

Abstract
‘The best way to predict the future is to create it.’ When we look at drugs that are used to control parasites, we see that new knowledge has been created (discovered) about their modes of action. This knowledge will allow us to predict combinations of drugs which can be used together rationally to increase the spectrum of action and to slow the development of anthelmintic resistance. In this paper we comment on some recent observations of ours on the modes of action of emodepside, diethylcarbamazine and tribendimidine. Emodepside increases the activation of a SLO-1 K+ current inhibiting movement, and diethylcarbamazine has a synergistic effect on the effect of emodepside on the SLO-1 K+ current, increasing the size of the response. The combination may be considered for further testing for therapeutic use. Tribendimidine is a selective cholinergic nematode B-subtype nAChR agonist, producing muscle depolarization and contraction. It has different subtype selectivity to levamisole and may be effective in the presence of some types of levamisole resistance. The new information about the modes of action may aid the design of rational drug combinations designed to slow the development of resistance or increase the spectrum of action.
Keywords: Anthelmintic, mode of action, tribendimidine, emodepside, diethylcarbamazine
1. INTRODUCTION
‘The best way to predict the future is to create it.’ This quote is usually ascribed to Abraham Lincoln and leads to some interesting thoughts when applied to different research fields including our own field of study, that is, anthelmintics. What are we creating and what can we see for the future for anti-parasitic drugs? We are creating new techniques for the screening of anthelmintic drugs, we are creating new methods for studying the modes of actions of anthelmintics and we are creating new ways for detecting anthelmintic resistance (Gilleard and Beech, 2007; Lanusse et al., 2014). We are also creating better awareness of the ‘Neglected Tropical Diseases’ (Hotez et al., 2008) of humans which include ascariasis, hookworm and trichuriasis and better awareness of the link between human and animal medicine, the ‘One Health’ concept. Our western economic system has also produced a more favorable economic environment for the development of animal anthelmintics, than anthelmintics for humans. Currently, we limit the damage done by nematode parasites of animals with pasture management, and improved targeted metaphylactic and therapeutic use of anthelmintics; for humans, clean water and sanitation are limiting factors (like clean pasture for animals) while mass drug administration (MDA) has similarities to the regular metaphylactic use of anthelmintics for animals. Vaccinations against parasites for both humans and animals are very desirable but so far they have limited efficacy.
The advances and creations listed above lead to a number of logical predictions for the future use and development of anthelmintics. We are confident that the development and better understanding of anthelmintic properties will continue, at a steady but modest pace, driven by economic, human and animal needs. We think that the economic pressures associated with animal medicine will remain greater than for human medicine and focused on the development of novel ‘resistance-busting’ anthelmintics. Success in research can be limited by funding as was said by the father of modern drug development, Paul Ehrlich Fig. 1 (1854–1915), who recognized success required the four Gs: Gluck, Geduld, Geshcick und Geld (luck, patience, skill and money, (PERUTZ, 1988). The ‘money’ for animal anthelmintics comes from the market for anti-parasitic drugs and chemicals for small animals and livestock which was estimated at $11 billion (Evans and Chapple, 2002). This contrasts with the market for human anthelmintics which is only $0.5 billion despite some 2 billion humans being infected in developing countries, around 25 cents per person annually! Given that the out of pocket costs of new human drugs may be $403 million (at year 2000 valuations (DiMasi et al., 2003) or more, we can see that the commercial development of human anthelmintic drugs is not favorable. It seems more likely that the economics of animal health will drive the development and advance of the knowledge base of animal anthelmintics and that these developments will be applied and adapted for human use (ivermectin and perhaps emodepside) unless private charities, governments and foundations overcome the financial limitations. In addition to the gradual development of resistance-busting anthelmintics we see: developments in our understanding of the modes of action of anthelmintics; we see more logical combinations of anthelmintics to slow down or counter the development of resistance and; also new methods for detecting anthelmintic resistance.
Fig. 1.

Paul Erlich in his Frankfurt office, circa 1900, the father of modern chemotherapy, who worked on trypanosome diseases and popularized the concept of the ‘magic bullet’ (magische Kugel, the perfect therapeutic agent).
Our lab has focused on understanding of the modes of action of anthelmintics and in this paper we illustrate some of our recent observations and developments in our understanding of the actions of emodepside, diethylcarbamazine and tribendimidine. The mode of action of these anthelmintics involves effects on membrane ion-channels and has required us to use electrophysiological techniques for their study (Martin et al., 1996b). We think that better knowledge of the mode of action of these compounds will allow rational combination with other anthelmintics to increase potency, spectra and allow a slowing of the speed of development of resistance in animal and human parasites. This paper is based on a lecture given to the World Association for the Advancement of Veterinary Parasitology (WAAVP) in Liverpool, 2015 and covers: 1) emodepside, an anthelmintic used for small animals, which has the potential for being used for human use to control filarial parasites; 2) diethylcarbamazine, a long serving anthelmintic still used for the control of filariasis in humans and; 3) tribendimidine, a recent anthelmintic developed by China for human use.
2. Emodepside
Emodepside is a cyclooctadepsipeptide developed by Bayer, Fig. 2, which is related to its parent compound, PF1022A (Martin et al., 1996a) which has broad-spectrum anthelmintic activity. Emodepside has an inhibitory effect on locomotion that allowed (Guest et al., 2007) to use a C. elegans mutagenesis screen to find that slo-1 (a Ca2+-dependent K+ channel) mutant alleles were resistant to the inhibitory effects of emodepside. (Crisford et al., 2011) also described transgenic experiments in which C. elegans SLO-1a channels were swapped for KCNMA1, the human orthologue of SLO-1 channels. These studies found that the sensitivity to emodepside in the rescues depended upon the origin of the SLO-1 channel: the human KCNMA1 channel was 10–100 times less sensitive to emodepside than the rescues expressing C. elegans SLO-1a channel. These experiments suggested that SLO-1 Ca2+-dependent K+ channels of nematodes are a major part of the target site of emodepside. Expression of C. elegans SLO-1a channels in Xenopus oocytes have revealed that emodepside can directly open these channels (Kulke et al., 2014) and that emodepside action on this splice variant of the channel does not require the presence of additional receptors like the latrophilin receptors for an emodepside effect (Willson et al., 2004). It does not however, rule out a contribution of latrophilin receptors to the overall mode of action of emodepside in parasites as indicated by the interaction of Lat-1 and emodepside (Saeger et al., 2001).
Fig. 2.
