

Abstract
Müller glia can be stimulated to de-differentiate and become proliferating progenitor cells that regenerate neurons in the retina. The signaling pathways that regulate the formation of proliferating Müller glia-derived progenitor cells (MGPCs) are beginning to be revealed. The purpose of this study was to investigate whether Hedgehog (Hh) signaling influences the formation of MGPCs in the chick retina. We find that Hh signaling is increased in damaged retinas where MGPCs are known to form. Sonic Hedgehog (Shh) is normally present in the axons of ganglion cells, but becomes associated with Müller glia and MGPCs following retinal damage. Activation of Hh signaling with recombinant human SHH (rhShh) or smoothened agonist (SAG) increased levels of Ptch1, Gli1, Gli2, Gli3, Hes1 and Hes5, and stimulated the formation of proliferating MGPCs in damaged retinas. In undamaged retinas, SAG or rhShh had no apparent effect upon the Müller glia. However, SAG combined with FGF2 potentiated the formation of MGPCs, whereas SAG combined with IGF1 stimulated the nuclear migration of Müller glia, but not the formation of MGPCs. Conversely, inhibition of Hh signaling with KAAD-cyclopamine, Gli antagonists or antibody to Shh reduced numbers of proliferating MGPCs in damaged and FGF2-treated retinas. Hh signaling potentiates Pax6, Klf4 and cFos expression in Müller glia during the formation of MGPCs. We find that FGF2/MAPK signaling recruits Hh signaling into the signaling network that drives the formation of proliferating MGPCs. Our findings implicate Hh signaling as a key component of the network of signaling pathways that promote the de-differentiation of Müller glia and proliferation of MGPCs.
KEY WORDS: Glia, Hedgehog, Proliferation, Regeneration, Retina
Summary: In the chick retina, Hedgehog signaling contributes to retinal repair by promoting the de-differentiation of Muller glia and the proliferation of Muller glia-derived progenitor cells.
INTRODUCTION
Müller glia are the predominant type of glial cell in the retina and normally provide structural, synaptic and metabolic support to neurons. However, Müller glia can also act as a source of proliferating progenitors to regenerate neurons. Müller glia are distinctly different from progenitor cells based on functions, yet share a significant overlap of transcriptomes with retinal progenitors (Blackshaw et al., 2004). Müller glia are capable of de-differentiating, acquiring a progenitor phenotype and proliferating in response to damage or exogenous growth factors (reviewed by Fischer and Bongini, 2010; Gallina et al., 2014a). Müller glia have been identified as the cellular source of retinal regeneration in birds (Fischer et al., 2002b; Fischer and Reh, 2001), zebrafish (Bernardos et al., 2007; Fausett and Goldman, 2006) and rodents (Karl et al., 2008; Ooto et al., 2004). Similar to the glia in mammalian retina, Müller glia in the chick retina have a limited capacity to generate new neurons. By comparison, Müller glia in the fish retina have an extraordinary capacity to regenerate retinal neurons (reviewed by Gallina et al., 2014a).
Proliferation is an integral step in the formation of Müller glia-derived progenitor cells (MGPCs; Fischer, 2005; Fischer and Reh, 2003b; Reh and Fischer, 2001). The identification of the signaling pathways that influence de-differentiation, proliferation and neurogenesis is key to harnessing the regenerative potential of MGPCs. The Hh signaling pathway may be important to the formation of MGPCs. Hh signaling is known to have many important pleiotropic actions during early eye and retinal development (Dakubo et al., 2003; Levine et al., 1997; Perron et al., 2003; Wang et al., 2002; Zhang and Yang, 2001b). In developing retina, Sonic Hedgehog (Shh) is expressed by the ganglion cells, is exported from the eye via ganglion cell axons, and stimulates the formation of glia in the optic nerve and chiasm (Gao and Miller, 2006; Wallace and Raff, 1999). In the rodent retina, patched (Ptch1; integral membrane receptor for Hh and antagonist of Smo) and smoothened (Smo; co-receptor and activator of intracellular signaling) are expressed by mature Müller glia (Moshiri and Reh, 2004; Nelson et al., 2011; Roesch et al., 2008, 2012). Studies in the embryonic chick have demonstrated regeneration in peripheral regions of prospective retina, and this regeneration requires coordination of Hh and FGF/MAPK signaling (Spence et al., 2007a, 2004). In the rodent, it has been proposed that MGPCs regenerate photoreceptors in response to Hh (Wan et al., 2007). However, this study was performed in rats with retinal damage elicited by systemic N-nitroso-methyl-urea on or before postnatal day (P) 7, with Shh applied before P21, when the proliferation of late-stage retinal progenitors progressively declines and Müller glia have not yet fully differentiated (Close et al., 2005; Nelson et al., 2011). Thus, it remains unknown whether Hh signaling influences mature Müller glia in the rodent retina. In the zebrafish, Shh produced by retinal neurons facilitates regeneration from MGPCs (Sherpa et al., 2014). Here, we investigate Hh signaling after retinal injury or in response to FGF2 treatment, and how Hh signaling impacts the formation of MGPCs in the chick retina in vivo.
RESULTS
Hedgehog signaling in normal and damaged retinas
Shh binds to the transmembrane receptor patched (Ptch) to relieve repression of the transmembrane protein smoothened (Smo) to permit activation of Gli proteins (Ruiz i Altaba et al., 2002). Activated Smo liberates Gli second messengers from a cytoplasmic retention complex permitting translocation of Gli proteins to the nucleus to influence transcription (Hooper and Scott, 2005). Levels of Ptch (Goodrich et al., 1996; Marigo and Tabin, 1996) and Gli proteins (Bai et al., 2002; Litingtung and Chiang, 2000; Marigo et al., 1996) are upregulated in response to Hh signaling. We analyzed whether components of the Hh pathway are expressed in the chick retina using RT-PCR. In normal undamaged retina, we detected mRNA for Shh, Gli1, Gli2, Gli3, Ptch1, Smo and Sufu (Fig. 1A), suggesting that Hh signaling is active in the avian retina under normal conditions. Similar to a previous report (Dakubo et al., 2003), we failed to detect mRNA for desert hedgehog and indian hedgehog in normal and NMDA-damaged retinas or in retinal pigmented epithelium (not shown).
Fig. 1.

Components of the Hh pathway in normal and damaged retinas. (A,B) RT-PCR (A) and qRT-PCR (B) were used to probe for Gapdh, Shh, Gli1, Gli2, Gli3, Ptch1, Smo and Sufu. Beginning at P7, RNA was extracted from normal retinas (A) or from retinas treated with saline or 2 µmol NMDA at 4, 24, 48 and 72 h after treatment (B). Significance of difference (*P<0.05, **P<0.01, ***P<0.005) was determined using a two-tailed Mann–Whitney U-test (n≥4). (C-E″) In situ hybridization was used to detect Ptch1 in control retinas (C) and NMDA-damaged retinas at 24 h (D) and 72 h (E-E″) after treatment. Immunolabeling for PCNA (red) was included to indicate proliferating cells (E-E″). Arrows indicate presumptive Müller glia and small double arrows indicate presumptive NIRG cells. GCL, ganglion cell layer; INL, inner nuclear layer; IPL, inner plexiform layer; ONL, outer nuclear layer. Scale bars: 50 µm (in D for C,D; in E for E alone).
We used quantitative RT-PCR to assay for components of the Hh pathway following NMDA treatment in regions where MGPCs are known to form. Shh was rapidly upregulated within 4 h of NMDA treatment, downregulated at 1 and 2 days after treatment, and returned to control levels by 3 days after treatment (Fig. 1B). By comparison, Gli1 and Gli2 were upregulated from 4 h to 3 days after treatment, and Gli3 was upregulated from 1 to 3 days after treatment (Fig. 1B). Levels of Ptch1 were increased after NMDA treatment with levels peaking at 1 day after treatment, and levels of Smo were elevated from 4 h to 3 days after treatment (Fig. 1B). In older animals (≥P27), we found that levels of Ptch1, Smo and Gli2, but not Gli1, were significantly increased at 1 day after NMDA treatment (supplementary material Fig. S2), suggesting that the upregulation of Hh signaling is not diminished with age. These data suggest that Hh signaling is rapidly upregulated after damage, with signaling peaking at 24 h after treatment. This peak in Hh signaling coincides with when Müller glia are known to de-differentiate and is prior to entry into the cell cycle following NMDA treatment (Fischer and Reh, 2001).
We next examined patterns of Ptch1 expression within normal and damaged retinas. Ptch is expressed at low levels by mature Müller glia and some cells (possibly astrocytes) in the ganglion cell layer (GCL) in the rodent retina (Moshiri and Reh, 2004; Wang et al., 2002). In addition, microarray data from single or sorted Müller glia indicate low levels of expression of Ptch, Smo and Gli genes in mature mouse retina (Roesch et al., 2008, 2012). Although Ptch and Smo are expressed by retinal progenitors in the embryonic chick (Zhang and Yang, 2001a), the identity of the cells that are receptive to Hh in the mature chick retina remains uncertain. In undamaged retina, weak signal for Ptch1 was observed in the proximal inner nuclear layer (INL) and GCL (Fig. 1C). By comparison, 24 h after NMDA-induced damage there was a robust induction of Ptch1 (Fig. 1D), consistent with data from qRT-PCR analysis. We found Ptch1 in the GCL, in cells scattered in the inner plexiform layer (IPL), and in the inner half of the INL (Fig. 1D), suggesting that Ptch1 may be upregulated by ganglion cells, amacrine cells, non-astrocytic inner retinal glial (NIRG) cells in the IPL, and Müller glia. NIRG cells have been characterized as a unique type of glial cell that reside in the inner retina (Fischer et al., 2010). At 72 h after NMDA treatment, Ptch1 remained prevalent in the GCL and INL, and in cells scattered across the IPL and nerve fiber layer (NFL; Fig. 1E). In addition to diffuse labeling across inner retinal layers, Ptch1 appeared to be colocalized with proliferating cell nuclear antigen (PCNA) cells in the IPL/GCL and INL (Fig. 1E), suggesting that Ptch1 is expressed by proliferating NIRG cells and MGPCs.
