

Abstract
In order to fully understand the orientation behaviour of migrating birds, it is important to understand when birds set their travel direction. Departure directions of migratory passerines leaving stopover sites are often assumed to reflect the birds' intended travel directions, but this assumption has not been critically tested. We used data from an automated radiotelemetry system and a tracking radar at Falsterbo peninsula, Sweden, to compare the initial orientation of departing songbirds (recorded by radiotelemetry) with the orientation of songbird migrants in climbing and level flight (recorded by radar). We found that the track directions of birds at high altitudes and in level flight were more concentrated than the directions of departing birds and birds in climbing flight, which indicates that the birds adjust their travelling direction once aloft. This was further supported by a wide scatter of vanishing bearings in a subsample of radio-tracked birds that later passed an offshore radio receiver station 50 km southeast of Falsterbo. Track directions seemed to be more affected by winds in climbing compared with level flights, which may be explained by birds not starting to partially compensate for wind drift until they have reached cruising altitudes.
Keywords: bird migration, orientation, departure direction, radiotelemetry, radar
1. Introduction
Birds are known to use multiple cues in order to determine directions during migration ([1], and references therein) and are expected to decide and take off in the migratory direction when leaving stopovers. This departure direction is often interpreted as the birds' intended travelling direction and has been demonstrated to be related to a number of external and internal factors. When and how often birds adjust their travel direction during migration are important factors to know in order to understand their orientation behaviour.
Wind conditions have been shown to have a major influence on bird migration [2,3] and birds en route are subjected to lateral drift caused by winds to varying degrees [2,4]. Shorebirds [5] and terns [6] have been observed not to compensate for lateral drift during climbing flights and departure over sea, but in contrast partially or fully compensate for drift at high altitudes and in flights over land. It is unclear whether this pattern is also true for passerines.
In this study, we tested the hypothesis that passerine migrants maintain their initial direction from departure also during their succeeding migratory flights by comparing radio telemetry and radar data of birds departing and migrating at the Falsterbo peninsula, Sweden. The alternative hypothesis was that birds make important course changes after take-off. We further examined the vanishing bearings of birds leaving the Falsterbo peninsula and later passing the same reference point (figure 1h), to see whether they adjusted their directions after departure. In order to investigate a possible cause of a shift in direction, we investigated how winds affected the paths of birds at departure and in climbing and level flight.
Figure 1.

(a) Mean vector lengths (r-values) of the track directions (radar) of birds in level and climbing flight and vanishing bearings (radiotelemetry) at different altitudes demonstrate a higher concentration of track directions at higher altitudes and in level flight. Circular distributions illustrating the pattern of adjustment of direction once at cruising altitudes: (b) vanishing bearing (mean ± 95% CI = 145 ± 16°), track direction of (c) climbing birds with altitude less than 800 m (171 ± 25°), (d) climbing birds with altitude more than 800 m (186 ± 10°), and birds with level flights at altitudes, (e) less than 300 m (212 ± 14°), (f) 300–800 m (210 ± 5°) and (g) more than 800 m (202 ± 4°). A map (h) of the southern Baltic Sea with Falsterbo (black star) and the offshore receiver station (white star), including the distribution of vanishing bearing of birds that passed the offshore station. Arrows indicate mean direction, r-values and 95% CI; all circular distributions have p(Rayleigh) < 0.001; for statistics of the concentrations, see table 1.
2. Material and methods
The study was performed in Falsterbo, Sweden (55°38′ N, 12°82′ E) during autumn 2009–2012 (radiotelemetry) and 2009–2011 (radar).
