η=1, light shade), intermediate (η=3, intermediate shade) or high (η=10, dark shade) degrees of male reproductive variance (the number of successful breeding females is set at 1,000). A sample of 5 trajectories is shown for each parameter value. Variance in heterogametic sex expression for (c) Z-linked and (d) X-linked genes after the end of the simulation is shown as a frequency distribution for 1,000 replicates. Selection is stronger on males than on females (Sm=1, Sf=0.2), inter-sexual correlation is relatively strong (ρ=0.6) and the mutation rate is μ=0.0003.
Differences in ecology or mating systems (and hence Ne) could underlie variation in the rate at which dosage compensation evolves. Under simplifying assumptions29, the effective population sizes of the Z and X chromosomes in a population reproducing under harem polygamy are NeZ=9Nf/(6+4η) and NeX=9Nf/(6+2η), where Nf is the number of successfully breeding females and η is the ratio of the number of successfully breeding females to successfully breeding males. As two-thirds of the Z chromosomes are transmitted by males each generation, compared with only one-third of X chromosomes, greater variance in male reproductive success (η>1) leads to a more marked diminution of NeZ than NeX. This predicts slower and more stochastic evolution of dosage compensation on the Z than on the X (Fig. 2).
Empirical tests of model predictions
The model predicts that whenever selection is more intense on males (Sm>Sf), the evolution of dosage compensation is more rapid and efficient in species with XY as opposed to ZW sex determining systems. This expectation fits broadly with the limited amount of available data on average sex chromosomal expression levels (Table 1). A more rigorous and powerful test can be performed by analysing variation in individual gene expression patterns within sex chromosomes. In particular, an important model prediction is that dosage compensation should be more prevalent among genes subject to strong selection, and in genes where selection is stronger in the heterogametic sex. We addressed these questions using transcriptome data from chicken, a species previously shown to have incomplete Z chromosome dosage compensation30. To gauge the strength of selection on individual genes in the two sexes, we measured the sex-specific variability of transcript levels due to differences in expression levels between biological replicates (biological coefficient of variation (BCV); see Methods)31,32. The assumption is that genes under stronger purifying selection show less expression variation between biological samples. Our approach follows an increasing number of studies using variance as an indicator of selection on gene expression levels31,32,33,34,35. It is important to note, however, that the assumptions have not been fully validated experimentally, and so should be interpreted with caution.
A set of preliminary analyses supported our estimates of BCV as informative measures for selection strength. First, we observed a negative relationship between BCV and average expression in all tissues and both sexes (Supplementary Table 3). This shows that our estimation procedure successfully removed the positive association between mean and variance that is expected in Poisson-distributed read count data. The negative association is also in line with data from other species36 showing that more highly expressed genes are under tighter selective constraint. Second, we also observed a positive association between BCV and the rate of protein evolution (measured as the rate of non-synonymous to synonymous substitutions, dN/dS, along the terminal chicken branch; Supplementary Tables 4 and 5). Such an association has been previously observed in vertebrates37,38,39 and insects33,40,41 and implies that genes under more intense purifying selection on expression level also tend to be under stronger purifying selection on protein sequence. Importantly, the positive relationship between expression variance and dN/dS occurs over and above a negative association between average expression level and rates of protein evolution that is observed in other species42.
Comparing estimates of male and female BCV, we find that across all autosomal and Z-linked genes expression variation is greater in females than males for all three tissues we analysed (Wilcoxon test on difference between male and female BCV; liver: V=851,261, P<0.0001, N=9,196; heart: V=21,797,411, P<0.0001, N=9,790; gonad: V=1,240,318, P<0.0001, N=11210). The same difference is also found when analysing autosomal genes alone (liver: V=698,999, P<0.0001, N=8,333; heart: V=17,969,183, P<0.0001, N=8,857; gonad: V=971,978, P<0.0001, N=10,086). These results support the notion that selection is more intense on male expression.
We analysed whether the degree of dosage compensation of individual Z-linked genes (measured as log2 of the ratio of male-to-female expression, see Methods) is related to our measure of selection strength on expression in the two sexes. Because the strength of selection in males and females is positively correlated across loci, we first transformed our measures of male and female expression variability into two perpendicular components, one that measures the strength of sexually concordant selection on expression and the other that measures male (or female) bias in selection intensity. We then related the pattern of dosage compensation across genes expressed in three tissues (heart, liver and gonad) to their value along these axes of selection.
We found that both measures of selection have a significant effect on the male-to-female expression ratio, but that this effect differs between the tissues (tissue-by-selection interactions in Supplementary Table 6). The nature of these differences is best illustrated by analyses run separately on data liver, heart and gonad. In liver, we found that the male-to-female expression ratio decreases with the strength of concordant selection and increases with male-biased selection (Supplementary Table 7, Fig. 3). A significant interaction between the two selection measures further showed that the effect of male-bias in selection intensity is more pronounced when the strength of concordant selection is weak. This means that, as predicted by our model, dosage compensation is most pronounced in genes that are under strong selection in both sexes and those where selection intensity is biased towards females. In contrast to liver, genes expressed in heart showed no relationship between male-to-female expression ratio and either selection measure, while in the gonads we detect only a weak signal for a positive relationship between male-to-female expression ratio and concordant selection strength (Supplementary Table 7, Supplementary Fig. 3).
Figure 3. Selection on gene expression and dosage compensation in the chicken liver.

The figure shows the strength of sexually concordant selection and the measure of male bias in selection strength for liver-expressed genes used in our analysis. Dosage compensated genes (0.8<male-to-female expression ratio<1.2) are shown in red, and non-dosage compensated genes are shown in grey.