We next characterized patterns of expression for Shh. Shh immunofluorescence is normally present in the axons of ganglion cells in the NFL (Fig. 2A), consistent with the notion that Shh is normally expressed by ganglion cells and exported out of the eye (Dakubo et al., 2003; Traiffort et al., 2001; Wallace and Raff, 1999). At 2 days after NMDA treatment, Shh immunoreactivity remains prominent in the NFL and appears as distinct puncta in the INL (Fig. 2B). By 3 days after treatment, Shh immunofluorescence is diminished in the NFL, further accumulates as puncta in the INL, and accumulates in the OPL (Fig. 2C). By 5 days after treatment, Shh is no longer present in the NFL, whereas Shh appears in the OPL and in puncta scattered across the INL and ONL (Fig. 2D). The Shh+ puncta were associated with Pax6-expressing MGPCs that accumulate in the distal INL and ONL at 3 days after NMDA treatment (Fig. 2E-I). The Shh immunoreactivity that accumulates in the OPL overlaps, in part, with the axon terminals of calbindin+ cone photoreceptors (Fig. 2J-M). It remains uncertain whether the Shh accumulates within or at the surface of photoreceptor terminals. To investigate further the expression patterns of Shh we performed in situ hybridization. We detected Shh in the GCL of control retinas (Fig. 2N), and this pattern of expression was prominent at 4 h after NMDA treatment (Fig. 2O). However, signal for Shh was diminished at 3 days after NMDA treatment (Fig. 2N), consistent with data from qRT-PCR and immunolabeling experiments.
Fig. 2.
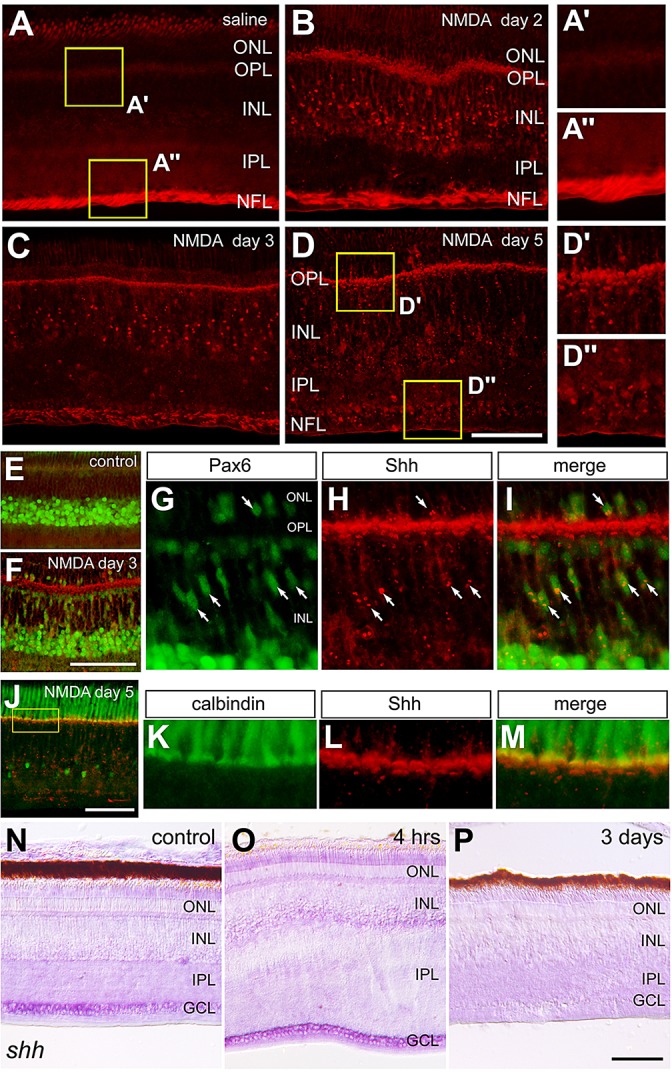
Shh expression in normal and NMDA-damaged retinas. (A-P) Retinas were obtained from eyes that were injected with saline (control; A,E,N) or 2 µmol NMDA and harvested 4 h (O), 2 days (B), 3 days (C,F-I,P) or 5 days (D,J-M) later. Retinal sections were labeled with antibodies to Shh (red; A-M), Pax6 (green; E-I), and calbindin (green; J-M). In situ hybridization for Shh was performed on control retina (N) and NMDA-treated retinas (O,P). Arrows indicate Müller glia labeled for Pax6 and Shh puncta. G-I show high-magnification images of Shh-immunoreactive puncta in Pax6+ nuclei of MGPCs. K-M show high-magnification images of the boxed area in J. GCL, ganglion cell layer; INL, inner nuclear layer; IPL, inner plexiform layer; ONL, outer nuclear layer. Scale bars: 50 µm (in D for A-D; in F for E,F; in J for J alone; in P for P alone).
There was no depletion of neurofilament+ axons or Brn3a+ ganglion cells in NMDA-treated retinas (supplementary material Fig. S1), indicating that the loss of Shh immunoreactivity from the NFL did not result from the destruction of ganglion cells, consistent with findings that NMDA does not destroy ganglion cells in the chick retina (Fischer et al., 1998). We found that the neurofilament+ axons in the NFL were displaced away from the vitreal surface of the retina by the end-feet of GFAP/TopAP+ Müller glia (supplementary material Fig. S1). This probably resulted from the swelling or expansion of glial end-feet in response to damage.
Hh signaling stimulates the proliferation of MGPCs in damaged retinas
We next examined whether Hh signaling influenced the formation of MGPCs. Four consecutive daily injections of rhShh failed to stimulate the formation of MGPCs in undamaged retinas (not shown). Thus, we investigated whether activation of Hh signaling influenced the formation of MGPCs following retinal injury in which components of the Hh pathway are upregulated. A low dose of NMDA failed to stimulate the accumulation of large numbers of BrdU+ MGPCs (Fig. 3A,E). Application of rhShh following a low dose of NMDA significantly increased numbers of proliferating MGPCs in central and peripheral regions of the retina (Fig. 3B,E). Numbers of proliferating NIRG cells and microglia/macrophages were unaffected by rhShh (Fig. 3C-E). Hh signaling has been shown to coordinate with Notch signaling effectors in retinal progenitor cells (Wall et al., 2009), and Notch signaling stimulates the formation of MGPCs (Ghai et al., 2010; Hayes et al., 2007). Thus, we tested whether Notch influenced the formation of MGPCs in retinas treated with NMDA and rhShh. We found that inhibition of Notch with DAPT reduced the number of proliferating MGPCs in retinas treated with NMDA and rhShh (Fig. 3F). We next tested whether smoothened agonist (SAG), which bypasses Ptch and activates Smo (Chen et al., 2002), influences the formation of MGPCs. Similar to rhShh, four consecutive daily intraocular injections of SAG failed to stimulate the formation of MGPCs in undamaged retinas (not shown). Using antibodies to pERK, p38 MAPK, cFos, pS6, vimentin, GFAP and transitin, we failed to find activation of different signaling pathways or reactivity in Müller glia treated with rhShh or SAG (data not shown), suggesting that Müller glia are normally not receptive to rhShh or SAG. However, following a low dose of NMDA, SAG stimulated the accumulation of BrdU+ MGPCs in central and peripheral regions of retina (Fig. 3G). Similar to rhShh, SAG had no influence upon the proliferation of NIRG cells or microglia/macrophages in NMDA-damaged retinas (not shown). In older animals (≥P27), we found that numbers of proliferating MGPCs were significantly increased by SAG following NMDA treatment (supplementary material Fig. S2), suggesting that Hh signaling retains the capacity to stimulate Müller glia to become MGPCs in older damaged retinas. Hh-mediated regeneration of embryonic chick retina is known to require MAPK signaling (Spence et al., 2007b, 2004). Accordingly, we tested whether MAPK signaling influenced the ability of SAG to stimulate the proliferation of MGPCs. We found that inhibition of MAPK signaling with U0126 reduced the number of BrdU-labeled MGPCs in retinas treated with NMDA+SAG (Fig. 3H). U0126 also reduced the number of Sox2+/PCNA+ cells after NMDA+SAG (treated 82.2±21.1 versus control 155±30.2, n=5; P=0.002). Consistent with this finding, we found that level of Ptch1, Gli1 and Gli2 were decreased in NMDA-damaged retinas treated with U0126 (Fig. 3I), suggesting that MAPK signaling is required to enable Hh signaling during the reprogramming of Müller glia into MGPCs.
Fig. 3.