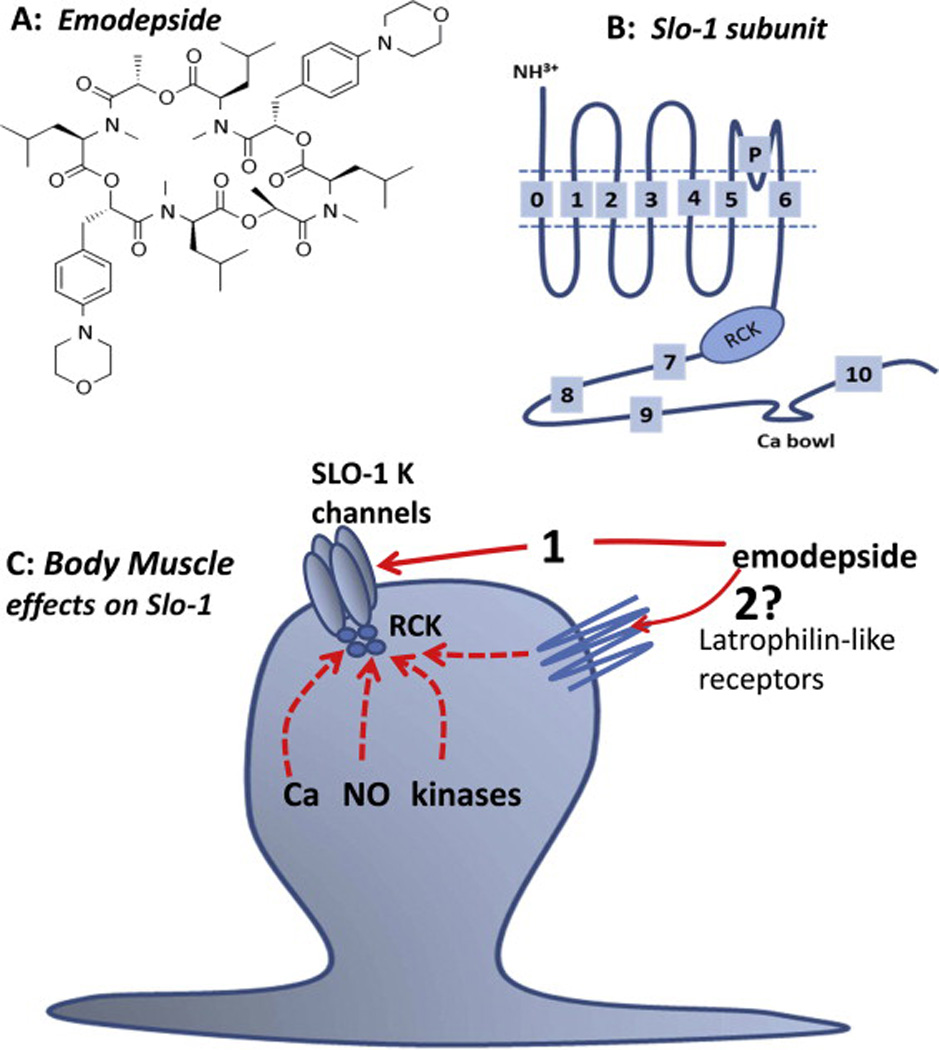
Summary diagrams of emodepside structure, Slo-1 subunit and a model of the mode action of emodepside on nematode body muscle. A: Emodepside. B: Line diagram of structure of one SLO-1 subunit; each SLO-1 K+ channel is made up of 4 of these subunits. C. Putative mode of action of emodepside on SLO-1 K+ channels in the muscle: act directly (1) or indirectly by stimulating latrophilin-like receptors (2) and signaling cascades that may involve NO, protein Kinase C and/or calcium. It is unlikely that emodepside acts at the extracellular surface of the SLO-1 K+ channel because of the slow time course of its action. It is very lipophilic and could act in the lipid membrane phase on the SLO-1 K+ channel or move into the cytoplasm and act intracellularly. A SLO-1 K+ channel (C) is shown composed of 4 subunits along with the ‘RCK’ cytoplasmic regulatory region of the channel. (Martin et al., 2012)
SLO-1 K+ channels have large (~200 pS) single-channel conductances and are also known as ‘big’ potassium (BK) channels. These channels serve to clamp the membrane potential near the potassium reversal potential and inhibit electrical excitability. The SLO-1 K+ channel, has a different function to the high conductance Ca2+-dependent Cl− channel (Thorn and Martin, 1987) which is associated with the transport of carboxylic acids from anaerobic metabolism of glucose (Valkanov and Martin, 1995). Subunits homologous to the vertebrate SLO-1 K+ channels are found in C. elegans and Ascaris suum (Buxton, 2011). The SLO-1 K+ channels are composed of 4 α-subunits; α-subunits have seven (S0—S6) transmembrane regions, a P-loop between S5 and S6, a large intracellular domain (S7-S-10) and a well conserved ‘calcium bowl’ between domains S9 and S10, Fig. 2. In addition, the regulator of the K+ channel conductance (RCK) domains (S7, S8) contains high and low affinity calcium binding sites. The SLO-1 α-subunits show alternative splicing, producing channels with different calcium sensitivities; in C. elegans there are up to 15 splice variants (including SLO-1a, SLO-1b and SLO-1c (Kulke et al., 2014; Wang et al., 2001)). Each α-subunit of the channel has at least two high affinity calcium binding sites and one low affinity calcium/magnesium binding site. The channel also combines with secondary, regulatory β-subunits in vertebrates (Knaus et al., 1994) but these subunits have not yet been identified in C. elegans. However, Drosophila, which also lacks β subunits, has Slob proteins, which appear to carry out similar functions to the vertebrate β-subunits and these may be present in nematodes (Claridge-Chang et al., 2001).
The suggested function of the SLO-1 K+ channels is that they adjust the resting membrane potential of electrically excitable cells and adjust the level of excitability, up or down, and so affect the response to other inputs. The opening of the SLO-1 K+ channel, Fig. 2, is regulated by at least ten factors including: 1) membrane potential; 2) calcium; 3) magnesium; 4) NO; 5) CO; 6) arachidonic acid; 7) prostaglandins; 8) phosphorylation by cAMP-dependent protein kinase A; 9) phosphorylation by diacylgycerol/Ca2+-dependent protein kinase C; and 10) phosphorylation by cGMP-dependent protein kinase G (Salkoff et al., 2006). The kinases allow SLO-1 to be coupled to multiple and quite diverse signaling cascades permitting different ways of adjusting the excitability of the cells. The different nematode neuropeptides could affect cAMP and cGMP levels (e.g. AF1, AP2, and PF1 & PF2) could then affect SLO-1 channels (Muhlfeld et al., 2009; Verma et al., 2007, 2009). A selective agonist for the PF1 and/or a selective antagonist of the AF1 receptor would activate a SLO-1 K+-like current and increase the efficacy of emodepside. The neuropeptides AF1, AF10 and PF2 also bind with low affinity to the latrophilin-like receptor, HC110-R of Haemonchus contortus, suggesting that emodepside could also have an indirect mode of action on SLO-1 K+ channels through these latrophilin-like receptors (Fig.2) that are neuropeptide receptors (Muhlfeld et al., 2009).
We found (Buxton et al., 2011) that Asu-slo-1 is an evolutionarily conserved homologue of the slo-1 genes, expressed in adult A. suum body muscle flaps. Using a two-micro-electrode current-clamp and voltage-clamp technique, Fig 3 A, we found in the nematode parasite A. suum, that emodepside activates SLO-1-like K+ channels to produce hyperpolarization under current-clamp, Fig. 3 B1, and an outward current when under voltage-clamp, Fig. 3 B2. We also found that emodepside increased voltage-activated K+ currents, Fig. 3C D, in a time dependent manner. Emodepside, Fig. 3C, unlike PF1 did not decrease calcium currents and so emodepside does not work by releasing PF1 as has been hypothesized. We found that the effects of emodepside on the voltage-activated K+ channels is Ca2+-dependent and were inhibited by 5 mM 4-aminopyridine, Fig. 4A. The membrane hyperpolarization and increase in voltage-activated K+ current produced by emodepside, Fig. 3B, are very slow in onset and increase over a period of more than 10 minutes (Buxton et al. 2011); the slow onset effect of emodepside might be due to its very lipophilic nature and a membrane partitioning effect. It may also be because the effects of emodepside are indirect and produced by activation of a slow signaling cascade. Figs. 4 B & C shows that the effects of emodepside are potentiated by sodium nitroprusside (a NO donor), antagonized by iNOS inhibitors (NNLA), and inhibited by 1 µM staurosporine, an inhibitor of protein kinase (Buxton et al., 2011). Interestingly, these signaling molecules are known as activators of SLO-1 in other cells (Bolotina et al., 1994b; Holden-Dye et al., 2007; Mistry and Garland, 1998; Wang et al., 1999) and therefore encourage the view that emodepside could act through either or both of these signaling cascades and the signaling cascades may be in series or parallel, Fig 2C. A number of studies on the mammalian orthologues of SLO-1 show that they are directly and alternately regulated by complex, multiple signaling cascades, involving NO and diacylglyerol or PKC activation (Ghatta et al., 2006; Salkoff et al., 2006). It is pointed out that NO signaling pathways in mammals may not be conserved in all species of nematode.
