

Summary
Natural sensory stimuli have a rich spatiotemporal structure and can often be characterized as a high frequency signal that is independently modulated at lower frequencies. This lower frequency modulation is known as the envelope. Envelopes are commonly found in a variety of sensory signals, such as contrast modulations of visual stimuli and amplitude modulations of auditory stimuli. While psychophysical studies have shown that envelopes can carry information that is essential for perception, how envelope information is processed in the brain is poorly understood. Here we review the behavioral salience and neural mechanisms for the processing of envelopes in the electrosensory system of wave-type gymnotiform weakly electric fishes. These fish can generate envelope signals through movement, interactions of their electric fields in social groups or communication signals. The envelopes that result from the first two behavioral contexts differ in their frequency content, with movement envelopes typically being of lower frequency. Recent behavioral evidence has shown that weakly electric fish respond in robust and stereotypical ways to social envelopes to increase the envelope frequency. Finally, neurophysiological results show how envelopes are processed by peripheral and central electrosensory neurons. Peripheral electrosensory neurons respond to both stimulus and envelope signals. Neurons in the primary hindbrain recipient of these afferents, the electrosensory lateral line lobe (ELL), exhibit heterogeneities in their responses to stimulus and envelope signals. Complete segregation of stimulus and envelope information is achieved in neurons in the target of ELL efferents, the midbrain torus semicircularis (Ts).
Keywords: second-order statistics, electrosensory system, jamming avoidance response, social envelope response
Introduction
How the brain processes natural stimuli for the control of behavior remains, generally speaking, poorly understood. This is in part because natural stimuli have complex spatiotemporal characteristics, many of which have not been fully described. One category of signal feature, envelopes, has only recently been appreciated for its widespread significance in the control of behavior. In practice, the envelope can be viewed as the line connecting successive maxima in the stimulus waveform and represents the ‘instantaneous amplitude’ of the stimulus (Fig. 1). We now know that most natural auditory (Heil, 2003; Zeng et al., 2005), visual (Derrington and Cox, 1998; Baker, 1999) and somatosensory (Lundstrom et al., 2010) stimuli contain salient slow time-varying envelopes. These envelopes are critical for perception and are used during a variety of tasks such as second-order visual processing (Grosof et al., 1993; Mareschal and Baker, 1998), stereopsis (Langley et al., 1999; Tanaka and Ohzawa, 2006), speech perception (Calhoun and Schreiner, 1998; Smith et al., 2002; Zeng et al., 2005; Gourévitch et al., 2008; Nourski et al., 2009) and sound localization (Lohuis and Fuzessery, 2000).
Fig. 1.

Envelope stimuli are found ubiquitously in sensory stimuli.
(A) Example of a contrast envelope stimulus in the visual system in which there is a fine-grain pattern (grating of high temporal frequency) whose contrast is modulated by a coarse-grain ‘envelope’ (red). (B) An excerpt from Tchaikovski’s 1812 Overture showing sound pressure (black) and its envelope (red). Note that the envelope corresponds to, in this case, the amplitude modulation (AM) of the pressure stimulus (black). (C) Electric organ discharge (EOD) waveform from Apteronotus leptorhynchus (black) with AM (green) and envelope (red) waveforms. We note that the envelope, for the electrosensory system, corresponds to the depth of modulation of the EOD AM. The inset (right) shows a snippet of all waveforms for a short time indicated by the cyan vertical bar.
Although envelopes are widely used in engineered systems such as radio communications, very little is known about how they are used and processed in biological systems. This is in part because natural envelope stimuli are complex and often cannot be captured in any simple mathematical expression (Heil and Neubauer, 2003). Further, the neural circuits that process envelopes are largely unknown.
In this review, we discuss advances made in understanding both the behavioral relevance and neural coding of envelope stimuli in the electrosensory system of wave-type Gymnotiform fishes. Specifically, electrosensory envelopes emerge as a consequence of the relative movement between fish (Yu et al., 2012), the interactions of their electric organ discharges (EODs) when in groups of three or more (Stamper et al., 2012), and the combination of movement and EOD interactions (Yu et al., 2012). Thus, these fish can influence the envelopes that they receive in two ways – through their own movement and by changing the frequency of their electric signal.
Weakly electric fish
Weakly electric fish use their electrosensory system for a variety of behaviors including navigation, prey capture, refuge tracking and obstacle avoidance (Caputi and Budelli, 2006; von der Emde, 2006). In addition, they also use their electrosensory system for conspecific communication (Hopkins, 1974; Fortune, 2006; Hupé and Lewis, 2008; Triefenbach and Zakon, 2008).
Wave-type electric fish have a specialized electric organ (EO), typically along the sides of their bodies and/or along the tail, composed of electrocytes derived from modified muscle cells (myogenic) or nerve cells (neurogenic) (Bennett, 1971). The EO generates a significant quasi-sinusoidal alternating current by summing ionic currents across cells that are simultaneously generated following command signals originating from the pacemaker nucleus (Pn) in the brain. The electric signal that results is known as the EOD (Babineau et al., 2006; Kelly et al., 2008; von der Emde et al., 2010), which is an electric field that surrounds the fish and may be detectable at distances of more than 1 m around the fish (Fig. 1C, inset, black).
The spatial and temporal features of the electric field are complex (Chen et al., 2005) and vary as a function of objects in the environment and the presence of other electric fish (Kelly et al., 2008; von der Emde et al., 2010). Historically, electrosensory stimuli in gymnotiform wave-type electric fish have been characterized in terms of the amplitude modulation (AM) and frequency modulation (FM) because there are two classes of electroreceptors, one for processing each attribute (Scheich, 1973). For this review, we will consider AM stimuli exclusively.
The AMs can be characterized in terms of their frequency content (Fig. 1C, green). In this regard, small prey items typically lead to low frequency (<10Hz) AMs (Nelson and MacIver, 1999; MacIver et al., 2001), whereas those caused by interaction with conspecifics lead to AMs with a wide frequency range (near 0 Hz to 400 Hz) (Heiligenberg, 1991; Zupanc and Maler, 1993; Ramcharitar et al., 2005; Tan et al., 2005; Ramcharitar et al., 2006).
However, these AMs can also be characterized in terms of their depth of modulation or contrast (Fig. 1C, red). In general, the depth of modulation is proportional to the distance of an object or conspecific from electroreceptors in the skin of an individual fish. The envelope, which is described in detail below, is the time-varying modulation of the depth of modulation of the AM and is a function of multiple environmental variables (Middleton et al., 2006; Middleton et al., 2007; Longtin et al., 2008; Savard et al., 2011; McGillivray et al., 2012; Stamper et al., 2012).