Activation of Hh signaling following retinal injury stimulates the formation of proliferating MGPCs. Eyes were injected with 60 nmol NMDA, followed by vehicle (control), rhShh or SAG (treated) 4 and 24 h later and BrdU at 48 h; retinas were harvested 24 h after injection of BrdU. (A-D) Sections of the retinas were immunolabeled for Sox2 (red), BrdU (green) and PCNA (magenta; A,B), BrdU (green) and Nkx2.2 (red; C), or BrdU (green) and CD45 (red; D). Arrows indicate proliferating MGPCs, small double arrows indicate proliferating microglia, and arrowheads indicate proliferating NIRG cells. Scale bars: 50 µm (in B for A,B; in D for C,D). GCL, ganglion cell layer; INL, inner nuclear layer; IPL, inner plexiform layer; ONL, outer nuclear layer. (E-H) Histograms illustrating the mean number (±s.d.) of proliferating MGPCs (BrdU/Sox2+ Nkx2.2−) in rhShh-treated (E), rhShh±DAPT-treated (F), SAG-treated (G) and SAG±U0126-treated (H) central and peripheral retinas (±s.d.; n≥6). Significance of difference (*P<0.05, **P<0.01, ***P<0.005) was determined using a two-tailed t-test. (I) qRT-PCR was used to measure relative levels of Ptch1, Gli1 and Gli2 in retinas at 2 days after treatment with NDMA±U0126. (J) qRT-PCR was used to measure relative levels of Ptch1, Gli1, Gli2, Gli3, Pax2, Hes1, Hes5, Notch1, Ascl1a, Lin28, Yap, c3aR and c3 in retinas at 3 days after treatment with NMDA±rhShh. The mean (±s.d.; n≥4) percentage change in mRNA levels was determined, and the significance of difference (*P<0.05) between NMDA/vehicle- and NMDA/U0126- or rhShh-treated samples was determined using a Mann–Whitney U-test. ns, not significant.
To understand better how rhShh stimulates the formation of MGPCs in damaged retinas, we used qRT-PCR to assess changes in expression of components of the Hh pathway and genes that are known to be upregulated in MGPCs. In NMDA-damaged retinas treated with rhShh, we found significant increases in mRNAs of components of the Hh pathway (Fig. 3J). We found that injections of rhShh following NMDA treatment significantly increased levels of Ptch1, Gli1, Gli2 and Gli3 (Fig. 3J). In addition, rhShh increased levels of transcription factors that are associated with progenitors, including Pax2, Hes1, Hes5 and Yap, whereas levels of Notch1, Ascl1a and Lin28 were unaffected (Fig. 3J).
To investigate further how activation of Hh signaling in damaged retinas stimulates the proliferation of MGPCs, we probed for expression of components of the complement system. In embryonic chick, the complement peptide C3a and the C3a Receptor (C3aR) stimulate retinal regeneration during early stages of development (Haynes et al., 2013). In post-hatch chicks, levels of c3 and c3aR are elevated in NMDA-damaged retinas, but are significantly reduced when the microglia were ablated; in the latter case, MGPCs fail to form (Fischer et al., 2014). We found that levels of c3 were not affected by rhShh, whereas levels of c3aR were significantly increased (Fig. 3J).
Levels of cell death and damage can impact the formation of MGPCs (Fischer and Reh, 2001; Fischer et al., 2004b). Accordingly, we tested whether rhShh influenced cell death when applied after NMDA. The terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) assay revealed no difference in the numbers of dying cells in retinas treated with rhShh compared with numbers of dying cells in retinas treated with vehicle (treated 46.3±11.1 versus control 43.1±11.6, n=6; P=0.64).
Inhibition of Hh signaling reduces the proliferation of MGPCs
We next tested whether KAAD-cyclopamine, which directly binds to and inhibits Smo, influenced the formation of MGPCs. KAAD-cyclopamine significantly reduced numbers of proliferating MGPCs in retinas damaged by a high dose of NMDA (Fig. 4A-C). This inhibition of proliferation was specific to MGPCs, as the proliferation of CD45+ microglia and Nkx2.2+ NIRG cells was unaffected by KAAD-cyclopamine (Fig. 4C). Inhibition of Hh signaling with KAAD-cyclopamine after NMDA resulted in significant decreases in retinal levels of Ptch1 and Gli1, whereas levels of Hes1 were unaffected (Fig. 4D). In older animals (≥P27), we found that numbers of proliferating MGPCs were significantly decreased by KAAD-cyclopamine following NMDA treatment (supplementary material Fig. S2), suggesting that Hh signaling retains the capacity to stimulate Müller glia to become proliferating MGPCs in older damaged retinas.
Fig. 4.

Inhibition of Hh signaling suppresses the formation of MGPCs. Eyes were injected with 500 nmol NMDA, followed by injections of vehicle (control) or KAAD-cyclopamine, GANT58, GANT61 or anti-Shh (treated) at 4 and 24 h later, and BrdU at 48 h later. Retinas were harvested 24 h after the last injection of BrdU. (A,B) Retinal sections were immunolabeled for Sox2 (red), BrdU (green) and PCNA (magenta; A,B). Arrows indicate proliferating MGPCs and small double arrows indicate proliferating NIRG cells. GCL, ganglion cell layer; INL, inner nuclear layer; IPL, inner plexiform layer; ONL, outer nuclear layer. Scale bar: 50 µm (in B for A,B). (C,E-G) Histograms illustrating the mean number (±s.d.; n≥6) of proliferating MGPCs (BrdU/Sox2+ Nkx2.2−) in central and peripheral retina, NIRG cells (BrdU/Sox2/Nkx2.2+) and microglia (BrdU/CD45+). (G) The effects of 5E1 monoclonal were relatively small and significant differences were found only when accounting for inter-individual differences (treated – control). Significance of difference (*P<0.05, **P<0.01) was determined using a two-tailed t-test. (D) qRT-PCR for Ptch1, Gli1 and Hes1 in retinas at 3 days after treatment with NMDA±KAAD-cyclopamine. The mean (±s.d.; n≥4) percentage change in mRNA levels was determined and the significance of difference (*P<0.05) was determined using a Mann–Whitney U-test. ns, not significant.
Cyclopamine can have off-target effects (Yauch et al., 2008) by acting independently of Smo function and activating neutral sphingomyelinase 2 and ceramide production (Ponnusamy et al., 2010). Accordingly, we tested whether inhibition of Hh signaling with drugs that target different components of the Hh pathway influences the formation of MGPCs. We inhibited Gli proteins with the small molecule inhibitors GANT58, which interferes with Gli1-DNA binding, and GANT61, which interferes with Gli1/2-DNA binding (Lauth et al., 2007). Similar to treatment with KAAD-cyclopamine, GANT58 or GANT61 suppressed the formation of proliferating MGPCs in NMDA-damaged retinas (Fig. 4E,F), whereas the proliferation of microglia and NIRG cells was not affected (not shown). To block Shh ligand, we injected the 5E1 monoclonal antibody into eyes with damaged retinas. Application of the 5E1 antibody after NMDA treatment caused a significant decrease in the number of proliferating MPGCs (Fig. 4G), whereas the proliferation of microglia and NIRG cells was unaffected (not shown). The decrease in proliferation of MGPCs with 5E1 treatment was not as pronounced as that observed with KAAD-cyclopamine or GANT treatment, and only became statistically significant when accounting for inter-individual variability (Fig. 4G). TUNEL was performed 3 days after NMDA treatment, 24 h after the last dose of BrdU±Hh inhibitor, to assess whether decreases in cell death might underlie decreases in proliferation of MGPCs. We found that the Hh inhibitors did not influence cell death in NMDA-damaged retinas (KAAD-cyclopamine: control 49.25±34.2 versus treated 64.5±31.8, P=0.45; GANT58: control 39.0±40.1 versus treated 71±36.4, P=0.27).
The Hh pathway synergizes with FGF/MAPK signaling to influence MGPCs
Previous work in regeneration of prospective retina in the embryonic chick has implicated a requirement and interdependence of Hh signaling and FGF/MAPK signaling (Spence et al., 2007b, 2004). How these pathways interact in the context of MGPCs remains to be determined. Recently, it has been shown that sustained FGF/MAPK signaling is sufficient to stimulate the formation of MGPCs in the absence of damage (Fischer et al., 2014). Furthermore, FGF2 is known to activate and upregulate components of the Notch pathway, including Notch1, Dll4, Jagged, Hes5, Hey1 and Hey2 (Ghai et al., 2010), and activation of this pathway is required for the formation of MGPCs (Ghai et al., 2010; Hayes et al., 2007). We sought to investigate whether Hh signaling is activated by FGF/MAPK signaling and whether the Hh pathway contributes to the formation of MGPCs in FGF2-treated retinas. We began by probing for changes in expression of components of the Hh pathway in response to Smo agonist (SAG), FGF2 or IGF1. Consistent with findings that intraocular injections of SAG fail to stimulate the formation of MGPCs in undamaged retinas, treatment with SAG failed to increase retinal levels of Smo, Ptch1 or Gli2, whereas levels of Gli1 were increased (Fig. 5A). We found that FGF2 treatment, which selectively activates MAPK signaling in Müller glia (Fischer et al., 2009a), upregulated levels of Smo and Gli2, but not Ptch1 or Gli1 (Fig. 5A). By comparison, we detected no significant increases in components of the Hh pathway in response to IGF1 (data not shown).
Fig. 5.

Hh signaling influences the formation of proliferating MGPCs in FGF2-treated retinas in the absence of damage. Eyes were treated with different regimens involving FGF2/vehicle (control), FGF2/SAG (treated) or FGF2/KAAD-cyclopamine (treated). The injection paradigms for each data set are shown. (A) qRT-PCR was used to measure retinal levels of components of the Hh pathway. (A) Histograms illustrate the mean (±s.d.; n≥4) percent change in levels for Smo, Ptch1, Gli1 and Gli2 in retinas treated with SAG or FGF2. Significance of difference (*P<0.05) was determined using a Mann–Whitney U-test. (B-E) Retinal sections were immunolabeled for BrdU (green) and Sox2 (red; B,D), or cFos (green) and Sox2 (red; C,E). Sox2 (red; C,E) is included as a partial-field overlay to indicate nuclear colocalization with cFos (green). The histograms in B and D illustrate the mean number (±s.d.; n=6) of proliferating MGPCs (BrdU/Sox2+ Nkx2.2−) in peripheral retina. The histograms in C and E illustrate the mean (±s.d.; n=6) density sum for cFos immunofluorescence in Sox2+ Müller glia. Arrows indicate the nuclei of Müller glia and/or MGPCs. GCL, ganglion cell layer; INL, inner nuclear layer; IPL, inner plexiform layer; ONL, outer nuclear layer. Scale bars: 50 µm. Significance of difference (*P<0.05, **P<0.01) was determined using a two-tailed t-test. ns, not significant.
