

Abstract
Background
Neurodevelopmental disorders are impairments of brain function that affect emotion, learning, and memory. Copy number variations of contactin genes (CNTNs), including CNTN3, CNTN4, CNTN5, and CNTN6, have been suggested to be associated with these disorders. However, phenotypes have been reported in only a handful of patients with copy number variations involving CNTNs.
Methods
From January 2009 to January 2013, 3724 patients ascertained through the University of Pittsburgh Medical Center were referred to our laboratory for clinical array comparative genomic hybridization testing. We screened this cohort of patients to identify individuals with the 3p26.3 copy number variations involving the CNTN6 gene, and then retrospectively reviewed the clinical information and family history of these patients to determine the association between the 3p26.3 copy number variations and neurodevelopmental disorders.
Results
Fourteen of the 3724 patients had 3p26.3 copy number variations involving the CNTN6 gene. Thirteen of the 14 patients with these CNTN6 copy number variations presented with various neurodevelopmental disorders including developmental delay, autistic spectrum disorders, seizures and attention deficit hyperactivity disorder. Family history was available for 13 of the 14 patients. Twelve of the thirteen families have multiple members with neurodevelopmental and neuropsychiatric disorders including attention deficit hyperactivity disorder, seizures, autism spectrum disorder, intellectual disability, schizophrenia, depression, anxiety, learning disability, and bipolar disorder.
Conclusions
Our findings suggest that deletion or duplication of the CNTN6 gene is associated with a wide spectrum of neurodevelopmental behavioral disorders.
Keywords: 3p26.3 CNV, Array CGH, CNTNs, CNTN6, Microdeletion, Microduplication, Neurodevelopmental disorders, Neuropsychiatric disorders
Background
Contactins (CNTNs) are members of a protein subfamily of neural immunoglobulin (Ig) domain-containing cell adhesion molecules, which may play a role in the formation of axon connections in the developing nervous system. The CNTN4 and CNTN6 genes are located at the distal part of the short arm of chromosome 3 (3p26.3). The CNTN4 gene has been proposed as one of the critical genes for chromosome 3pter-p25 deletion syndrome [1]. The characteristic features associated with this syndrome include prenatal and postnatal growth retardation, developmental delay (DD), intellectual disabilities (ID), hypotonia, and microcephaly (OMIM #613792). Gene association studies have shown the involvement of contactin genes (CNTNs) in autism spectrum disorders (ASDs) [2, 3]. The disruption of a single copy of the CNTN4 by a de novo balanced translocation has been reported in a boy with some features of 3p deletion syndrome including DD, mild ID, hypotonia, ptosis, and ASD [4, 5]. In addition, chromosome 3 copy number variations (CNVs) involving the CNTN4 gene have been reported to be associated with ASD in a few patients without any other classic 3p deletion syndrome phenotype in three independent studies using genome-wide SNP genotyping or microarray analysis [6–8]. The CNTN6 gene, which encodes another member of the contactin family, mapped just distal to the CNTN4 gene is also deleted in the 3p deletion syndrome. A study by Cui et al. [9] shows that the CNTN6 (NB-3) participates in the generation of oligodendrocytes by acting as a ligand of NOTCH1 to promote NOTCH1 activation through the released notch intracellular domain (NICD) and subsequent translocation to the nucleus. The expression of Cntn6 in the mouse brain is at a low level during embryogenesis but increases significantly after birth [10]. Northern blot analysis showed that the level of CNTN6 expression in human adult brain was the highest in the cerebellum, followed by the thalamus and subthalamic nucleus, and was lower in the corpus callosum, caudate nucleus, and spinal cord [11]. Moreover, studies have shown that Cntn6−/− deficient mice have impaired motor coordination and abnormal apical dendrite projections of deep layer pyramidal neurons in the visual cortex [12, 13]. In addition, a recent study indicated that CNTN4, CNTN5, and CNTN6 proteins may be a part of the heteromeric receptor complexes and serve as ligands themselves [14]. Therefore, deletion or duplication of the CNTN6 gene may affect the function of the receptor complex and cause malfunction of the brain and nervous system. These CNVs involving the CNTN5 or CNTN6 gene alone have also been reported in a handful of patients with either ASD or ID or DD [15–18]. However, this remains controversial. A study involving a single family concluded that the deletion of the CNTN6 gene is not associated with dysmorphic features and ID [19]. In this study, we report 3p26.3 CNVs encompassing the CNTN6 gene in 14 patients. Thirteen of the 14 patients have neurodevelopmental disorders (NDDs) and seven of the 14 patients have dysmorphic features.
Methods
Patient ascertainment
From January 2009 to January 2013, 3724 patients ascertained through the University of Pittsburgh Medical Center (UPMC) were referred to our laboratory for clinical array comparative genomic hybridization (aCGH) testing because of the presence of multiple congenital anomalies, heart defect, short stature, DD, ID, ASD, seizures (SZs), or other unexplained anomalies. The clinical information and family history of the patients with the 3p26.3 CNV were retrospectively reviewed. This study was approved by the University of Pittsburgh Institutional Review Board (IRB#: PRO13090288).
