

Abstract
We determined the vascular and airway effects of PGF2α and its mechanism of action on isolated-perfused lungs of rats were isolated and perfused at 50 ml/kg/min with Krebs-Henseleit bicarbonate buffer solution containing 3% bovine serum albumin. The lungs were ventilated with 21% O2 and 5% CO2 at a tidal volume of 2 ml, frequency of 60 per minute and positive end expiratory pressure of 3 cmH2O. Following injection of 50 μg PGF2α into the afferent pulmonary catheter, there was a marked rise in pulmonary arterial pressure (Ppa) and in resistance to airflow across the lung (RL) and a fall in dynamic lung compliance (Cdyn). Double vascular occlusion technique revealed that 29% of the rise in Ppa was due to an increase in upstream and 71% to downstream resistance. Nω-nitro-L-arginine. 100 μm, a NO synthase inhibitor potentiated the Ppa response two-fold with significant change in airway mechanics. Rat atrial natriuretic factor (r-ANF), 40 μg, quickly reversed the changes in Ppa, RL and Cdyn. Infusion of r-ANF prior to PGF2α attenuated the Ppa response by 38%, RL by 44% and Cdyn by 12%. SQ 29548, a thromboxane receptor blocker and Cl, a protein kinase C (PKC) inhibitor, fully blocked both the vascular and airway responses to PGF2α. PGF2α is a constrictor of pulmonary vessels and airways in rat lungs via thromboxane SQ 29548 receptors, thansduced by intracellular PKC.
Keywords: Pulmonary Circulation, Prostaglandins, Vasoconstriction, Bronchoconstriction
INTRODUCTION
Prostanoids exert a variety of actions in the lung and have been implicated in the pathogenesis of a number of pulmonary diseases. Of the biological actions of prostanoids, the best documented are their effects on smooth muscles. PGF2α is one of the most frequently studied cyclooxygenase-derived prostanoid for its contractile action on the smooth muscle. Lung is one of the major sites of prostanoid synthesis and degradation. PGF2α causes bronchoconstriction and pulmonary vasoconstriction in vivo in all species of animals studied1). However, in vitro, the action of PGF2α is not uniform among different species of animals. PGF2α contracts trachea in man, both the trachea and parenchymal strips in the guinea pig, parenchymal strips only in dogs, with no effect on trachea. Data in rats, one of the most widely utilized species of laboratory animals, are limited to vascular effects of PGF2α2–4), although the site of vasoconstriction is unknown. Little is known about the effect of PGF2α on airway mechanics in rats.
The purpose of this study was to determine both the vascular and airway effects of PGF2α and its mechanism of action in isolated rat lungs.
MATERIAL AND METHODS
1. Animal Preparation
Sprague-Dawley rats weighing 250–400g were used. Rats were anesthetized by intraperitoneal injection of thiamylal sodium (150 mg/kg). After tracheostomy and the insertion of a tracheal cannula, the animals were ventilated with a Harvard respirator (Model 683). The respirator was set at a tidal volume (VT) of 2 ml and a breathing rate of 60 per minute. The lungs were hyperinflated at 20 cmH2O PEEP by a water seal to prevent atelectasis. Ventilatory gas concentrations were 21% O2 and 5% CO2, and 74% N2. The chest was opened by midline incision, followed by an intracardiac injection of heparin (1,000 IU/kg). The lungs were initially perfused in an open, non-recirculating manner for 10 minutes and the residual pulmonary blood was slowly washed out with 50 ml of Krebs-Henseleit bicarbonate buffer solution containing 3% bovine serum albumin at 37.5°C. The pulmonary artery was cannulated through an incision in the right ventricle and the cannula containing a bubble trap ws secured in place with a ligature around both the pulmonary artery and aorta. A cannula was also placed in the left ventricle and secured in place with a silk ligature. The heart and lungs were then removed from the thoracic cavity and suspended in a humidified. water-jacketed chamber at 39°C.
Pulmonary arterial and left ventricular catheters were connected to Validyne pressure transducers to continuously record inflow and outflow pressure on a strip chart recorder (Astro-Med Model MT 9500). The perfusion chamber (reservoir) had a volume of 50 ml and was maintained at 37–39°C. The height of the reservoir was adjusted to −4 cmH2O relative to the left atrium. Perfusion was then changed to a recirculating mode. The cannulas were manipulated until resistance to flow was less than 1 mmHg/ml per minute.