Fig. 3.
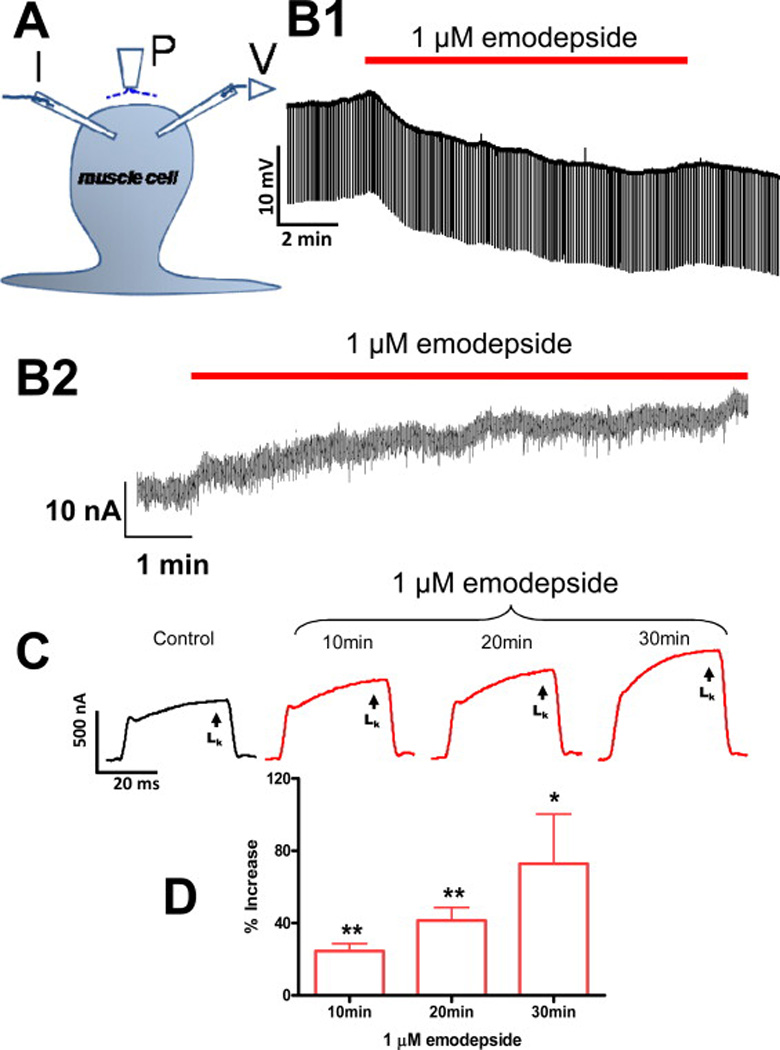
Electrophysiological techniques (two micropipette current-clamp and voltage-clamps) for recording from Ascaris suum. A: A. suum muscle bag showing the current (I) and voltage (V) micropipettes in the bag, and the perfusion needle (P). B1: Representative current-clamp trace showing the slow hyperpolarizing membrane potential during and after 10 min application of 1 µM emodepside. B2: Outward current response to 1 µM emodepside at higher time resolution. Holding potential −35 mV. Notice that emodepside produces a gradually increasing current after a delay of some 30 seconds. The response does not plateau in the time period of this recording. C: Voltage-clamp traces of control K+ current and the time-dependent effects of 1 µM emodepside on the K+ currents, all to a step potential of 0 mV from a holding potential of −35 mV. D: Bar chart (mean ± S.E.) of 1 µM emodepside effect on steady state (LK) currents. Comparison was made between the control 0 mV step current at 30 – 40 ms and the corresponding current increased by emodepside at 10, 20 and 30 min. Emodepside increased LK currents at 10 min (p < 0.01, n = 4, paired t-test), 20 min (p < 0.01, n = 4, paired t-test) and 30 min (p < 0.05, n = 4, paired t-test). (Martin et al., 2012)
Fig. 4.
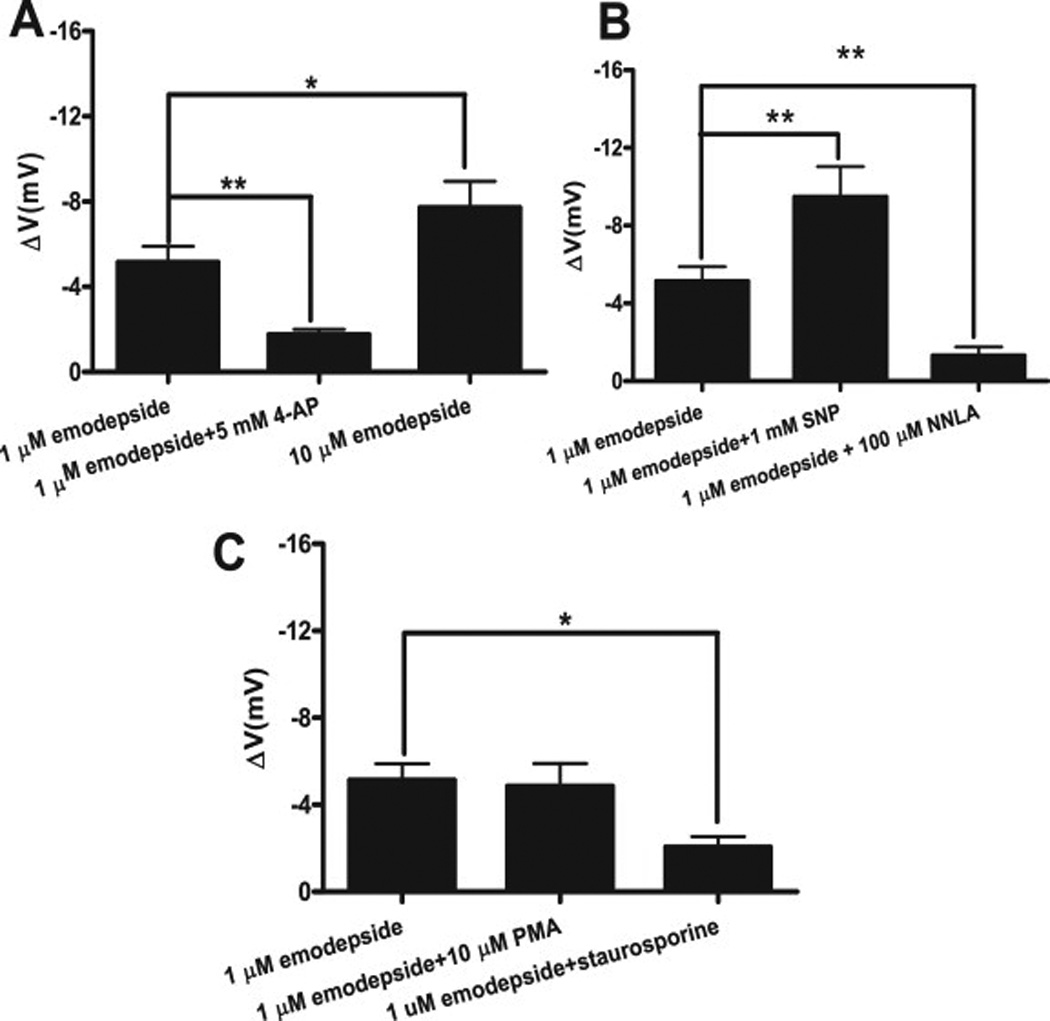
Effect of 1 µM emodepside on membrane potential. A: Comparison was made between membrane potential before and during emodepside application. 1 µM emodepside caused a significant membrane hyperpolarization (p < 0.001, n = 10, paired t-test) which was reduced in the presence of 5 mM 4-aminopyridine (p < 0.01, n = 8, unpaired t-test). 10 µM emodepside caused an increased hyperpolarization in comparison to 1 µM emodepside (p < 0.05, unpaired t-test). B Bar chart (mean ± s.e) of NO influence on emodepside-induced hyperpolarization. In the presence of 1 mM SNP, 1 µM emodepside caused an increased hyperpolarization (p < 0.01, n = 4, paired t-test). 100 µM NNLA decreased the hyperpolarization caused by 1 µM emodepside (p < 0.01, unpaired t-test). C: Bar chart (mean ± s.e) of effect of protein kinase modulators on emodepside-induced hyperpolarization. 10 µM PMA had no significant effects on the hyperpolarization caused by 1 µM emodepside. However, 1 µM staurosporine decreased the hyperpolarization caused by 1 µM emodepside (p < 0.05, unpaired t-test). (Martin et al., 2012)