Array-based comparative genomic hybridization analysis
Oligonucleotide-based whole-genome aCGH was performed using a NimbleGen 135K oligonucleotide array, SignatureChip Oligo Solution version 2.0, which was custom-designed by Signature Genomic Laboratories (Spokane, WA, USA) and made by Roche NimbleGen (Madison, WI, USA) as previously described [20]. Results were displayed by custom aCGH analysis software Genoglyphix version 2.6 (Signature Genomic Laboratories).
Chromosome analysis
High-resolution G-banded chromosome analysis was performed on the peripheral blood specimen following routine protocols. A minimum of 20 metaphase cells were examined for both numerical and structural chromosomal anomalies, and two or more karyograms were created using the Ikaros Karyotyping System (MetaSystems, Waltham, MA, USA) on each patient.
Fluorescence in situ hybridization analysis
Fluorescence in situ hybridization (FISH) analysis was performed on metaphase spreads of cultured peripheral blood lymphocytes from patients and their parents using standard procedures. FISH probes were made from the RPCI-11 human genomic library (Invitrogen, Carlsbad, CA, USA) using Nick Translation Kit (Abbott Molecular Inc., Des Plaines, IL, USA). Images were captured using the Isis FISH Imaging System v5.3 software (MetaSystems, Waltham, MA, USA).
Results
Fourteen of the 3724 patients (0.4 %) were found to have CNVs involving the CNTN6 gene. Seven of the 14 patients had single copy loss in the 3p26.3 region involving deletion of the entire CNTN6 gene (patients 1 and 2) or intragenic CNTN6 deletion (patients 3–7). Five of the 14 patients had a single copy gain in the 3p26.3 region involving an intragenic duplication of the CNTN6 gene (patients 8–12). The remaining two patients (patients 13 and 14) had duplications encompassing CHL1/CNTN6 and CHL1/CNTN6/CNTN4 genes, respectively. The representative aCGH profiles (patient 2 and patient 9) are shown in Fig. 1a, b. The schematic representation of the array-CGH results in 14 patients is shown in Fig. 2. The size of the deletions/duplications involving the CNTN6 gene alone ranged from 93.95 kb to 1.23 Mb. No other known pathogenic CNVs or CNVs with unclear clinical significance were observed in any of these patients. The laboratory and clinical findings of these patients and their family histories are summarized in Tables 1 and 2. The genomic positions of the deletion or the duplication for each patient are illustrated in Fig. 2 and summarized in Tables 1 and 2. The karyotype was normal in all 14 patients. Parental FISH analysis was offered to the families. However, only four families completed the FISH testing (family 6, 8, 11, and 13). The results of the parental FISH analysis indicated that these CNVs are inherited.
Fig. 1.
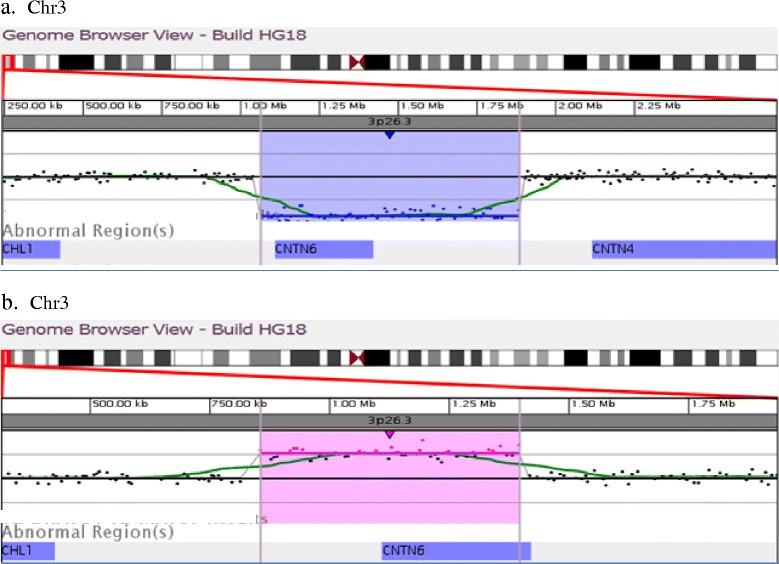
a aCGH profile showing an approximately 821-kb deletion in the 3p26.3 region (1,063,289–1,884,842) detected in patient 2; genome browser [hg18] showing the CNTN6 in the deleted region. b aCGH profile showing an approximately 541-kb duplication in the 3p26.3 region (855,662–1,397,384) detected in patient 9; genome browser [hg18] showing CNTN6 in the duplicated region
Fig. 2.
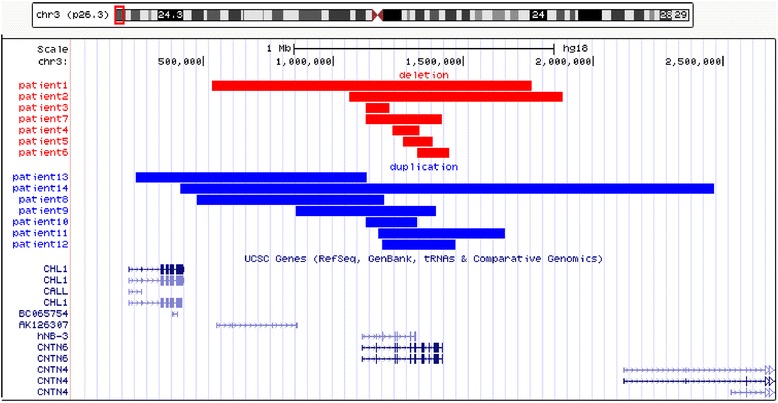
Schematic representation of the array-CGH results of the 14 patients with deletion or duplication of the 3p26.3 region. The ideogram of chromosome 3 and the 3p26.3 highlighted in a small red box is shown on top. The red bars represent deletion sizes, and the blue bars represent duplication sizes. The genes in the interval are shown at the bottom
Table 1.