Perfusion rate was advanced in small steps to 0.05 ml/gm per minute and then held constant throughout the experiment using a Masterflex peristaltic pump5). During studies, samples of effluent perfusate were collected from the left ventricular cannula for determination of blood gases and pH. Partial pressure of CO2 was maintained in the range of 30–35 mmHg and pH at 7.3–7.4.
Proximal airway opening pressure (Pao) was measured via a side port in the tracheal cannula. Tidal airflow was continuously monitored by measuring the pressure difference across a heated pneumotachograph (Hans Rudolph, Model 8300) connected to the tracheal cannula. All pressure signals were measured with Validyne differential pressure transducers. Airflow was calibrated by using a rotameter. VT was Obtained by electrical integration of the flow signal by using a Buxco Pulmonary Mechanics Computer (Model 6). The latter also computed on a breath-by-breath basis the dynamic lung compliance (Cdyn)and the resistance to airflow across the lung (RL) from the input signals of volume (VT), pressure (Pao) and flow. Cdyn was calculated as VT divided by Pao at ponits of zero flow and expressed as milliliters per centimeter of water. RL was calculated as the difference between the inspiratory and expiratory Pao at mid-VT divided by the difference between the inspiratory and expiratory flow at mid-VT6). RL was expressed as centimeters of water per milliliter per second. RL measures total pulmonary resistance, of which approximately 80% is airway resistance and 20% is tissue viscous resistance. Pao, flow, VT, Cdyn and RL were all recorded continuously on the strip chart recorder.
Before experiments were started, a 30-minute equilibration perfusion period was allowed to establish stable values of pulmonary arterial pressure, dynamic lung compliance and lung resistance to airflow. Based on our previous experiments7), a single dose of 50 μg of PGF2α was used in all experiments. This dose of PGF2α injected into the afferent pulmonary arterial catheter produced maximal vascular and airway responses in the isolated-perfused lungs of rats. The following procedures were carried out with a minimum interval of 10 minutes between the procedures or until the vascular and airway responses returned to the baseline values following a particular procedure.
2. Effect of PGF2α on Ppa and Microvascular Pressure
To obtain an estimate of the microvascular pressure of the experimental animals, a double vascular occlusion technique8,9) was applied in 15 rat lungs at the end of exhalation by simultaneously occluding the pulmonary artery and the left atrial catheters and turning off the ventilatory pump. The vascular occlusion technique was applied first under stable baseline conditions and then at the peak Ppa response to PGF2α
3. Effects of Different Vasoactive and Blocking Agents on Vascular and Airway Responses to PGF2α
Atrial Natriuretic Factor (ANF)
ANF is a potent short-lived peptide pulmonary vasodilator that we used to determine the reversibility of the contractile response to PGF2α10,11). Two sets of experiments were performed with ANF, with each set containing 6 rat lungs. In one set, 40 μg of r-ANF (r-ANF, 28 amino acids: Peninsula Laboratories, Belmont, CA) were added when the Ppa reached its peak after the injection of 50 μg of PGF2α. In the second set, 40 μg of rat ANF were injected into the pulmonary artery chatheter 2 minutes before the injection of 50 μg of PGF2α. The dose of ANF chosen was based on our previous experiments for maximal response15).
Nω-nitro-L-arginine(NNLA)
In 12 rat lungs, following two consecutive doses of 50 μg of PGF2α. NNLA, a nitric oxide synthase inhibitor was added to the perfusate at a final concentration of 100 μM12,13). Three minutes later, another dose of 50 μg of PGF2α was injected into the pulmonary artery catherter.
Thromboxane (TX) Receptor Inhibitor, SQ 29548
This compound was provided by Squibb. 50 μg of PGF2α was injected into the pulmonary arterial catheter of 4 rat lungs before and after the addition of SQ 29548 to the perfusate at a final concentration of 40 μM7,14)
1-(5-isoquinolinesulfonyl)Piperazine (Cl)
50 μg of PGF2α were injected into the pulmonary arterial catheter of 4 rat lungs before and after the addition in the perfusate of 0.2 μM of Cl, an inhibitor of protein kinase C15,16).
4. Statistical Analyses
Means and standard errors of mean are presented for values obtained in each series of experiments. The statistical significance of diffenrences among the means was analyzed by Student’s t-test and paired t-test where applicable.
