

Abstract
Objectives: This study aims to explore whether Huperzine A (HupA) could protect neural stem cells against amyloid beta-peptide Aβ induced apoptosis in a neural stem cells (NSCs) and microglia co-culture system. Methods: Rat NSCs and microglial cells were isolated, cultured and identified with immunofluorescence Assays (IFA). Co-culture systems of NSCs and microglial cells were employed using Transwell Permeable Supports. The effects of Aβ1-42 on NSCs were studied in 4 groups using co-culture systems: NSCs, Aβ+NSCs, co-culture and Aβ+co-culture groups. Bromodeoxyuridine (BrdU) incorporation and flow cytometry were utilized to assess the differences of proliferation, differentiation and apoptosis of NSCs between the groups. LQ test was performed to assess the amounts of IL-6, TNF-α and MIP-α secreted, and flow cytometry and Western blotting were used to assess apoptosis of NSCs and the expressions of Bcl-2 and Bax in each group. Results: IFA results showed that isolated rat NSCs were nestin-positive and microglial cells were CD11b/c-positive. Among all the groups, the Aβ+co-culture group has the lowest BrdU expression level, the lowest MAP2-positive, ChAT-positive cell counts and the highest NSC apoptosis rate. Smaller amounts of IL-6, TNF-α and MIP-α were being secreted by microglial cells in the HupA+Aβ+co-culture group compared with those in the Aβ+ co-culture group. Also the Bcl-2: Bax ratio was much higher in the HupA+Aβ+co-culture group than in the Aβ+co-culture group. Conclusions: HupA inhibits cell apoptosis through restraining microglia’s inflammatory response induced by Aβ1-42.
Keywords: Huperzine A, Alzheimer’s disease, amyloid beta-peptide, neural stem cells, microglial cells
Introduction
Alzheimer’s disease (AD) is a progressive, neurodegenerative disease, the most common form of progressive intellectual failure in humans [1]. To date, its mechanism is not clear yet, but it is considered to be associated with loss of neurons in the brain resulted from inflammatory response caused by Amyloid beta-peptide (Aβ) deposits activating microglial cells [2]. However, it has not been confirmed yet whether inflammation caused by Aβ-induced microglia activation makes the environment in the brain not suitable to live and differentiate in for neural cells, leading to insufficient supplementation of neural cells.
Huperzine A (HupA), extracted from Chinese club moss Huperzia serrata (Qian Ceng Ta), is a potent, reversible and selective inhibitor of acetylcholinesterase (AChE) with a rapid absorption and penetration into the brain in animal tests [3,4]. HupA has been used for AD treatment widely in China and in clinical trials for the treatment of age-related memory deficiencies in the United States (http://www.clinicaltrials.gov/show/NCT00083590). At the moment, the mechanism of treating AD with HupA is not fully understood yet. Studies have shown that preincubation with HupA could protect rat neurons against Aβ-mediated apoptosis [5,6]. Most existing studies on treating AD with HupA target mature functional cells, only a few investigate the effects of HupA on neural stem cells (NSCs).
NSCs are neural precursor cells with capacity for self-renewal and multipotency. Neurons, astrocytes and oligodendrocytes are among cells that NSCs can differentiate into [7]. Along with loss of neurons caused by Aβ-induced microglia activation, studies have shown that Aβ can suppress neural progenitor cell proliferation and differentiation, and induce apoptosis [8]. Based on this, we speculate that Aβ-induced microglia activation may indirectly harm NSCs’ chances of survival, while HupA protects them against such an effect.
To test our hypothesis, we established co-culture systems of NSCs and microglia with Transwell Permeable Supports, added Aβ1-42 into microglial cell layers to simulate the inflammatory microenvironment in AD patients’ brains. Effects of Aβ treated microglia on the survival of NSCs were studied in vitro to determine whether Aβ inhibits the production of neural cells through microglial cells. Also, effects of HupA on inflammatory response were evaluated in NSC-microglia co-culture systems. Meanwhile, the apoptosis rates of NSCs were assessed to determine the protective effects of HupA on NSCs.
Materials and methods
Immunofluorescence assays (IFA)
The cells were fixed with 4% paraformaldehyde for 15 min and then blocked with goat serum for 30 min. Nestin (Rat-401) mouse mAb (Cell Signaling Technology, USA) was added to indentify NSCs and anti-CD11b/c antibodies (Abcam, USA) to identify microglial cells. The cells were rinsed with PBS after placing at 4°C overnight. FITC-labeled (green) and TRITC-labeled (red) goat anti-mouse IgG (Cell Signaling Technology, USA) secondary antibodies were then added for microglial cells and NSCs, respectively. Together, they were incubated for 1 h in the dark. DAPI was used to stain the nuclei for 5 min and images were taken with a fluorescent microscope (Olympus, Japan).