We found that three consecutive daily injections of the combination of FGF2 and SAG stimulated increased numbers of proliferating MGPCs compared with injection of FGF2 alone (Fig. 5B). To understand better the mechanism of the synergistic effects of FGF2 and SAG we assayed for cell-level changes in Müller glia following an acute dose of these factors. FGF2 is known to stimulate MAPK signaling in Müller glia (Fischer et al., 2009a). Thus, we probed for changes in expression levels of the MAPK effectors pERK, p38 MAPK, pCREB, Egr1 and cFos. Müller glia treated with FGF2 and SAG had elevated levels of the immediate early gene cFos compared with levels seen in Müller glia treated with FGF2 alone (Fig. 5C), whereas levels of pERK (supplementary material Fig. S3), Egr1, p38 MAPK, pS6 or pCREB (data not shown) in Müller glia were unaffected.
Because Hh agonist stimulated the formation of MGPCs in FGF2-treated retinas whereas Hh agonist alone had no effect, we tested whether Hh signaling was part of a signaling network activated by FGF2. Four consecutive daily doses of FGF2 are known to stimulate the formation of MGPCs in the absence of damage (Fischer et al., 2014). Co-application of KAAD-cyclopamine with FGF2 significantly reduced numbers of proliferating MGPCs compared with numbers seen in retinas treated with FGF2 alone (Fig. 5D). To understand better how KAAD-cyclopamine attenuates the formation of MGPCs, we investigated cell-level expression of signaling components in Müller glia following FGF/KAAD treatment. One day after two consecutive daily injections of FGF2 all of the Sox2+ Müller glia (320 of 320 cells, n=4) were positive for cFos (Fig. 5E). KAAD-cyclopamine significantly reduced FGF-induced cFos in Müller glia (Fig. 5D). In contrast to the embryonic chick (Spence et al., 2007a), we observed no differences between FGF-induced pERK in Müller glia following KAAD-cyclopamine treatment (supplementary material Fig. S3). Furthermore, we observed no KAAD-cyclopamine-induced differences in p38 MAPK, Egr1 and pCREB (not shown). Collectively, these data suggest that: (1) Hh signaling amplifies the ability of FGF2/MAPK signaling to stimulate the formation of MGPCs; (2) FGF2/MAPK signaling renders Müller glia receptive to Hh signaling; and (3) inhibition of Hh signaling disrupts FGF2/MAPK-mediated formation of MGPCs downstream of MAPK signaling, possibly at the level of cFos expression.
Hedgehog and IGF1 signaling
It has recently been shown that Shh and PI3K/mTOR pathways synergize to promote proliferation of human glioblastoma cells (Gruber Filbin et al., 2013). Accordingly, we tested whether co-activation of PI3K/mTor and Hh pathways stimulates proliferation of MGPCs in the uninjured retina. We applied four consecutive daily doses of SAG, IGF1, or SAG and IGF1. There was no nuclear migration of Müller glia in retinas treated with SAG alone, whereas there was some nuclear migration in retinas treated with IGF alone (Fig. 6A,B), consistent with a previous report (Fischer et al., 2010). By comparison, treatment with SAG+IGF1 caused widespread migration of Müller glia nuclei (Fig. 6A,B). The nuclei that migrated were ovoid shape in retinas treated with IGF1+SAG, compared with the more fusiform nuclei of Müller glia treated with SAG or IGF1, which did not undergo widespread nuclear migration (Fig. 6A,B). Proliferating MGPCs are known to undergo nuclear migration, reminiscent of proliferating embryonic retinal progenitors (Fischer et al., 2002b; Fischer and Reh, 2001). Despite widespread nuclear migration, Müller glia failed to incorporate BrdU or express significant levels of PCNA, GFAP or vimentin (not shown). Few BrdU+ MGPCs were found in far peripheral regions of the retina (Fig. 6C,D). Although numbers of Sox2+ Müller glia appeared to be increased in retinas treated with IGF1+SAG, cell counts revealed no significant difference in total numbers of Müller glia in retinas treated with IGF1 versus IGF1+SAG (Fig. 6E), consistent with the notion that treatment with IGF1+SAG fails to stimulate the formation of proliferating MGPCs.
Fig. 6.

The combination of IGF1 and Hh agonist (SAG) stimulates nuclear migration of Müller glia and proliferation of CMZ progenitors, but fails to stimulate the formation of MGPCs. (A-G) Retinal sections were immunolabeled for Sox2 (red; A-D) and BrdU (green; C,D,G). Arrows indicate the nuclei of Müller glia/MGPCs, and small double arrows indicate BrdU/Sox2+ NIRG cells. GCL, ganglion cell layer; INL, inner nuclear layer; IPL, inner plexiform layer; ONL, outer nuclear layer. (E) Mean (±s.d.; n≥8) number of Sox2+ Müller glia in the INL of retinas treated with IGF1 or IGF1+SAG. (F) Mean (±s.d.) number of BrdU+ cells within the temporal CMZ treated with saline, IGF1, SAG or IGF1+SAG. Significance of difference (P<0.001) among the treatment groups was determined using one-way ANOVA. Significance of difference (*P<0.05) between treatment groups was determined using a two-tailed t-test with Bonferroni's correction. ns, not significant. Scale bars: 50 µm (in D for A,C,D; in G for G alone).
To examine further the effects of IGF1 and SAG on proliferation, we tested whether the proliferation of progenitors in the circumferential marginal zone (CMZ) was affected. Intraocular injections of IGF1 or SAG stimulated the CMZ progenitors and the addition of new cells to the far peripheral edge of the retina by stimulating the proliferation of retinal progenitors (Fig. 6F,G), consistent with previous reports (Fischer and Reh, 2000; Moshiri et al., 2005). By comparison, the combination of IGF1 and SAG further stimulated the proliferation of retinal progenitors in the CMZ (Fig. 6F,G). These findings indicate that the doses of IGF1 and SAG were sufficient to stimulate the proliferation of progenitors that were capable of responding, unlike the Müller glia.
Most, if not all, proliferating MGPCs undergo nuclear migration that is accompanied by proliferation (Fischer and Reh, 2001). Thus, we investigated whether delaminated Müller glia de-differentiated to upregulate Pax6 in response to IGF1 and SAG. Müller glia express little or no Pax6 (Fischer et al., 2014), whereas MGPCs upregulate a number of different transcription factors, including Pax6, that are characteristic of retinal progenitors [reviewed by Gallina et al. (2014a); Fischer et al. (2010)]. In Müller glia treated with four consecutive daily doses of SAG, levels of Pax6 remain low and nuclei remain laminated in the middle of the INL (Fig. 7A), consistent with the notion that Hh signaling does not influence Müller glia. By comparison, nuclei migrated away from the middle of the INL and levels of Pax6 remained relatively low in Müller glia treated with four consecutive daily doses of SAG and IGF1 (Fig. 7A). Levels of Pax6 appeared elevated with relatively little nuclear migration in Müller glia treated with three consecutive daily doses of FGF2 (Fig. 7A), a treatment regime in which relatively few MGPCs proliferate (see Fig. 5). Levels of Pax6 were increased most and nuclear migration was widespread in Müller glia treated with three doses of FGF2+SAG or four doses of FGF2 alone (Fig. 7A), coincident with significant numbers of proliferating MGPCs (Fig. 5; Fischer et al., 2014). By comparison, levels of Pax6 were reduced and nuclear migration remained prominent in Müller glia treated with four doses of FGF2+KAAD-cyclopmine (Fig. 7A), suggesting that inhibition of Hh signaling might, in part, suppress the proliferation of MGPCs by inhibiting Pax6 expression.
Fig. 7.

Müller glia and MGPCs express Pax6 and Klf4 in response to treatment with different growth factors and/or Hh agonists/antagonists. (A,B) Retinas were treated with consecutive daily doses of saline ×4, SAG ×4, SAG and IGF1 ×4, FGF2 ×3, FGF2 and SAG ×3, FGF2 ×4 or FGF2 and KAAD-cyclopamine ×4. Retinas were immunolabeled for Pax6 (green; A) and Sox9 (red; A), or Klf4 (green; B) and TopAP (red; B). Arrows indicate the nuclei of Müller glia and/or MGPCs. Images were obtained by confocal microscopy, projecting four optical sections; therefore, there was some overlap of adjacent amacrine and glial nuclei through the z axis, indicated by small double arrows. GCL, ganglion cell layer; INL, inner nuclear layer; IPL, inner plexiform layer; ONL, outer nuclear layer. Scale bars: 50 µm.
To understand better how different combinations of growth factors influence the de-differentiation of Müller glia into progenitor cells, we probed for the expression of Klf4. Klf4 is one of the key transcription factors, when combined with Sox2, Myc and Oct4, that is capable of reprogramming cells into iPSCs (Takahashi and Yamanaka, 2006). We found that Klf4 is not expressed at detectable levels in normal Müller glia (Fig. 7B). By comparison, treatment with three consecutive daily intraocular injections of FGF2, IGF1 or the combination of IGF1+SAG induced robust expression of Klf4 in Müller glia (Fig. 7B). The upregulation of Klf4 in Müller glia appeared to be uniform across central and peripheral regions of retinas treated with FGF2, IGF1 or IGF1+SAG (data not shown). In central regions of retinas treated with FGF2+KAAD-cyclopamine, levels of Klf4 in Müller glia appeared to be diminished, and the nuclei remained laminated near the middle of the INL (Fig. 7B). By comparison, in peripheral regions of retinas treated with FGF2+KAAD-cyclopamine, levels of Klf4 in the Müller glia appeared to be reduced compared with those seen in glia treated with FGF2 alone, and the nuclei migrated away from the middle of the INL (Fig. 7B).