Laboratory findings, clinical features, and family histories of seven patients with 3p26.3 deletion involving CNTN6 gene
| Patients | Age (year) | CNV sizes | Coordinates (hg18) | Inheritance | OMIM genes | Types of deletion | Reasons for referral | Other features | Family history of NDDs and/or neuropsychiatric disorders |
|---|---|---|---|---|---|---|---|---|---|
| 1 | 5 | 1.23 Mb | 535,618–1,763,192 | Unknown | CNTN6 | Whole gene | ASD, DD | Tantrums, aggression | Father: LD; mother: SZs, hearing loss, and mental health issue |
| 2 | 3 | 821.55 kb | 1,063,289–1,88,4842 | Unknown | CNTN6 | Whole gene | DD, SZs | Nystagmus, macrocephaly, frontal bossing, down-slanting palpebral fissures, high-arched palate | Paternal aunt: ID; maternal aunt, cousin, grandmother, and grandaunt: SZs |
| 3 | 11 | 93.95 kb | 1,124,286–1,218,241 | Unknown | CNTN6 | Exon 2 | DD, SZs, ID | Strabismus, regression in skills, headache | Father: LD; mother: SZs, schizophrenia, migraines; multiple members of maternal side: LD, ASD, SZs, depression, anxiety, bipolar disorder, schizophrenia |
| 4 | 6 | 106.77 kb | 1,227,323–1,334,091 | Unknown | CNTN6 | Exons 3–7 | DD, ASD | Tantrums | Maternal grandmother: depression |
| 5 | 2 | 115.87 kb | 1,266,963–1,382,828 | Unknown | CNTN6 | Exons 5–12 | DD, SZs | Schizencephaly, hydrocephaly, hydronephrosis, diabetes insipidus, hypothyroidism, right-sided spasticity and hemiparesis, cardiomyopathy | Adopted into family; adopted sibling: schizophrenia |
| 6 | 6 | 125.24 kb | 1,322,292–1,447,530 | Paternally inherited | CNTN6 | Exons 8–23 | SZs | Abnormal EEG | Father: del. CNTN6, normal phenotype; sister: del. CNTN6, SZs; mother and brother: without del. CNTN6, phenotypically normal; a maternal aunt: SZs |
| 7 | 1 | 244 kb | 1,124,286–1,419,226 | Unknown | CNTN6 | Exons 2–23 | Heart block | No | No |
ASD autism spectrum disorder, CNV copy number variation, del. deletion, DD developmental delay, EEG electroencephalogram, ID intellectual disability, LD learning disability, NDD neurodevelopmental disorder, SZs seizures
Table 2.
Laboratory findings, clinical features, and family histories of seven patients with 3p26.3 duplication involving CNTN6 gene
| Patients | Age (year) | CNV sizes | Coordinates (hg18) | Inheritance | OMIM genes | Types of deletion | Reasons for referral | Other features | Family history of NDDs and/or neuropsychiatric disorders |
|---|---|---|---|---|---|---|---|---|---|
| 8 | 7 | 718.82 kb | 476,636–1,195,459 | Maternally inherited | CNTN6 | Upstream and exons 1–2 | DD, SZs | ADHD, DBD, abnormal EEG, macrocephaly, epicanthal folds, high and wide nasal bridge, broad nasal tip, large central incisors | Mother: normal phenotype; father: schizophrenia, alcoholic; sister: DD; maternal half brother: ADHD, hearing loss, anxiety |
| 9 | 8 | 541.62 kb | 855,662–1,397,284 | Unknown | CNTN6 | Upstream and exons 1–17 | DD | Short stature, reflux | Father: depression; mother: ADHD, anxiety |
| 10 | 6 | 199.60 kb | 1,124,286–1,323,884 | Unknown | CNTN6 | Exons 2–7 | DD | Short stature, reflux, joint hypermobility, microcephaly, low anterior hairline, second and third toe syndactyly | A sister: ASD, microcephaly; another sister: microcephaly; brother: ASD; paternal half brother: ADHD; both parents: microcephaly; paternal grandmother: seizures |
| 11 | 7 | 489.60 kb | 1,172622–1,662,216 | Paternally inherited | CNTN6 | Exons 3–23 and downstream | DD, ASD, SZs | LD, ADHD, ODD, VSD, feeding difficulties, failure to thrive, brachycephaly, upswept anterior hairline, unusual large hallux, and short second toe | Father: bipolar disorder and ADHD; brother: ADHD, ODD; paternal twin half sisters: DD; mother: bipolar disorder, migraines; maternal grandaunt: migraine |