Electrosensory envelopes
Electrosensory envelopes can be generated through movement, the interactions of three or more EODs of conspecifics in close proximity (<1 m), or when fish emit a particular subtype of communication signal called a type I chirp in which the AM amplitude (i.e. the envelope) decreases by 50% for a brief period of time (50 ms) (Zakon et al., 2002). Observations in the natural habitat have shown that fish often form groups of two or more conspecifics (Tan et al., 2005; Stamper et al., 2010), are also commonly in motion, especially at night, and routinely emit communication signals (Hupé and Lewis, 2008). As such, envelopes are ubiquitous features of the natural electrosensory environment.
However, envelopes in the electrosensory system differ somewhat from those found in other (e.g. visual, auditory) senses. Indeed, the envelopes found in the visual (Fig. 1A, red) and in the auditory (Fig. 1B, red) systems correspond to the AM of the signal (i.e. a first order envelope). In contrast, the envelopes in the electrosensory system refer to the depth of modulation (Fig. 1C, red) of EOD AMs and would be equivalent to second order envelopes in the auditory and visual systems. It is important to note that, unlike vision and audition, electrosensation is a sensory modality in which perturbations of a self-generated signal carry the relevant information. As such, the EOD AM (Fig. 1C, green) is actually the stimulus waveform here.
Movement envelopes
When two wave-type electric fish are in close proximity, their EODs sum and the combined signal will display AMs and FMs. The rate of these modulations occurs at the frequency difference (dF) between the two fish. The depth of modulation of the AMs is determined by the relative positions of the two fish. For fish that are stationary, the depth of modulation is constant, whereas in fish that are moving, the depth of modulation varies with the movements of the fish (Fig. 2). In this way, movement creates an envelope by changing the depth of modulation, which occurs in relation to changes in both the distance and orientation between the two fish (Yu et al., 2012). Specifically, the depth of modulation increases when the distance between the two fish decreases (Fig. 2). In general (Figs 2, Figs 3, and4), the envelopes (red) caused by movement tend to contain lower temporal frequencies than the AMs (green). The information extracted from the time-varying envelope signal might give information as to the relative position between the two fish (Yu et al., 2012). In contrast, the AM frequency gives information as to the difference between the EOD frequencies, and can give information as to whether the other fish is the same sex (low dF) or opposite sex (high dF) in some species including Apteronotus leptorhynchus and Eigenmannia virescens.
Fig. 2.

Envelope generated by the relative movement of two weakly electric fish. (A) The movement trajectories of two individual fish where fish 1 is stationary and fish 2 is swimming (purple) is shown in the inset box. The distance (magenta) between individuals varies over time as a function of the movement of fish 2. The combined signal (black) is the sum of the EOD signal of fish 1 (S1; 300 Hz) and the EOD of fish 2 (S2; 350 Hz). This combined signal has an emergent amplitude modulation (green) and higher order envelope (red). Note that the black dots show the position of fish 2 (inset) and the distance between the two fish at time 0, respectively. (B) A Fourier transform (FT) analysis shows the power (i.e. the modulus squared of FT) for each of the signals – the combined signal (black), the AM (green) and the envelope (red). (C) A close-up of B, highlighting the power at low frequencies in the envelope due to the relative movement of the fish.
Fig. 3.

Envelope generated by the interaction of the EODs of three weakly electric fish. (A) Three fish that are not moving are shown in the inset. The magenta and orange lines indicate the constant distance of the two outer fish to the middle fish. The combined signal (black) is the sum of the EOD signals of the three fish, S1 (300 Hz), S2 (350 Hz) and S3 (395 Hz). The combined signal has an amplitude modulation (green) and a higher order envelope (red). (B) A Fourier analysis shows the power for each of the signals – the combined signal (black), the AM (green) and the envelope (red). (C) A close-up of B, highlighting the power at 5 Hz in the envelope due to our choice of EOD frequencies in this simulation.
Fig. 4.
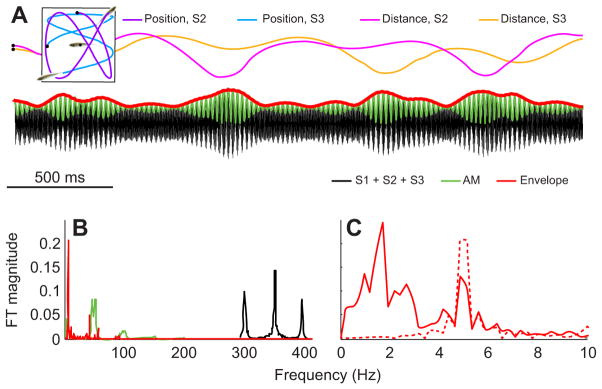
Interactions of movement and EODs for producing envelopes. In naturalistic conditions, weakly electric fish are often in groups of three or more individuals and are moving relative to one another, producing a complex envelope that contains a social and movement-generated component. (A) There are three fish, with one fish that is stationary and two fish (purple and blue) that are swimming (inset). The distance from fish 1 to each of the swimming fish (magenta and orange) varies as a function of the relative movement. The combined signal (black) contains the EODs of all three fish, an AM (green) and a complex envelope (red), which is composed of both the movement and the difference between the AM frequencies (ddF), from the social EOD interactions. (B) A Fourier analysis shows the power for each of the signals – the combined signal (black), the AM (green) and the envelope (red; consisting of movement and social components). (C) A close-up of B, highlighting the power below 5 Hz, which is due to the relative movement of the three fish, and at 5 Hz, which is due to the social envelope, and results from our choice of EOD frequencies (300, 350, 395 Hz) in this simulation. We have also replotted the power spectrum from the non-moving three fish case (red dotted line; from Fig. 2C) to highlight the potential separation between social and movement-generated envelopes.
Social envelopes
When three or more fish are in close proximity and are stationary, the EODs sum and the combined signal then consists of two prominent AMs [see Appendix in Stamper et al. (Stamper et al., 2012)]. If these two AMs have different frequencies (i.e. dFs), then the interaction between the two as they cycle in and out of phase with respect to one another can create an envelope that oscillates at the difference between the two AM frequencies (i.e. the ddF). Such ‘social envelopes’ do not require movement and are illustrated in Fig. 3. For example, if there are three stationary fish with EOD frequencies of 300 Hz (fish 1), 350 Hz (fish 2) and 395 Hz (fish 3), then fish 2 would experience AMs (green) with frequencies dF1=50 and dF2=45 Hz whose interference creates an envelope (red) at frequency ddF=5 Hz. In general, ‘social’ envelopes tend to contain higher temporal frequencies (>15Hz) (Stamper et al., 2010).
Combined movement and social envelopes
Weakly electric fish typically move with respect to one another in their natural environment and are often found in groups of three or more. As such, these fish experience both envelopes that result from interaction between their electric fields (i.e. ‘social envelopes’) as well as those that result from movement (i.e. ‘movement envelopes’). Thus, envelope signals in the natural environment will contain power due to movement and to interaction between the EODs of three or more fish. A specific case is illustrated in Fig. 4 where the temporal frequency content of the movement envelope component is lower but still overlaps with that of the social envelope component.