Isolating and culturing of rat NSCs
Cells derived from hippocampus of newborn SD rats were resuspended with DMEM/F12 (Gibco) mixed with EGF (20 μg/L), BFGF (20 μg/L), B27 (2%), L-Gln (2%) and penicillin/streptomycin (1%). They were then seeded in 25 cm2 vented culture flasks (1×109/L per flask) placed at 37°C in a 5% CO2 incubator. Half of the solution was changed every 3-4 days. Subculturing started on Day 7 and repeated once every 6 to 7 days. NSCs obtained from secondary cultures underwent IFA. Second-generation neurospheres were obtained to prepare single cell suspension. Bromodeoxyuridine (BrdU, 10 μmol/L) was added and labeling was allowed for 24 h before the solution was replaced with a fresh batch. The neurospheres were transferred after 4 days to a 24 well culture plate coated with polylysine. IFA tests were performed to assess cell proliferation 2 days later. Second-generation NSCs were treated with DMEM/12F containing 10% FBS. After 10 days, immunofluorescence staining for GFAP and MAP2 was used to evaluate cell multipotency.
Purifying and culturing of rat microglial cells
Hippocampus of newborn SD rats was centrifuged after digested with 0.125% trypsin. The supernatant was discarded and the pellet was resuspended in DMEM/F12 containing 10% FBS. The single cell suspension was then transferred at a cell density of 5×108 to 75 cm2 vented culture flasks coated with polylysine and incubated at 37°C in a 5% CO2 incubator. After 2 days, the medium was replaced with a fresh batch. The flask was shaken by hand after 7 to 9 days to complete the separation of purified microglial cells. IFA tests were then performed on the microglial cells with CD11b/c, specific markers for microglia. Meanwhile, Hoechst 33258 was used for nuclear staining so that microglial cell numbers can be measured and purity can be assessed.
Co-culturing rat NSCs and microglial cells
Co-culture systems where cells shared medium without making any contact were employed with 6-well Transwell Permeable Supports. NSCs were seeded on Transwell inserts and purified microglial cells were cultured in 6-well culture plates at the lower compartment. To evaluate effects of microglial cells on the survival of NSCs before and after microglial cells were activated by Aβ1-42, for NSCs group, NSCs were cultured on a Transwell insert and medium was added on a 6-well plate underneath; for Aβ+NSCs group, NSCs were seeded on a Transwell insert and medium containing 10 μmol/L Aβ1-42 (US Biological) was added on the 6 well plate; for both co-culture and Aβ+co-culture groups, NSCs and microglial cells were seeded at a ratio of 1:1 on Transwell inserts and in the wells of culture plates, respectively. For Aβ+co-culture group, 10 μmol/L Aβ1-42 was also added to the lower compartment. All four groups were incubated at 37°C in a 5% CO2 incubator for 96 h before undergoing tests.
To examine the protective effects of HupA on NSCs grown in co-culture systems, NSCs and microglial cells were added at a ratio of 1:1 onto Transwell inserts and culture plates underneath, respectively for co-culture, Aβ+co-culture and HupA+Aβ+co-culture groups. Aβ1-42 (10 μmol/L) was added to the lower compartments of Aβ+co-culture and HupA+Aβ+co-culture groups. For HupA+Aβ+co-culture group, the microglial cells were treated with 1μmol/L HupA 4 hours ahead of seeding. Concentrations of IL-6, TNF-α and MIP-1α were measured 24, 48 and 72 h after co-culturing started, and flow cytometry and Western blotting were performed to assess NSC apoptosis 72 h after co-culturing started.
Assessment of NSC proliferation with BrdU incorporation assays
Neurospheres were obtained to prepare single cell suspension. BrdU (10 μmol/L) was added to label cells. Cells were collected after 24 h and resuspended. Cell suspension was then used in each group in co-culture systems. Neurospheres from all the groups were transferred after 96 hours’ co-culturing to a 24-well culture plate coated with polylysine and cultured for 2 h. IFA was then performed as follows to examine expressions of BrdU in each group. The cells were fixed with 4% paraformaldehyde at room temperature and then treated with 0.3%Triton X-100. They were blocked with 10% goat serum before incubated with mouse anti-BrdU monoclonal antibodies (Santa Cruz Biotech, Dallas, TX, USA) overnight at 4°C in a humidified chamber and then for another 1.5 h in the dark with FITC-conjugated goat anti-mouse IgG (Santa Cruz Biotech, Dallas, TX, USA). After nuclear staining with Hoechst 33258, the cells were observed and images were taken with a fluorescence microscope.
Assessment of cell apoptosis with flow cytometry
NSCs were collected from each group. A flow cytometer was used and instructions of an AnnexinV-FITC/PI detection kit (sigma) were followed to assess cell apoptosis.