| 12 | 9 | 281 kb | 1,189,367–1,470,327 | Unknown | CNTN6 | Exons 4–23 and downstream | LD | No | Father: ID, ADHD, bipolar disorder, depression; mother: bipolar disorder, depression; sibling: LD; sister: ID |
| 13 | 15 | 886.76 kb | 243,741–1,130,505 | Maternally inherited | CHL1, CNTN6 | Whole CHL1 and exons 1 of CNTN6 | Obesity, ADHD, bipolar | OCD, migraine, scoliosis, fibromyalgia, rheumatoid arthritis, asthma, irritable bowel syndrome | Mother: migraines, fibromyalgia, psychiatric problem; twin brother and sister: bipolar disorder; maternal half sister: ADHD, psychiatric problems; paternal grandmother: migraines |
| 14 | 16 | 2.05 Mb | 413,294–2,465,270 | Unknown | CHL1, CNTN6, CNTN4 | Exons 23–25 of CHL1, CNTN6 and exons 1–2 of CNTN4 | DD, SZs, dysmorphic | ADHD, OCD, sensorineural hearing loss, asymmetric face, left esotropia, bilateral ptosis, high-arched palate, short stature, single palmar creases, fifth finger clinodactyly, cranial nerve palsy, micropenis | Father: depression; paternal cousin: ASD |
ADHD attention deficit hyperactivity disorder, ASD autism spectrum disorder, CNV copy number variation, DBD disruptive behavior disorders, DD developmental delay, dup duplication, EEG electroencephalography, ID intellectual disability, LD learning disability, NDD neurodevelopmental disorder, OCD, obsessive–compulsive disorder, ODD oppositional defiant disorder, VSD ventricular septal defect, SZs seizures
Thirteen of the 14 patients with these CNVs presented with variable clinical manifestation of neurodevelopmental disorders including ASD in three patients (patients 1, 4, and 11), DD in 10 patients (patients 1–5, 8–11, and 14), SZs in seven patients (patients 2, 3, 5, 6, 8, 11, and 14), and ADHD in four patients (patients 8, 11, 13 and 14). The remaining patient with intragenic deletion of the CNTN6 was a newborn whose development and behavior at 1 year of age was normal. He was tested because of heart block. In addition, two patients had macrocephaly (patients 2 and 8), one had hydrocephaly (patient 5), and one had microcephaly (patient 10). Other malformations/dysmorphisms were also found in six of the 14 patients, which include craniofacial abnormalities (asymmetric face: patient 14; high-arched palate: patients 2 and 14; schizencephaly: patient 5; brachycephaly: patient 11), digital anomalies (second and third toe syndactyly: patient 10; unusual large hallux and short second toe: patient 11; fifth finger clinodactyly: patient 14), and eye problems (nystagmus: patient 2; esotropia: patient 14; strabismus: patient 3). Family history was available for 13 of the 14 patients, except patient 5 who was adopted. Family 7 had maternal family history of Sjogren’s syndrome. The remaining 12 families had multiple members with NDDs and neuropsychiatric disorders including ADHD, SZs, ASD, ID, schizophrenia, depression, anxiety, learning disability, and bipolar disorder. The pedigree for each family is presented in Fig. 3, and only neurodevelopmental or psychiatric issues are marked in these pedigrees. More detailed family information is summarized in Tables 1 and 2.
Fig. 3.
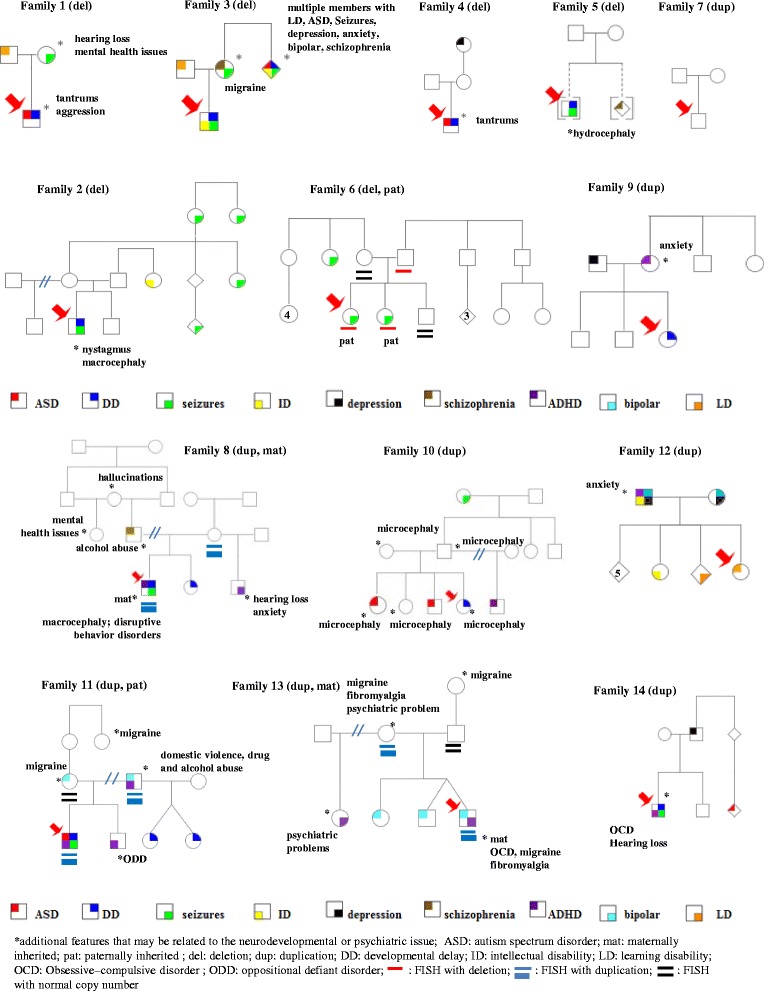
Pedigrees of the 14 families
Discussion