The model makes the additional prediction that the degree of dosage compensation should increase over time. We assessed this by comparing male-to-female expression ratios across three well-supported strata identified within the chicken Z chromosome. These have been estimated to have diverged from the W chromosome for 75–106 mya (Stratum 1), 45–71 mya (Stratum 2) and 36–68 mya (Stratum 3)43. Our model makes the qualitative prediction that if dosage compensation evolves slowly (relative to the age of these strata), then genes on older strata should show more similar (less male biased) expression between the sexes than genes on younger strata. However, our data show no relationship between dosage compensation and the age of the strata (Supplementary Table 8). We obtained qualitatively identical results when we examined the female Z to autosomal expression across strata (Supplementary Table 9); however, the power of these analyses is limited due to the low number of genes in each stratum. This suggests that the evolution of effective dosage compensation occurs within a time range shorter than the age of the youngest chicken stratum (36–68 mya). Furthermore, the persistence of many genes that lack dosage compensation across the strata and the absence of a decrease in their frequency through time suggests either that the genes without dosage compensation have no or little effect on fitness or that persistent sex differences in their expression are accommodated by changes elsewhere in the genome.
Discussion
Our model predicts that XY and ZW systems can differ substantially in the rate with which dosage compensation evolves and the trajectory taken by male and female expression over the course of evolution. In particular, the evolution of dosage compensation will be slower and more stochastic in ZW than XY systems whenever the inter-sexual correlation in mutational effects is strongly positive and coupled to greater selection in males and/or higher variance in male mating success (Figs 1 and 2).
The conditions that are predicted to lead to different evolutionary trajectories of X- and Z-dosage compensation are biologically plausible. First, more intense selection in males due to sexual selection is a standard expectation supported by much empirical evidence19 and this is also supported by our chicken transcriptome data. Importantly, the greater intensity of male selection potentially affects a large number of genes. Thus, genes under sexual selection need not directly code for male reproductive traits, such as ornaments or weaponry. More often than not, male reproductive success depends on general condition44, which has a much broader genetic basis, including genes with general effects on viability. In many cases, these genes are also important for female function and therefore under selection for similar expression in both sexes. However, sexual selection tightens the selection pressure for expression in males to be close to the optimum compared with selection on female expression. Second, the situation where sexual selection on males is associated with higher variance in male mating success is frequent in natural populations18,19. And finally, it is plausible to expect a strong positive correlation of mutational effects on gene expression in males and females. The sexes typically show positive genetic correlations for the expression of genes (for example, refs 13, 14) and new mutations tend to have positively correlated effects on male and female fitness and phenotypes45,46.
The predictions of our model find some support in our analysis of expression variation on the chicken Z chromosome. Our analyses show that in the liver, the degree of dosage compensation of individual genes is more pronounced in genes that are presumably under strong selection in both sexes, and therefore show reduced expression variance, and where the intensity of selection is greater in females than males (Fig. 3, Supplementary Table 7). Similar relationships were not detected in the other two tissues analysed. In the gonad, expression evolution is likely to be strongly influenced by sex-specific selection pressures that could mask the effects we are interested in. However, the difference between liver and heart is not straightforward to explain. It is conceivable that the function of the liver is more sensitive to changes in gene expression. In line with this argument, liver-expressed genes are on average more dosage compensated than those expressed in the heart (results not shown). More research is needed to elucidate the ultimate causes for tissue differences in regulatory evolution.
Our analysis of gene expression on the chicken Z chromosome also revealed that the prevalence of dosage compensation is independent of the age of evolutionary strata in which the genes reside. This suggests that dosage compensation is saturated within the time-frame of the youngest stratum of the chicken Z chromosome (36–68 mya) and there does not appear to be compensation of additional loci beyond this time. One possible reason for this is that the expression of other Z-linked genes is not or is no longer sensitive to gene copy number; perhaps because other unlinked compensatory changes have already taken place. Alternatively, selection against hypertranscription in the homogametic sex may be sufficiently strong to retard further evolution of compensation in the heterogametic sex. In addition, it is possible that the low effective population size of the Z chromosome limits the efficacy of selection for dosage compensation, and slows its evolution in genes whose expression only mildly depends on dosage, perhaps again favouring the evolution of compensatory mechanisms elsewhere in the genome. However, we note that the youngest stratum in chicken is several tens of millions of years old, so is not young on an absolute scale. Further studies are needed in species with younger sex chromosomes or younger strata to investigate how quickly a plateau is reached in the rate of dosage compensation evolution of Z chromosomes.
The model shows that dosage compensation on the X chromosome evolves via a phase of sexually antagonistic gene expression due to a positive inter-sexual correlation in mutational effects and stronger selection on males. Selection on the heterogametic sex to increase expression towards the optimum is predicted to result in overshooting of the optimum in the homogametic sex. There is some evidence for this in the flour beetle Tribolium castaneum, where the female X chromosome is thought to be hypertranscribed relative to autosomal expression levels but the X and autosomes are in dosage parity in males47. Sexual conflict over optimal expression may also be the biological explanation for compensatory mechanisms that reduce expression of the X in the homogametic sex. Female-limited mechanisms that re-establish expression parity between the female X chromosomes and autosomes are selected once XY males have approached the optimum expression level (Fig. 1). Female X inactivation in eutherian mammals48 and the Dpy complex in C. elegans hermaphrodites49 might be examples of such mechanisms, as could be the repression of X hypertranscription during the embryogenesis of female Drosophila50.
The model also predicts an important role for reproductive variance and genetic drift in generating differences between species with XY systems and those with ZW systems. In monogamous species, the sex-specific reproductive variance is more often close to equal between males and females (η≈1), resulting in similar sex chromosome to autosomal effective population sizes for X and Z chromosomes (NeX/NeA≈NeZ/NeA≈3/4). Based on our model, we then expect dosage compensation to evolve at similar rates on the X and Z. In polygynous species, however, the greater reproductive skew in males decreases the effective size of the Z much more than that of the X, as there are two copies of the Z in males (NeX/NeA>3/4, NeZ/NeA<3/4; ref. 51). This asymmetric change in effective population sizes is expected to result in more rapid and less stochastic evolution of dosage compensation on the X compared with the Z (Fig. 2).