Thus emodepside appears to selectively activate the SLO-1 K+ current that is present in nematodes but has less effect on the SLO-1 K+ currents of vertebrates. This can explain the selective toxicity of the drug. The existence and mechanisms of resistance to emodepside in nematode parasites remain to be studied but could involve several different mechanisms including changes in the splice forms of the SLO-1 K+ channel or its regulation by different signaling cascades. However, such resistance, depending on the specific mechanism, might be countered (Verma et al., 2007, 2009) by a selective agonist for the FMRFamide PF1 receptor and/or a selective antagonist of the FMRFamide AF1 receptor which would activate a SLO-1 K+-like current and increase the efficacy of emodepside. An increased knowledge of the physiology of target site of emodepside provides a new way of designing drug combinations. The complex nature of SLO-1 K+ regulation suggests that if there were resistance to emodepside it would be complex in nature and therefore likely to be polygenic involving multiple mechanisms.
3. Diethylcarbamazine
Diethylcarbamazine, Fig 5A, is an antifilarial drug that has been used since 1947 against lymphatic filariasis and loiasis. It is still an important and effective antifilarial drug but its mode of action is not fully described. Diethylcarbamazine has been suggested to have an indirect, host mediated mode of action: it appears to alter host arachidonic acid, nitric oxide metabolic pathways and inhibits NF-κB, which together in an unknown way leads to immobilization and sequestration of the microfilariae (Maizels and Denham, 1992; Peixoto and Silva, 2014). Diethylcarbamazine activity against B. malayi microfilariae is abolished in inducible nitric oxide synthase knockout mice (iNOS−/−), suggesting that diethylcarbamazine activity is dependent on host inducible nitric oxide synthase (iNOS) and nitric oxide (McGarry et al., 2005).
Fig 5.
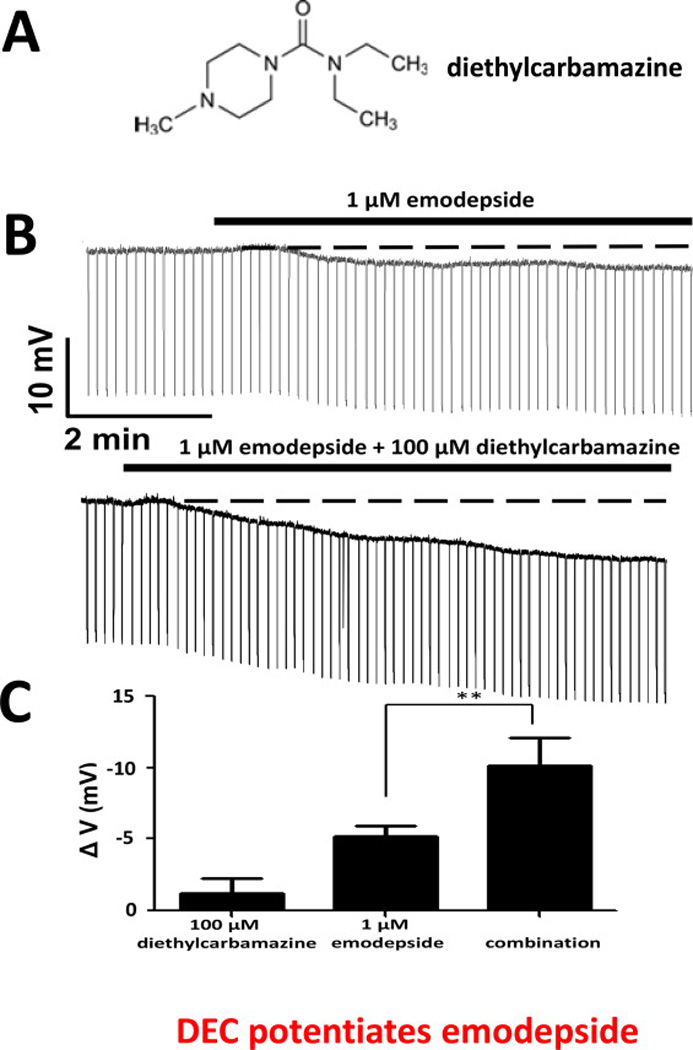
The combined effect of emodepside and diethylcarbamazine is greater than the effect of emodepside alone on A. suum membrane potential. A: Chemical structure of diethylcarbamazine. B: Representative current-clamp traces showing the membrane potential before, during 1 µM emodepside application (top trace) and during application of 1 µM emodepside plus 100 µM diethylcarbamazine (lower trace). The time delay between the end of the application of 100 µM diethylcarbamazine and 1 µM emodepside plus 100 µM diethylcarbamazine was 10 minutes. The change in membrane potential between the beginning of the 100 µM diethylcarbamazine and application of 1 µM emodepside plus 100 µM diethylcarbamazine was a hyperpolarization of 1.5 mV. C: Bar chart (mean ± i.e.) of diethylcarbamazine effect on emodepside-induced hyperpolarization. 100 µM diethylcarbamazine increased the hyperpolarization caused by 1 µM emodepside (p < 0.01, n = 5, unpaired t-test (Buxton et al., 2014a).