Communication envelopes
Weakly electric fish also emit communication signals called chirps (Zakon et al., 2002). It is important to note that these signals typically occur on top of the AMs that are created when two fish are in close proximity to one another. Chirps are typically associated to a transient (~50ms) increase in one of the fish’s EOD frequencies. However, for one particular chirp subtype (type I), the increase in EOD frequency is also associated with a ~50% decrease in EOD amplitude. This has the net effect of creating an envelope as the depth of modulation of the AM drops during the type I chirp. Type I chirps tend to be produced during opposite sex encounters, which are typically associated with a high frequency AM (Bastian et al., 2001).
Behavioral responses to envelopes
The social envelopes that have been observed in groups of three or more fish in natural habitats and in laboratory settings tend to have power at higher frequencies (>15 Hz) and not lower frequencies (Stamper et al., 2010). These data suggested that fish avoid producing low frequency social envelopes in their natural environment. A recent study has shown that weakly electric fish change their EOD frequencies in response to low frequency social envelopes (Stamper et al., 2012). In this behavior, fish shift their EOD frequency to increase the frequency of the social envelope, as discussed below.
The jamming avoidance response
Before describing behavioral responses to envelopes, it is useful to briefly review how some species (e.g. Eigenmannia and Apteronotus) of weakly electric fish respond to constant AMs (i.e. not envelopes) that are generated when two fish are in close proximity. When the AM frequency (dF) is low (<15Hz), the animal’s electrolocation abilities are impaired (Heiligenberg, 1974; Matsubara and Heiligenberg, 1978; Bastian, 1987) as the frequency content of the signals used in electrolocation and of the AM caused by the interaction of EODs overlap. This is known as ‘jamming’ and is thought to occur because neural responses to salient signals are then degraded (Ramcharitar et al., 2005).
Some species, including Eigenmannia and Apteronotus, solve this problem by shifting their EOD frequencies in order to increase the AM frequency (dF) to move it away from the frequency content of other behaviorally relevant electrosensory signals (e.g. those caused by prey). This behavior is known as the jamming avoidance response (JAR) (Watanabe and Takeda, 1963; Bullock et al., 1972). The underlying algorithm and the neural circuits that support the JAR have been fully elucidated (Heiligenberg, 1991; Metzner, 1993). It was previously shown that the JAR as observed in Eigenmannia requires integration of AM and FM information streams. Additionally, Eigenmannia respond to EOD AMs without the concurrent FM stimuli but this response differs from the JAR (Takizawa et al., 1999; Carlson and Kawasaki, 2007).
The social envelope response
Recent experiments by Stamper and colleagues show that Eigenmannia will either increase or decrease their EOD frequency in response to social envelope stimuli (Stamper, et al., 2012). This behavior was termed the social envelope response (SER). Indeed, when an individual fish is presented with two AMs whose interferences gives rise to a low frequency envelope, the animal shifts its EOD frequency by 2–10 Hz. The amount by which the animal changes its EOD frequency decreases with increasing envelope frequency (Fig. 5A) and increases with stimulus amplitude (Fig. 5B). The change in EOD frequency increases the envelope frequency to between 5 and 15Hz (Fig. 5C) (Stamper et al., 2012). This suggests that the SER may be a mechanism for avoiding low frequency social envelopes in the natural electrosensory environment, much like the proposed function of the JAR.
Fig. 5.
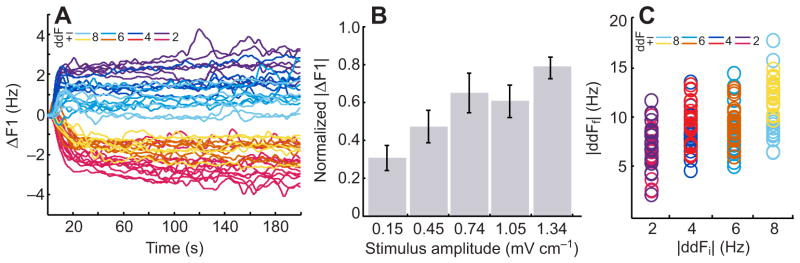
The social envelope response (SER). (A) Eigenmannia shift their EOD frequency (ΔF1) in response to envelope stimuli, but the response is dependent on the initial envelope frequency (color coded). Fish show stronger responses to initial envelopes that are lower (i.e. 2 and 4 Hz) in frequency. The response is bidirectional, whereby the frequency shift is down for positive ddFs and up for negative ddFs. (B) The strength of the response (normalized |ΔF1|) varies as a function of combined stimulus amplitude (S2 + S3), where fish show an increased change in EOD frequency for higher stimulus amplitudes. Under natural conditions, increased stimulus amplitude would be achieved by decreasing the distance between individuals, as often happens when they are swimming relative to one another. (C) The SER increases the frequency of the envelope, with the final envelope frequencies (|ddFf|) typically ending up in a range of 5 to 15 Hz for all initial envelope frequencies (|ddFi|) tested.
Additional studies are necessary to determine whether low frequency envelopes, like low frequency AMs, are detrimental to electrolocation behaviors. Alternatively, it is worth noting that in a complex envelope, in which there are both social and movement components, the movement envelope often has a lower frequency content and the social envelopes a higher frequency content (Fig. 4). The SER may also function to increase the separation between these two frequency bands to permit separate processing of movement and social envelope information in brain circuits.
An algorithm that would mediate the SER has also been determined (Stamper et al., 2012). When three or more EODs sum, plotting the amplitude and phase modulations of the EOD signal can, with additional low-pass filtering, give information as to the sign of the ddF, which is necessary in order for the fish to know whether to increase or decrease its own EOD frequency (Stamper et al., 2012). Further studies are needed to understand the neural circuits that perform the necessary computations required for the SER.
Coding of envelopes in the electrosensory system
There have been several recent advances towards understanding the neural algorithms for the extraction of behaviorally relevant information from envelope stimuli in electrosensory circuits. P-type tuberous electroreceptors project to pyramidal cells within the electrosensory lateral line lobe (ELL). Pyramidal cells then project to the midbrain torus semicircularis (Ts). Ts neurons project to the nucleus electrosensorius (nE), which projects to the prepacemaker nucleus (PPn). The PPn projects to the pacemaker nucleus (Pn), which sends commands to the EO, thereby completing the sensorimotor loop. This circuitry is well described elsewhere (Heiligenberg, 1991; Metzner, 1993).