Deletions of both CNTN4 and CNTN6 genes have been reported in the 3p deletion syndrome, which is characterized by low birth weight, growth restriction, DD, ID, hypotonia, and microcephaly. We present here 13 of the 14 patients with neurodevelopmental disorders and CNVs in the 3p26.3 region encompassing the CNTN6 gene. The clinical features include ASD, DD, SZs, and ADHD. These CNVs involving the CNTN6 gene or CNTN6 and CHL1 genes have been reported in five patients in three previous independent studies [15–18]. The clinical findings are consistent with the features in our patients. Moreover, macrocephaly presented in two of the 14 patients has been reported in patients with ASD, while microcephaly presented in one of the 14 patients has been reported in chromosome 3p deletion syndrome [21, 22]. Furthermore, microcephaly was also reported in two previously reported patients with deletion of CNTN6 [16]. In addition to microcephaly, other manifestations of chromosome 3pter-p25 deletion syndrome also exist in one or two of our patients, such as high-arched palate, second and third toe syndactyly, fifth finger clinodactyly, ptosis, joint laxity. and scoliosis (Tables 1 and 2). Syndactyly, clinodactyly and scoliosis, joint laxity and high arched palate were previously reported in patients who have the CNTN6 deletions [16, 18]. Since the genomic location of the CNTN6 gene is within the deletion interval for chromosome 3pter-p25 deletion syndrome, losses of the CNTN6 gene may contribute to the phenotype observed in chromosome 3pter-p25 deletion syndrome. In addition, among the eight patients with gain of copy number in this region five of them had CNV involving partial duplication of the CNTN6 gene. It is unclear whether the expression of CNTN6 gene has been altered in these patients as there is a lack of published functional studies of these CNVs. However, partial duplications of other gene, such as partial duplications of the CHRNA7 gene, have shown to alter the function of the gene product by dominant negative regulation [23]. Unlike 3p terminal deletions which are often de novo events, these CNTN6 deletions or duplications reported previously and found in our patients are interstitial and are more likely inherited. These deletions and duplications vary in size and in location of breakpoints. The lack of segmental duplications in the surrounding sequences suggests that nonallelic homologous recombination is not the mechanism underlying these deletion and duplication events.
It is striking that 12 of the 13 families have positive family history of various NDDs and neuropsychiatric disorders including ADHD, SZs, ASD, ID, schizophrenia, depression, anxiety, learning disability, and bipolar disorder (Fig. 3; Tables 1 and 2). It is known that the NDDs and psychiatric disorders appear to present a disease spectrum and the outcome of the neurodevelopmental process in each individual patient is determined by interactions among genetic, sociocultural, medical, and environmental factors. Therefore, intrafamilial and interfamilial phenotypic heterogeneity and possible incomplete penetrance are expected. Individual and familial comorbidity among SZs, ASD, bipolar disorder, major depression, ADHD, and other psychiatric diagnoses have been documented by large-scale epidemiological studies [24–27]. These studies suggest a possible genetic overlap between these disorders, which could attribute to the familial vulnerability to NDDs and psychiatric disorders [28]. Risk loci shared by major psychiatric disorders have been reported [29, 30]. Recurrent CNVs in synaptic and neurodevelopmental genes have been found to predispose to a wide spectrum of developmental or psychiatric disorders [30–33]. Similar to other CNVs involving synaptic and neurodevelopmental genes, the CNVs disrupting the CNTN6/CNTN4 region have also been reported in patients with intellectual disabilities, cognitive disorders, severe anorexia nervosa, and bipolar disorder in addition to ASD [16, 19, 26, 34]. These published records support our hypothesis of variable expression of the CNVs involving the CNTN6 gene.