Unlike differences in selection that act on individual genes on the X and Z, differences in the mating system, and hence NeX and NeZ, affect the evolution of dosage compensation across the whole X and Z chromosome. They are therefore more apt at explaining differences in average chromosome-wide dosage compensation between species. Our preliminary comparison suggests that dosage compensation is more pronounced in species with XY systems than those with ZW systems (Table 1). Testing the contribution of reproductive variance and genetic drift will require improved comparative analyses, on a larger sample of XY, as well as ZW species that show lineage-specific variation in mating systems. One complication of such analyses will be that the average difference in selection intensity between the sexes is expected to covary with the mating system. Polygynous species are expected to feature increased male reproductive variance, as well as more intense selection on males. As a consequence, disentangling the effects of selection and drift may not be straightforward.
We used a number of assumptions in our modelling to reach tractable results (considered further in the Supplementary Discussion). The model's predictions show a striking similarity to currently described patterns of dosage compensation, and to differences in dosage compensation among genes on the chicken Z. This suggests that the marked difference in dosage compensation between male and female heterogametic species can be explained by the combination of genetic correlation for expression across the sexes, with greater male specific selection and high male reproductive variance. Sexual selection on males simultaneously increases the strength of selection and reproductive variance in males, and thus provides a simple and general explanation for the differences in the evolution of dosage compensation.
Methods
Population genetic model of dosage compensation evolution
We model the co-evolution of male and female gene expression level for X- and Z-linked dose-sensitive genes. It is assumed that the Y and W copies have undergone degradation and have no relevant gene expression. In the following, we consider X-linked genes; the Z-linked case is derived in a similar manner.
Gene expression evolves by the recurrent substitutions of mutations that have small sex-specific effects, in a population that in effect remains monomorphic (for example, see refs 52, 53). The population is initially genetically homogeneous, and expression of the X-linked gene is set at zm in males and zf in females. A cis-mutation arises and changes male and female expression by δm and δf respectively (Table 2). Mutant effects on expression are additive, in agreement with data for a large proportion of genes54, and appropriate to model dosage compensation, which can be generalized as a cis-regulatory additive mechanism55,56. Mutations are assumed to be sufficiently rare for one mutation to be fixed or lost before another arises52. So we can characterize the expected change in expression in the population by considering only the probability of fixation of the mutant, which depends on the life cycle of the population and the fitness effects of the mutation.
The population is composed of N males and N females, and the life cycle follows a standard Wright–Fisher process57. Generations are non-overlapping. Male and female adults produce large numbers of gametes that are randomly paired to produce zygotes. The zygotes are then sampled with replacement and with a selective bias to form the males and females of the next generation. Since the population evolves according to a Wright–Fisher process and selection is weak, we can use Kimura's equation for sex-linked genes to determine the fate of the mutants57.
Mutation changes male and female expression level. Whether a mutant is selectively favoured depends on its effects on male and female fitness. It is assumed that there is an optimum expression around which fitness declines. The rate at which fitness decreases from the optimum may differ across the sexes. More precisely, male and female fitness are given as Gaussian functions of the level z of expression of the gene, with fitness 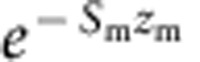 in males and by
in males and by 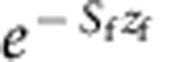 in females, and the optimum arbitrarily set at zero (zm=0, zf=0). Greater values for Sm and Sf reflect stronger sex-specific stabilizing selection. It is possible to have different optimal levels in males and females; however, this does not change the qualitative features of our observations.
in females, and the optimum arbitrarily set at zero (zm=0, zf=0). Greater values for Sm and Sf reflect stronger sex-specific stabilizing selection. It is possible to have different optimal levels in males and females; however, this does not change the qualitative features of our observations.
If the effects of mutations on the level of expression (δm, δf) are small and the population N is large (so that the initial mutation frequency is small), it is reasonable to linearize Kimura's equation for the probability that a single copy X-linked mutant fixes in the population57,
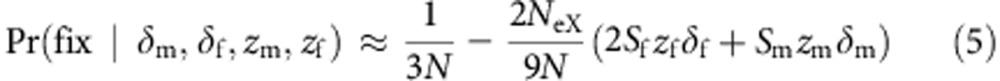 |
where NeX is the effective population size of the X chromosome, and captures the effect of genetic drift, and N is the actual population size, capturing the initial frequency of a mutant. The first term of equation (5) is the probability of fixation of a neutral X-linked mutant. The terms inside the bracket are the effects of selection on females and males. Selection increases the probability of fixation if the mutant pushes the expression rate closer to the optimal level (that is, if zfδf<0 or zmδm<0). The increase is proportional to the strength of selection in each sex Sf and Sm, but is twice as strong in females as they have twice as many X chromosomes compared with males. This lends females a greater importance in the spread of X mutants.
The probability of fixation determines the expected change in expression over one particular substitution in each sex. If the mutant is lost, expression does not change, if the mutant is fixed, male and female expression become zm+δm and zf+2δf, respectively (Table 2). To calculate the expected change in expression, we consider all possible mutational sizes and their frequencies,
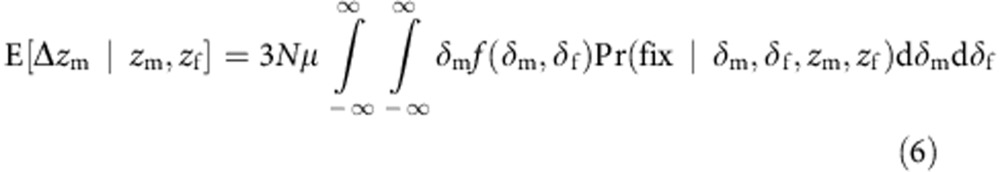 |
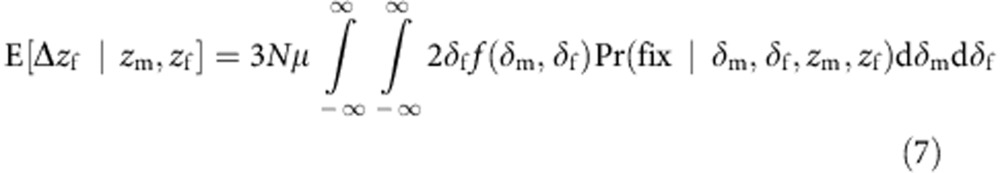 |
where 3N is the total number of genes in the population, μ is the mutation rate and f(δm,δf) is the probability density function for the effects of mutants on male and female expression. We assume that f(δm,δf) is a bivariate normal distribution with mean (0, 0) and covariance matrix
 |
In this case, equations (6) and (7) read
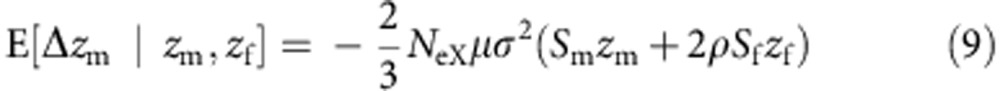 |
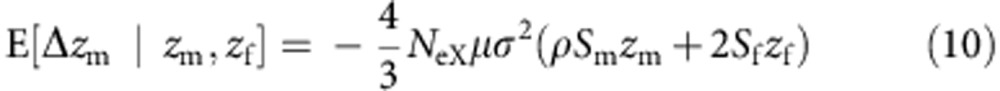 |
and the second moments of the change in male and female expressions are approximately E[(Δzm)2|zm, zf]=E[(Δzf)2|zm, zf ]=σ2, and E[(Δzm)(Δzf )|zm, zf ]=ρσ2. For simplicity, we set σ2=0.1. Setting different values for σ2 changes the impact of mutations, but since it does so with the same intensity in males and females, it does not change our results. However, increasing σ2 increases the overall rate of evolution.