NSC potency assessment
Immunofluorescent staining was performed 96 h after co-culturing started. The following antibodies were used: mouse anti-MAP2 (Abcam, USA) and rabbit anti-ChAT polyclonal antibody (Abcam, USA) as primary antibodies, FITC-labeled anti-mouse IgG and anti-rabbit IgG (Abcam, USA) as secondary antibodies. NSC potency was analyzed with a flow cytometer (FACS) and a CellQuest 3.0 analysis software.
LQ test
Supernatant was collected from each group. Instructions of LINCOplex Kit (Millipore, USA) were followed as follows to assess the concentrations of IL-6, TNF-α and MIP-α. Detection reagents were brought to room temperature. Buffer was pipette onto an assay plate to coat the filter plate. The plate was then shaken for 10 min at room temperature. 25 μL supernatant, diluted standard sample, blank sample and quality control sample were added to wells on the plate, separately. Microplate mixtures were added too. Samples were incubated for 2 h at room temperature on a microplate shaker. The plate was rinsed twice with wash buffer twice before detection reagents were added to each well to detect antibodies. The cells were then incubated for 2 h at room temperature on a microplate shaker. Afterwards, streptavidin was added and the cells were incubated for 30 min. The plate was rinsed again before detection reagents were added. Luminex LiquiChip analytical system was utilized to detect the concentrations of inflammatory factors.
Western blot
The nuclear proteins of NSCs were extracted with a nuclear protein extraction kit (KeyGen bioTECH, China). The bicinchoninic acid (BCA) assay was used for protein quantization. A sample containing 15 μg proteins were separated by 10% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE). Semi-dry transfer was performed and proteins were transferred to PVDF membranes (Bio-Rad Laboratories, USA). Proteins were blocked with 5% milk made out of skim milk powder and 1 h later incubated with Bcl-2 (1:100), Bax (1:100) and β-actin (Cell Signaling, USA) primary antibodies and secondary antibodies.
Statistical analysis
SPSS 16.0 was utilized for statistical analysis. Data are presented as mean ± standard deviation (SD). Statistical significance among groups was determined by one way ANOVA. LSD tests were performed to determine differences between groups. Differences were considered to be statistically significant where P<0.05.
Results
Identification of NSCs and microglial cells
To identify isolated rat NSCs and microglial cells, IFA assays were performed. Nestin, a marker that expresses in neuroepithelial stem cells, is widely employed for identification of NSCs [9]. Neurospheres were tested positive for nestin (Figure 1A). Microglial cells were identified by immunofluorescence staining with CD11b/c. Microglial cells can be seen-flat or oval-shaped with slender elongated processes projecting from cell bodies. IFA results showed more than 95% of microglial cells were CD11b/c- positive (Figure 1B).
Figure 1.
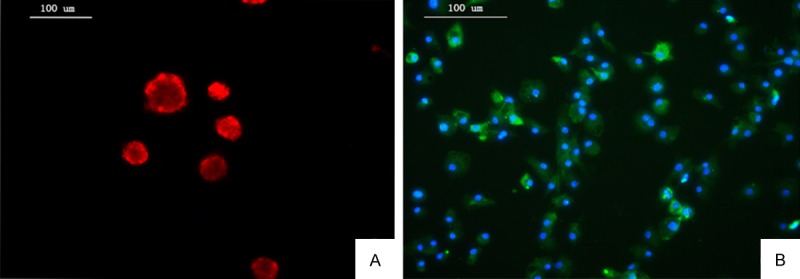
Identification of NSCs and microglial cells. A: NSCs were nestin-positive (red). B: Microglial cells were CD11b/c-positive (green). Nuclei were stained with DIPA (blue).
Aβ1-42 inhibits proliferation and differentiation and promote apoptosis of NSCs in co-culture systems
To study effects of Aβ1-42 on proliferation of NSCs in co-culture systems, BrdU incorporation assays were used to assess NSC proliferations in each group. Results showed that Aβ1-42 treatment significantly inhibited NSC proliferation. No significant difference was observed between the co-culture group and the NSCs group (P>0.05), while differences between the Aβ+co-culture group, the Aβ+NSCs group and the co-culture group were all statistically significant (P<0.01). This suggests microglial cells that were not treated with Aβ1-42 have no significant effect on NSC proliferation and that Aβ1-42 could not only exert direct effects on NSC proliferation but also magnify the inhibitory effects of microglial cells on NSC proliferation (Figure 2A-E).
Figure 2.