DISCUSSION
Collectively, our findings indicate that the Hh pathway plays an important role in the signaling network that underlies the formation of MGPCs. We found that components of the Hh signaling pathway are present in normal retina. It is possible that Hh signaling remains active in mature glia in order to maintain homeostasis (Alvarez et al., 2011). Deletion of Smo in adult astrocytes results in reactive gliosis, suggesting that baseline Hh signaling maintains homeostasis or glial quiescence (Garcia et al., 2010). In response to NMDA-induced damage, there was a rapid and sustained increase in retinal levels of Hh signaling components. Similarly, injury to the cerebral cortex induces Hh signaling (Amankulor et al., 2009; Sirko et al., 2013). Our data suggests that in NMDA-treated retinas, neuron-to-glia signaling mechanisms exists to direct Shh from ganglion cells to Müller glia and/or MGPCs. Re-distribution of Shh from the axons of ganglion cells to retinal cells following damage is similar to the re-distribution of Shh observed in zebrafish retina in response to injury (Sherpa et al., 2014). Additionally, our data are consistent with other reports on the nervous system demonstrating that neurons release Hh, whereas glia are receptive to Hh (Garcia et al., 2010; Sirko et al., 2013; Traiffort et al., 1999).
Although Hh agonists failed to influence Müller glia in normal retinas, Hh agonists stimulated the proliferation of MGPCs in damaged retinas and in retinas treated with FGF2 in the absence of damage. Our findings suggest that damage or FGF2 treatment render Müller glia responsive to Hh signaling and, thereafter, activation of Hh signaling stimulates the proliferation of MGPCs. We find that Hh signaling in Müller glia is downstream of MAPK in retinas damaged by NMDA: inhibition of MAPK signaling downregulated components of the Hh pathway and inhibited the ability of Hh agonists to stimulate the proliferation of MGPCs. In undamaged retinas, by comparison, activation of FGF2/MAPK signaling in Müller glia upregulated components of the Hh pathway and enabled Hh agonists/antagonists to influence the formation of MGPCs. Our findings suggest that rhShh activates a proliferative program through Gli effectors and/or Notch signaling and the Notch effectors Hes1 and Hes5 (Fig. 8). Additionally, inhibition of Notch signaling attenuated the ability of rhShh to stimulate the proliferation of MGPCs. Hh signaling is known to stimulate proliferation of neuronal precursors via activation of D-type cyclins, and Gli2 is able to regulate these cyclins directly (Kenney and Rowitch, 2000; Mill et al., 2003). In addition, Hh signaling is known to stimulate the expression of Hes1 and increase the proliferation of retinal progenitor cells (Wall et al., 2009) and subventricular zone neural stem cells (Ferent et al., 2014). We did not observe upregulation of Ascl1, a factor known to stimulate the formation of MGPCs in zebrafish and mouse retinas (Pollak et al., 2013; Ramachandran et al., 2010). It is possible that we failed to detect increases in Ascl1a owing to the time point of our analysis; following NMDA damage Ascl1a upregulation is maximal at 4 days after damage (Hayes et al., 2007), whereas our analysis took place 3 days after damage. We observed increases in Pax2 after treatment with rhShh in the damaged retina. During development, the expression of Pax2 is stimulated in optic nerve glia by Shh (Dakubo et al., 2003; Take-uchi et al., 2003; Wallace and Raff, 1999). By comparison, Pax2 is expressed by Müller glia in central regions of the chick and zebrafish retina (Boije et al., 2010; Stanke et al., 2010). Interestingly, we observed Hh-induced upregulation of C3aR, the receptor recently discovered to mediate the regeneration of embryonic chick retina in response to complement factor C3 (Haynes et al., 2013). These data suggest that C3aR-mediated signaling could play a role in Hh-stimulated regeneration of mature retina.
Fig. 8.

Schematic illustrating the induction of MGPCs by NMDA and FGF2 treatments, activation of MAPK signaling, subsequent activation of Hh and Notch signaling, and sites of action of different compounds used in the study. Agonists shown in green; antagonists in red.
We found that inhibition of Hh signaling by targeting the Hh ligand, receptor or transcriptional effectors significantly reduced numbers of proliferating MGPCs in damage retinas. Blockade of Hh signaling pharmacologically in retinal progenitors is known to suppress progression through the cell cycle (Locker et al., 2006). Similarly, the FGF2-mediated regeneration of prospective retina in the embryonic chick fails when Hh signaling is inhibited (Spence et al., 2007a). In mature Müller glia in vivo, FGF2 selectively activates MAPK signaling (Fischer et al., 2009a), and stimulates the formation of proliferating MGPCs in the absence of damage (Fischer et al., 2014). Collectively, these data suggest that FGF2/MAPK signaling ‘kick-starts’ a network of interdependent pathways to stimulate the formation of proliferating MGPCs. These pathways are known to include upregulation of Notch signaling (Ghai et al., 2010), upregulation of Hh signaling (current study) and downregulation of glucocorticoid signaling (Gallina et al., 2014b). Our current findings suggest that Notch signaling is downstream of both MAPK and Hh pathways (Fig. 8).
In the chick eye, CMZ progenitors normally respond to Hh signaling (Fig. 6; Moshiri et al., 2005), whereas Müller glia must be ‘primed’ by damage or FGF2/MAPK signaling to become responsive to Hh. Our data indicate that IGF1 is not sufficient to stimulate Müller to respond to the mitogenic effects of Hh agonists. In response to the combination of IGF1 and Hh agonist, we observed widespread nuclear migration of Müller glia nuclei and changes in nuclear morphology, but no proliferation. These data suggest that nuclear migration of Müller glia can occur independently of cell cycle re-entry. By comparison, proliferation of MGPCs may require nuclear migration away from the center of the INL. For example, the vast majority of pHisH3+ mitotic figures for MGPCs are observed in the distal INL or ONL (Fischer and Reh, 2001). In retinal development, nuclear migration of progenitor cells is coordinated with cell cycle regulation. However, nuclear migration can be uncoupled from cell cycle re-entry (reviewed by Baye and Link, 2008).
Stimulation of Müller glia with IGF1 and Hh agonist is not sufficient to upregulate Pax6 to levels seen in proliferating MGPCs. In normal Müller glia, Pax6 is expressed at very low levels but is rapidly induced during the formation of proliferating MGPCs (Fischer and Reh, 2001). Upregulation of Pax6 is symptomatic of Müller glial de-differentiation. In addition, Pax6 plays a key role in the proliferation and multipotency of retinal progenitor cells in development (reviewed by Ashery-Padan and Gruss, 2001) and is required for Müller glia-mediated retinal regeneration in zebrafish (Thummel et al., 2010). Consistent with the notion that upregulation of Pax6 is an important step in the formation of MGPCs, inhibition of Hh signaling suppressed the formation of MGPCs and prevented the upregulation Pax6, in addition to Klf4, in Müller glia treated with FGF2.
Our data suggest that in response to IGF1 and Hh agonist Müller glia make initial steps to becoming ‘progenitor-like’ by migrating away from the INL and upregulating low levels of Pax6 and Klf4, but fail to fully de-differentiate and re-enter the cell cycle as MGPCs. Alternatively, Müller glia that respond to IGF1 and Hh agonist may migrate as part of a gliotic response. Inhibition of Hh signaling in FGF2-treated retinas suppressed Pax6 expression in Müller glia, but does not suppress nuclear migration, suggesting that Hh signaling and FGF2/MAPK signaling synergize to stimulate Pax6 expression during the transition in phenotype from Müller glia to MGPC. Müller glia are known to undergo nuclear migration in response to various retinopathies [reviewed by Bringmann et al. (2006); Gruber Filbin et al. (2013)]. However, we failed to detect markers of glial reactivity, such as elevated vimentin or GFAP, in retinas treated with IGF1 and Hh agonist.
We cannot exclude the possibility that the Hh agonists and antagonists act through amacrine or ganglion cells to influence the proliferation of MGPCs in damaged retinas. We find that Ptch1 is expressed by neurons in addition to Müller glia and/or MPGCs in damaged retinas. Thus, it is possible that Hh signaling through inner retinal neurons impacts the formation of proliferating MGPCs. Consistent with this hypothesis, a recent study in zebrafish retina demonstrated that regeneration from MGPCs is delayed with a partial loss-of-function mutation in Shh, which is produced by inner retinal neurons (Sherpa et al., 2014). In FGF2-treated retinas in the absence of damage, where Müller glia are known to be selectively activated (Fischer et al., 2009a), we find that activation or inhibition of Hh signaling influences the expression of cFos and the proliferation of MGPCs. It remains uncertain whether the influences of Hh signaling on FGF2-induced MGPCs are manifested directly through the Müller glia or through inner retinal neurons.
Conclusions
We conclude that Hh signaling influences the proliferation of MGPCs in NMDA-damaged retinas and in FGF2-treated retinas in the absence of damage. In damaged retinas, Shh signaling is dynamically regulated and is correlated with a redistribution of Shh from ganglion cell axons to MGPCs. In addition, we find that FGF2/MAPK signaling stimulates Müller glia to become responsive to Hh signaling, and that Hh signaling contributes to the network of signaling pathways and to Pax6 expression to stimulate the de-differentiation and proliferation of MGPCs. We find that the combined activation of IGF1 and Hh signaling is sufficient to stimulate some aspects of de-differentiation, such as nuclear migration and induction of low levels of Pax6, but fails to promote fully the de-differentiation of Müller glia into proliferating MGPCs.