RESULTS
1. Effects of PGF2α on Microvascular Pressure
PGF2α caused significant vascular and airway responses in isolated rat lungs. The results of double occlusion studies are shown in Fig. 1. Ppa rose from 8.6±0.3 to 14.2±0.6 cmH2O after 50 μg of PGF2α (p<0.01) and microvascular pressure (Pmv) increased from 2.7 ± 0.3 to 6.7 ± 0.5 cmH2O (p<0.01). Of the rise in Ppa, 29% was due to an increase in upstream and 71% due to an increase in downstream resistance. Venous resistance could not be calculated, because the lungs were under zone II conditions (outflow pressure less than alveolar pressure). Yet, following the application of the double vascular occlusion technique, pulmonary arterial pressure dropped and left atrial pressure rose to the identical value, giving an estimate of the microvascular pressure. Following 50 μg PGF2α, RL increased from baseline value of 0.21±0.02 to 0.32+0.04 cmH2O/ml/sec (p<0.01) and Cdyn declined from 0.29±0.02 to 0.22±0.01 ml/cmH2O (p<0.01).
Fig. 1.
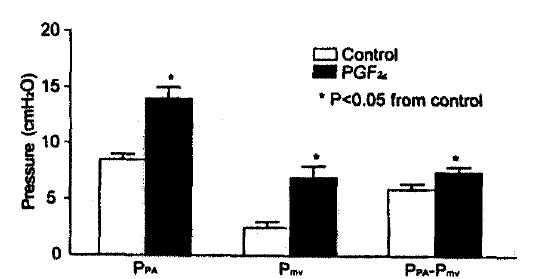
Change in longitudinal vascular segment pressure to a bolus dose of 50 μg PGF2α. in 15 isolated-perfused rat lungs, measured by double occlusion. PPA, pulmonary arterial pressure: PmV, microvascular pressure: PPA-PmV, difference. Most of the pressure change was in the downstream segment.
2. Effects of ANF on Vascular and Airway Responses
Rat atrial natriuretic factor quickly reserved the changes in Ppa, Cdyn and RL induced by PGF2α. When 40 μg of r-ANF were injected at the peak Ppa rise after 50 μg of PGF2α in 6 rat lungs, the half time for the decline of Ppa to the baseline was 0.3 min as opposed to 4 min when no r-ANF was added (Table 1). The return time to baseline Ppa was shortened by 73%. Injection of r-ANF at peak response caused a 78% recovery in RL and 48% recovery in Cdyn within one minute (Table 2). Injection of 40 μ or r-ANF in another set of 6 rat lungs before the injection of 50 μg PGF2α caused a 38% reduction in peak Ppa rise, a 44% reduction in the rise in RL and a 12% reduction in the decline in Cdyn, compared with the pre-ANF injection response to 50 μg PGF2α in the same rat lung (Fig. 2). The ANF effect was greater on RL than on Cdyn.
Table 1.
ANF Caused a More Rapid Decay of Pulmonary Arterial Pressure Following PGF2α
| CONTROL | ANF | |
|---|---|---|
| Basal PPA (cm H2O) | 8.7±0.3 | 8.3±0.3 |
| Peak PPA (cm H2O) | 14.0±0.9 | 14.0±1.0 |
| Time to Baseline (min) | 8.2±0.6 | 1.2±0.1* |
| Time to Half of Peak (min) | 4.2±0.4 | 0.3±0.1* |
Values are means ±SE of 6 experiments. ANF, atrial natriuretic factor.
p < 0.05
Table 2.
ANF at Peak Response to PGF2α Caused a Partial Recovery of Lung Resistance (RL) and Dynamic Compliance (Cdyn) Values
| RL (cm H2O/ml/sec) | Cdyn(ml/cmH2O) | |
|---|---|---|
| Baseline | 0.21±0.02 | 0.29±0.02 |
| Peak Response | 0.32±0.04 | 0.22±0.01 |
| After ANF | 0.24±0.03 | 0.25±0.02 |
| % Recovery (1 min) | 78.33±4.87 | 47.67±1.96 |
Values are means ± SE of 6 experiments
Fig. 2.