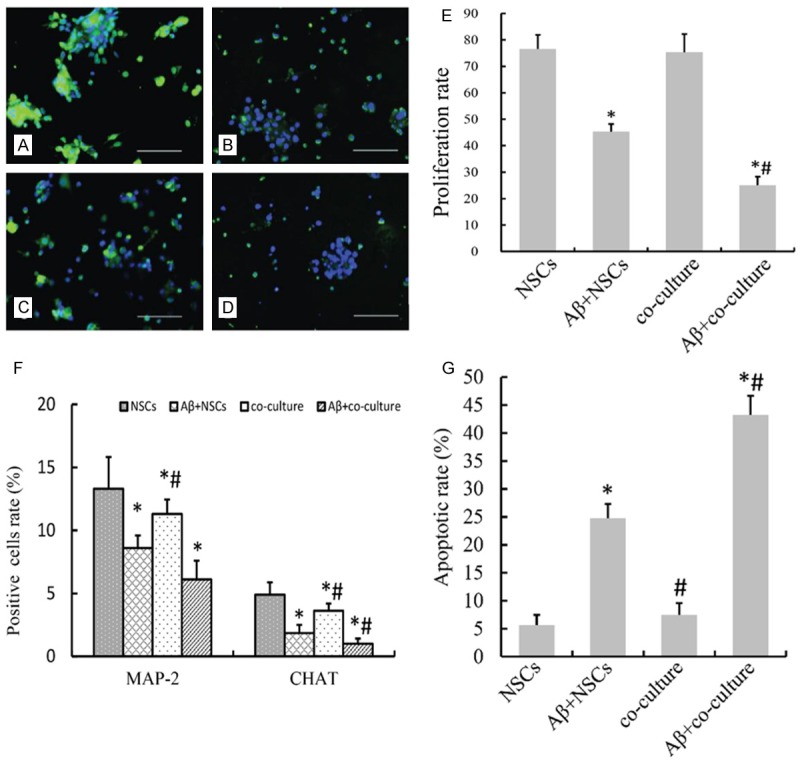
Aβ1-42 inhibits proliferation and differentiation and promotes apoptosis of NSCs in co-culture systems. (A-E) BrdU incorporation assays were utilized to examine effects of Aβ1-42 on NSC proliferation (scale bar =100 μm). (A) NSCs; (B) NSCs treated with Aβ1-42; (C) NSCs and microglial cells; (D) NSCs and microglial cells treated with Aβ1-42. (E) NSC proliferation rate of (A-D) group showed in BrdU incorporation assays. *P<0.05 vs. NSCs, #P<0.05 vs. Aβ+NSCs. F: Flow cytometry was performed to detect MAP2 and ChAT positive cells. *P<0.05 vs. NSCs, #P<0.05 vs. Aβ+NSCs. The experiment was repeated 5 times. (G) NSC apoptosis rate. *P<0.05 vs. NSCs, #P<0.05 vs. Aβ+NSCs. The experiment was repeated 5 times and the mean of the results was adopted.
To investigate effects of Aβ1-42 on differentiation of neurons in co-culture systems, IFA was performed to detect MAP2-positive and ChAT-positive cells. Results showed there were more MAP2-positive and ChAT-positive cells in the NSCs group than in the co-culture group (P<0.01, Figure 2F). Also, there were less MAP2-positive and ChAT-positive cells in the Aβ+NSCs group compared with the co-culture group, and the differences are statistically significant (ALL P<0.05).
Results from flow cytometry showed that there was no significant differences between the NSC apoptosis rates in the co-culture group and the NSCs group (P>0.05) while NSC apoptosis rate in the Aβ+co-culture group was higher than those in the co-culture group and the Aβ+NSCs group (P<0.05). The results indicate that Aβ1-42 promotes differentiation and apoptosis while inhibits proliferation of NSCs (Figure 2G).
HupA inhibits Aβ1-42-induced secretion of IL-6, TNF-α and MIP-1α by microlia
Secretions of IL-6, TNF-α and MIP-1α were compared between the co-culture, the Aβ+co-culture and the HupA+Aβ+co-culture groups using LQ test. Results showed that the concentrations of IL-6, TNF-α and MIP-1α were much higher 48 h after co-culturing started than 24 h after treatments. The concentrations of IL-6, TNF-α and MIP-1α in the Aβ group were 2.6, 4.2 and 3.4 times as high as those in NSCs group, respectively, 48 h after co-culturing started (P<0.05), while the concentrations of the three cytokines in the HupA group were 2.1, 1.8 and 2.3 times as high as the concentrations of those in the NSCs group, respectively, and much lower than those in Aβ group (P<0.05). The differences were bigger 72 h after co-culturing started. The concentrations of the cytokines in Aβ were 2.7, 5.8 and 4.8 times as high as the concentrations of those in NSCs group, respectively (P<0.05), while the concentrations of the cytokines in HupA were much lower than those in Aβ group (P<0.05, Figure 3). The results suggest that HupA restrains inflammatory factors from being secreted.
Figure 3.