MATERIALS AND METHODS
Animals
The use of animals in these experiments was in accordance with the guidelines established by the National Institutes of Health and the Ohio State University. Newly hatched male and female wild-type leghorn chickens (Gallus gallus domesticus) were obtained from Meyer Hatchery (Polk, OH, USA). Chicks were kept on a cycle of 12 h light, 12 h dark (lights on at 08:00 h). Chicks were housed in a stainless steel brooder at ∼25°C and received water and Purina chick starter ad libitum. All of the chicks used in these studies were between 6 and 28 days of age.
Intraocular injections
Chickens were anesthetized and intraocular injections were performed as described previously (Fischer et al., 2008, 2009a). Injected compounds included NMDA (6.4 or 154 µg/dose; Sigma-Aldrich), FGF2 (250 ng/dose; Sigma-Aldrich), IGF1 (400 ng/dose; R&D Systems), rhShh (500 ng/dose; R&D Systems), KAAD-cyclopamine (1 µg/dose; Selleck Chemicals; CAS 306387-90-6), 5E1 anti-Shh (20 µl/dose; Developmental Studies Hybridoma Bank), Gli antagonist GANT58 (1 µg/dose; Selleck Chemicals), Gli antagonist GANT61 (1 µg/dose; Tocris Bioscience), smoothened agonist (SAG) (500 ng/dose; EMD Millipore; CAS 364590-63-6), DAPT {N-[N-(3,5-difluorophenacetyl-L-alanyl)]-S-phenylglycine t-butyl ester; 865 ng/dose; Sigma-Aldrich}, UO126 (1.7 μg/dose; Calbiochem) and 5-bromo-2′-deoxyuridine (BrdU; 2 µg/dose; Sigma-Aldrich). Hydrophobic compounds were diluted and injected in 30% DMSO in saline. Injection paradigms are included in the figures and legends.
PCR
Tissue dissections, RNA isolation, reverse transcriptase reactions and PCR reactions were performed as described previously (Fischer et al., 2004a, 2010; Ghai et al., 2010). PCR primers were designed by using the Primer-BLAST primer design tool at NCBI (http://www.ncbi.nlm.nih.gov/tools/primer-blast/). Primer sequences are included in supplementary material Table S1. qPCR reactions were performed using SYBR Green Master Mix and StepOnePlus Real-Time system (Applied BioSystems). Samples were run in triplicate on a minimum of four individuals. Ct values obtained from real-time PCR were normalized to GAPDH for each sample and the fold change between control and treated samples was determined using the 2−ΔΔCt method [=fold change 2(−ΔΔCt)] and represented as a percentage change from the control, which was assigned a value of 100. Significance of difference for percent change was determined by using a non-parametric Mann–Whitney U-test.
TUNEL
To identify dying cells that contained fragmented DNA, the TUNEL method was used. We used an In Situ Cell Death Kit (TMR red; Roche Applied Science), as per the manufacturer's instructions.
In situ hybridization
The plasmid carrying a cDNA fragment of Ptch1 was provided by Dr Clifford Tabin (Department of Genetics, Harvard University, Boston, USA). The riboprobe to Ptch1 covered nucleotides 900 to 3.4 kb. Riboprobe to Shh was generated from PCR product using primers that included RNA polymerase initiation sites, as described elsewhere (Stanke et al., 2010). The riboprobe to Shh covered nucleotides 332 to 1415. Digoxigenin-labeled riboprobes were synthesized using the DIG RNA Labeling Kit (Roche), and stored at −80°C until use. Eyes were dissected, fixed, sectioned and processed for in situ hybridization as described previously (Fischer et al., 2002a, 2004a; Ghai et al., 2009, 2010).
Fixation, sectioning and immunocytochemistry
Tissues were fixed, sectioned and immunolabeled as described previously (Fischer and Reh, 2000, 2002; Fischer et al., 1998). Primary and secondary antibodies are listed in supplementary material Table S2.
Photography, cell counts and statistics
Digital photomicroscopy was performed as described in previous studies (Fischer et al., 2008, 2010; Ghai et al., 2009, 2010). Central retina was defined as the region within a 3 mm radius of the posterior pole of the eye, and peripheral retina was defined as an annular region between 3 mm and 0.5 mm from the CMZ. The identity of BrdU-labeled cells was determined based on previous findings that all proliferating cells in the chick retina are Sox2/9+ Müller glia in the INL/ONL, Sox2/9/Nkx2.2+ NIRG cells in the IPL, GCL and NFL (the NIRG cells do not migrate distally into the retina), and CD45+ (Sox2/9−) microglia (Fischer et al., 2010; Zelinka et al., 2012). Sox2+ nuclei in the INL were identified as Müller glia based on their large size and fusiform shape, which was distinctly different from the Sox2+ nuclei of cholinergic amacrine cells, which are small and round (Fischer et al., 2010). None of the proliferating Sox2/Sox9-positive cells within peripheral regions of the retina was derived from CMZ progenitors; cells produced by CMZ progenitors do not migrate laterally into the retina (Fischer and Reh, 2003a). Similar to previous studies (Fischer et al., 2009a,b, 2010; Ghai et al., 2009), immunofluorescence was quantified by using ImagePro 6.2 (Media Cybernetics, Bethesda, MD, USA). The mean area, intensity and density sum was calculated for the pixels within threshold regions from at least five retinas for each experimental condition. GraphPad Prism 6 was used for statistical analyses.
Supplementary Material
Acknowledgements
The authors thank Donika Gallina and Chris Zelinka for comments on the manuscript; and Lilianna Suarez and Natalie Squires for technical assistance. The antibodies to transitin, Nkx2.2, Pax6, neurofilament, Shh and BrdU developed by Drs G. J. Cole, T. M. Jessell, A. Kawakami, J. Wood and S. J. Kaufman, respectively, were obtained from the Developmental Studies Hybridoma Bank, which was developed under the auspices of the NICHD and is maintained by the University of Iowa, Department of Biological Sciences, Iowa City, IA 52242, USA.
Footnotes
Competing interests
The authors declare no competing or financial interests.
Author contributions
L.T. designed and executed experiments, gathered data, constructed figures and contributed to writing the manuscript. A.J.F. designed and executed experiments, gathered data, constructed figures and contributed to writing the manuscript.
Funding
This work was supported by a grant from the National Institutes of Health, National Eye Institute [EY022030-03]. Deposited in PMC for release after 12 months.
Supplementary material
Supplementary material available online at http://dev.biologists.org/lookup/suppl/doi:10.1242/dev.121616/-/DC1
References
- Alvarez J. I., Dodelet-Devillers A., Kebir H., Ifergan I., Fabre P. J., Terouz S., Sabbagh M., Wosik K., Bourbonniere L. and Bernard M. et al. (2011). The Hedgehog pathway promotes blood-brain barrier integrity and CNS immune quiescence. Science 334, 1727-1731. 10.1126/science.1206936 [DOI] [PubMed] [Google Scholar]
- Amankulor N. M., Hambardzumyan D., Pyonteck S. M., Becher O. J., Joyce J. A. and Holland E. C. (2009). Sonic hedgehog pathway activation is induced by acute brain injury and regulated by injury-related inflammation. J. Neurosci. 29, 10299-10308. 10.1523/JNEUROSCI.2500-09.2009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ashery-Padan R. and Gruss P. (2001). Pax6 lights-up the way for eye development. Curr. Opin. Cell Biol. 13, 706-714. 10.1016/S0955-0674(00)00274-X [DOI] [PubMed] [Google Scholar]
- Bai C. B., Auerbach W., Lee J. S., Stephen D. and Joyner A. L. (2002). Gli2, but not Gli1, is required for initial Shh signaling and ectopic activation of the Shh pathway. Development 129, 4753-4761. [DOI] [PubMed] [Google Scholar]
- Baye L. M. and Link B. A. (2008). Nuclear migration during retinal development. Brain Res. 1192, 29-36. 10.1016/j.brainres.2007.05.021 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bernardos R. L., Barthel L. K., Meyers J. R. and Raymond P. A. (2007). Late-stage neuronal progenitors in the retina are radial Muller glia that function as retinal stem cells. J. Neurosci. 27, 7028-7040. 10.1523/JNEUROSCI.1624-07.2007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blackshaw S., Harpavat S., Trimarchi J., Cai L., Huang H., Kuo W. P., Weber G., Lee K., Fraioli R. E., Cho S.-H. et al. (2004). Genomic analysis of mouse retinal development. PLoS Biol. 2, e247 10.1371/journal.pbio.0020247 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boije H., Ring H., López-Gallardo M., Prada C. and Hallböök F. (2010). Pax2 is expressed in a subpopulation of Müller cells in the central chick retina. Dev. Dyn. 239, 1858-1866. 10.1002/dvdy.22309 [DOI] [PubMed] [Google Scholar]
- Bringmann A., Pannicke T., Grosche J., Francke M., Wiedemann P., Skatchkov S. N., Osborne N. N. and Reichenbach A. (2006). Müller cells in the healthy and diseased retina. Prog. Retin. Eye Res. 25, 397-424. 10.1016/j.preteyeres.2006.05.003 [DOI] [PubMed] [Google Scholar]
- Chen J. K., Taipale J., Young K. E., Maiti T. and Beachy P. A. (2002). Small molecule modulation of Smoothened activity. Proc. Natl. Acad. Sci. USA 99, 14071-14076. 10.1073/pnas.182542899 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Close J. L., Gumuscu B. and Reh T. A. (2005). Retinal neurons regulate proliferation of postnatal progenitors and Muller glia in the rat retina via TGF beta signaling. Development 132, 3015-3026. 10.1242/dev.01882 [DOI] [PubMed] [Google Scholar]