We were interested to determine how diethylcarbamazine would affect calcium-dependent SLO-1 K+ currents in isolated A. suum muscle flap preparations, and how diethylcarbamazine interacts with emodepside. The interest was prompted by observations in vertebrates (Bolotina et al., 1994a) which show that nitric oxide activates SLO-1 K+ channels and observations on A. suum indicating the presence of nitric oxide synthase and of SLO-1 K+ channels which show positive modulation by a nitric oxide pathway (Buxton et al., 2011). We hypothesized that diethylcarbamazine, with effects on arachidonic acid and nitric oxide pathways, would increase activation of SLO-1 K+ currents in A. suum muscle and potentiate effects of emodepside on membrane potential (Buxton et al., 2014a). We conducted current-clamp and voltage-clamp electrophysiological experiments, using A. suum body muscle in the presence of sufficient calcium to allow activation of the SLO-1 K+ currents. We found that indeed diethylcarbamazine, by itself, can increase activation of SLO-1 K+ currents and potentiate the hyperpolarizing effects of emodepside. This to us was very interesting since it showed that diethylcarbamazine has a direct effect on the nematode parasite and its effects are not exclusively mediated via the host as has been suggest by earlier experiments. It seems likely that the effects of diethylcarbamazine may involve effects on NO or arachidonic acid metabolites both in the host and in the parasite. Although the mechanism of action of diethylcarbamazine remains to be further defined, the synergistic effect of emodepside and diethylcarbamazine suggests that the combination of the two drugs could be considered for therapeutic use for the treatment of filarial nematode infections.
4. Tribendimidine
Tribendimidine, Fig. 6, is a symmetrical diamidine derivative, of amidantel which was developed in China for use in humans in the mid-1980s. It is a broad-spectrum anthelmintic effective against soil-transmitted helminthiasis including hookworm, pinworms, roundworms, Strongyloides and flatworms (Xiao et al., 2005; Xu et al., 2014) of humans. Molecular studies (Hu et al., 2009) on C. elegans using null-mutants of the levamisole receptor subunits strongly suggested that tribendimidine is a cholinergic agonist that is selective for the same nematode muscle nAChR as levamisole. When we looked at these studies we found that there are no direct electrophysiological observations in nematode parasites that had been made to test this hypothesis and we were aware that sometimes observations on the model Clade V nematode were not always the same in parasitic nematodes, particularly for the parasites from a different Clade. We were also interested in trying to explain why tribendimidine is effective against some nematode parasites when levamisole is not.
Fig. 6.
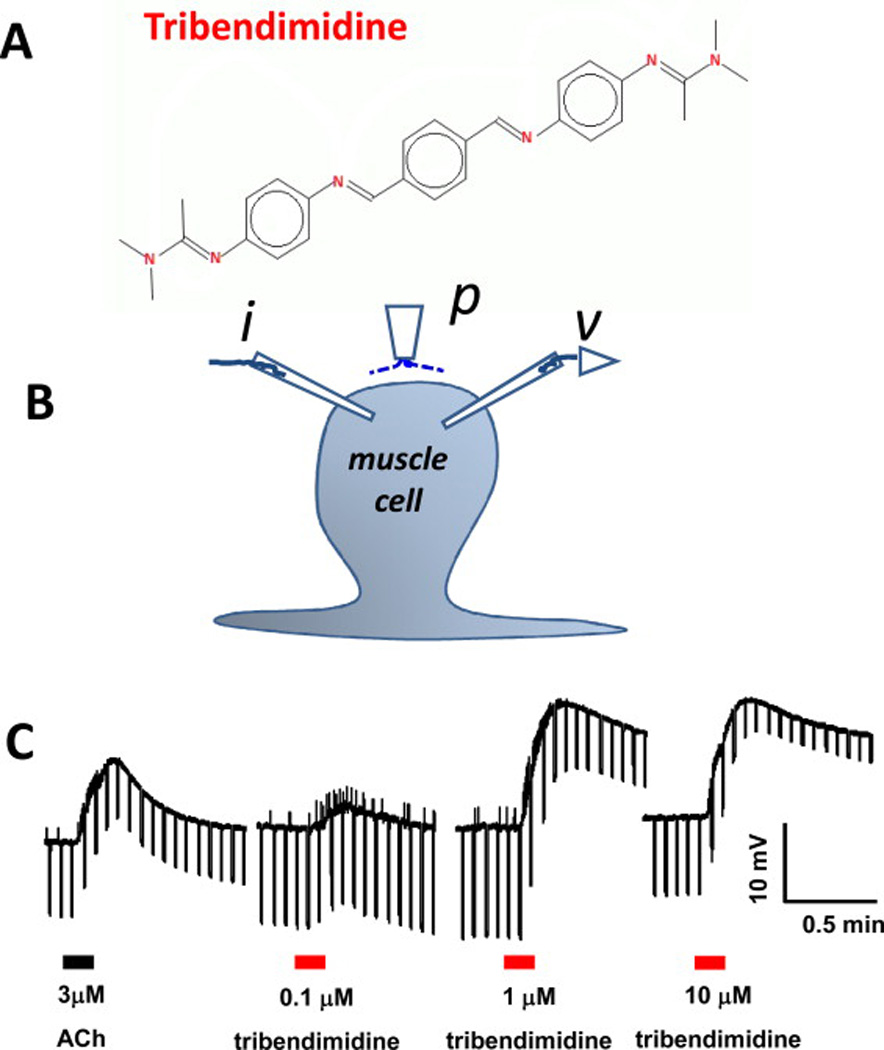
A: Chemical structure of tribendimidine. B: Diagram of the two-micropipette current-clamp technique used to record the membrane potential (v) and to inject 40 nA hyperpolarizing 500 ms current pulses (i) at 0.3 Hz. p is the microperfusion pipette used to apply and wash off the drugs. C: Application of 3 µM acetylcholine and then 0.1, 1 and 10 µM tribendimidine to the same preparation. 1 µM tribendimidine produces a bigger depolarization response (upward movement) and conductance increase (reduction in the voltage responses to current injection, producing a narrowing of the width of the trace) than 3 µM acetylcholine (Robertson et al., 2015).
We tested the effects of tribendimidine on the electrophysiology, Fig. 6, and contraction of A. suum muscle (Robertson et al., 2015) and found that tribendimidine produces a dose-dependent depolarization associated with an increase in the conductance of the muscle membrane as the nicotinic receptors (nAChRs) open, Fig 6. We found that tribendimidine was more potent than acetylcholine and had an EC50 of 0.8µM. We also tested the effects of the nicotinic antagonist mecamylamine, a potent antagonist of muscle nAChRs of parasitic nematodes and found that 3µM was potent against tribendimidine; the antagonist effects of mecamylamine confirmed the cholinergic action of tribendimidine on the nAChRs of the parasite A. suum.