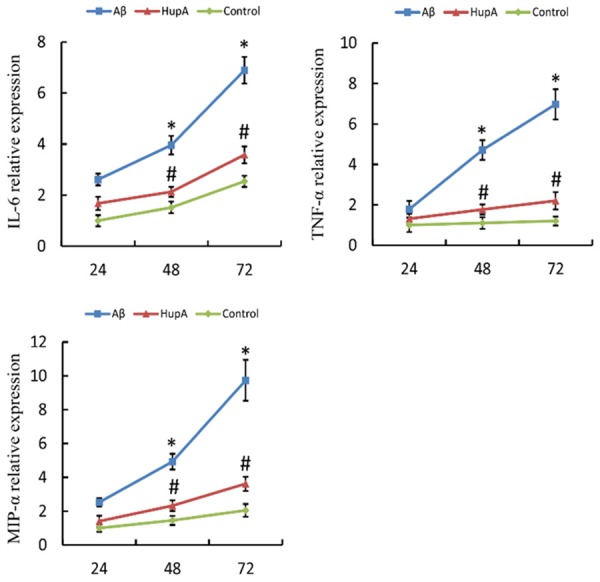
Concentrations of IL-6, TNF-α and MIP-1α in each group. *P<0.05 vs. NSCs, #P<0.05 vs. Aβ+NSCs. The experiment was repeated 5 times and the mean of the results was adopted. Aβ: Aβ+co-culture group; HupA: HupA+Aβ+co-culture group; Control: co-culture group.
HupA reduces Aβ1-42-induced NSC apoptosis
IFA and Western blot were performed to study the effects of HupA on NSC apoptosis in NSC-microglia co-culture systems with the presence of Aβ1-42. Flow cytometry results showed that 72 h after co-culturing started, the apoptosis rate of NSCs in co-culture group was 4.3%, that in Aβ group was 24.6% and that in HupA group was 9.8%. The differences between the co-culture group and the Aβ+co-culture group, and that between the Aβ group and the HupA+Aβ+co-culture group are statistically significant (P<0.05, Figure 4).
Figure 4.
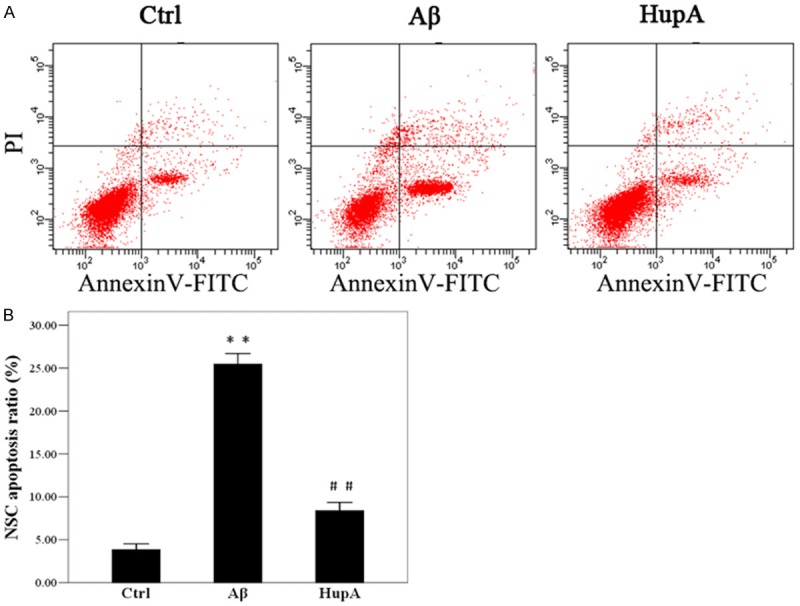
Flow cytometry to assess NSC apoptosis. Aβ: Aβ+co-culture group; HupA: HupA+Aβ+co-culture group; Control: co-culture group. (B) NSC apoptosis ratio showed in (A). *P<0.05 vs. Control; #P<0.05 vs. Aβ. The experiment was repeated 5 times and the mean of the results was adopted.
A study by Wong R, Zhang HY and Tang XC showed that HupA inhibits apoptosis induced by Aβ through raising the expression of anti-apoptotic protein Bcl-2 and decreasing the expression of pro-apoptotic protein Bax [6]. Expressions of Bcl-2 and Bax were analyzed by Western blot. Results showed that expressions of Bcl-2 and Bax proteins were the highest in the Aβ+co-culture group 72 h after treatment, with the lowest Bcl-2: Bax ratio (P<0.05). The Bcl-2: Bax ratio is much higher in the HupA+Aβ+co-culture group compared with the Aβ+co-culture group. The difference is statistically significant (P<0.05, Figure 5).
Figure 5.