Since selection is weak, the first (equations (9) and (10)) and second moments of the distribution of infinitesimal change in male and female expression are sufficient to describe the evolution of male and female expression over many substitutions52,53. In continuous time, and ignoring the time taken for segregation to occur and the time between mutations, we obtain that the random variables Zm(t) and Zf(t), which describe male and female expression at time t, satisfy the stochastic differential equation,
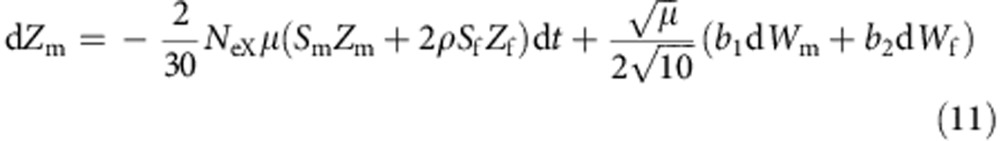 |
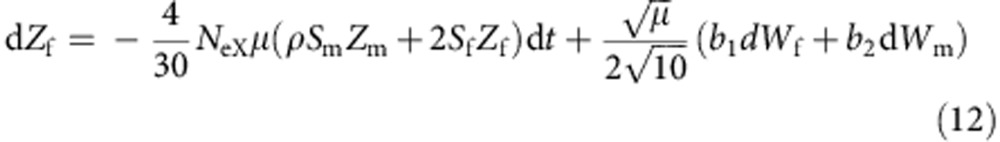 |
where dWm and dWf are standard independent Brownian motions, and b1 and b2 are scaled variance terms given by 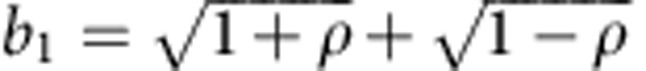 and
and 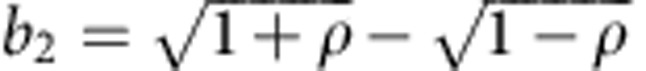 .
.
Unless selection on male and female expression is very weak (very small NeXSm, NeXSf), and evolution is dominated by genetic drift, the qualitative features of the evolution of dosage compensation are captured by the expected trajectory of equations (11) and (12). Writing the expected male and female expression as zm and zf, we find that their evolution is given by equations (1) and (2), where time is rescaled according to τ=2μt/30. This scaling eases the comparison between X and Z expression when differences between mutation rates in the two systems are ignored.
The expected values for male and female expression through time, zm(τ) and zf(τ), may be found exactly by solving equations (1) and (2). If we assume that degradation of the Y-linked gene copy leads to an initial diminution in expression in males of −z0, and female expression is unperturbed by degradation and remains at the ancestral optimal level zero, we have
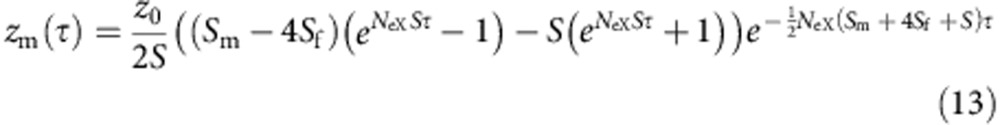 |
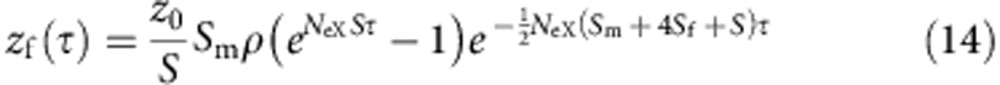 |
where 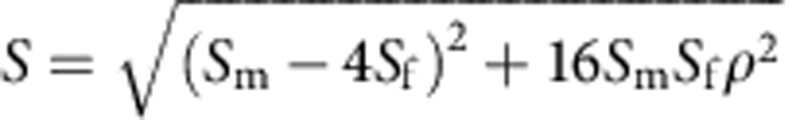 . Similar expressions for Z-linked genes can be found by replacing NeX by NeZ, and m subscripts by f and vice versa. Plots in Fig. 1 correspond to equations (13) and (14) with respect to t rather than τ to compare more easily the deterministic paths with the stochastic replicates of equations (11) and (12) that are shown in Fig. 2.
. Similar expressions for Z-linked genes can be found by replacing NeX by NeZ, and m subscripts by f and vice versa. Plots in Fig. 1 correspond to equations (13) and (14) with respect to t rather than τ to compare more easily the deterministic paths with the stochastic replicates of equations (11) and (12) that are shown in Fig. 2.