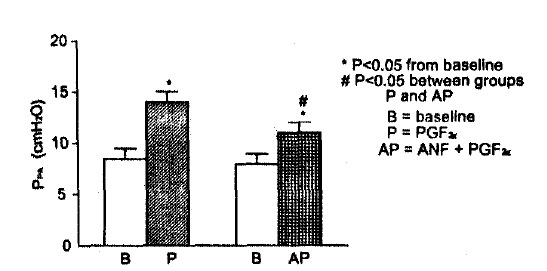
Pulmonary arterial pressure response to PGF2α without and with pre-treatment with atrial natriuretic factor(ANF). Values are means±SE of 6 experiments.
3. Mechanism of Ppa Response to PGF2α
NNLA increased baseline Ppa only at doses>100 μM. Subsequent injection of PGF2α caused pulmonary edema associated with a pronounced increase (>60 mmH2O) in Ppa. The dose of NNLA we chose did not affect baseline Ppa, nor was there any change in airway mechanics after the addition of NNLA in the perfusate. 100 μM of NNLA potentiated the peak Ppa response to 50 μg PGF2α with no statistically significant change in RL and Cdyn (Fig. 3).
Fig. 3.
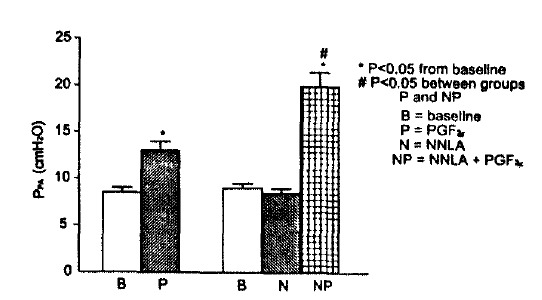
Pulmonary arterial pressure response to PGF2α without and with pretreatment with Nw-nitro-L-arginine(NNLA).
Values are means±SE of 12 experiments.
Before and after addition of NNLA, the response to 50 μg PGF2α were as follows : Ppa values increased from 8.7±0.3 to 13.1± 0.5 cmH2O without NNLA and from 9.0±0.3 to 20.0±1.1 cmH2O with NNLA. The difference in the rise in Ppa with and without NNLA was significant at p<0.01. Similarly, PGF2α increased RL values from 0.20±0.01 to 0.31±0.01 cmH2O/ml/sec without NNLA and from 0.22±0.01 to 0.31 ±0.01 cmH2O/ml/sec with NNLA. Cdyn decreased from 0.28 ±0.02 to 0.21 ±0.01 ml/cmH2O without NNLA and from 0.26±0.01 to 0.20±0.01 ml/cmH2O with NNLA. The addition of SQ 29548 and Cl fully blocked the pulmonary vascular and airway effects of PGF2α (data not shown).
DISCUSSION
PGF2α caused both pulmonary vasoconstriction and bronchoconstriction in rat lungs which were isolated and perfused with Krebs-Henseleit bicarbonate buffer solution. The vascualr effects are similar to those reported by others1). It has been shown that PGF2α increases Ppa with an increase in pulmonary vascular resistance but has no effect on pulmonary vascular permeability either in vitro or in vivo in dogs and sheep. In addition, PGF2α causes bronchoconstriction both in vitro and in vivo in guinea pigs, dogs and humans. Although we wanted a maximal pulmonary response following PGF2α injection into rat lungs, we limited the PGF2α dose to 50 μg, because severe pulmonary hypertension and pulmonary edema developed at higher doses. Only a small part (29%) of the pulmonary arterial pressure rise was due to an increase in arterial resistance. With an F2-isoprostane, 8-epi-PGF2α, we also found that only 28% of the rise in Ppa in rat lungs was due to an increase in arterial resistance7). On the other hand, in rabbits, 78% of the 8-epi-PGF2α-induced increase in Ppa was contributed by the increase in arterial resistance11). Ducharme et al17) found that the primary site of activity of PGF2α in dogs was the smooth muscle of the small veins2). In contrast, taylor and his co-workers pavititioned the effect of PGF2α on vascalar resistance of isolated rat lungs into segments of large and small arteries and veins and found that the primary site of action was the small arteries2). In rat lungs, PGF2α caused contraction of the airways, as observed with other species of animals1). However, PGF2α caused a larger change in lung resistance than in dynamic compliance, which suggests that effects of PGF2α is greater on central than on peripheral airways of rats18). This needs to be confirmed with tissue bath experiments containing different segments of the airways.