Electroreceptor responses to envelopes
Electroreceptors of gymnotiform wave-type species are characterized by high baseline firing rates (150–600Hz) and firing rates monotonically increase with increasing EOD amplitude. Of course, receptors have the standard neural non-linearities that can influence their responses to envelopes including cut-off (i.e. their firing rate cannot be negative) and saturation (i.e. their firing rate cannot be greater than the EOD frequency as electroreceptors can fire at most one spike per EOD cycle) (Scheich et al., 1973; Bastian, 1981; Xu et al., 1996; Gussin et al., 2007). Further analysis has shown that their gains increase with stimulus frequency (Chacron et al., 2005; Xu et al., 1996). Therefore, electroreceptors have non-linear responses to electrosensory stimuli (AMs) that have amplitudes large enough to elicit non-linear saturation and/or cut-off. Further, as a consequence of increased gain to high frequency stimuli, electroreceptors display non-linear phase locking as well as increased synchronization to higher frequency AMs (Wessel et al., 1996; Nelson et al., 1997; Benda et al., 2005; Chacron et al., 2011).
As responses to envelope stimuli require a non-linear operation, mathematical models suggested that electroreceptors could respond to envelopes provided that the envelope signals exceed a certain threshold amplitude value (Longtin et al., 2008; Yu et al., 2012). Experimental recordings have confirmed these predictions by showing that electroreceptors can respond to envelopes arising from band-pass filtered white noise when the filter band contained high (>100 Hz) frequencies (Savard et al., 2011). Further analysis revealed that electroreceptor heterogeneities played a strong role in determining whether a given electroreceptor would respond to envelopes or not. Indeed, Savard and colleagues showed that it was the electroreceptor with the lowest baseline firing rate that tended to respond to envelopes because these units displayed the greatest tendency to reach cut-off during these stimuli (Savard et al., 2011).
The fact that electroreceptors can respond to envelopes when the amplitude of envelope signals exceeds a given threshold can be demonstrated by examining their responses to a sinusoidal AM stimulus whose depth of modulation (i.e. envelope) increases linearly with time (Fig. 6A). Indeed, electroreceptors tend to respond to these AMs with modulations in firing rate that are linearly related to the stimulus when the amplitude is low and with non-linear phase-locking when the amplitude is high (Fig. 6A). This has important consequences for determining whether the electroreceptor responds to the envelope or not. If the envelope amplitude is low enough, then the increase in firing rate during one half of the AM cycle will be compensated by the decrease in firing rate during the other half of the AM cycle. As such, the average firing rate during each AM cycle is then the same (Fig. 6B). However, the situation changes when the envelope amplitude is large enough to elicit static non-linearities such as cut-off. When cut-off is elicited, the increase in firing rate during one half of the AM cycle is not fully compensated by the decrease in firing rate during the other half, and the average firing rate increases (Fig. 6B).
Fig. 6.

Peripheral electrosensory neurons respond to envelopes. (A) Left: example stimulus in which the envelope (red) increases linearly as a function of time with a 20 Hz sinusoidal AM (green). a.u., arbitrary units. Right: example recording from an electroreceptor in response to this stimulus. It can be seen that the electroreceptor phase locks to high but not to low envelopes. Vm, membrane potential. (B) Peri-stimulus time histogram (PSTH) from the same electroreceptor. It can be seen that the firing rate is constant for low envelope values and increases linearly when the envelope is greater than a threshold value (dashed line). (C) Phase histograms when the envelope is lower than threshold (left) and higher than threshold (right). Phase locking (i.e. the absence of firing at some carrier phases) is observed only when the envelope is higher than threshold.
The amplitude level at which the firing rate starts either increasing or decreasing is therefore a threshold for the detection of envelope signals. In other words, electroreceptors do not respond to envelope stimuli when they are below the threshold and do respond to envelopes when they are above threshold due, at least in part, to non-linear phase locking (Fig. 6C). The exact threshold value is dependent on multiple factors including the baseline firing rate as well as the amplitude of the AM (Savard et al., 2011). As such, electroreceptors can either respond solely to the AM or to the AM as well as the envelope.
Electroreceptors also respond strongly to type I chirps. Indeed, the high frequency AM (i.e. constant non-zero envelope) causes non-linear synchronization in the electroreceptor population while the decrease in envelope during the chirp causes a transient desynchronization (Benda et al., 2006).
ELL pyramidal cell responses to envelopes
Pyramidal cells are the sole output neurons of the ELL. There are two pyramidal cell types, ON-cells that are excited (E-type) and OFF-cells that are inhibited (I-type) by increases in EOD amplitude (Saunders and Bastian, 1984). Like electroreceptors, the population of pyramidal cells is heterogeneous (Fig. 7A) (for review, see Maler, 2009). Pyramidal cells whose somata are found most superficially tend to display the largest dendritic trees and the smallest baseline firing rates while pyramidal cells whose somata are found most deeply tend to display the smallest dendritic trees and the largest baseline firing rates (Fig. 7A). This strong negative correlation between baseline firing rate and dendritic morphology allows one to easily identify a given pyramidal cell based on physiological measurements (Bastian and Nguyenkim, 2001; Bastian et al., 2004).
Fig. 7.

Hindbrain electrosensory neurons respond to envelopes. (A) Left: morphology of different electrosensory lateral line lobe (ELL) pyramidal cell classes. Note the large differences in dendritic tree length. Superficial pyramidal cells have the largest dendritic trees and are found superficially while deep pyramidal cells have the smallest dendritic trees and are found deep in the pyramidal cell layer. Intermediate pyramidal cells are in between. Right: firing rate and dendritic tree length are strongly negatively correlated. (B) Responses of example superficial (left) and deep (right) ELL pyramidal cells. (C) Phase histograms from these same cells to the AM stimulus. Note that only the superficial pyramidal cell (left) displays strong phase locking because the count is zero for phases between −π/2 and π/2. (D) Envelope response as a function of AM response for ELL pyramidal cells. Note that all cells respond to both. (E) Proposed neural circuit by which ELL pyramidal cells can respond to envelopes. An inhibitory interneuron, the ovoid cell, responds to the envelope via the non-linear action potential generation mechanism and sends this information primarily to superficial ELL pyramidal cells.
While it has been known for some time that pyramidal cells respond to envelopes (Middleton et al., 2006), recent experiments have shown a large heterogeneity in these responses (Marsat and Maler, 2010; McGillivray et al., 2012). Indeed, McGillivray and colleagues (McGillivray et al., 2012) showed that, while all pyramidal cells respond to both AMs and envelopes, superficial pyramidal cells tend to respond most strongly to envelopes while deep pyramidal cells tend to respond least strongly to envelopes (Fig. 7B). This is because superficial pyramidal cells, with their low baseline firing rates, tend to display the most cut-off in response to AMs (Fig. 7C). Nevertheless, all pyramidal cells respond to both carrier and envelope, but to different degrees based on heterogeneities in baseline firing rates (Fig. 7D). These findings are consistent with those of Marsat and Maler, who showed that ELL pyramidal cells respond differentially to type I chirps (Marsat and Maler, 2010).