Using standard results of stochastic systems, the properties of equations (11) and (12) show that after a sufficiently long time, male and female expression will reach an equilibrium distribution that is normally distributed, with mean zero, and a covariance matrix which can be computed explicitly58. In particular, the variance in male expression of the X chromosome is given by
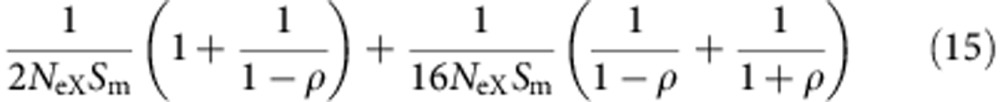 |
The variance in female expression of the Z chromosome is found by replacing NeX by NeZ and by substituting Sf for Sm in equation (15).
Effect of dominant mutations
Our model assumes that the expression of X- and Z-linked genes evolve by the accumulation of cis-acting mutations, which act locally on the sex chromosomes and therefore are expected to have additive effects on overall expression in the homogametic sex (Table 2). Another non-exclusive possibility is that expression evolves by trans-acting mutations. In contrast to cis mutants, trans mutants are likely to affect the expression of both gene copies in the homogametic sex, and therefore correspond to dominant mutants on overall gene expression.
To illustrate the effect of dominant mutations on the evolution of dosage compensation, we also derive the dynamics of male and female expression when mutants are completely dominant so that expression in heterozygotes is equal to expression in mutant homozygotes of the homogametic sex. In that case, the probability of fixation of a dominant mutant of an X-linked gene is
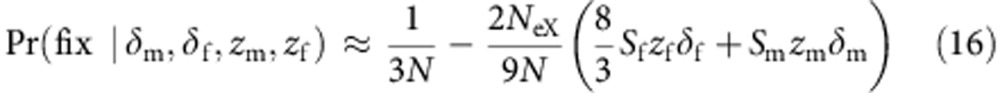 |
In contrast to the additive case (equation (5)), equation (16) shows that selection on the homogametic females is even stronger. For the evolution of dosage compensation in the presence of correlation in mutational effects across males and females, this implies that counter-selection in the homogametic sex is inflated.
Effect of continuous expression decrease in the heterogametic sex
Our model investigates the case where the heterogametic sex suffers a decrease in expression due to the loss of expression of the gene copy on the W or Y. An alternative possibility is that once recombination ceases between the sex chromosomes the loss of expression is a result of gradual gene deterioration. To study the effect of a continuous decrease in expression on the evolution of dosage compensation, a time-dependent decline in the expression level of the W- or Y-gene copy is incorporated.
We consider the case where gene expression in the heterogametic sex is initially equal on both sex chromosomes. Starting from half the optimal level (z0/2), the expression of the W- or Y-gene copy then declines over time t according to
 |
which models an asymptotic decrease approaching total shut-down as time progresses. The parameter α>0 controls the rate of decrease in expression of the gene copy. We can then study cis-regulatory evolution of the X- or Z-linked copy in males and females in the context of the declining Y- or W-linked gene expression. Fitness in the heterogametic sex then depends on expression from the evolving expression of the Z- or X-gene copy and time-dependent level of expression from the W- or Y-gene copy, while fitness in the homogametic sex depends on expression from both Z- or X-gene copies (Supplementary Table 2).
Expression analysis
All samples were collected with approval by the Zoology Ethical Review Committee and in accordance with national guidelines. We obtained fertilized eggs of the White Leghorn chicken breed and kept them under standard incubator conditions. The left gonad, heart and liver were collected at embryonic day 19 and stored in RNAlater (Qiagen) until preparation. RNA from each tissue was prepared with the Animal Tissue RNA Kit (Qiagen). Library and RNA-sequence samples were prepared by the Wellcome Trust Centre for Human Genetics Facility at Oxford University using standard methods. Four samples per sex were prepared for each of the four tissues, and each individual was barcoded so that individual variation in coding sequence and expression level could be tracked. Samples were sequenced using Illumina HiSeq as paired-end 100-bp reads, resulting in 16 million paired-end mappable reads per sample, on average. To minimize technical sources of variance, the same HiSeq machine was used for all samples.
Resulting RNA-Seq data were assessed for quality using FastQC v0.10.1 (ref. 59) and filtered using Trimmomatic v0.22 (ref. 60). Filtered reads were mapped to the chicken reference genome (ENSEMBL version 68 WASHUC2) using TopHat v2.05 (refs 61, 62) and Bowtie2 v2.0.0-beta7 (ref. 63), allowing up to two mismatches per alignment. For each tissue separately, aligned reads were sorted and indexed using Samtools v0.1.18 (ref. 64) and raw read counts extracted using Htseq-count v0.5.4p3 (ref. 59). After removing genes for which no read was detected in any sample, raw read counts were normalized across libraries. We used the weighted trimmed mean of M-values method in EdgeR v3.2.4 (ref. 65). To limit the impact of incomplete dosage compensation on this step, normalization factors were estimated based on autosomal genes only. Relative transcript abundances were then estimated for all expressed genes as reads per kilobase of exon per million mapped reads (RPKM).
Statistical analysis of expression data
To filter out noisy gene expression for each tissue, genes were excluded if they were not expressed in all individuals of at least one sex (log2(RPKM)<1 for any sample)32. For all genes retained, we determined the male-to-female expression ratio as log2(average male RPKM/average female RPKM)=log2(average male RPKM)−log2(average female RPKM). We further estimated the variability of gene expression as an inverse measure of stabilizing selection on expression levels. To quantify expression variability, we used log2 of the BCV as estimated by EdgeR. The package implements Bayesian procedures that—based on a negative binomial model—decompose the across-sample variance in read counts for a gene into a part of technical, Poisson-distributed variance and a part of biological variation that stems from true expression differences between samples. The BCV is the square-root of the estimate biological variation divided by the mean expression level of a gene. We estimated BCVs separately for each tissue and sex. The value of the prior, which governs the relative importance of expression variance across all genes and gene-specific expression variance in the estimation of BCVs, was set to 10. Based on preliminary analysis, this value achieved a good balance between capturing gene-specific effects, while minimizing the number of genes with zero estimates of BCV. Using different priors produced qualitatively equivalent results.