We characterized the pharmacological profile of tribendimidine using our Ascaris muscle contraction assay systems (Qian et al., 2006; Robertson et al., 2002) with tribendimidine as the agonist and methyllycaconitine, paraherquamide and derquantel as antagonists to calculate the potency, pA2, of the antagonists and the subtype of the selectivity of tribendimidine. We found that tribendimidine was more selective for the B-subytpe (bephenium preferring) nAChRs than the L-subtype (levamisole preferring) or the N-subtype (nicotine preferring). These observations showed the selectivity for the different groups of nAChRs present on the Ascaris muscle was not the same as levamisole and it was possible that levamisole-resistant parasites would remain sensitive to tribendimidine. To test this we used, Oesophagostomum dentatum L3 larval migration inhibition assays with levamisole-sensitive isolates and levamisole-resistant isolates and tested the effects of tribendimidine on motility. Levamisole was less effective (p< 0.001, F-test) in inhibiting migration of levamisole-resistant larvae than the migration of levamisole-sensitive larvae. We tested the effects of tribendimidine to the limits of its solubility, ~30 µM, and found that tribendimidine was actually more potent on the levamisole-resistant isolate than on the levamisole-sensitive isolates (p < 0.001, F-test). The larger effect of tribendimidine on the levamisole-resistant, than the levamisole-sensitive isolates at lower tribendimidine concentrations, supports the view that levamisole and tribendimidine do not activate identical nAChR receptor subtypes. The higher efficacy of tribendimidine on the levamisole resistant O. dentatum isolate might indicate a case of negative cross resistance (Miltsch et al., 2013)
We have been able to express, in Xenopus oocytes a range of different nAChR receptor subtypes derived from anthelmintic sensitive Oesophagostomum dentatum clones of their nAChR receptor subunits (Buxton et al., 2014b). Fig. 7 shows a diagram of the subunit composition of two nAChR subtypes expressed in oocytes, one composed of Ode-UNC-29:Ode-UNC-63:Ode-UNC-38, the other composed of Ode-UNC-29:Ode-UNC-63:Ode-UNC-38:Ode-ACR-8 subunits. The Ode-UNC-29:Ode-UNC-63:Ode-UNC-38 receptor was more sensitive to tribendimidine and pyrantel but not to levamisole or nicotine. We can see that the receptor subunit arrangements affect their pharmacology and that not all cholinergic anthelmintics have the same selectivity.
Fig. 7.
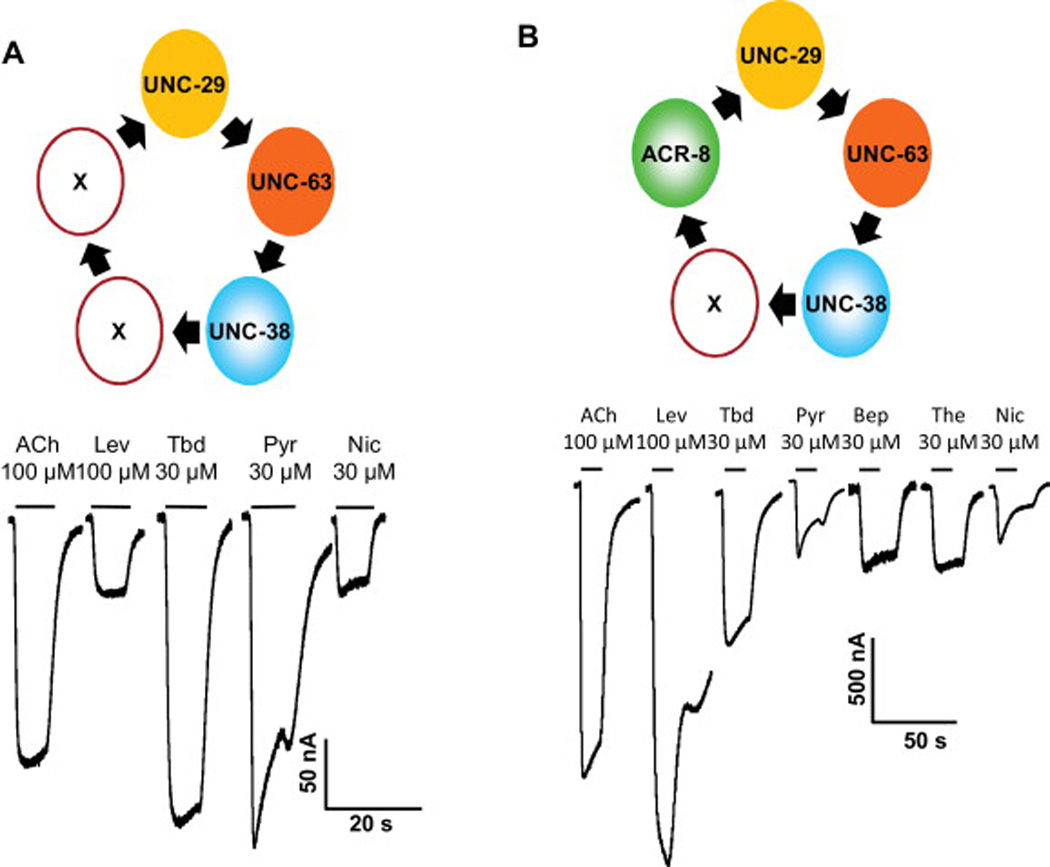
Figure 4. Tribendimidine as an agonist of Ode-(29 – 8 – 38 – 63) and Ode-(29-63-38) receptors expressed in oocytes, currents recorded under voltage-clamp at −60 mV (Buxton et al., 2014b) A: Tribendimidine and pyrantel at 100 µM is more potent than levamisole on the Ode(29 – 38 – 63) receptor. B: Levamisole at 100 µM is more potent than acetylcholine tribendimidine, pyrantel and nicotine on the expressed Ode(29 – 8 – 38 – 63) receptor. Note that derquantel produces competitive antagonism of the effects of levamisole but a non-competitive antagonism of pyrantel, suggesting that the mode of action of levamisole and pyrantel are not identical.
Our understanding to date is that there are a range of pharmacologically different nAChR subtypes present in parasitic nematodes and the pharmacology of the receptor subtypes varies with the subunit composition of the receptor which varies with the tissue and species of the nematode parasite. There will therefore be cholinergic anthelmintics that act on different nAChR subtypes and it is therefore possible that there will not always be cross-resistance between the cholinergic anthelmintics.
5. Discussion
Diethylcarbamazine is used mostly for treatment of filariasis in humans but has effects against hookworm and ascariasis, intestinal nematode parasites (Meyrowitsch and Simonsen, 2001). In addition to the treatment of filariasis, diethylcarbamazine, as a single dose treatment, has modest effects on intestinal nematode parasite infections including ascariasis and trichuriasis but is more effective when combined with ivermectin or albendazole (Belizario et al., 2003). We have seen in our experiments that emodepside increases a Slo-1 K+ current in the parasite and that diethylcarbamazine can potentiate the effect of emodepside. Both of these compounds have been used separately as single anthelmintics but we do not have data on their therapeutic effect when used in combination. This information is desirable to obtain and if therapeutic combination of emodepside and diethylcarbamazine is safe and more potent than either compound in isolation, such synergism could slow the development of resistance and extend the spectrum of action of the therapy.
Levamisole and tribendimidine have different selectivities for nAChR subtypes. In Ascaris tribendimidine is more selective for the B-subtype rather than the L-subtype of nAChR. Tribendimidine is more potent on expressed O. dentatum receptor subtypes composed of Ode-UNC-29:Ode-UNC-63:Ode-UNC-38 subunits, than on subtypes composed of Ode-UNC-29:Ode-UNC-63:Ode-UNC-38:Ode-ACR-8 subunits (Buxton et al., 2014b). The difference in selectivity suggests that tribendimidine has the potential to be effective against nematode parasites that are not sensitive to levamisole, including those parasites that have developed resistance.. A careful combination of different cholinergic anthelmintics has the potential to extend the spectrum of action of the drug treatment. Tribendimidine has an interesting and promising pharmacology and has the potential for single-dose MDA with its broad-spectrum of action. Although tribendimidine appears safe and has broad-spectrum activity, a large-scale clinical study is advocated to further verify human safety.