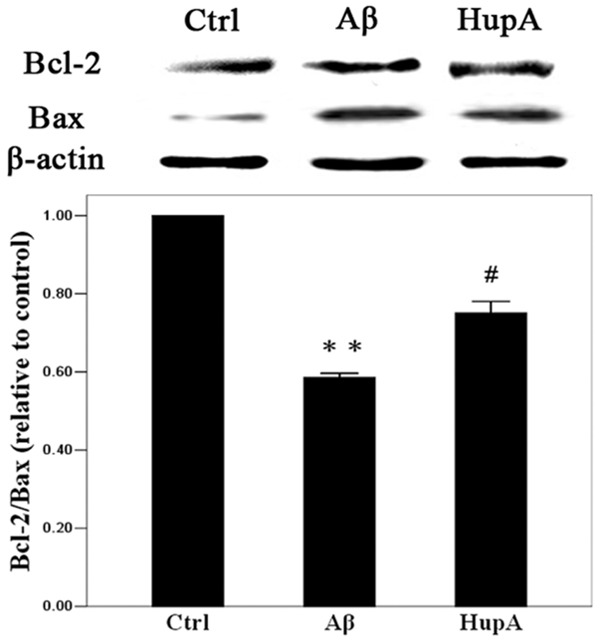
Western blot assay of Bcl-2 and Bax expressions. Western blotting was repeated 3 times and the mean of the results was adopted. *P<0.05 vs. Control; #P<0.05 vs. Aβ.
Discussion
In vivo and in vitro studies have shown that Aβ can activate microglial cells [10], and that activated microglial cells can release several types of cytokines, chemokines, complement and activation products of complement, leading to nonspecific inflammatory cell infiltration and causing chronic inflammation [11]. It still has not been confirmed whether this makes the environment in the brain not suitable for neural cells to live and differentiate in, resulting in endogenous and transplanted exogenous NSCs not being able to function properly, and eventually leading to insufficient supplementation of neural cells in AD patients’ brain. This study established co-culture systems of NSCs and microglial cells. Proliferation and differentiation of NSCs were evaluated before and after microglial cells were treated with Aβ1-42. Results showed that microglial cells not treated with Aβ1-42 has no significant effects on proliferation of NSCs in co-culture systems, just as reported by past researches [12]. That the percentage of NSCs differentiating into neurons and cholinergic neurons was lower than normal level may have to do with some factors secreted by microglial cells under normal circumstances. Studies have shown that Aβ1-42 can inhibit proliferation of NSCs, promote their apoptosis and lower the percentage of NSCs differentiating into neurons and cholinergic neurons [13-15]. NSCs’ capacity of self-renewal and multipotency was markedly restrained by Aβ1-42 in NSC-microglia co-culture systems, indicating inhibitive effects of Aβ1-42 on NSC survival may be magnified through microglial cells. These factors may be to blame for the hostile microenvironment inside the brains of AD patients that threatens the survival of endogenous and exogenous NSCs. This poses a challenge on tackling AD using NSCs but also sheds some light on it.
In this study, as significantly smaller amounts of inflammatory factors such as IL-6, TNF-α and MIP-1α were secreted with the presence of HupA, we presume this may be the way in which HupA exerts its protective effects on NSCs. Aβ that accumulates in the brain tissue of AD patients acts on microglia constantly, resulting in chronic inflammation. Chronic inflammation then leads to neuron apoptosis and inhibits neural cells from being produced, causing massive loss of neural cells in the central nervous system. This could be the cause of AD and the main reason that it deteriorates. HupA, as a cholinesterase inhibitor, has been applied in AD treatment and is effective to certain extent. A study by Wang J et al. suggested HupA makes the inflammatory response less severe through acting on cholinergic receptors [16]. This study utilized Transwell Permeable Supports and established co-culture systems of NSCs and microglial cells. The inflammatory microenvironment in AD patients’ brains was simulate by exposing microglial cells to Aβ to examine the protective effects of HupA and NSCs. Results showed that, with the stimulus of Aβ1-42, microglial cells secreted IL-6, TNF-α and MIP-α. These inflammatory factors passed through Transwell semipermeable membranes, triggering apoptosis of NSCs. IL-6, a cytokine that expresses largely in AD patients, can promote inflammatory response through inducing the expression of phospholipase A2 gene. TNF-α is an important pro-inflammatory cytokine that triggers and regulates cytokine storm in the inflammatory response. It can stimulate the expressions of cytokines such as IL-1, and these cytokines, in turn, continuously magnify the inflammatory response in various ways [17]. Therefore, inhibiting TNF-α and IL-6 from being produced is crucial for the control of chronic inflammation and the improvement of microenvironment for AD patients. Unlike TNF-α and IL-6, MIP-1α is a member of the C-C subfamily of chemokines. MIP-1α chemotaxis can cause microglial cells to accumulate around Aβ plaques in AD patients. Microglial cells, stimulated by Aβ, release inflammatory factors including MIP-ld, forming a vicious circle (ieroglia activation in retinal degeneration). With the presence of HupA, significantly smaller amounts of IL-6, TNF-α and MIP-1α were secreted by microglial cells, exerting strong anti-inflammatory effects. Studies conducted in the past by researchers showed this is realized as HupA restrains cholinesterase activities, increases acetylcholine, activates cholinergic receptors and eventually reduces the expressions of cytokines and chemokines [16,18,19]. Our study showed HupA can act on microglial cells directly, retraining inflammatory factors from being secreted by microglial cells, thus inhibit inflammatory response.