In our experiments, NNLA, an NO synthase inhibitor, potentiated the pulmonary vascular response to PGF2α with no change in airways mechanics. This suggests that the effect of PGF2α on pulmonary vasculature can be modulated by the release of NO. It is somewhat controversial whether NO contributes to normal low pulmonary arterial tone19). In isolated perfused lungs of pigs, sheep and humans. Cremona et al20) showed that release of NO regulates basal pulmonary vascular tone. On the other hand, Hasunuma et al21). like us, found that NO inhibitors did not increase the baseline perfusion pressure in isolated rat lungs. Also, Nishiwaki et al22) showed in conscious dogs that NNLA had no effect on the baseline pulmonary vascular pressure-flow relationship but did potentiate the pulmonary vasoconstrictor response to the TX mimetic. U-46619. However, Thomas et al23) showed in dogs, in vivo, an increase in pulmonary vascular tone in both normoxic and hypoxic lung after the administration of N-methyl arginine, another NO blocker. The use of methyl arginine, as an adequate probe to study the role of endogenous EDRF has recently been challenged by Hyman and his co-workers24). These authors failed to demonstrate the inhibition of NO in adult feline pulmonary vascular bed in vivo, by this enzyme which contains a methyl group at the quanidino-nitrogen position. The same methyl group, containing enzyme, has recently been found to paradoxically relax preconstricted canine intrapulmonary arteries and a role of an intermediate dliator-prostaglandin has been suggested for its action25). We conclude that NO has variable effects on resting tone, depending on species and baseline conditions, but that NO synthase inhibition by NNLA is associated with augmented vasoconstrictor responses.
Both the airway hemodynamic effects of PGF2α were fully prevented by SQ 29548, an endoperoxide-TX receptor blocker. This strongly suggests that the mechanism of PGF2α-induced pulmonary vasoconstriction and bronchoconstriction is due to TX receptor activation. We showed earlier, in sheep, that TX receptor blockade antagonized the effect of PGF2α and PGF2α on the pulmonary vasculature, with no effect on either the systemic vasculature or the airways26). Evidence for TX receptor-mediated contraction by PGF2α PGF2α and PGF2α has been shown in guinea pig and human airways in vitro by several investigators20,27–29). Finally, at the cellular level, the muscular contraction of pulmonary vasculature and the airways may be due to the involvement of protein kinase C (PKC), since the use of CI. a PKC inhibitor completely blocked the pulmonary response to PGF2α. There is evidence for the participation of both intracellular calcium and PKC in PGF2α-induced vasoconstriction4,30). Involvement of PKC in the tonic phase of smooth muscle contraction has also been suggested by other investigators31–33). However, the pulmonary response of isolated-perfused lungs of rats to PGF2α was more of a phasic rather than a tonic nature. Thus, we cannot assert fully that these data are sufficient to prove the definitive role of PKC in the observed response.
In summary, PGF2α is both pulmonary vasoconstrictor, primarily a venoscostrictor, and a bronchoconstrictor in isolated perfused lungs of rats. The effect of PGF2α on rat lungs appears to be greater on central than on peripheral airways. The vaso-, but not the bronchoconstrictor effect, is potentiated by the use of NNLA, a nitric oxide (EDRF) synthase inhibitor. However, both the vaso and bronchoconstrictor effects are partially reversed by the use of atrial natriuretic peptide. The mechanism of action of PGF2α on the pulmonary vasculature and the airways appears to be due to the activation of SQ 29548-responsive thromboxane receptors. Also, it is likely that protein kinase C may play a role in the smooth muscle response of pulmonary vessels and airways to PGF2α intracellular.