Most of the families in our study presented with NDDs and/or psychiatric disorders in both maternal and paternal sides, which makes it difficult to determine the segregation patterns of these CNVs. In family 13, the asymptomatic father has normal copy number for CNTN6; the proband with ADHD and bipolar disorder inherited the duplication of CNTN6 from the mother who has psychiatric problems. In addition, the proband’s maternal half sister has ADHD, and twin brother and full sister have bipolar disorder. However, the copy number for CNTN6 in these individuals was not determined. In family 6, the proband and her sister with seizures have a paternally inherited deletion of CNTN6, and her phenotypically normal mother and brother have normal copy number of CNTN6. Although the father of the proband 6 is an asymptomatic carrier of the deletion, it may indicate an incomplete penetrance. Incomplete penetrance was also observed in family 8 (Fig. 3) and in previously reported families [15, 16, 19]. Parental FISH analysis and family history in families 13 and 6 may suggest that the CNVs segregate with the abnormal phenotypes. However, more extended family studies are needed for segregation analysis in the future. Furthermore, most patients have both maternal and paternal family histories of NDDs and/or psychiatric disorders. Therefore, the possibility of a double hit model for inheritance also exists. Based on our study, the CNTN6 CNVs are rare and presented in about 4 out of 1,000 (14/3,724) patients. At least 40 patients with similar small duplication or deletion involving CNTN6 gene or CNTN6 and CNTN4 genes are reported in the DECIPHER database. These CNVs have also been reported in some phenotypically normal individuals [35]. An expanded CNV morbidity map generated from 29,085 children with developmental delay in comparison to 19,584 healthy controls by Coe et al. [35] showed a frequency of 0.4 % for CNTN6 CNVs (deletion and duplication) in patients with neurodevelopmental disorders and a frequency of 0.3 % in normal controls. Incomplete penetrance and variable expression are common in neurodevelopmental and neurological disorders. Some patients with these CNVs could have mild or normal phenotype. Therefore, these CNVs can be detected in “healthy individuals.” Finding of the CNVs involving CNTN6 gene in normal individuals does not exclude the possibility that these CNVs are risk factors for NDDs. Individuals having a mild phenotype may not be recognized.
Unlike other patients, patients 13 and 14 have a CNV involving duplications of CHL1 and CNTN6 or CHL1, CNTN6, and CNTN 4 genes, respectively. Both patients 13 and 14 have DD, ADHD, oppositional defiant disorder (ODD) and dysmorphic features. In addition, patient 13 has bipolar disorder. Both CHL1 and CNTN4 are expressed in the brain and have been proposed as candidate genes for NDDs [1, 36]. Patient 14 has more dysmorphic features including short stature, high-arched palate, bilateral ptosis, clinodactyly, and micropenis, which are commonly present in patient with chromosome 3pter-p25 deletion syndrome. Our study and previous studies found dysmorphic features are also commonly seen in patients who have the CNV involving CNTN6 gene only [16, 18]. These findings indicate that dosage alterations of CNTN genes may affect the normal development of other tissues or organs in addition to the brain.
Conclusions
We identified 3p26 CNVs involving the deletion/intragenic deletion or duplication/intragenic duplication of CNTN6 gene in 14 patients. Twelve of the 14 patients have the CNV encompassing CNTN6 gene alone. Thirteen of the 14 patients have neurodevelopmental behavioral disorders and remarkable family history for neurodevelopment or neuropsychiatric disorders. Seven of the 14 patients presented with dysmorphic features. Our findings provide more evidence to support that deletion or duplication of the CNTN6 gene is associated with a wide spectrum of neurodevelopmental behavioral disorders. These genotype phenotype correlations pave the way to further investigate the role of CNTN6 in neurodevelopmental disorders.
Abbreviations
- aCGH
array comparative genomic hybridization
- ADHD
attention deficit hyperactivity disorder
- ASD
autism spectrum disorder
- CNTNs
genes for contactin
- CNVs
copy number variations
- DBD
disruptive behavior disorders
- DD
developmental delay
- dup
duplication
- EEG
electroencephalography
- ID
intellectual disability
- LD
learning disability
- NDD
neurodevelopmental disorder
- OCD
obsessive–compulsive disorder
- ODD
oppositional defiant disorder
- VSD
ventricular septal defect
- SZs
seizures
Footnotes
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
JH designed this study; carried out the microarray analysis, medical chart review, and data analysis; and drafted the manuscript. JL carried out the screening of the array database to identify those patients with 3p26.3 CNVs and literature search. JS helped in the review of the patients’ medical charts. MS, SAY, and US carried out the array analysis and participated in the data analysis and preparation of the manuscript. SK, MS, SAY, and US carried out the critical reading and editing of the manuscript. All the authors have read and approved the final manuscript.
Contributor Information
Jie Hu, Email: jhu@upmc.edu.
Jun Liao, Email: jun.liao@mssm.edu.
Malini Sathanoori, Email: MSathanoori@pathgroup.com.
Sally Kochmar, Email: kochsj@upmc.edu.
Jessica Sebastian, Email: Jessica.Hartman@chp.edu.
Svetlana A. Yatsenko, Email: yatsenkosa@mail.magee.edu
Urvashi Surti, Email: surtus@mail.magee.edu.