(a). Radiotelemetry
The radiotelemetry system consisted of three automatic receiver stations (SRX600; Lotek Wireless, Newmarket Ontario, Canada) with an additional offshore receiver placed 47 km southeast from Falsterbo (for set-up, monitoring regime and transmitter information, see [7]). Only night-time departures (18.00–06.00 CET) were included in the analysis, and a vanishing bearing was calculated from the bird's last 10 min on the peninsula (mean measurement bias of vanishing bearings was 9.5°; for details, see the electronic supplementary material). It should be noted that departure directions based on radiotelemetry data are rather coarse estimates, because of the above-mentioned measurement bias and also because of the parallax error (deviation between vanishing and departure direction). However, we expect the parallax error in our data to be small since the birds are expected to start their departure flights close to the receiver stations. This is supported by a high directional consistency in the vast majority of cases during the final 10 min radiotelemetry recordings of the migrants' departure flights. Hence, radiotelemetry data can be used to add supportive evidence to the more precise directional radar data about the birds' flight directions at the peninsula (electronic supplementary material).
Four species of nocturnal migrants were equipped with radio transmitters: song thrush (Turdus philomelos), European robin (Erithacus rubecula), willow warbler (Phylloscopus trochilus) and garden warbler (Sylvia borin). All are considered southwesterly migrants from southern Sweden according to ringing recoveries [8]. We selectively tagged individuals of different ages (based on plumage characteristics) and with different amounts of stored fat (fat class 1–8 on a 10° scale of 0–9). We used weather data collected by the Swedish Meteorological and Hydrological Institute (SMHI) at Falsterbo Bird Observatory every third hour. The data from the weather observation nearest in time to each bird's departure from the peninsula were included in the analysis.
(b). Tracking radar
We used a customized X-band tracking radar to track individual nocturnal migrants as they flew over the Falsterbo peninsula. The wing beat signature of the radar echo was used to identify targets using bounding flight (exclusively used by passerines) that were included in this analysis. The radar was operated from sunset to a few hours after midnight on selected nights. Every tracking night, exact wind profiles were recorded by tracking weather balloons. For details on radar operation and data processing, see [9]. The radar tracks were subdivided into climbing flights (vertical speed, Vz > 0.75 m s−1) and level flights (−0.75 < Vz < 0.75 m s−1). Descending flights (Vz < −0.75 m s−1) were excluded. Climbing flight was further divided into two altitude categories: less than 800 m and more than 800 m; and level flights divided into three categories: less than 300 m, 300–800 m and more than 800 m in the analysis on track concentration. Only tracks for which a wind measurement at the same altitude (±150 m) and within 2 h was available were included in the analysis of wind effects.
(c). Statistics
All circular distributions were assessed using Rayleigh tests, and the mean vector length (r) was used as a measure of concentration (0 = uniform circular distribution, 1 = total consistency in directions). To test whether the concentration of departure directions and track directions differed between the different altitudes and flight behaviours, Mardia & Jupp tests of equality of concentration parameters were performed (table 1; [10]). To investigate the differences in wind effects between the different categories, we performed pairwise Mardia–Watson–Wheeler tests within each wind category (electronic supplementary material, table S2). All statistical analyses were performed in Oriana 4.0 (Kovach Computing Services, Anglesey, UK) or Matlab R2014b (MathWorks, Natick, MA, USA).
Table 1.
Sample sizes and differences in concentration between the different methods and categories. Mardia & Jupp [10] test of equality of concentration parameter. Above diagonal cells p-values, below diagonal cells z-values.
| radar |
|||||||||
|---|---|---|---|---|---|---|---|---|---|
| climbing flight |
level flight |
||||||||
| N | radio telemetry | <800 m | >800 m | <300 m | 300–800 m | >800 m | |||
| radiotelemetry | 102 | — | <0.05 | n.s. | n.s. | n.s. | <0.001 | ||
| radar | climbing flight | <800 m | 142 | 2.58a | — | <0.001 | <0.05 | <0.001 | <0.001 |
| >800 m | 214 | 0.54b | 3.73a | — | n.s. | n.s. | <0.001 | ||
| level flight | <300 m | 149 | 0.29a | 2.53a | 0.97b | — | <0.05 | <0.001 | |
| 300–800 m | 597 | 1.75b | 5.34b | 1.55b | 2.47b | — | <0.001 | ||
| >800 m | 529 | 3.76b | 7.61b | 4.26b | 4.82b | 3.72b | — | ||
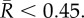
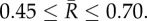
3. Results
Track directions (radar) were more concentrated at higher compared with lower altitudes and in level compared with climbing flight (figure 1 and table 1). The scatter of orientation was largest in birds in climbing flight at altitudes lower than 800 m. Birds in level flight at altitudes exceeding 800 m showed the highest concentration of orientation. Vanishing bearings of the radio-tracked birds were more concentrated than track directions of climbing birds at low altitudes and less concentrated than track directions of birds in level flight at more than 800 m. Furthermore, birds in level flight (radar) were more directed in the expected migratory direction at all altitudes compared to departing (radiotelemetry) and climbing birds.