How do pyramidal cells respond to the envelope? Envelope responses in pyramidal cells are not solely due to the threshold non-linearity described above: their subthreshold membrane activity already contains information about the envelope (Middleton et al., 2006). This subthreshold envelope information is presumably transmitted via the synaptic inputs to pyramidal cells. Middleton and colleagues showed that a class of inhibitory interneurons, called ovoid cells, responds to envelopes (Middleton et al., 2006). These responses are also most likely due to a threshold non-linearity. Middleton and colleagues (Middleton et al., 2006) concluded that inhibitory inputs from ovoid cells might be a mechanism underlying envelope responses in pyramidal cells (Fig. 7E).
While the respective contributions of inhibitory inputs and the threshold non-linearity towards determining the envelope responses of pyramidal cells have not been fully elucidated, it is possible that these mechanisms might act synergistically. This is because superficial pyramidal cells with the lowest baseline firing rates also receive the greatest proportion of inhibitory inputs, while deep pyramidal cells with the highest baseline firing rates receive the least inhibitory inputs (Bastian et al., 2002). However, further studies are necessary to fully understand the mechanisms by which pyramidal cells respond to envelopes. Again, while pyramidal cell heterogeneities can influence envelope responses, all pyramidal cells still respond to both envelope and AMs (Middleton et al., 2006; McGillivray et al., 2012).
Ts neural responses to envelopes
Electrophysiological studies have shown that Ts neurons can also respond to envelope stimuli (Fig. 8A–C). However, the responses of Ts neurons fall into three distinct categories. Ts neurons can respond (1) to both the AM and the envelope (Fig. 8D, blue), or (2) selectively to only the AM (Fig. 8D, green) or (3) to the envelope (Fig. 8D, red) (McGillivray et al., 2012). Some Ts neurons can also respond selectively to type I chirps (Vonderschen and Chacron, 2011). Therefore, segregation of information about envelope and AM stimuli appears to emerge at the level of the Ts. This segregation potentially resolves ambiguities as to whether a response is caused by the AM or the envelope. This is important as behavioral studies have shown that AM and envelope stimuli can overlap in their frequency ranges (Yu et al., 2012).
Fig. 8.

Midbrain electrosensory neurons respond to envelopes. (A) The stimulus consists of a noisy AM (green) with corresponding envelope (red).
(B) Response from an example Ts neuron to the stimulus shown in A. This neuron responds strongly to the envelope. (C) PSTH response from this same Ts neuron to the stimulus shown in A. (D) Envelope response as a function of AM response from Ts neurons. In contrast with ELL pyramidal cells, three distinct clusters are seen. Some Ts neurons respond selectively to either the envelope (red) or the AM (green) while some respond to both (blue). (E) Model in which a Ts neuron receives input from both ON- and OFF-type ELL pyramidal cells. The strength of the input from ON-cells is given by ρON while the strength of the input from OFF-cells is given by 1–ρON. Both inputs are thus balanced in strength when ρON=0.5. (F) Model results showing the AM (green) and envelope (red) responses as a function of ρON. (G) Model results showing the AM (green) and envelope (red) responses as a function of the baseline firing rate. (H) Proposed schematic diagram by which Ts neurons acquire their response selectivity to envelopes. ON-and OFF-type ELL pyramidal cells respond during the positive and negative phases of the AM, respectively (brown and green). However, both tend to respond more when the envelope is high. Summing these responses gives a response that is independent of AM but will still depend on the envelope (black).
How can Ts neurons respond selectively to either the envelope or the AM? Anatomical studies have shown that Ts neurons receive excitatory inputs from both ON- and OFF-type ELL pyramidal cells although it is not known whether a given Ts neuron can receive input from both ON- and OFF-type cells at the same time (Carr and Maler, 1985). McGillivray and colleagues (McGillivray et al., 2012) used spike-triggered covariance analysis in order to investigate the different sources (ON-, OFF-, or both ON- and OFF-type) of inputs received by Ts neurons. Spike-triggered averages of the stimulus showed that envelope-selective neurons apparently did not respond to features of the AM. However, covariance analysis showed that envelope-selective Ts neurons, and not the other two categories of Ts neurons, exhibited great variability in their responses that could not be explained by the variability of the stimulus. Further analysis revealed that such neurons did not respond to the AM because they spiked in response to both the increases as well as the decreases in the AM. Indeed, because those features were matched in strength, they essentially canceled out and, when averaged, resulted in no effective response to the AM. However, because the neuron is responding to both phases of the AM, changes in the depth of modulation (i.e. envelope) will result in changes in the magnitude of firing. In this way, the neuron responds to the changes in the envelope and not the AM.
McGillivray and colleagues (McGillivray et al., 2012) tested this interesting idea by building a mathematical model in which they could vary the relative balance of inputs from ON- and OFF-type ELL pyramidal cells (Fig. 8E). They found that the model neuron responded selectively to the envelope only when the two sources of input were matched in strength (Fig. 8F). When there was a mismatch, the model neuron instead responded to both AM and envelope. Moreover, this model made an important prediction. When they increased the baseline firing rate of the model ELL neurons, thereby mimicking the baseline firing rates of deep ELL pyramidal cells, they found that the model Ts neuron tended to respond most selectively to the AM and not to the envelope.
Thus, these results strongly suggest that Ts neurons acquire selectivity to the envelope stimuli by combining balanced inputs from ON- and OFF-type pyramidal cells. The algorithm is the following: ON- and OFF-type pyramidal cells display rectification in their responses but at opposite phases of the AM stimulus, but are essentially in-phase with respect to the envelope stimulus (Fig. 8H). These responses are then summed and low-pass filtered in order to give rise to a selective envelope response in ON–OFF Ts neurons (Fig. 8H). Further experiments performed by McGillivray and colleagues (McGillivray et al., 2012) show that these correspond to the ON–OFF (‘E–I’) neurons that were previously reported (Partridge et al., 1981) and that they thus most likely form a distinct cell class within the Ts. Ts neurons that respond to both AM and envelope are thought to receive unbalanced input from ON- and OFF-type ELL pyramidal cells.
Thus, the following neural circuit is most likely responsible for segregation of envelope and carrier information in Ts. Envelope-selective Ts neurons are thought to receive balanced input from ON- and OFF-type superficial ELL pyramidal cells. This is because these cells tend to give the strongest responses to envelopes. In contrast, AM-selective Ts neurons most likely receive input from deep ON-type ELL pyramidal cells (Fig. 9).
Fig. 9.

Summary of the electrosensory neural circuits by which envelope and AM information streams are segregated. Electroreceptors synapse onto deep and superficial ELL pyramidal cells. Envelope-selective Ts neurons receive balanced input from superficial ON- and OFF-type pyramidal cells while AM-selective Ts neurons receive input from ON-type deep pyramidal cells that tend to respond the least to envelope stimuli.