- Dakubo G. D., Wang Y. P., Mazerolle C., Campsall K., McMahon A. P. and Wallace V. A. (2003). Retinal ganglion cell-derived sonic hedgehog signaling is required for optic disc and stalk neuroepithelial cell development. Development 130, 2967-2980. 10.1242/dev.00515 [DOI] [PubMed] [Google Scholar]
- Fausett B. V. and Goldman D. (2006). A role for alpha1 tubulin-expressing Muller glia in regeneration of the injured zebrafish retina. J. Neurosci. 26, 6303-6313. 10.1523/JNEUROSCI.0332-06.2006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ferent J., Ruat M. and Traiffort E. (2014). [Symmetric or asymmetric division: sonic Hedgehog controls the fate of neural stem cells]. Med. Sci. 30, 705-708. 10.1051/medsci/20143006026 [DOI] [PubMed] [Google Scholar]
- Fischer A. J. (2005). Neural regeneration in the chick retina. Prog. Retin. Eye Res. 24, 161-182. 10.1016/j.preteyeres.2004.07.003 [DOI] [PubMed] [Google Scholar]
- Fischer A. J. and Bongini R. (2010). Turning Müller glia into neural progenitors in the retina. Mol. Neurobiol. 42, 199-209. 10.1007/s12035-010-8152-2 [DOI] [PubMed] [Google Scholar]
- Fischer A. J. and Reh T. A. (2000). Identification of a proliferating marginal zone of retinal progenitors in postnatal chickens. Dev. Biol. 220, 197-210. 10.1006/dbio.2000.9640 [DOI] [PubMed] [Google Scholar]
- Fischer A. J. and Reh T. A. (2001). Müller glia are a potential source of neural regeneration in the postnatal chicken retina. Nat. Neurosci. 4, 247-252. 10.1038/85090 [DOI] [PubMed] [Google Scholar]
- Fischer A. J. and Reh T. A. (2002). Exogenous growth factors stimulate the regeneration of ganglion cells in the chicken retina. Dev. Biol. 251, 367-379. 10.1006/dbio.2002.0813 [DOI] [PubMed] [Google Scholar]
- Fischer A. J. and Reh T. A. (2003a). Growth factors induce neurogenesis in the ciliary body. Dev. Biol. 259, 225-240. 10.1016/S0012-1606(03)00178-7 [DOI] [PubMed] [Google Scholar]
- Fischer A. J. and Reh T. A. (2003b). Potential of Müller glia to become neurogenic retinal progenitor cells. Glia 43, 70-76. 10.1002/glia.10218 [DOI] [PubMed] [Google Scholar]
- Fischer A. J., Pickett Seltner R. L., Poon J. and Stell W. K. (1998). Immunocytochemical characterization of quisqualic acid- and N-methyl-D-aspartate-induced excitotoxicity in the retina of chicks. J. Comp. Neurol. 393, 1-15. 10.1002/(SICI)1096-9861(19980330)393:1<1::AID-CNE1>3.0.CO;2-3 [DOI] [PubMed] [Google Scholar]
- Fischer A. J., Dierks B. D. and Reh T. A. (2002a). Exogenous growth factors induce the production of ganglion cells at the retinal margin. Development 129, 2283-2291. [DOI] [PubMed] [Google Scholar]
- Fischer A. J., McGuire C. R., Dierks B. D. and Reh T. A. (2002b). Insulin and fibroblast growth factor 2 activate a neurogenic program in Muller glia of the chicken retina. J. Neurosci. 22, 9387-9398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fischer A. J., Omar G., Eubanks J., McGuire C. R., Dierks B. D. and Reh T. A. (2004a). Different aspects of gliosis in retinal Muller glia can be induced by CNTF, insulin and FGF2 in the absence of damage. Mol. Vis. 10, 973-986. [PubMed] [Google Scholar]
- Fischer A. J., Schmidt M., Omar G. and Reh T. A. (2004b). BMP4 and CNTF are neuroprotective and suppress damage-induced proliferation of Müller glia in the retina. Mol. Cell. Neurosci. 27, 531-542. 10.1016/j.mcn.2004.08.007 [DOI] [PubMed] [Google Scholar]
- Fischer A. J., Ritchey E. R., Scott M. A. and Wynne A. (2008). Bullwhip neurons in the retina regulate the size and shape of the eye. Dev. Biol. 317, 196-212. 10.1016/j.ydbio.2008.02.023 [DOI] [PubMed] [Google Scholar]
- Fischer A. J., Scott M. A., Ritchey E. R. and Sherwood P. (2009a). Mitogen-activated protein kinase-signaling regulates the ability of Müller glia to proliferate and protect retinal neurons against excitotoxicity. Glia 57, 1538-1552. 10.1002/glia.20868 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fischer A. J., Scott M. A. and Tuten W. (2009b). Mitogen-activated protein kinase-signaling stimulates Müller glia to proliferate in acutely damaged chicken retina. Glia 57, 166-181. 10.1002/glia.20743 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fischer A. J., Scott M. A., Zelinka C. and Sherwood P. (2010). A novel type of glial cell in the retina is stimulated by insulin-like growth factor 1 and may exacerbate damage to neurons and Müller glia. Glia 58, 633-649. 10.1002/glia.20950 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fischer A. J., Zelinka C., Gallina D., Scott M. A. and Todd L. (2014). Reactive microglia and macrophage facilitate the formation of Müller glia-derived retinal progenitors. Glia 62, 1608-1628. 10.1002/glia.22703 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gallina D., Todd L. and Fischer A. J. (2014a). A comparative analysis of Müller glia-mediated regeneration in the vertebrate retina. Exp. Eye Res. 123, 121-130. 10.1016/j.exer.2013.06.019 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gallina D., Zelinka C. and Fischer A. J. (2014b). Glucocorticoid receptors in the retina, Muller glia and the formation of Muller glia-derived progenitors. Development 141, 3340-3351. 10.1242/dev.109835 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gao L. and Miller R. H. (2006). Specification of optic nerve oligodendrocyte precursors by retinal ganglion cell axons. J. Neurosci. 26, 7619-7628. 10.1523/JNEUROSCI.0855-06.2006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Garcia A. D. R., Petrova R., Eng L. and Joyner A. L. (2010). Sonic hedgehog regulates discrete populations of astrocytes in the adult mouse forebrain. J. Neurosci. 30, 13597-13608. 10.1523/JNEUROSCI.0830-10.2010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ghai K., Zelinka C. and Fischer A. J. (2009). Serotonin released from amacrine neurons is scavenged and degraded in bipolar neurons in the retina. J. Neurochem. 111, 1-14. 10.1111/j.1471-4159.2009.06270.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ghai K., Zelinka C. and Fischer A. J. (2010). Notch signaling influences neuroprotective and proliferative properties of mature Muller glia. J. Neurosci. 30, 3101-3112. 10.1523/JNEUROSCI.4919-09.2010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goodrich L. V., Johnson R. L., Milenkovic L., McMahon J. A. and Scott M. P. (1996). Conservation of the hedgehog/patched signaling pathway from flies to mice: induction of a mouse patched gene by Hedgehog. Genes Dev. 10, 301-312. 10.1101/gad.10.3.301 [DOI] [PubMed] [Google Scholar]
- Gruber Filbin M., Dabral S. K., Pazyra-Murphy M. F., Ramkissoon S., Kung A. L., Pak E., Chung J., Theisen M. A., Sun Y., Franchetti Y. et al. (2013). Coordinate activation of Shh and PI3K signaling in PTEN-deficient glioblastoma: new therapeutic opportunities. Nat. Med. 19, 1518-1523. 10.1038/nm.3328 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hayes S., Nelson B. R., Buckingham B. and Reh T. A. (2007). Notch signaling regulates regeneration in the avian retina. Dev. Biol. 312, 300-311. 10.1016/j.ydbio.2007.09.046 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haynes T., Luz-Madrigal A., Reis E. S., Echeverri Ruiz N. P., Grajales-Esquivel E., Tzekou A., Tsonis P. A., Lambris J. D. and Del Rio-Tsonis K. (2013). Complement anaphylatoxin C3a is a potent inducer of embryonic chick retina regeneration. Nat. Commun. 4, 2312 10.1038/ncomms3312 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hooper J. E. and Scott M. P. (2005). Communicating with Hedgehogs. Nat. Rev. Mol. Cell Biol. 6, 306-317. 10.1038/nrm1622 [DOI] [PubMed] [Google Scholar]
- Karl M. O., Hayes S., Nelson B. R., Tan K., Buckingham B. and Reh T. A. (2008). Stimulation of neural regeneration in the mouse retina. Proc. Natl. Acad. Sci. USA 105, 19508-19513. 10.1073/pnas.0807453105 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kenney A. M. and Rowitch D. H. (2000). Sonic hedgehog promotes G1 cyclin expression and sustained cell cycle progression in mammalian neuronal precursors. Mol. Cell. Biol. 20, 9055-9067. 10.1128/MCB.20.23.9055-9067.2000 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lauth M., Bergstrom A., Shimokawa T. and Toftgard R. (2007). Inhibition of GLI-mediated transcription and tumor cell growth by small-molecule antagonists. Proc. Natl. Acad. Sci. USA 104, 8455-8460. 10.1073/pnas.0609699104 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Levine E. M., Roelink H., Turner J. and Reh T. A. (1997). Sonic hedgehog promotes rod photoreceptor differentiation in mammalian retinal cells in vitro. J. Neurosci. 17, 6277-6288. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Litingtung Y. and Chiang C. (2000). Specification of ventral neuron types is mediated by an antagonistic interaction between Shh and Gli3. Nat. Neurosci. 3, 979-985. 10.1038/79916 [DOI] [PubMed] [Google Scholar]