The mean vanishing bearing (±95% CI; 138 ± 27°) of the 18 birds that departed from Falsterbo and later passed the offshore receiver station (after flight durations between 33 and 123 min) corresponds to the direction towards the offshore receiver (153°) from the peninsula. However, the birds showed a surprisingly large spread in directions (r = 0.61; figure 1h) with respect to their known mutual reference point less than 50 km after departure.
Winds clearly affected the patterns of track directions recorded by radar (figure 2; electronic supplementary material, S2) with mean directions reflecting a degree of wind drift in the tracks. In northeasterly and southeasterly winds, birds in both climbing and level flight seemed equally affected by the winds, while the tracks of climbing birds were more strongly deflected towards east in northwesterly and southwesterly winds. The birds' vanishing bearings differed from track directions of climbing and level flight in all but southwesterly winds by being more scattered.
Figure 2.

Departure and track directions in different winds during autumn migration. Distributions of orientation are shown for four different groups with wind directions from the four different quadrants. Arrows indicate mean direction, r-values and 95% CI; all circular distributions have p(Rayleigh) < 0.001, except the vanishing bearings in NE and SE winds that have p(Rayleigh) > 0.05. Bar charts illustrate percentage of tracks at different altitudes. Significant differences in the concentration of circular distributions are indicated by asterisks; *p < 0.05, **p < 0.01, ***p < 0.001 (for detailed statistics, see electronic supplementary material, S2).
4. Discussion
The differences in concentration between birds in climbing and level flight, as well as between birds at different altitudes, in the radar sample indicate that the migrants made significant adjustments to their travelling direction once they reached cruising altitudes. Even though the radiotelemetry gives a coarser estimate of departure directions than the precise radar data (see the electronic supplementary material), the difference in directional concentration between radio-tracked birds departing the peninsula and birds in level flight at high altitudes adds further support to this conclusion. Furthermore, this pattern is also indicated by earlier observations on the migration pattern at Falsterbo peninsula [11] with higher concentration of movements at higher altitudes. We also indirectly observed substantial changes in direction after departure in the radio-tracked birds that passed the offshore receiver station at the Baltic Sea (figure 1h) after having departed in different directions from Falsterbo. This sheds considerable doubt on the hypothesis that birds maintain the same direction as during take-off, and instead supports the alternative hypothesis that birds adjust their directions once aloft.
The adjustment of direction once aloft may be explained by birds fine-tuning the orientation once in the air or by birds not compensating for wind drift while climbing. Birds climbing in northwesterly and southwesterly winds were deflected more towards the east than birds in level flight in these wind conditions which could imply a larger degree of drift (figure 2). Such a drift effect supports that adjustments of headings are partly owing to wind compensation once aloft. Also, it agrees with earlier observations of shorebirds and terns that did not compensate for wind drift during departure [5,6]. However, birds in both ascending and level flight seemed to be similarly drifted by northeasterly and southeasterly winds.
5. Conclusion
There have been numerous studies of individual and environmental factors affecting birds' departure directions from stopovers, but it is not well known if and for how long birds keep to this direction once aloft. The differences in the concentration of directions during departure and level flight indicate an adjustment of the migratory flight direction once at cruising altitudes. This pattern could only partly be explained by a difference in wind drift compensation between birds in ascending and level flight. When using vanishing bearings (or directions in orientation cages) as intended travel directions, one should keep in mind that they will probably show larger variation than the true travel directions. In the future, longer tracks of departing small passerines, with better resolution, will shed further light on how and when the birds adjust their direction once aloft. This adjustment may be of fundamental importance in the orientation system of nocturnally migrating passerines.