6. Conclusion
We have been excited by our discovery of further details of the modes of action of the different anthelmintics that we work on and believe that the information will be useful for defining mechanisms of resistance to the anthelmintic drugs that we have studied. We also think that the improved understanding of their mechanisms of action will lead to better use of these compounds and logical design of synergistic combinations. We do not look back, but forward, to the future.
Highlights.
The best way to predict the future is to create it.
Mode of action of anthelmintics required:
for rational design of synergistic anthelmintic combinations;
for slowing down the development of anthelmintic resistance.
Recent observations on emodepside, diethylcarbamazine & tribendimidine described.
Acknowledgments
Funding: The research project culminating in this paper was funded by the National Institute of Allergy and Infectious Diseases (NIH) grant R 01 A1 047194 to RJM and R21 AI092185 to APR. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Belizario VY, Amarillo ME, De Leon WU, De los Reyes AE, Bugayong MG, Macatangay BJC. A comparison of the efficacy of single doses of albendazole, ivermectin, and diethylcarbamazine alone or in combinations against Ascaris and Trichuris spp. Bull World Health Organ.: Int J. Public Health. 2003;81(1):35–42. [PMC free article] [PubMed] [Google Scholar]
- Bolotina V, Najibi S, Palacino J, Pagano P, Cohen R. Nitric oxide directly activated calcium-dependent potassium channels in vascular smooth muscle. Nature. 1994a;368(6474):850–853. doi: 10.1038/368850a0. [DOI] [PubMed] [Google Scholar]
- Bolotina VM, Najibi S, Palacino JJ, Pagano PJ, Cohen RA. Nitric oxide directly activates calcium-dependent potassium channels in vascular smooth muscle. Nature. 1994b;368:850–853. doi: 10.1038/368850a0. [DOI] [PubMed] [Google Scholar]
- Buxton S, Neveu C, Charvet C, Robertson AP, Martin RJ. On the mode of action of emodepside: slow effects on membrane potential and voltage-activated currents in Ascaris suum. British Journal of Pharmacology. 2011;164:453–470. doi: 10.1111/j.1476-5381.2011.01428.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Buxton S, Robertson A, Martin R. Diethylcarbamazine Increases Activation of Voltage-Activated Potassium (SLO-1) Currents in Ascaris suum and Potentiates Effects of Emodepside. Plos Neglected Tropical Diseases. 2014a;8 doi: 10.1371/journal.pntd.0003276. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Buxton SK, Charvet CL, Neveu C, Cabaret J, Cortet J, Peineau N, Abongwa M, Courtot E, Robertson AP, Martin RJ. Investigation of acetylcholine receptor diversity in a nematode parasite leads to characterization of tribendimidine- and derquantel-sensitive nAChRs. PLoS Pathog. 2014b;10:e1003870. doi: 10.1371/journal.ppat.1003870. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Buxton SK, Robertson AP, Martin RJ. On the mode of action of emodepside: Effects on membrane potential and voltage-activated currents in Ascaris suum. 2011 doi: 10.1111/j.1476-5381.2011.01428.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Claridge-Chang A, Wijnen H, Naef F, Boothroyd C, Rajewsky N, Young MW. Circadian regulation of gene expression systems in the Drosophila head. Neuron. 2001;32:657–671. doi: 10.1016/s0896-6273(01)00515-3. [DOI] [PubMed] [Google Scholar]
- Crisford C, Marray C, O'Connor V, Edwards RJ, Kruger N, Samson-Himmelstjerna Gv, Harder A, Walker RJ, Holden-Dye L. Selective Toxicity of the Anthelmintic Emodepside Revealed by Heterologous Expression of Human KCNMA1 in Caenorhabditis elegans. Molecular Pharmacology. 2011 doi: 10.1124/mol.111.071043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- DiMasi J, Hansen R, Grabowski H. The price of innovation: new estimates of drug development costs. Journal of Health Economics. 2003;22:151–185. doi: 10.1016/S0167-6296(02)00126-1. [DOI] [PubMed] [Google Scholar]
- Evans T, Chapple N. The animal health market. Nature Reviews Drug Discovery. 2002;1:937–938. doi: 10.1038/nrd975. [DOI] [PubMed] [Google Scholar]
- Ghatta S, Nimmagadda D, Xu X, O'Rourke ST. Large-conductance, calcium-activated potassium channels: Structural and functional implications. Pharmacology & Therapeutics. 2006;110:103–116. doi: 10.1016/j.pharmthera.2005.10.007. [DOI] [PubMed] [Google Scholar]
- Gilleard J, Beech R. Population genetics of anthelmintic resistance in parasitic nematodes. Parasitology. 2007;134:1133–1147. doi: 10.1017/S0031182007000066. [DOI] [PubMed] [Google Scholar]
- Guest M, Bull K, Walker RJ, Amliwala K, O'Connor V, Harder A, Holden-Dye L, Hopper NA. The calcium-activated potassium channel, SLO-1, is required for the action of the novel cyclo-octadepsipeptide anthelmintic, emodepside, in Caenorhabditis elegans. Int. J. Parasitol. 2007 doi: 10.1016/j.ijpara.2007.05.006. [DOI] [PubMed] [Google Scholar]
- Holden-Dye L, O'Connor V, Hopper NA, Walker RJ, Harder A, Bull K, Guest M. SLO, SLO, quick, quick, slow: calcium-activated potassium channels as regulators of Caenorhabditis elegans behaviour and targets for anthelmintics. Invert Neurosci. 2007;7:199–208. doi: 10.1007/s10158-007-0057-z. [DOI] [PubMed] [Google Scholar]
- Hotez PJ, Brindley PJ, Bethony JM, King CH, Pearce EJ, Jacobson J. Helminth infections: the great neglected tropical diseases. J Clin Invest. 2008;118:1311–1321. doi: 10.1172/JCI34261. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hu Y, Xiao SH, Aroian RV. The new anthelmintic tribendimidine is an L-type (levamisole and pyrantel) nicotinic acetylcholine receptor agonist. PLoS Negl Trop Dis. 2009;3:e499. doi: 10.1371/journal.pntd.0000499. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Knaus HG, Garcia-Calvo M, Kaczorowski GJ, Garcia ML. Subunit composition of the high conductance calcium-activated potassium channel from smooth muscle, a representative of the mSlo and slowpoke family of potassium channels. J Biol Chem. 1994;269:3921–3924. [PubMed] [Google Scholar]
- Kulke D, von Samson-Himmelstjerna G, Miltsch S, Wolstenholme A, Jex A, Gasser R, Ballesteros C, Geary T, Keiser J, Townson S, Harder A, Krucken J. Characterization of the Ca2+-Gated and Voltage-Dependent K+-Channel Slo-1 of Nematodes and Its Interaction with Emodepside. Plos Neglected Tropical Diseases. 2014;8 doi: 10.1371/journal.pntd.0003401. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lanusse C, Alvarez L, Lifschitz A. Pharmacological knowledge and sustainable anthelmintic therapy in ruminants. Veterinary Parasitology. 2014;204:18–33. doi: 10.1016/j.vetpar.2013.11.003. [DOI] [PubMed] [Google Scholar]