Discussion
Electrosensory envelopes arise as a result of interactions between the EODs of three or more motionless fish (‘social envelopes’), movement when two or more fish are in close proximity (‘movement envelopes’), and the production of specific types of intermittent social signaling. In general, envelopes found in the natural electrosensory environment likely result from a combination of each of these categories. Some species of weakly electric fish exhibit behavioral responses to social envelopes (the SER) in order to increase the frequency content of the envelope. Finally, electrosensory circuits appear to be devoted to the processing of envelopes as some midbrain neurons respond selectively to them. However, there remain many questions related to the perception and coding of envelope stimuli by weakly electric fish.
First, it is important to realize that both Middleton and colleagues (Middleton et al., 2006) as well as McGillivray and colleagues (McGillivray et al., 2012) used stimulus intensities that did not elicit envelope responses from electroreceptors. As such, how ELL pyramidal cells and subsequently Ts neurons integrate envelope information from multiple sources is unknown and will be a fascinating area of research.
Second, how electrosensory neurons in higher order areas (e.g. nE) respond to envelopes is currently unknown. It will be critical to record from these higher order neurons in order to determine where and how information from envelope-selective Ts neurons is processed in higher order brain areas.
Third, while Stamper and colleagues (Stamper et al., 2012) have proposed an algorithm that would explain the SER, further studies should investigate whether the neural circuits that mediate the SER are indeed similar to those that mediate the JAR.
Finally, while social envelopes may interfere with the animal’s electrolocation abilities, this is not the case for movement envelopes that might contain information related to environmental variables (e.g. the distance between both fish). Further studies should focus on whether these movement envelopes interfere with the animal’s electrolocation abilities, whether electrosensory neurons can respond to these very low frequency signals, and whether weakly electric fish actually exhibit behavioral responses to movement envelopes.
Acknowledgments
We would like to thank M. G. Metzen for help in making Fig. 1.
Funding
This material is based upon work supported by the Canada Research Chairs (M.J.C.), the Canadian Institutes of Health Research (M.J.C.), the Natural Sciences and Engineering Research Council (M.J.C.), the National Science Foundation [grant nos IOS-0817918 to E.S.F.; CMMI-0941674, E.S.F.; CISE-0845749 to E.S.F.) and the Office of Naval Research [grant no. N000140910531 to E.S.F.].
List of symbols and abbreviations
- AM
amplitude modulation
- ddF
difference of dFs
- dF
frequency difference
- ELL
electrosensory lateral line lobe
- EO
electric organ
- EOD
electric organ discharge
- FM
frequency modulation
- FT
Fourier transform
- JAR
jamming avoidance response
- nE
nucleus electrosensorius
- Pn
pacemaker nucleus
- PPn
pre-pacemaker nucleus
- PSTH
peri-stimulus time histogram
- SER
social envelope response
- Ts
torus semicircularis
Footnotes
Author contributions
S.A.S., E.S.F. and M.J.C. wrote the manuscript.
Competing interests
No competing interests declared.
References
- Babineau D, Longtin A, Lewis JE. Modeling the electric field of weakly electric fish. J Exp Biol. 2006;209:3636–3651. doi: 10.1242/jeb.02403. [DOI] [PubMed] [Google Scholar]
- Baker CL., Jr Central neural mechanisms for detecting second-order motion. Curr Opin Neurobiol. 1999;9:461–466. doi: 10.1016/S0959-4388(99)80069-5. [DOI] [PubMed] [Google Scholar]
- Bastian J. Electrolocation I. How the electroreceptors of Apteronotus albifrons code for moving objects and other electrical stimuli. J Comp Physiol A. 1981;144:465–479. [Google Scholar]
- Bastian J. Electrolocation in the presence of jamming signals: behavior. J Comp Physiol A. 1987;161:811–824. doi: 10.1007/BF00610223. [DOI] [PubMed] [Google Scholar]
- Bastian J, Nguyenkim J. Dendritic modulation of burst-like firing in sensory neurons. J Neurophysiol. 2001;85:10–22. doi: 10.1152/jn.2001.85.1.10. [DOI] [PubMed] [Google Scholar]
- Bastian J, Schniederjan S, Nguyenkim J. Arginine vasotocin modulates a sexually dimorphic communication behavior in the weakly electric fish Apteronotus leptorhynchus. J Exp Biol. 2001;204:1909–1923. doi: 10.1242/jeb.204.11.1909. [DOI] [PubMed] [Google Scholar]
- Bastian J, Chacron MJ, Maler L. Receptive field organization determines pyramidal cell stimulus-encoding capability and spatial stimulus selectivity. J Neurosci. 2002;22:4577–4590. doi: 10.1523/JNEUROSCI.22-11-04577.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bastian J, Chacron MJ, Maler L. Plastic and nonplastic pyramidal cells perform unique roles in a network capable of adaptive redundancy reduction. Neuron. 2004;41:767–779. doi: 10.1016/s0896-6273(04)00071-6. [DOI] [PubMed] [Google Scholar]
- Benda J, Longtin A, Maler L. Spike-frequency adaptation separates transient communication signals from background oscillations. J Neurosci. 2005;25:2312–2321. doi: 10.1523/JNEUROSCI.4795-04.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Benda J, Longtin A, Maler L. A synchronization-desynchronization code for natural communication signals. Neuron. 2006;52:347–358. doi: 10.1016/j.neuron.2006.08.008. [DOI] [PubMed] [Google Scholar]
- Bennett MVL. Electrolocation in fish. Ann New York Acad Sci. 1971;188:242–269. doi: 10.1111/j.1749-6632.1971.tb13102.x. [DOI] [PubMed] [Google Scholar]
- Bullock TH, Hamstra JR, Scheich H. The jamming avoidance response of high frequency electric fish. J Comp Physiol A. 1972;77:1–22. [Google Scholar]
- Calhoun BM, Schreiner CE. Spectral envelope coding in cat primary auditory cortex: linear and non-linear effects of stimulus characteristics. Eur J Neurosci. 1998;10:926–940. doi: 10.1046/j.1460-9568.1998.00102.x. [DOI] [PubMed] [Google Scholar]
- Caputi AA, Budelli R. Peripheral electrosensory imaging by weakly electric fish. J Comp Physiol A. 2006;192:587–600. doi: 10.1007/s00359-006-0100-2. [DOI] [PubMed] [Google Scholar]