HupA is effective in AD treatment. A study by Zhang HY and Tang XC showed that HupA protect neurons through reducing Aβ production and acting on the signaling pathways of nerve growth factor [20]. This study focuses on the protective effects of HupA on NSCs. Results from flow cytometry and Western blotting showed that the Bcl-2: Bax ratio in NSCs in the HupA+Aβ+co-culture group was higher than that in the Aβ+co-culture group. Bcl-2 is an important anti-apoptotic protein while Bax is a pro-apoptotic one in Bcl-2 protein family and can antagonize the protective effects of Bcl-2. Whether cells live or die after stimulated by apoptosis signals depends on the ratio of Bcl-2 protein family’s pro-apoptotic and anti-apoptotic proteins. When Bcl-2: Bax ratio is low, Bax forms homodimers to induce cell apoptosis; when Bcl-2: Bax ratio is elevated, Bax and Bcl-2 form heterodimers, inhibiting cell apoptosis [21].
We found that stimulated by Aβ1-42-mediated inflammatory response, the expressions of Bcl-2 and Bax were the highest in the Aβ+co-culture group, with the lowest Bcl-2: Bax ratio. LQ test revealed that treated with HupA, the amount of inflammatory factors being secreted was smaller in the HupA+Aβ+co-culture group than in the Aβ+co-culture group, with a much higher Bcl-2: Bax ratio, exerting inhibitive effects on NSC apoptosis. Results from Western blotting and flow cytometry both indicate HupA can inhibit NSC apoptosis. HupA reduces the amount of inflammatory factors being secreted, decrease NSC apoptosis rates, favoring neural cell survival.
To conclude, this study proved Aβ1-42 inhibits NSC proliferation, apoptosis and differentiation, suggesting Aβ1-42 magnifies its inhibitive effects on neural cell survival through microglial cells. Also, smaller amount of IL-6, TNF-α and MIP-1α was secreted by microglial cells and the apoptosis level of NSCs was lower with the presence of HupA, indicating one mechanism for HupA to treat AD may be through restraining microglial cells from secreting cytokines and chemokines, and thus making inflammatory response less severe and the microenvironment more friendly for neural cells to live in and eventually raise their survival rate. However, the specific signaling pathways through which HupA acts on microglial cells, the mechanism and effects of HupA on proliferation and differentiation of NSCs await further studies.
Acknowledgements
This work was supported in part by grants from the National Natural Science Foundation of China (No. 30973162) and the National Natural Science Foundation of Guangdong Province (No. S2013010015546).
Disclosure of conflict of interest
None.
References
- 1.Selkoe DJ. Developing preventive therapies for chronic diseases: lessons learned from Alzheimer’s disease. Nutr Rev. 2007;65:S239–S243. doi: 10.1111/j.1753-4887.2007.tb00370.x. [DOI] [PubMed] [Google Scholar]
- 2.Farias GG, Godoy JA, Vazquez MC, Adani R, Meshulam H, Avila J, Amitai G, Inestrosa NC. The anti-inflammatory and cholinesterase inhibitor bifunctional compound IBU-PO protects from beta-amyloid neurotoxicity by acting on Wnt signaling components. Neurobiol Dis. 2005;18:176–183. doi: 10.1016/j.nbd.2004.09.012. [DOI] [PubMed] [Google Scholar]
- 3.Wang R, Yan H, Tang XC. Progress in studies of huperzine A, a natural cholinesterase inhibitor from Chinese herbal medicine. Acta Pharmacol Sin. 2006;27:1–26. doi: 10.1111/j.1745-7254.2006.00255.x. [DOI] [PubMed] [Google Scholar]
- 4.Bai DL, Tang XC, He XC. Huperzine A, a potential therapeutic agent for treatment of Alzheimer’s disease. Curr Med Chem. 2000;7:355–374. doi: 10.2174/0929867003375281. [DOI] [PubMed] [Google Scholar]
- 5.Xiao XQ, Zhang HY, Tang XC. Huperzine A attenuates amyloid beta-peptide fragment 25-35-induced apoptosis in rat cortical neurons via inhibiting reactive oxygen species formation and caspase-3 activation. J Neurosci Res. 2002;67:30–36. doi: 10.1002/jnr.10075. [DOI] [PubMed] [Google Scholar]