- Maizels R, Denham D. Diethylcarbamazine (DEC): immunopharmacological interactions of an anti-filarial drug. Parasitology. 1992;105(Suppl):S49–S60. doi: 10.1017/s0031182000075351. [DOI] [PubMed] [Google Scholar]
- Martin R, Buxton S, Neveu C, Charvet C, Robertson A. Emodepside and SL0-1 potassium channels: A review. Experimental Parasitology. 2012;132:40–46. doi: 10.1016/j.exppara.2011.08.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martin RJ, Harder A, Londerhausen M, Jenschke P. Anthelmintic actions of the cyclic depsipeptide PF1022A and its electrophysiological effects on muscle cells of Ascaris suum. Pesticide Science. 1996a;38:343–349. [Google Scholar]
- Martin RJ, Valkanov MA, Dale VM, Robertson AP, Murray I. Electrophysiology of Ascaris muscle and anti-nematodal drug action. Parasitology. 1996b;113(Suppl):S137–S156. doi: 10.1017/s0031182000077945. [DOI] [PubMed] [Google Scholar]
- McGarry HF, Plant LD, Taylor MJ. Diethylcarbamazine activity against Brugia malayi microfilariae is dependent on inducible nitric-oxide synthase and the cyclooxygenase pathway. Filaria Journal. 2005;4:4. doi: 10.1186/1475-2883-4-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meyrowitsch D, Simonsen P. Efficacy of DEC against Ascaris and hookworm infections in schoolchildren. Tropical Medicine & International Health. 2001;6:739–742. doi: 10.1046/j.1365-3156.2001.00766.x. [DOI] [PubMed] [Google Scholar]
- Miltsch S, Krucken J, Demeler J, Ramunke S, Harder A, von Samson-Himmelstjerna G. Interactions of anthelmintic drugs in Caenorhabditis elegans neuro-muscular ion channel mutants. Parasitology International. 2013;62:591–598. doi: 10.1016/j.parint.2013.05.006. [DOI] [PubMed] [Google Scholar]
- Mistry DK, Garland CJ. Nitric Oxide (NO)-induced activation of large conductance Ca2+-dependent K+ channels (BKCa) in smooth muscle cells isolated from the rat mesenteric artery. British Journal of Pharmacology. 1998;124:1131–1140. doi: 10.1038/sj.bjp.0701940. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Muhlfeld S, Schmitt-Wrede H, Harder A, Wunderlich F. FMRFamide-like neuropeptides as putative ligands of the latrophilin-like HC110-R from Haemonchus contortus. Molecular and Biochemical Parasitology. 2009;164:162–164. doi: 10.1016/j.molbiopara.2008.12.003. [DOI] [PubMed] [Google Scholar]
- Peixoto C, Silva B. Anti-inflammatory effects of diethylcarbamazine: A review. European Journal of Pharmacology. 2014;734:35–41. doi: 10.1016/j.ejphar.2014.03.046. [DOI] [PubMed] [Google Scholar]
- Perutz M. IN PRAISE OF IMPERFECTION - MY LIFE AND WORK - LEVIMONTALCINI, R. Nature. 1988;332:791–791. [Google Scholar]
- Qian H, Martin RJ, Robertson AP. Pharmacology of N-, L- and B-subtypes of nematode nAChR resolved at the single-channel level in Ascaris suum. FASEB J. 2006 doi: 10.1096/fj.06-6264fje. [DOI] [PubMed] [Google Scholar]
- Robertson AP, Clark CL, Burns TA, Thompson DP, Geary TG, Trailovic SM, Martin RJ. Paraherquamide and 2-deoxy-paraherquamide distinguish cholinergic receptor subtypes in Ascaris muscle. J. Pharmacol. Exp. Ther. 2002;302:853–860. doi: 10.1124/jpet.102.034272. [DOI] [PubMed] [Google Scholar]
- Robertson AP, Puttachary S, Buxton SK, Martin RJ. Tribendimidine: Mode of Action and nAChR Subtype Selectivity in Ascaris and Oesophagostomum. In PLOS Neglected Tropical Diseases. 2015 doi: 10.1371/journal.pntd.0003495. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saeger B, Schmitt-Wrede HP, Dehnhardt M, Benten WP, Krucken J, Harder A, Von Samson-Himmelstjerna G, Wiegand H, Wunderlich F. Latrophilin-like receptor from the parasitic nematode Haemonchus contortus as target for the anthelmintic depsipeptide PF1022A. FASEB J. 2001;15:1332–1334. doi: 10.1096/fj.00-0664fje. [DOI] [PubMed] [Google Scholar]
- Salkoff L, Butler A, Ferreira G, Santi C, Wei A. High-conductance potassium channels of the SLO family. Nature Reviews Neuroscience. 2006;5:921–931. doi: 10.1038/nrn1992. [DOI] [PubMed] [Google Scholar]
- Thorn P, Martin RJ. A high-conductance calcium-dependent chloride channel in Ascaris suum muscle. Q. J. Exp. Physiol. 1987;72:31–49. doi: 10.1113/expphysiol.1987.sp003053. [DOI] [PubMed] [Google Scholar]
- Valkanov MA, Martin RJ. A Cl channel in Ascaris suum selectivity conducts dicarboxylic anion product of glucose fermentation and suggests a role in removal of waste organic anions. J. Membr. Biol. 1995;148:41–49. doi: 10.1007/BF00234154. [DOI] [PubMed] [Google Scholar]
- Verma S, Robertson AP, Martin RJ. The nematode neuropeptide, AF2 (KHEYLRF-NH(2)), increases voltage-activated calcium currents in Ascaris suum muscle. Br. J. Pharmacol. 2007;151:888–899. doi: 10.1038/sj.bjp.0707296. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Verma S, Robertson AP, Martin RJ. Effects of SDPNFLRF-amide (PF1) on voltage-activated currents in Ascaris suum muscle. Int J Parasitol. 2009;39:315–326. doi: 10.1016/j.ijpara.2008.07.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang J, Zhou Y, Wen H, Levitan IB. Simultaneous binding of two protein kinases to a calcium-dependent potassium channel. J Neurosci. 1999;19:1–7. doi: 10.1523/JNEUROSCI.19-10-j0005.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang ZW, Saifee O, Nonet ML, Salkoff L. SLO-1 potassium channels control quantal content of neurotransmitter release at the C. elegans neuromuscular junction. Neuron. 2001;32:867–881. doi: 10.1016/s0896-6273(01)00522-0. [DOI] [PubMed] [Google Scholar]
- Willson J, Amliwala K, Davis A, Cook A, Cuttle MF, Kriek N, Hopper NA, O'Connor V, Harder A, Walker RJ, Holden-Dye L. Latrotoxin receptor signaling engages the UNC-13-dependent vesicle-priming pathway in C. elegans. Curr. Biol. 2004;14:1374–1379. doi: 10.1016/j.cub.2004.07.056. [DOI] [PubMed] [Google Scholar]
- Xiao SH, Hui-Ming W, Tanner M, Utzinger J, Chong W. Tribendimidine: a promising, safe and broad-spectrum anthelmintic agent from China. Acta Trop. 2005;94:1–14. doi: 10.1016/j.actatropica.2005.01.013. [DOI] [PubMed] [Google Scholar]
- Xu L, Jiang B, Duan J, Zhuang S, Liu Y, Zhu S, Zhang L, Zhang H, Xiao S, Zhou X. Efficacy and Safety of Praziquantel, Tribendimidine and Mebendazole in Patients with Co-infection of Clonorchis sinensis and Other Helminths. Plos Neglected Tropical Diseases. 2014;8 doi: 10.1371/journal.pntd.0003046. [DOI] [PMC free article] [PubMed] [Google Scholar]