- Carlson BA, Kawasaki M. Behavioral responses to jamming and phantom jamming stimuli in the weakly electric fish Eigenmannia. J Comp Physiol A. 2007;193:927–941. doi: 10.1007/s00359-007-0246-6. [DOI] [PubMed] [Google Scholar]
- Carr CE, Maler L. A Golgi study of the cell types of the dorsal torus semicircularis of the electric fish Eigenmannia: functional and morphological diversity in the midbrain. J Comp Neurol. 1985;235:207–240. doi: 10.1002/cne.902350206. [DOI] [PubMed] [Google Scholar]
- Chacron MJ, Maler L, Bastian J. Electroreceptor neuron dynamics shape information transmission. Nat Neurosci. 2005;8:673–678. doi: 10.1038/nn1433. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chacron MJ, Longtin A, Maler L. Efficient computation via sparse coding in electrosensory neural networks. Curr Opin Neurobiol. 2011;21:752–760. doi: 10.1016/j.conb.2011.05.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen L, House JL, Krahe R, Nelson ME. Modeling signal and background components of electrosensory scenes. J Comp Physiol A. 2005;191:331–345. doi: 10.1007/s00359-004-0587-3. [DOI] [PubMed] [Google Scholar]
- Derrington A, Cox M. Temporal resolution of dichoptic and second-order motion mechanisms. Vision Res. 1998;38:3531–3539. doi: 10.1016/s0042-6989(98)00050-9. [DOI] [PubMed] [Google Scholar]
- Fortune ES. The decoding of electrosensory systems. Curr Opin Neurobiol. 2006;16:474–480. doi: 10.1016/j.conb.2006.06.006. [DOI] [PubMed] [Google Scholar]
- Gourévitch B, Le Bouquin Jeannès R, Faucon G, Liégeois-Chauvel C. Temporal envelope processing in the human auditory cortex: response and interconnections of auditory cortical areas. Hear Res. 2008;237:1–18. doi: 10.1016/j.heares.2007.12.003. [DOI] [PubMed] [Google Scholar]
- Grosof DH, Shapley RM, Hawken MJ. Macaque V1 neurons can signal illusory contours. Nature. 1993;365:550–552. doi: 10.1038/365550a0. [DOI] [PubMed] [Google Scholar]
- Gussin D, Benda J, Maler L. Limits of linear rate coding of dynamic stimuli by electroreceptor afferents. J Neurophysiol. 2007;97:2917–2929. doi: 10.1152/jn.01243.2006. [DOI] [PubMed] [Google Scholar]
- Heil P. Coding of temporal onset envelope in the auditory system. Speech Commun. 2003;41:123–134. [Google Scholar]
- Heil P, Neubauer H. A unifying basis of auditory thresholds based on temporal summation. Proc Natl Acad Sci USA. 2003;100:6151–6156. doi: 10.1073/pnas.1030017100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heiligenberg W. Electrolocation and jamming avoidance in a Hypopygus (Rhamohichthyidae, Gymnotoidei), an electric fish with pulse-type discharges. J Comp Physiol A. 1974;91:223–240. [Google Scholar]
- Heiligenberg W. Neural Nets in Electric Fish. Cambridge: MIT Press; 1991. [Google Scholar]
- Hopkins CD. Electric communication: functions in the social behavior of Eigenmannia virescens. Behaviour. 1974;50:270–304. [Google Scholar]
- Hupé GJ, Lewis JE. Electrocommunication signals in free swimming brown ghost knifefish, Apteronotus leptorhynchus. J Exp Biol. 2008;211:1657–1667. doi: 10.1242/jeb.013516. [DOI] [PubMed] [Google Scholar]
- Kelly M, Babineau D, Longtin A, Lewis JE. Electric field interactions in pairs of electric fish: modeling and mimicking naturalistic inputs. Biol Cybern. 2008;98:479–490. doi: 10.1007/s00422-008-0218-0. [DOI] [PubMed] [Google Scholar]
- Langley K, Fleet DJ, Hibbard PB. Stereopsis from contrast envelopes. Vision Res. 1999;39:2313–2324. doi: 10.1016/s0042-6989(98)00271-5. [DOI] [PubMed] [Google Scholar]
- Lohuis TD, Fuzessery ZM. Neuronal sensitivity to interaural time differences in the sound envelope in the auditory cortex of the pallid bat. Hear Res. 2000;143:43–57. doi: 10.1016/s0378-5955(00)00021-6. [DOI] [PubMed] [Google Scholar]
- Longtin A, Middleton JW, Cieniak J, Maler L. Neural dynamics of envelope coding. Math Biosci. 2008;214:87–99. doi: 10.1016/j.mbs.2008.01.008. [DOI] [PubMed] [Google Scholar]
- Lundstrom BN, Fairhall AL, Maravall M. Multiple timescale encoding of slowly varying whisker stimulus envelope in cortical and thalamic neurons in vivo. J Neurosci. 2010;30:5071–5077. doi: 10.1523/JNEUROSCI.2193-09.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- MacIver MA, Sharabash NM, Nelson ME. Prey-capture behavior in gymnotid electric fish: motion analysis and effects of water conductivity. J Exp Biol. 2001;204:543–557. doi: 10.1242/jeb.204.3.543. [DOI] [PubMed] [Google Scholar]
- Maler L. Receptive field organization across multiple electrosensory maps. I. Columnar organization and estimation of receptive field size. J Comp Neurol. 2009;516:376–393. doi: 10.1002/cne.22124. [DOI] [PubMed] [Google Scholar]
- Mareschal I, Baker CL., Jr Temporal and spatial response to second-order stimuli in cat area 18. J Neurophysiol. 1998;80:2811–2823. doi: 10.1152/jn.1998.80.6.2811. [DOI] [PubMed] [Google Scholar]
- Marsat G, Maler L. Neural heterogeneity and efficient population codes for communication signals. J Neurophysiol. 2010;104:2543–2555. doi: 10.1152/jn.00256.2010. [DOI] [PubMed] [Google Scholar]
- Matsubara JA, Heiligenberg W. How well do electric fish electrolocate under jamming. J Comp Physiol A. 1978;125:285–290. [Google Scholar]
- McGillivray P, Vonderschen K, Fortune ES, Chacron MJ. Parallel coding of first- and second-order stimulus attributes by midbrain electrosensory neurons. J Neurosci. 2012;32:5510–5524. doi: 10.1523/JNEUROSCI.0478-12.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Metzner W. The jamming avoidance response in Eigenmannia is controlled by two separate motor pathways. J Neurosci. 1993;13:1862–1878. doi: 10.1523/JNEUROSCI.13-05-01862.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Middleton JW, Longtin A, Benda J, Maler L. The cellular basis for parallel neural transmission of a high-frequency stimulus and its low-frequency envelope. Proc Natl Acad Sci USA. 2006;103:14596–14601. doi: 10.1073/pnas.0604103103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Middleton JW, Harvey-Girard E, Maler L, Longtin A. Envelope gating and noise shaping in populations of noisy neurons. Phys Rev E Stat Nonlin Soft Matter Phys. 2007;75:021918. doi: 10.1103/PhysRevE.75.021918. [DOI] [PubMed] [Google Scholar]