REFERENCES
- 1.Shore SA, Austen KF, Drazen JM, editors. Lung cell biology. New York: Dekker; 1989. p. 1011. [Google Scholar]
- 2.Bernard JW, Ward RA, Tayler AE. Evaluation of prostaglandin F2α and prostacyclin in the isolated perfused rat lung. J Appl Physiol. 1992;72:2469. doi: 10.1152/jappl.1992.72.6.2469. [DOI] [PubMed] [Google Scholar]
- 3.Ruttner E, Huszar E, Barat E, et al. Effects of arachidonic acid and prostaglandin F2α in isolated rat lungs. Acta Physiol Hungarica. 1990;75:53. [PubMed] [Google Scholar]
- 4.Santoian EC, Angerio AD, Schneidkraut MJ, et al. Role of calcium in U46619 and PGF2α pulmonary vasoconstriction in rat lungs. Am J Physiol. 1989;257(Heart Circ Physiol 26):H2001. doi: 10.1152/ajpheart.1989.257.6.H2001. [DOI] [PubMed] [Google Scholar]
- 5.Newman JH, MeMurtry IF, Reeves JT. Blunted pulmonary pressor responses to hypoxia in blood perfused, ventilated lungs isolated from oxygen toxic rats : Possible role of prostaglandins. Prostaglandins. 1981;22:11. doi: 10.1016/0090-6980(81)90050-2. [DOI] [PubMed] [Google Scholar]
- 6.Amdur MO, Mead J. Mechanics of respiration in unanesthetized guinea pigs. Am J Physiol. 1958;192:364. doi: 10.1152/ajplegacy.1958.192.2.364. [DOI] [PubMed] [Google Scholar]
- 7.Kang KH, Morrow JD, Roberts LJ, II, et al. Airway and vascular effects of 8-epi-prostaglandin F2α in isolated perfused rat lung. J Appl Physiol. 1993;74:460. doi: 10.1152/jappl.1993.74.1.460. [DOI] [PubMed] [Google Scholar]
- 8.Hakim TS, Michel RP, Chang HK. Partitioning of pulmonary vascular resistance in dogs by arterial and venous occlusion. J Appl Physiol. 1992;52:710. doi: 10.1152/jappl.1982.52.3.710. [DOI] [PubMed] [Google Scholar]
- 9.Townsley MI, Korthuis RJ, Rippe B, et al. Validation of double vascular occlusion method for Pc, in lung and skeletal muscle. J Appl Physiol. 1986;61:127. doi: 10.1152/jappl.1986.61.1.127. [DOI] [PubMed] [Google Scholar]
- 10.Atals SA, Laragh JH. In: Atrial hormones and other natriuretic factors. Mulrow PJ, Schrier R, editors. Bethesda MD: 1987. Am Physiol Soc chapt 6p 53 Clin Physiol Ser. [Google Scholar]
- 11.Banerjee M, Kang K, Morrow JD, et al. Effects of a novel prostaglandin, 8-epi-PGF2α in rabbit lung in situ. Am j Physiol. 1992;263(heart Circ Physiol 32):H660. doi: 10.1152/ajpheart.1992.263.3.H660. [DOI] [PubMed] [Google Scholar]
- 12.Ishi K, Chang B, kerwin JF, et al. Nω-nitro-L-arginine-: A potent inhibitor of endothelium-dependent relaxing factor formation. Eur J Pharmacol. 1990;176:219. doi: 10.1016/0014-2999(90)90531-a. [DOI] [PubMed] [Google Scholar]
- 13.Lamontagne D, Pohi U, Russe R. Nw-NITRO-L-arginine antagonizes endothelium-dependent dilator responses by inhibiting endothelium-dependent relaxing factor release in the isolated rabbit heart. Pfuegers Arch. 1991;418:266. doi: 10.1007/BF00370525. [DOI] [PubMed] [Google Scholar]
- 14.Wagerle LC, Busija D. Effect of thromboxane A2 endoperoxide antagonist SQ 29548 on the contractile response to acetylcholine in newborn piglet cerebral arteries. Circ Res. 1990;66:824. doi: 10.1161/01.res.66.3.824. [DOI] [PubMed] [Google Scholar]
- 15.Harvath L. Inhibition of human chemotzxis by the protein kinase inhibitor [1-(5-isoquinolinesulfonyl) piperazine] J Immunol. 1987;139:3055. [PubMed] [Google Scholar]
- 16.Salzer W, Gerard C, McCall C. Effects of an inhibitor of protein kinase C on human polymorphonuclear leukocyte degranulation. Biochem Biophys Res Commun. 1987;148:747. doi: 10.1016/0006-291x(87)90939-9. [DOI] [PubMed] [Google Scholar]
- 17.Ducharme DW, Weeks JR, Montagomery RG. Studies on the mechanism of the hypertensive effect of prostaglandin F2α. J Pharmacol Exp Ther. 1968;160:1. [PubMed] [Google Scholar]