- Locker M., Agathocleous M., Amato M. A., Parain K., Harris W. A. and Perron M. (2006). Hedgehog signaling and the retina: insights into the mechanisms controlling the proliferative properties of neural precursors. Genes Dev. 20, 3036-3048. 10.1101/gad.391106 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marigo V. and Tabin C. J. (1996). Regulation of patched by sonic hedgehog in the developing neural tube. Proc. Natl. Acad. Sci. USA 93, 9346-9351. 10.1073/pnas.93.18.9346 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marigo V., Johnson R. L., Vortkamp A. and Tabin C. J. (1996). Sonic hedgehog differentially regulates expression of GLI and GLI3 during limb development. Dev. Biol. 180, 273-283. 10.1006/dbio.1996.0300 [DOI] [PubMed] [Google Scholar]
- Mill P., Mo R., Fu H., Grachtchouk M., Kim P. C. W., Dlugosz A. A. and Hui C.-C. (2003). Sonic hedgehog-dependent activation of Gli2 is essential for embryonic hair follicle development. Genes Dev. 17, 282-294. 10.1101/gad.1038103 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moshiri A. and Reh T. A. (2004). Persistent progenitors at the retinal margin of ptc+/- mice. J. Neurosci. 24, 229-237. 10.1523/JNEUROSCI.2980-03.2004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moshiri A., McGuire C. R. and Reh T. A. (2005). Sonic hedgehog regulates proliferation of the retinal ciliary marginal zone in posthatch chicks. Dev. Dyn. 233, 66-75. 10.1002/dvdy.20299 [DOI] [PubMed] [Google Scholar]
- Nelson B. R., Ueki Y., Reardon S., Karl M. O., Georgi S., Hartman B. H., Lamba D. A. and Reh T. A. (2011). Genome-wide analysis of Müller glial differentiation reveals a requirement for Notch signaling in postmitotic cells to maintain the glial fate. PLoS ONE 6, e22817 10.1371/journal.pone.0022817 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ooto S., Akagi T., Kageyama R., Akita J., Mandai M., Honda Y. and Takahashi M. (2004). Potential for neural regeneration after neurotoxic injury in the adult mammalian retina. Proc. Natl. Acad. Sci. USA 101, 13654-13659. 10.1073/pnas.0402129101 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Perron M., Boy S., Amato M. A., Viczian A., Koebernick K., Pieler T. and Harris W. A. (2003). A novel function for Hedgehog signalling in retinal pigment epithelium differentiation. Development 130, 1565-1577. 10.1242/dev.00391 [DOI] [PubMed] [Google Scholar]
- Pollak J., Wilken M. S., Ueki Y., Cox K. E., Sullivan J. M., Taylor R. J., Levine E. M. and Reh T. A. (2013). Ascl1 reprograms mouse Muller glia into neurogenic retinal progenitors. Development 140, 2619-2631. 10.1242/dev.091355 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ponnusamy S., Meyers-Needham M., Senkal C. E., Saddoughi S. A., Sentelle D., Selvam S. P., Salas A. and Ogretmen B. (2010). Sphingolipids and cancer: ceramide and sphingosine-1-phosphate in the regulation of cell death and drug resistance. Future Oncol. 6, 1603-1624. 10.2217/fon.10.116 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ramachandran R., Fausett B. V. and Goldman D. (2010). Ascl1a regulates Müller glia dedifferentiation and retinal regeneration through a Lin-28-dependent, let-7 microRNA signalling pathway. Nat. Cell Biol. 12, 1101-1107. 10.1038/ncb2115 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reh T. A. and Fischer A. J. (2001). Stem cells in the vertebrate retina. Brain Behav. Evol. 58, 296-305. 10.1159/000057571 [DOI] [PubMed] [Google Scholar]
- Roesch K., Jadhav A. P., Trimarchi J. M., Stadler M. B., Roska B., Sun B. B. and Cepko C. L. (2008). The transcriptome of retinal Müller glial cells. J. Comp. Neurol. 509, 225-238. 10.1002/cne.21730 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roesch K., Stadler M. B. and Cepko C. L. (2012). Gene expression changes within Muller glial cells in retinitis pigmentosa. Mol. Vis. 18, 1197-1214. [PMC free article] [PubMed] [Google Scholar]
- Ruiz i Altaba A., Palma V. and Dahmane N. (2002). Hedgehog–Gli signalling and the growth of the brain. Nat. Rev. Neurosci. 3, 24-33. 10.1038/nrn704 [DOI] [PubMed] [Google Scholar]
- Sherpa T., Lankford T., McGinn T. E., Hunter S. S., Frey R. A., Sun C., Ryan M., Robison B. D. and Stenkamp D. L. (2014). Retinal regeneration is facilitated by the presence of surviving neurons. Dev. Neurobiol. 74, 851-876. 10.1002/dneu.22167 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sirko S., Behrendt G., Johansson P. A., Tripathi P., Costa M. R., Bek S., Heinrich C., Tiedt S., Colak D., Dichgans M. et al. (2013). Reactive glia in the injured brain acquire stem cell properties in response to sonic hedgehog. Cell Stem Cell 12, 426-439. 10.1016/j.stem.2013.01.019 [DOI] [PubMed] [Google Scholar]
- Spence J. R., Madhavan M., Ewing J. D., Jones D. K., Lehman B. M. and Del Rio-Tsonis K. (2004). The hedgehog pathway is a modulator of retina regeneration. Development 131, 4607-4621. 10.1242/dev.01298 [DOI] [PubMed] [Google Scholar]
- Spence J. R., Aycinena J.-C. and Del Rio-Tsonis K. (2007a). Fibroblast growth factor-hedgehog interdependence during retina regeneration. Dev. Dyn. 236, 1161-1174. 10.1002/dvdy.21115 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spence J. R., Madhavan M., Aycinena J. C. and Del Rio-Tsonis K. (2007b). Retina regeneration in the chick embryo is not induced by spontaneous Mitf downregulation but requires FGF/FGFR/MEK/Erk dependent upregulation of Pax6. Mol. Vis. 13, 57-65. [PMC free article] [PubMed] [Google Scholar]
- Stanke J., Moose H. E., El-Hodiri H. M. and Fischer A. J. (2010). Comparative study of Pax2 expression in glial cells in the retina and optic nerve of birds and mammals. J. Comp. Neurol. 518, 2316-2333. 10.1002/cne.22335 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Takahashi K. and Yamanaka S. (2006). Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 126, 663-676. 10.1016/j.cell.2006.07.024 [DOI] [PubMed] [Google Scholar]
- Take-uchi M., Clarke J. D. W. and Wilson S. W. (2003). Hedgehog signalling maintains the optic stalk-retinal interface through the regulation of Vax gene activity. Development 130, 955-968. 10.1242/dev.00305 [DOI] [PubMed] [Google Scholar]
- Thummel R., Enright J. M., Kassen S. C., Montgomery J. E., Bailey T. J. and Hyde D. R. (2010). Pax6a and Pax6b are required at different points in neuronal progenitor cell proliferation during zebrafish photoreceptor regeneration. Exp. Eye Res. 90, 572-582. 10.1016/j.exer.2010.02.001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Traiffort E., Charytoniuk D., Watroba L., Faure H., Sales N. and Ruat M. (1999). Discrete localizations of hedgehog signalling components in the developing and adult rat nervous system. Eur. J. Neurosci. 11, 3199-3214. 10.1046/j.1460-9568.1999.00777.x [DOI] [PubMed] [Google Scholar]
- Traiffort E., Moya K. L., Faure H., Hassig R. and Ruat M. (2001). High expression and anterograde axonal transport of aminoterminal sonic hedgehog in the adult hamster brain. Eur. J. Neurosci. 14, 839-850. 10.1046/j.0953-816x.2001.01708.x [DOI] [PubMed] [Google Scholar]
- Wall D. S., Mears A. J., McNeill B., Mazerolle C., Thurig S., Wang Y., Kageyama R. and Wallace V. A. (2009). Progenitor cell proliferation in the retina is dependent on Notch-independent Sonic hedgehog/Hes1 activity. J. Cell Biol. 184, 101-112. 10.1083/jcb.200805155 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wallace V. A. and Raff M. C. (1999). A role for Sonic hedgehog in axon-to-astrocyte signalling in the rodent optic nerve. Development 126, 2901-2909. [DOI] [PubMed] [Google Scholar]
- Wan J., Zheng H., Xiao H.-L., She Z.-J. and Zhou G.-M. (2007). Sonic hedgehog promotes stem-cell potential of Müller glia in the mammalian retina. Biochem. Biophys. Res. Commun. 363, 347-354. 10.1016/j.bbrc.2007.08.178 [DOI] [PubMed] [Google Scholar]
- Wang Y. P., Dakubo G., Howley P., Campsall K. D., Mazarolle C. J., Shiga S. A., Lewis P. M., McMahon A. P. and Wallace V. A. (2002). Development of normal retinal organization depends on Sonic hedgehog signaling from ganglion cells. Nat. Neurosci. 5, 831-832. [DOI] [PubMed] [Google Scholar]
- Yauch R. L., Gould S. E., Scales S. J., Tang T., Tian H., Ahn C. P., Marshall D., Fu L., Januario T., Kallop D. et al. (2008). A paracrine requirement for hedgehog signalling in cancer. Nature 455, 406-410. 10.1038/nature07275 [DOI] [PubMed] [Google Scholar]
- Zelinka C. P., Scott M. A., Volkov L. and Fischer A. J. (2012). The reactivity, distribution and abundance of Non-Astrocytic Inner Retinal Glial (NIRG) cells are regulated by microglia, acute damage, and IGF1. PLoS ONE 7, e44477 10.1371/journal.pone.0044477 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang X. M. and Yang X. J. (2001a). Regulation of retinal ganglion cell production by Sonic hedgehog. Development 128, 943-957. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang X.-M. and Yang X.-J. (2001b). Temporal and spatial effects of Sonic hedgehog signaling in chick eye morphogenesis. Dev. Biol. 233, 271-290. 10.1006/dbio.2000.0195 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.