Validation of BCV as a measure for selection strength
We first performed analyses to validate our estimates of BCVs and their interpretation as meaningful measures of selection on expression levels. First, we verified that the estimation of expression variance had successfully removed Poisson-distributed error in read counts by correlating sex- and tissue-specific estimates of BCV with average expression levels across autosomal and Z-linked genes. Next, we verified the relationship between BCV and selective constraint on protein coding sequences, measured as the rate of non-synonymous to synonymous substitutions (dN/dS). Gallus gallus, Meleagris gallopavo, Taeniopygia guttata and Anolis carolinensis complementary DNA sequences were obtained from Ensembl, and orthologues were established using reciprocal blastn66. Orthologous sequences were aligned using PRANK67 and only included in subsequent analyses if the length of the alignment without gaps was >33 amino acids. dN/dS for each gene for the terminal G. gallus branch was estimated using the branch model in the CODEML package in PAML68. Orthologues were excluded if dS>2 as double hits and mutational saturation can lead to inaccurate divergence estimates69. To avoid noise due to genes with poor rate estimates, orthologues were also only included in subsequent analyses if there was at least one non-synonymous or one synonymous substitution, and the total number of substitutions was greater than three on the terminal G. gallus branch.
Because estimates of BCV were negatively correlated with expression levels, and high expression levels have been shown to be associated with selective constraint on coding sequence42, we performed multiple regression analysis of rates of protein evolution (dN/dS) as a function of average expression level (average RPKM across samples) and BCV (all variables log2 transformed). The two predictor variables were entered in this specific order to estimate the association between BCV and rates of protein evolution over and above the effect of average expression levels. Analyses were performed separately for each sex with expression levels and BCV averaged across males and females, as well as separately for each tissue and sex.
Dosage compensation across Z-chromosomal strata
To analyse variation in dosage compensation across the Z chromosome, we filtered genes based on their male-to-female expression ratio. To avoid biasing our analyses by the confounding effects of sex-biased or sex-limited expression, we removed all genes that had strongly sex-biased expression. Specifically, we removed genes with expression ratios >log2 (1.25/0.5) and <log2 (1/1.25). This filtering removed a limited number of genes from the somatic tissues (422 of 433 genes retained for the heart and 364 of 381 retained for the liver), but more from the gonads (427/534 retained). For all genes passing this filter, we projected male and female expression variability (log2 BCV) onto two perpendicular axes, one describing the intensity of sexually concordant selection strength (with values increasing as selection becomes stronger in males and female) and one describing the male bias in selection intensity (values increasing as selection becomes stronger in males and/or weaker in females). The coordinates of the genes on the two axes were then used to determine the effects of selection on male-to-female expression ratios. Transforming selection intensities in this way eliminates problems in subsequent analyses that arise from the correlation between male and female selection and facilitates the interpretation of results.
We analysed variation in dosage compensation across all Z-linked genes and tissues using linear models of male-to-female expression ratio (dependent variable, as log2(male RPKM/female RPKM)) as a function of concordant selection intensity, male bias in selection intensity, tissue and their interactions. In line with previous analyses of dosage compensation, our approach assumes statistical independence between genes expressed in the same tissue. We also assume independence between expression measures from the same genes measured in different tissues. However, this assumption does not affect our conclusions; all reported tissue-specific effects of the selection measures are also detected when performing analyses separately on each tissue-specific data set.
We tested whether the age of chromosomal strata had an effect on the level of dosage compensation, over and above that of the two measures of selection strength. We ran an analysis modelling log2 male-to-female expression ratio as a function of the two measures of selection strength, tissue, the age of each gene's chromosomal stratum, as well as all the interactions between these terms. Genes that could not be unambiguously assigned to the three major strata were excluded from this analysis. In addition to assessing dosage compensation using the Z-linked log2 male-to-female expression ratio, we also calculated the expression ratio of the single Z chromosome and autosomes in females (Z:AA) and males (ZZ:AA) across each stratum.
Additional information
Accession codes: The raw Illumina reads are deposited at the NCBI Sequence Read Archive under Project number: PRJNA284655.
How to cite this article: Mullon, C. et al. Evolution of dosage compensation under sexual selection differs between X and Z chromosomes. Nat. Commun. 6:7720 doi: 10.1038/ncomms8720 (2015).
Supplementary Material
Supplementary Figures 1-3, Supplementary Tables 1-9, Supplementary Discussion and Supplementary References
Acknowledgments
We thank Beatriz Vicoso, Doris Bachtrog, Laurent Keller, Laurent Lehmann, Charlotte Soneson and Eric Lucas for helpful discussions, and Fabian Zimmer for providing gene expression data. C.M. was supported by the Swiss National Science Foundation (grant to Laurent Keller), A.E.W. and J.E.M. by the European Research Council (grant agreement 260233), and A.E.W. by a National Environment Research Council PhD studentship, M.R. by funding from the Natural Environment Research Council (NE/D009189/1, NE/G019452/1), and A.P. by grants from the Natural Environment Research Council (NE/G00563X/1) and the Engineering and Physical Sciences Research Council (EP/F500351/1, EP/I017909/1, EP/K038656/1).
Footnotes
Author contributions All authors conceived the model. C.M. analysed the model. A.E.W. collected the data. A.E.W., M.R. and J.E.M. analysed the data. All authors wrote the paper.