References
- 1.Dijkhuizen T, van Essen T, van der Vlies P, Verheij JB, Sikkema-Raddatz B, van der Veen AY, et al. FISH and array-CGH analysis of a complex chromosome 3 aberration suggests that loss of CNTN4 and CRBN contributes to mental retardation in 3pter deletions. Am J Med Genet A. 2006;140(22):2482–2487. doi: 10.1002/ajmg.a.31487. [DOI] [PubMed] [Google Scholar]
- 2.Arking DE, Cutler DJ, Brune CW, Teslovich TM, West K, Ikeda M, et al. A common genetic variant in the neurexin superfamily member CNTNAP2 increases familial risk of autism. Am J Hum Genet. 2008;82:160–164. doi: 10.1016/j.ajhg.2007.09.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Poot M. A candidate gene association study further corroborates involvement of contactin genes in autism. Mol Syndromol. 2014;5(5):229–35. doi: 10.1159/000362891. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Fernandez T, Morgan T, Davis N, Klin A, Morris A, Farhi A, et al. Disruption of contactin 4 (CNTN4) results in developmental delay and other features of 3p deletion syndrome. Am J Hum Genet. 2004;74(6):1286–1293. doi: 10.1086/421474. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Fernandez T, Morgan T, Davis N, Klin A, Morris A, Farhi A, et al. Disruption of contactin 4 (CNTN4) results in developmental delay and other features of 3p deletion syndrome. Am J Hum Genet. 2008;82(6):1385. doi: 10.1016/j.ajhg.2008.04.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Cottrell CE, Bir N, Varga E, Alvarez CE, Bouyain S, Zernzach R, et al. Contactin 4 as an autism susceptibility locus. Autism Res. 2011;4(3):189–199. doi: 10.1002/aur.184. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Glessner JT, Wang K, Cai G, Korvatska O, Kim CE, Wood S, et al. Autism genome-wide copy number variation reveals ubiquitin and neuronal genes. Nature. 2009;459(7246):569–573. doi: 10.1038/nature07953. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Guo H, Xun G, Peng Y, Xiang X, Xiong Z, Zhang L, et al. Disruption of contactin 4 in two subjects with autism in Chinese population. Gene. 2012;505(2):201–205. doi: 10.1016/j.gene.2012.06.051. [DOI] [PubMed] [Google Scholar]
- 9.Cui XY, Hu QD, Tekaya M, Shimoda Y, Ang BT, Nie DY, et al. NB-3/Notch1 pathway via Deltex1 promotes neural progenitor cell differentiation into oligodendrocytes. J Biol Chem. 2004;279(24):25858–25865. doi: 10.1074/jbc.M313505200. [DOI] [PubMed] [Google Scholar]
- 10.Lee S, Takeda Y, Kawano H, Hosoya H, Nomoto M, Fujimoto D, et al. Expression and regulation of a gene encoding neural recognition molecule NB-3 of the contactin/F3 subgroup in mouse brain. Gene. 2000;245(2):253–266. doi: 10.1016/S0378-1119(00)00031-7. [DOI] [PubMed] [Google Scholar]
- 11.Kamei Y, Tsutsumi O, Taketani Y, Watanabe K. cDNA cloning and chromosomal localization of neural adhesion molecule NB-3 in human. J Neurosci Res. 1998;51(3):275–283. doi: 10.1002/(SICI)1097-4547(19980201)51:3<275::AID-JNR1>3.0.CO;2-D. [DOI] [PubMed] [Google Scholar]
- 12.Takeda Y, Akasaka K, Lee S, Kobayashi S, Kawano H, Murayama S, et al. Impaired motor coordination in mice lacking neural recognition molecule NB-3 of the contactin/F3 subgroup. J Neurobiol. 2003;56(3):252–265. doi: 10.1002/neu.10222. [DOI] [PubMed] [Google Scholar]
- 13.Ye H, Tan YL, Ponniah S, Takeda Y, Wang SQ, Schachner M, et al. Neural recognition molecules CHL1 and NB-3 regulate apical dendrite orientation in the neocortex via PTP alpha. EMBO J. 2008;27(1):188–200. doi: 10.1038/sj.emboj.7601939. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Zuko A, Bouyain S, van der Zwaag B, Burbach JP. Contactins: structural aspects in relation to developmental functions in brain disease. Adv Protein Chem Struct Biol. 2011;84:143–180. doi: 10.1016/B978-0-12-386483-3.00001-X. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.van Daalen E, Kemner C, Verbeek NE, van der Zwaag B, Dijkhuizen T, Rump P, et al. Social Responsiveness Scale-aided analysis of the clinical impact of copy number variations in autism. Neurogenetics. 2011;12(4):315–323. doi: 10.1007/s10048-011-0297-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Kashevarova AA, Nazarenko LP, Schultz-Pedersen S, Skryabin NA, Salyukova OA, Chechetkina NN, et al. Single gene microdeletions and microduplication of 3p26.3 in three unrelated families: CNTN6 as a new candidate gene for intellectual disability. Mol Cytogenet. 2014;7(1):97. doi: 10.1186/s13039-014-0097-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Pinto D, Pagnamenta AT, Klei L, Anney R, Merico D, Regan R, et al. Functional impact of global rare copy number variation in autism spectrum disorders. Nature. 2010;466(7304):368–72. doi: 10.1038/nature09146. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Te Weehi L, Maikoo R, Mc Cormack A, Mazzaschi R, Ashton F, Zhang L, et al. Microduplication of 3p26.3 implicated in cognitive development. Case Rep Genet. 2014;2014:295359. doi: 10.1155/2014/295359. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Moghadasi S, van Haeringen A, Langendonck L, Gijsbers AC, Ruivenkamp CA. A terminal 3p26.3 deletion is not associated with dysmorphic features and intellectual disability in a four-generation family. Am J Med Genet A. 2014;164A(11):2863–8. doi: 10.1002/ajmg.a.36700. [DOI] [PubMed] [Google Scholar]