Supplementary Material
Acknowledgements
We thank Thomas Alerstam and Rachel Muheim for support throughout the entire project, Susanne Åkesson and Timothy Coppack for cooperation on the radiotelemetry project and all the staff at Falsterbo Bird Observatory. This is report no. 296 from Falsterbo Bird Observatory.
Ethics
The study was performed with permission from Malmö-Lund Ethical Committee, Sweden (M 27–10).
Data accessibility
Raw data are available via Dryad (http://dx.doi.org/10.5061/dryad.86020) and assigned to the title of this article.
Authors' contributions
S.S. and C.N. initiated the study and collected the data; S.S. did the analyses and wrote the paper with input from C.N.
Competing interests
We declare we have no competing interests.
Funding
This work was supported by grants from the Swedish Research Council (2007-5700 to R.M., 621-2010-5584 and 621-2013-4361 to S.Å., 2009-3589 to T.A. and Linnaeus grant 349-2007-8690 to the Center of Animal Movement Research, CAnMove) from the Royal Physiographical Society in Lund, The Crafoord Society and from Lunds djurskyddsfond. The offshore radiotelemetry station was funded by the German Federal Ministry for the Environment, Nature Conservation, Building and Nuclear Safety (BMUB).
References
- 1.Wiltschko R, Wiltschko W. 2003. Mechanisms of orientation and navigation in migratory birds. In Avian Migration (eds Berthold P, Gwinner E, Sonnenschein E.), pp. 433–456. Berlin, Germany: Springer. [Google Scholar]
- 2.Richardson W. 1990. Wind and orientation of migrating birds: a review. Experientia 46, 416–425. ( 10.1007/BF01952175) [DOI] [PubMed] [Google Scholar]
- 3.Alerstam T. 1979. Wind as selective agent in bird migration. Ornis Scand. 10, 76–93. ( 10.2307/3676347) [DOI] [Google Scholar]
- 4.Liechti F. 2006. Birds: blowin’ by the wind? J. Ornithol. 147, 202–211. ( 10.1007/s10336-006-0061-9) [DOI] [Google Scholar]
- 5.Grönroos J, Green M, Alerstam T. 2012. Orientation of shorebirds in relation to wind: both drift and compensation in the same region. J. Ornithol. 154, 385–392. ( 10.1007/s10336-012-0902-7) [DOI] [Google Scholar]
- 6.Alerstam T. 1985. Strategies of migratory flight, illustrated by arctic and common terns, Sterna paradisea and Sterna hirundo. Migr. Mech. Adapt. Significance 27, 580–603. [Google Scholar]
- 7.Sjöberg S, Alerstam T, Åkesson S, Schulz A, Weidauer A, Coppack T, Muheim R. 2015. Weather and fuel reserves determine departure and flight decisions in passerines migrating across the Baltic Sea. Anim. Behav. 104, 59–68. ( 10.1016/j.anbehav.2015.02.015) [DOI] [Google Scholar]
- 8.Fransson T, Karlsson-Hall S. 2008. Svensk Ringmärkningsatlas. Tättingar, vol. 3 Stockholm, Sweden: SOF förlag. [Google Scholar]
- 9.Nilsson C, Bäckman J, Alerstam T. 2014. Seasonal modulation of flight speed among nocturnal passerine migrants: differences between short- and long-distance migrants. Behav. Ecol. Sociobiol. 68, 1799–1807. ( 10.1007/s00265-014-1789-5) [DOI] [Google Scholar]
- 10.Mardia KV, Jupp PE. 2009. Directional statistics. Hoboken, NJ, USA: Wiley. [Google Scholar]
- 11.Zehnder S, Akesson S, Liechti F, Bruderer B. 2001. Nocturnal autumn bird migration at Falsterbo, South Sweden. J. Avian Biol. 32, 239–248. ( 10.1111/j.0908-8857.2001.320306.x) [DOI] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
Raw data are available via Dryad (http://dx.doi.org/10.5061/dryad.86020) and assigned to the title of this article.