References
- Bachtrog D. Y-chromosome evolution: emerging insights into processes of Y-chromosome degeneration. Nat. Rev. Genet. 14, 113–124 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pollack J. R. et al. Microarray analysis reveals a major direct role of DNA copy number alteration in the transcriptional program of human breast tumors. Proc. Natl Acad. Sci. USA 99, 12963–12968 (2002). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Torres E. M., Williams B. R. & Amon A. Aneuploidy cells losing their balance. Genetics 179, 737–746 (2008). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ohno S. Sex Chromosomes and Sex Linked Genes Springer-Verlag (1967). [Google Scholar]
- Charlesworth B. Model for the evolution of Y chromosomes and dosage compensation. Proc. Natl Acad. Sci. USA 75, 5618–5622 (1978). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gelbart M. E. & Kuroda M. I. Drosophila dosage compensation: a complex voyage to the X chromosome. Development 136, 1399–1410 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Straub T. & Becker P. B. Dosage compensation: the beginning and end of generalization. Nat. Rev. Genet. 8, 47–57 (2007). [DOI] [PubMed] [Google Scholar]
- Reinke V., Gil I. S., Ward S. & Kazmer K. Genome-wide germline-enriched and sex-biased expression profiles in Caenorhabditis elegans. Development 131, 311–323 (2004). [DOI] [PubMed] [Google Scholar]
- Ercan S. et al. X chromosome repression by localization of the C elegans dosage compensation machinery to sites of transcription initiation. Nat. Genet. 39, 403–408 (2007). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith G., Chen Y.-R., Blissard G. W. & Briscoe A. D. Complete dosage compensation and sex-biased gene expression in the moth Manduca sexta. Genome Biol. Evol. 6, 526–537 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bachtrog D. et al. Sex determination: why so many ways of doing it? PLoS Biol. 12, e1001899 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marín I., Siegal M. L. & Baker B. S. The evolution of dosage-compensation mechanisms. Bioessays 22, 1106–1114 (2000). [DOI] [PubMed] [Google Scholar]
- Griffin R. M., Dean R., Grace J. L., Rydén P. & Friberg U. The shared genome is a pervasive constraint on the evolution of sex-biased gene expression. Mol. Biol. Evol. 30, 2168–2176 (2013). [DOI] [PubMed] [Google Scholar]
- Innocenti P. & Chenoweth S. F. Interspecific divergence of transcription networks along lines of genetic variance in Drosophila: dimensionality, evolvability, and constraint. Mol. Biol. Evol. 30, 1358–1367 (2013). [DOI] [PubMed] [Google Scholar]
- Rice W. R. Sex chromosomes and the evolution of sexual dimorphism. Evolution 38, 735–742 (1984). [DOI] [PubMed] [Google Scholar]
- Mullon C., Pomiankowski A. & Reuter M. The effects of selection and genetic drift on the genomic distribution of sexually antagonistic alleles. Evolution 66, 3743–3753 (2012). [DOI] [PubMed] [Google Scholar]
- Mank J. E. The W, X, Y and Z of sex chromosome dosage compensation. Trends Genet. 25, 226–233 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clutton-Brock T. H. The Evolution of Parental Care Princeton Univ. Press (1991). [Google Scholar]
- Andersson M. Sexual Selection Princeton Univ. Press (1994). [Google Scholar]
- Charlesworth B., Coyne J. A. & Barton N. H. The relative rates of evolution of sex chromosomes and autosomes. Am. Nat. 130, 113–146 (1987). [Google Scholar]
- Mank J. E., Vicoso B., Berlin S. & Charlesworth B. Effective population size and the Faster-X Effect: Empirical results and their interpretation. Evolution 64, 663–674 (2010). [DOI] [PubMed] [Google Scholar]
- Mank J. E., Nam K., Brunström B. & Ellegren H. Ontogenetic complexity of sexual dimorphism and sex-specific selection. Mol. Biol. Evol. 27, 1570–1578 (2010). [DOI] [PubMed] [Google Scholar]
- Brawand D. et al. The evolution of gene expression levels in mammalian organs. Nature 478, 343–348 (2011). [DOI] [PubMed] [Google Scholar]
- Nunney L. The influence of mating system and overlapping generations on effective population size. Evolution 47, 1329–1341 (1993). [DOI] [PubMed] [Google Scholar]
- Arnold S. J. & Wade M. J. On the measurement of natural and sexual selection: applications. Evolution 38, 720–734 (1984). [DOI] [PubMed] [Google Scholar]
- Agrawal A. F. Sexual selection and the maintenance of sexual reproduction. Nature 411, 692–695 (2001). [DOI] [PubMed] [Google Scholar]
- Rowe L. & Houle D. The Lek Paradox and the capture of genetic variance by condition dependent traits. Proc. R Soc. Lond. Ser. B 263, 1415–1421 (1996). [Google Scholar]
- Cotton S., Fowler K. & Pomiankowski A. Do sexual ornaments demonstrate heightened condition-dependent expression as predicted by the handicap hypothesis? Proc. R Soc. Lond. Ser. B 271, 771–783 (2004). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nomura T. Effective size of populations with unequal sex ratio and variation in mating success. J. Anim. Breed. Genet. 119, 297–310 (2002). [Google Scholar]
- Arnold A. P., Itoh Y. & Melamed E. A bird's-eye view of sex chromosome dosage compensation. Ann. Rev. Genomics Hum. Genet. 9, 109–127 (2008). [DOI] [PubMed] [Google Scholar]
- Romero I. G., Ruvinsky I. & Gilad Y. Comparative studies of gene expression and the evolution of gene regulation. Nat. Rev. Genet. 13, 505–516 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moghadam H. K., Pointer M. A., Wright A. E., Berlin S. & Mank J. E. W chromosome expression responds to female-specific selection. Proc. Natl Acad. Sci. USA 109, 8207–8211 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nuzhdin S. V., Wayne M. L., Harmon K. L. & McIntyre L. M. Common pattern of evolution of gene expression level and protein sequence in Drosophila. Mol. Biol. Evol. 21, 1308–1317 (2004). [DOI] [PubMed] [Google Scholar]
- Ometto L., Shoemaker D., Ross K. G. & Keller L. Evolution of gene expression in fire ants: the effects of developmental stage, caste, and species. Mol. Biol. Evol. 28, 1381–1392 (2011). [DOI] [PubMed] [Google Scholar]