- 20.Liao J, DeWard SJ, Madan-Khetarpal S, Surti U, Hu J. A small homozygous microdeletion of 15q13.3 including the CHRNA7 gene in a girl with a spectrum of severe neurodevelopmental features. Am J Med Genet A. 2011;155A(11):2795–2800. doi: 10.1002/ajmg.a.34237. [DOI] [PubMed] [Google Scholar]
- 21.Fidler DJ, Bailey JN, Smalley SL. Macrocephaly in autism and other pervasive developmental disorders. Dev Med Child Neurol. 2000;42(11):737–740. doi: 10.1017/S0012162200001365. [DOI] [PubMed] [Google Scholar]
- 22.Gunnarsson C, Foyn Bruun C. Molecular characterization and clinical features of a patient with an interstitial deletion of 3p25.3-p26.1. Am J Med Genet A. 2010;152A(12):3110–3114. doi: 10.1002/ajmg.a.33353. [DOI] [PubMed] [Google Scholar]
- 23.Araud T, Graw S, Berger R, Lee M, Neveu E, Bertrand D, Leonard S. The chimeric gene CHRFAM7A, a partial duplication of the CHRNA7 gene, is a dominant negative regulator of α7*nAChR function. Biochem Pharmacol. 2011;82(8):904–14. doi: 10.1016/j.bcp.2011.06.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Clarke MC, Tanskanen A, Huttunen MO, Clancy M, Cotter DR, Cannon M. Evidence for shared susceptibility to epilepsy and psychosis: a population-based family study. Biol Psychiatry. 2012;71(9):836–839. doi: 10.1016/j.biopsych.2012.01.011. [DOI] [PubMed] [Google Scholar]
- 25.Daniels JL, Forssen U, Hultman CM, Cnattingius S, Savitz DA, Feychting M, et al. Parental psychiatric disorders associated with autism spectrum disorders in the offspring. Pediatrics. 2008;121(5):e1357–1362. doi: 10.1542/peds.2007-2296. [DOI] [PubMed] [Google Scholar]
- 26.Kerner B, Lambert CG, Muthen BO. Genome-wide association study in bipolar patients stratified by co-morbidity. PLoS One. 2011;6(12):e28477. doi: 10.1371/journal.pone.0028477. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Qin P, Xu H, Laursen TM, Vestergaard M, Mortensen PB. Risk for schizophrenia and schizophrenia-like psychosis among patients with epilepsy: population based cohort study. BMJ. 2005;331(7507):23. doi: 10.1136/bmj.38488.462037.8F. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Mitchell KJ. The genetics of neurodevelopmental disease. Curr Opin Neurobiol. 2011;21(1):197–203. doi: 10.1016/j.conb.2010.08.009. [DOI] [PubMed] [Google Scholar]
- 29.Cross-Disorder Group of the Psychiatric Genomics Consortium. Identification of risk loci with shared effects on five major psychiatric disorders: a genome-wide analysis. Lancet. 2013; 10.1016/S0140-6736(12)62129-1. [DOI] [PMC free article] [PubMed]
- 30.Serretti A, Fabbri C. Shared genetics among major psychiatric disorders. Lancet. 2013;381(9875):1339–41. doi: 10.1016/S0140-6736(13)60223-8. [DOI] [PubMed] [Google Scholar]
- 31.Ching MS, Shen Y, Tan WH, Jeste SS, Morrow EM, Chen X, et al. Deletions of NRXN1 (neurexin-1) predispose to a wide spectrum of developmental disorders. Am J Med Genet B Neuropsychiatr Genet. 2010;153B(4):937–947. doi: 10.1002/ajmg.b.31063. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Burbach JP, van der Zwaag B. Contact in the genetics of autism and schizophrenia. Trends Neurosci. 2009;32(2):69–72. doi: 10.1016/j.tins.2008.11.002. [DOI] [PubMed] [Google Scholar]
- 33.Friedman JI, Vrijenhoek T, Markx S, Janssen IM, van der Vliet WA, Faas BH, et al. CNTNAP2 gene dosage variation is associated with schizophrenia and epilepsy. Mol Psychiatry. 2008;13(3):261–266. doi: 10.1038/sj.mp.4002049. [DOI] [PubMed] [Google Scholar]
- 34.Wang K, Zhang H, Bloss CS, Duvvuri V, Kaye W, Schork NJ, et al. A genome-wide association study on common SNPs and rare CNVs in anorexia nervosa. Mol Psychiatry. 2011;16(9):949–959. doi: 10.1038/mp.2010.107. [DOI] [PubMed] [Google Scholar]
- 35.Coe BP, Witherspoon K, Rosenfeld JA, van Bon BW, Vulto-van Silfhout AT, Bosco P. Refining analyses of copy number variation identifies specific genes associated with developmental delay. Nat Genet. 2014;46(10):1063–71. doi: 10.1038/ng.3092. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Frints SG, Marynen P, Hartmann D, Fryns JP, Steyaert J, Schachner M, et al. CALL interrupted in a patient with non-specific mental retardation: gene dosage-dependent alteration of murine brain development and behavior. Hum Mol Genet. 2003;12(13):1463–1474. doi: 10.1093/hmg/ddg165. [DOI] [PubMed] [Google Scholar]