- Nelson ME, MacIver MA. Prey capture in the weakly electric fish Apteronotus albifrons: sensory acquisition strategies and electrosensory consequences. J Exp Biol. 1999;202:1195–1203. doi: 10.1242/jeb.202.10.1195. [DOI] [PubMed] [Google Scholar]
- Nelson ME, Xu Z, Payne JR. Characterization and modeling of P-type electrosensory afferent responses to amplitude modulations in a wave-type electric fish. J Comp Physiol A. 1997;181:532–544. doi: 10.1007/s003590050137. [DOI] [PubMed] [Google Scholar]
- Nourski KV, Reale RA, Oya H, Kawasaki H, Kovach CK, Chen H, Howard MA, 3rd, Brugge JF. Temporal envelope of time-compressed speech represented in the human auditory cortex. J Neurosci. 2009;29:15564–15574. doi: 10.1523/JNEUROSCI.3065-09.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Partridge BL, Heiligenberg W, Matsubara J. The neural basis of a sensory filter in the jamming avoidance response: No grandmother cells in sight. J Comp Physiol A. 1981;145:153–168. [Google Scholar]
- Ramcharitar JU, Tan EW, Fortune ES. Effects of global electrosensory signals on motion processing in the midbrain of Eigenmannia. J Comp Physiol A. 2005;191:865–872. doi: 10.1007/s00359-005-0008-2. [DOI] [PubMed] [Google Scholar]
- Ramcharitar JU, Tan EW, Fortune ES. Global electrosensory oscillations enhance directional responses of midbrain neurons in Eigenmannia. J Neurophysiol. 2006;96:2319–2326. doi: 10.1152/jn.00311.2006. [DOI] [PubMed] [Google Scholar]
- Saunders J, Bastian J. The physiology and morphology of two classes of electrosensory neurons in the weakly electric fish Apteronotus leptorhynchus. J Comp Physiol A. 1984;154:199–209. [Google Scholar]
- Savard M, Krahe R, Chacron MJ. Neural heterogeneities influence envelope and temporal coding at the sensory periphery. Neuroscience. 2011;172:270–284. doi: 10.1016/j.neuroscience.2010.10.061. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scheich H, Bullock TH, Hamstra RH., Jr Coding properties of two classes of afferent nerve fibers: high-frequency electroreceptors in the electric fish, Eigenmannia. J Neurophysiol. 1973;36:39–60. doi: 10.1152/jn.1973.36.1.39. [DOI] [PubMed] [Google Scholar]
- Smith ZM, Delgutte B, Oxenham AJ. Chimaeric sounds reveal dichotomies in auditory perception. Nature. 2002;416:87–90. doi: 10.1038/416087a. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stamper SA, Carrera-G E, Tan EW, Fugère V, Krahe R, Fortune ES. Species differences in group size and electrosensory interference in weakly electric fishes: implications for electrosensory processing. Behav Brain Res. 2010;207:368–376. doi: 10.1016/j.bbr.2009.10.023. [DOI] [PubMed] [Google Scholar]
- Stamper SA, Madhav MS, Cowan NJ, Fortune ES. Beyond the jamming avoidance response: weakly electric fish respond to the envelope of social electrosensory signals. J Exp Biol. 2012;215:4196–4207. doi: 10.1242/jeb.076513. [DOI] [PubMed] [Google Scholar]
- Takizawa Y, Rose GJ, Kawasaki M. Resolving competing theories for control of the jamming avoidance response: the role of amplitude modulations in electric organ discharge decelerations. J Exp Biol. 1999;202:1377–1386. doi: 10.1242/jeb.202.10.1377. [DOI] [PubMed] [Google Scholar]
- Tan EW, Nizar JM, Carrera-G E, Fortune ES. Electrosensory interference in naturally occurring aggregates of a species of weakly electric fish, Eigenmannia virescens. Behav Brain Res. 2005;164:83–92. doi: 10.1016/j.bbr.2005.06.014. [DOI] [PubMed] [Google Scholar]
- Tanaka H, Ohzawa I. Neural basis for stereopsis from second-order contrast cues. J Neurosci. 2006;26:4370–4382. doi: 10.1523/JNEUROSCI.4379-05.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Triefenbach F, Zakon H. Changes in the signalling during agnostic interactions between male weakly electric knifefish, Apteronotus leptorhynchus. Anim Behav. 2008;75:1263–1272. [Google Scholar]
- von der Emde G. Non-visual environmental imaging and object detection through active electrolocation in weakly electric fish. J Comp Physiol A. 2006;192:601–612. doi: 10.1007/s00359-006-0096-7. [DOI] [PubMed] [Google Scholar]
- von der Emde G, Behr K, Bouton B, Engelmann J, Fetz S, Folde C. 3-Dimensional scene perception during active electrolocation in a weakly electric pulse fish. Front Behav Neurosci. 2010;4:26. doi: 10.3389/fnbeh.2010.00026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vonderschen K, Chacron MJ. Sparse and dense coding of natural stimuli by distinct midbrain neuron subpopulations in weakly electric fish. J Neurophysiol. 2011;106:3102–3118. doi: 10.1152/jn.00588.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Watanabe A, Takeda K. The change of discharge frequency by A.C. stimulus in a weak electric fish. J Exp Biol. 1963;40:57–66. [Google Scholar]
- Wessel R, Koch C, Gabbiani F. Coding of time-varying electric field amplitude modulations in a wave-type electric fish. J Neurophysiol. 1996;75:2280–2293. doi: 10.1152/jn.1996.75.6.2280. [DOI] [PubMed] [Google Scholar]
- Xu Z, Payne JR, Nelson ME. Logarithmic time course of sensory adaptation in electrosensory afferent nerve fibers in a weakly electric fish. J Neurophysiol. 1996;76:2020–2032. doi: 10.1152/jn.1996.76.3.2020. [DOI] [PubMed] [Google Scholar]
- Yu N, Hupé GJ, Garfinkle C, Lewis JE, Longtin A. Coding conspecific identity and motion in the electric sense. PLOS Comput Biol. 2012;8:e1002564. doi: 10.1371/journal.pcbi.1002564. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zakon HH, Oestreich J, Tallarovic S, Triefenbach F. EOD modulations of brown ghost electric fish: JARs, chirps, rises, and dips. J Physiol Paris. 2002;96:451–458. doi: 10.1016/S0928-4257(03)00012-3. [DOI] [PubMed] [Google Scholar]
- Zeng FG, Nie K, Stickney GS, Kong YY, Vongphoe M, Bhargave A, Wei C, Cao K. Speech recognition with amplitude and frequency modulations. Proc Natl Acad Sci USA. 2005;102:2293–2298. doi: 10.1073/pnas.0406460102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zupanc GKH, Maler L. Evoked chirping in the weakly electric fish Apteronotus leptorhynchus: a quantitative biophysical analysis. Can J Zool. 1993;71:2301–2310. [Google Scholar]