- Harrison P. W. et al. Sexual selection drives evolution and rapid turnover of male gene expression. Proc. Natl Acad. Sci. USA 112, 4393–4398 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gout J.-F., Kahn D. & Duret L. Paramecium Post-Genomics Consortium. The relationship among gene expression, the evolution of gene dosage, and the rate of protein evolution. PLoS Genet. 6, e1000944 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jordan I. K., Mariño-Ramírez L. & Koonin E. V. Evolutionary significance of gene expression divergence. Gene 345, 119–126 (2005). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Khaitovich P. et al. Parallel patterns of evolution in the genomes and transcriptomes of humans and chimpanzees. Science 309, 1850–1854 (2005). [DOI] [PubMed] [Google Scholar]
- Blekhman R., Oshlack A., Chabot A. E., Smyth G. K. & Gilad Y. Gene regulation in primates evolves under tissue-specific selection pressures. PLoS Genet. 4, e1000271 (2008). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lemos B., Bettencourt B. R., Meiklejohn C. D. & Hartl D. L. Evolution of proteins and gene expression levels are coupled in Drosophila and are independently associated with mRNA abundance, protein length, and number of protein-protein interactions. Mol. Biol. Evol. 22, 1345–1354 (2005). [DOI] [PubMed] [Google Scholar]
- Hunt B. G., Ometto L., Keller L. & Goodisman M. A. D. Evolution at two levels in fire ants: the relationship between patterns of gene expression and protein sequence evolution. Mol. Biol. Evol. 30, 263–271 (2012). [DOI] [PubMed] [Google Scholar]
- Drummond D. A., Bloom J. D., Adami C., Wilke C. O. & Arnold F. H. Why highly expressed proteins evolve slowly. Proc. Natl Acad. Sci. USA 102, 14338–14343 (2005). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wright A. E., Moghadam H. K. & Mank J. E. Trade-off between selection for dosage compensation and masculinization on the avian Z chromosome. Genetics 192, 1433–1445 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pomiankowski A. & Moller A. P. A resolution of the Lek Paradox. Proc. R Soc. Lond. Ser. B 260, 21–29 (1995). [Google Scholar]
- Sharp N. P. & Agrawal A. F. Male-biased fitness effects of spontaneous mutations in Drosophila melanogaster. Evolution 67, 1189–1195 (2013). [DOI] [PubMed] [Google Scholar]
- Connallon T. & Clark A. G. Association between sex-biased gene expression and mutations with sex-specific phenotypic consequences in Drosophila. Genome Biol. Evol. 3, 151–155 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Prince E. G., Kirkland D. & Demuth J. P. Hyperexpression of the X chromosome in both sexes results in extensive female bias of X-linked genes in the Flour Beetle. Genome Biol. Evol. 2, 336–346 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pessia E., Makino T., Bailly-Bechet M., McLysaght A. & Marais G. A. B. Mammalian X chromosome inactivation evolved as a dosage-compensation mechanism for dosage-sensitive genes on the X chromosome. Proc. Natl Acad. Sci. USA 109, 5346–5351 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chuang P.-T., Lieb J. D. & Meyer B. J. Sex-specific assembly of a dosage compensation complex on the nematode X chromosome. Science 274, 1736–1739 (1996). [DOI] [PubMed] [Google Scholar]
- Gladstein N., McKeon M. N. & Horabin J. I. Requirement of male-specific dosage compensation in Drosophila females—implications of early X chromosome gene expression. PLoS Genet. 6, e1001041 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vicoso B. & Charlesworth B. Effective population size and the faster-X effect: an extended model. Evolution 63, 2413–2426 (2009). [DOI] [PubMed] [Google Scholar]
- Champagnat N. & Lambert A. Evolution of discrete populations and the canonical diffusion of adaptive dynamics. Ann. Appl. Prob. 17, 102–155 (2007). [Google Scholar]
- Lehmann L. The stationary distribution of a continuously varying strategy in a class-structured population under mutation–selection–drift balance. J. Evol. Biol. 25, 770–787 (2012). [DOI] [PubMed] [Google Scholar]
- Hughes K. A. et al. Segregating variation in the transcriptome: cis regulation and additivity of effects. Genetics 173, 1347–1355 (2006). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kelley R. L. et al. Epigenetic spreading of the Drosophila dosage compensation complex from roX RNA genes into flanking chromatin. Cell 98, 513–522 (1999). [DOI] [PubMed] [Google Scholar]
- Bachtrog D. Positive selection at the binding sites of the male-specific lethal complex involved in dosage compensation in Drosophila. Genetics 180, 1123–1129 (2008). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ewens W. J. Mathematical Population Genetics: I. Theoretical Introduction Springer (2004). [Google Scholar]
- Gardiner C. Stochastic Methods: A Handbook for the Natural and Social Sciences Springer (2009). [Google Scholar]
- Andrew S. Babraham Bioinformatics http://www.bioinformatics.babraham.ac.uk/projects/fastqc/.
- Lohse M. et al. RobiNA: a user-friendly, integrated software solution for RNA-Seq-based transcriptomics. Nucleic Acids Res. 40, W622–W627 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Trapnell C., Pachter L. & Salzberg S. L. TopHat: discovering splice junctions with RNA-Seq. Bioinformatics 25, 1105–1111 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim D. et al. TopHat2: accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol. 14, 1–13 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Langmead B., Trapnell C., Pop M. & Salzberg S. Ultrafast and memory-efficient alignment of short DNA sequences to the human genome. Genome Biol. 10, 1–10 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li H. et al. The sequence alignment/map format and SAMtools. Bioinformatics 25, 2078–2079 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Robinson M. D., McCarthy D. J. & Smyth G. K. edgeR: a Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 26, 139–140 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Altschul S. F., Gish W., Miller W., Myers E. W. & Lipman D. J. Basic local alignment search tool. J. Mol. Biol. 215, 403–410 (1990). [DOI] [PubMed] [Google Scholar]
- Löytynoja A. & Goldman N. An algorithm for progressive multiple alignment of sequences with insertions. Proc. Natl Acad. Sci. USA 102, 10557–10562 (2005). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang Z. PAML 4: phylogenetic analysis by maximum likelihood. Mol. Biol. Evol. 24, 1586–1591 (2007). [DOI] [PubMed] [Google Scholar]
- Axelsson E. et al. Natural selection in avian protein-coding genes expressed in brain. Mol. Ecol. 17, 3008–3017 (2008). [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplementary Figures 1-3, Supplementary Tables 1-9, Supplementary Discussion and Supplementary References