- 18.Banerjee MR, Newman JH. Acute effects of atrial natriuretic peptide on lung mechanics and hemodynamics in awake sheep. J Appl Physiol. 1990;69:728. doi: 10.1152/jappl.1990.69.2.728. [DOI] [PubMed] [Google Scholar]
- 19.Peach MJ, Johns RA, Rose CE Jr, editors. pulmonary Vascular Physiology Pathophysiology. New York: Dekker; 1989. p. 643. [Google Scholar]
- 20.Cremona G, Higenbottam T, Xuan ATD, et al. Basal release of endothelium-derived relaxing factors (EDRF) regulates the pulmonary vascular resistance (PVR) in isolated-perfused porcine, sheep and human lungs. Am Rev Resp Dis. 1991;143:A771. [Google Scholar]
- 21.Hasunuma K, Yamaguchi T, Rodman DM, et al. Effects of inhibitors of EDRF: and EDHF on vasoreactivity of perfused rat lungs. Am J Physiol. 1991;260(Lung Cell Mol Physiol 4):L97. doi: 10.1152/ajplung.1991.260.2.L97. [DOI] [PubMed] [Google Scholar]
- 22.Nishiwaki K, Nyhan DP, Rock P, et al. Nω-nitro-L-arginine and pulmonary vascular pressure flow relationship in conscious dogs. Am J Physiol. 1992;262(Heart Circ Physiol 31):H1331. doi: 10.1152/ajpheart.1992.262.5.H1331. [DOI] [PubMed] [Google Scholar]
- 23.Thomas HM, Carson R, Gupta G, et al. The effect of N-methyl arginine(NMA) on the pulmonary circulation in the presence of regional hypoxia. Am Rev Resp Dis. 1991;143:A775. [Google Scholar]
- 24.Lippton HL, Hao Q, Hyman A. L-NAME enhances pulmonary vasoconstriction inhibiting EDRF-dependent vasodilation. J Appl Physiol. 1992;73:2432. doi: 10.1152/jappl.1992.73.6.2432. [DOI] [PubMed] [Google Scholar]
- 25.Tseng C, Godman LW, Rubin LJ, et al. NG-monomethyl-L-arginine paradoxically relaxes preconstricted canine intrapulmonary arteries. J Appl Physiol. 1993;74:549. doi: 10.1152/jappl.1993.74.2.549. [DOI] [PubMed] [Google Scholar]
- 26.King LS, Fukushima M, Banerjee M, et al. Pulmonary vascular effects of prostaglandin D2, but not its systemic vascular and airway effects, are mediated through thromboxane receptor activation. Circ Res. 1991;68:352–358. doi: 10.1161/01.res.68.2.352. [DOI] [PubMed] [Google Scholar]
- 27.Armour CL, Johnson PRA, Alfredson ML, et al. Characterization of contractile prostanoid receptors on human airway smooth muscle. Eur J Pharmacol. 1989;165:215. doi: 10.1016/0014-2999(89)90715-2. [DOI] [PubMed] [Google Scholar]
- 28.Coleman RA, Sheldric RLG. Prostanoid-induced contraction of human bronchial smooth muscle is mediated by TP-receptros. Br J Pharmacol. 1989;96:688. doi: 10.1111/j.1476-5381.1989.tb11869.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Featherstone RL, Robinson C, Holgate ST, et al. Evidence for thromboxane receptor mediated contraction of guinea pig and human airways in vitro by prostaglandin (PG)D2 9 alpha, 11 beta-PGF2α and PGF-alpha. Life Sci. 1990;46:1301. doi: 10.1007/BF00176337. [DOI] [PubMed] [Google Scholar]
- 30.Morimoto S, Kim S, Fukuo K, et al. Participation of both intracellular free Ca2+ and protein kinase C in tonic vasoconstriction induced by prostaglandin F2α. Eur J Pharmacol. 1990;188:369. doi: 10.1016/0922-4106(90)90197-6. [DOI] [PubMed] [Google Scholar]
- 31.Nakaki T, Roth BL, Chuang D, et al. Phasic and tonic components in 5-HT2 receptor-mediated rat aorta contraction: Participation of Ca2+ channels and phospholipase. Cl. J Pharmacol Exp Ther. 1985;234:442. [PubMed] [Google Scholar]
- 32.Rasmussen H. The calcium messenger system. N Engl J Med. 1986;314:1094. doi: 10.1056/NEJM198604243141707. [DOI] [PubMed] [Google Scholar]
- 33.Rasmussen H. The calcium messenger system. N Engl J Med. 1986;314:1164. doi: 10.1056/NEJM198605013141807. [DOI] [PubMed] [Google Scholar]