- 6.Wang R, Zhang HY, Tang XC. Huperzine A attenuates cognitive dysfunction and neuronal degeneration caused by beta-amyloid protein-(1-40) in rat. Eur J Pharmacol. 2001;421:149–156. doi: 10.1016/s0014-2999(01)01030-5. [DOI] [PubMed] [Google Scholar]
- 7.Price J, Williams BP. Neural stem cells. Curr Opin Neurobiol. 2001;11:564–567. doi: 10.1016/s0959-4388(00)00250-6. [DOI] [PubMed] [Google Scholar]
- 8.Haughey NJ, Liu D, Nath A, Borchard AC, Mattson MP. Disruption of neurogenesis in the subventricular zone of adult mice, and in human cortical neuronal precursor cells in culture, by amyloid beta-peptide: implications for the pathogenesis of Alzheimer’s disease. Neuromolecular Med. 2002;1:125–135. doi: 10.1385/NMM:1:2:125. [DOI] [PubMed] [Google Scholar]
- 9.Gage FH, Ray J, Fisher LJ. Isolation, characterization, and use of stem cells from the CNS. Annu Rev Neurosci. 1995;18:159–192. doi: 10.1146/annurev.ne.18.030195.001111. [DOI] [PubMed] [Google Scholar]
- 10.Rothwell NJ, Luheshi GN. Interleukin 1 in the brain: biology, pathology and therapeutic target. Trends Neurosci. 2000;23:618–625. doi: 10.1016/s0166-2236(00)01661-1. [DOI] [PubMed] [Google Scholar]
- 11.Calafiore M, Battaglia G, Zappala A, Trovato-Salinaro E, Caraci F, Caruso M, Vancheri C, Sortino MA, Nicoletti F, Copani A. Progenitor cells from the adult mouse brain acquire a neuronal phenotype in response to beta-amyloid. Neurobiol Aging. 2006;27:606–613. doi: 10.1016/j.neurobiolaging.2005.03.019. [DOI] [PubMed] [Google Scholar]
- 12.Villeda S, Wyss-Coray T. Microglia--a wrench in the running wheel? Neuron. 2008;59:527–529. doi: 10.1016/j.neuron.2008.08.005. [DOI] [PubMed] [Google Scholar]
- 13.Fonseca MB, Sola S, Xavier JM, Dionisio PA, Rodrigues CM. Amyloid beta peptides promote autophagy-dependent differentiation of mouse neural stem cells: Abeta-mediated neural differentiation. Mol Neurobiol. 2013;48:829–840. doi: 10.1007/s12035-013-8471-1. [DOI] [PubMed] [Google Scholar]
- 14.Itokazu Y, Yu RK. Amyloid beta-peptide 1-42 modulates the proliferation of mouse neural stem cells: upregulation of fucosyltransferase IX and notch signaling. Mol Neurobiol. 2014;50:186–196. doi: 10.1007/s12035-014-8634-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Itokazu Y, Kato-Negishi M, Nakatani Y, Ariga T, Yu RK. Effects of amyloid beta-peptides and gangliosides on mouse neural stem cells. Neurochem Res. 2013;38:2019–2027. doi: 10.1007/s11064-013-1108-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Wang J, Zhang HY, Tang XC. Huperzine a improves chronic inflammation and cognitive decline in rats with cerebral hypoperfusion. J Neurosci Res. 2010;88:807–815. doi: 10.1002/jnr.22237. [DOI] [PubMed] [Google Scholar]
- 17.Perry RT, Collins JS, Wiener H, Acton R, Go RC. The role of TNF and its receptors in Alzheimer’s disease. Neurobiol Aging. 2001;22:873–883. doi: 10.1016/s0197-4580(01)00291-3. [DOI] [PubMed] [Google Scholar]
- 18.Wang J, Chen F, Zheng P, Deng W, Yuan J, Peng B, Wang R, Liu W, Zhao H, Wang Y, Wu G. Huperzine A ameliorates experimental autoimmune encephalomyelitis via the suppression of T cell-mediated neuronal inflammation in mice. Exp Neurol. 2012;236:79–87. doi: 10.1016/j.expneurol.2012.03.024. [DOI] [PubMed] [Google Scholar]
- 19.Wang ZF, Tang XC. Huperzine A protects C6 rat glioma cells against oxygen-glucose deprivation-induced injury. FEBS Lett. 2007;581:596–602. doi: 10.1016/j.febslet.2007.01.016. [DOI] [PubMed] [Google Scholar]
- 20.Zhang HY, Tang XC. Neuroprotective effects of huperzine A: new therapeutic targets for neurodegenerative disease. Trends Pharmacol Sci. 2006;27:619–625. doi: 10.1016/j.tips.2006.10.004. [DOI] [PubMed] [Google Scholar]
- 21.Oltvai ZN, Milliman CL, Korsmeyer SJ. Bcl-2 heterodimerizes in vivo with a conserved homolog, Bax, that accelerates programmed cell death. Cell. 1993;74:609–619. doi: 10.1016/0092-8674(93)90509-o. [DOI] [PubMed] [Google Scholar]