

Abstract
Despite the improvement in chemotherapeutic agents, the outcome of patients with prostate cancer remains poor. It is therefore imperative that new anticancer drugs are explored. The aim of the present study was to investigate the inhibitory effect of bortezomib on DU145 prostate cancer cells. The DU145 cell proliferation rate was detected via MTT assay prior to and following exposure to various concentrations of bortezomib, and the level of cell apoptosis and the cell cycle distribution were tested using flow cytometry. In addition, western blotting was used to measure the expression of Bcl-2-interacting killer (Bik) and active-caspase-3. The results showed that bortezomib inhibited the proliferation of DU145 cells in a time- and dose-dependent manner. Following treatment with 1.6 µmol/l bortezomib, the DU145 cells showed marked nuclear condensation, chromatin condensation and fragmentation. Analysis of the cell cycle revealed a significantly increased percentage of cells in the G0/G1 phase and a decreased percentage in the S and G2/M phases. The rate of DU145 cell apoptosis was significantly higher in the bortezomib group than that in the control group, and this was accompanied by an enhanced expression of Bik and active-caspase-3. It can be concluded that bortezomib inhibits the proliferation of DU145 cells by inducing apoptosis. The underlying mechanism may involve the upregulation of Bik and active-caspase-3 expression.
Keywords: prostate cancer, bortezomib, apoptosis
Introduction
As the second most common type of cancer in men worldwide, prostate cancer mainly affects elderly male patients (1). In the United States, prostate cancer is the most frequently occurring tumor type in men, with ~217,730 new cases annually (2). Prostate cancer accounts for ~11% of all types of tumor in European men and is responsible for 9% of the cases of cancer-related mortality (3). Patients with prostate cancer lack specific clinical symptoms during the early stages of the disease, meaning that, when a diagnosis of prostate cancer is reached, the majority of patients are at an advanced stage, when the tumor is no longer resectable. Despite the improvement in chemotherapeutic agents, the outcome of patients with prostate cancer remains poor, and it is therefore imperative that new anticancer drugs are explored.
As a proteasome inhibitor, bortezomib was first approved by the Food and Drug Administration for the treatment of relapsed or refractory multiple myeloma (4). Bortezomib has additionally been confirmed to elicit a good response in the treatment of certain solid tumors (5). The drug selectively binds to and inhibits the chymotryptic-like activity of the proteasome at nanomolar concentrations. Proteasome inhibition is associated with decreased tumor cell proliferation rates and increased apoptosis (6). A previous study indicated that Bcl-2-interacting killer (Bik)/natural born killer (NBK) is one of the mediators of proteasome inhibitor-induced apoptosis in several cell lines (7). Furthermore, bortezomib reverses the proliferative and anti-apoptotic effect of neuropeptides, particularly nuclear factor-κB (NF-κB), in prostate cancer cells (8). By contrast, bortezomib treatment alone is not effective for the treatment of metastatic castration-resistant prostate cancer (9). The available results concerning the effect of bortezomib on prostate cancer are therefore conflicting. To help resolve this conflict, the present study further investigated the inhibitory effect of bortezomib on prostate cancer by treating DU145 prostate cancer cells with bortezomib and observing the induced inhibitory effect and the rate of DU145 cell apoptosis, with the aim of providing a theoretical basis for the clinical treatment of prostate cancer with bortezomib.
Materials and methods
Drugs and reagents
Bortezomib was obtained from Xi'an Janssen Pharmaceutical Co., Ltd. (Xi'an, China). MTT and propidium iodide (PI) dye were purchased from Sigma-Aldrich (St. Louis, MO, USA). Fetal bovine serum and RPMI-1640 medium were obtained from Evergreen Biotech Co., Ltd. (Hangzhou, China). The Annexin V apoptosis kit was purchased from Unitech Biotechnology Co., Ltd. (Nanjing, China). Rabbit monoclonal anti-human Bik (1:500), active-caspase-3 (1:500) and β-actin (1:1,000) antibodies were obtained from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA, USA), while goat anti-rabbit IgG secondary antibody (1:3,000) was purchased from Boster Biological Technology Co., Ltd. (Wuhan, China).
Cell culture
DU145 human prostate cancer cells were obtained from the Chinese Academy of Medical Sciences (Beijing, China). The cells were maintained in RPMI-1640 medium supplemented with 10% fetal bovine serum in a chamber with 5% CO2 at 37°C.
Cell proliferation assay
DU145 cells in the logarithmic phase were seeded into a 96-well, round-bottomed culture plate (2×104 cells per well) with the total volume of 100 µl. According to the experimental design, the cells were divided into phosphate-buffered saline (PBS)-treated and bortezomib-treated (0.2, 0.4, 0.8, 1.6, 3.2 and 6.4 µmol/l) groups. The cells were incubated for 0, 12, 24 and 48 h, respectively. After the indicated times, an MTT assay microplate reader (Wuhan Boster Biological Technology Ltd., Wuhan, China) was used to measure the absorbance value (A) at a wavelength of 570 nm. The growth inhibition rate was calculated as follows: Growth inhibition rate (%) = (Acontrol group - Aexperimental group)/(Acontrol group - Ablank group) × 100.
Cell cycle analysis
The DU145 cells were seeded into 12-well plates (5×105 cells per well) with a 2-ml total volume and treated with PBS or 1.6 µmol/l bortezomib. The morphology of the anchorage-dependent cells was observed 24 h after exposure to bortezomib. The cells were fixed with 70 ml/l cold ethanol, digested with RNase (Wuhan Boster Biological Technology Ltd.), dyed with PI and the cell cycle status was detected using flow cytometry (FCM).
Cell apoptosis analysis
The DU145 cells were trypsinized 24 h after treatment with 1.6 µmol/l bortezomib and then washed with PBS. Those cells were then suspended in binding buffer and the number of the cells was adjusted to 5×105/ml. Annexin V-fluorescein isothiocyanate and 10 µl PI were added to 100 µl cell suspension and mixed thoroughly; this mixture was then kept in the dark for 15 min. Finally, FCM was used to detect the apoptosis rate of the DU145 cell.
Western blot analysis
Following treatment with 1.6 µmol/l bortezomib for 24 h, the cells were washed with cold PBS and lysed in lysis buffer containing 150 mM NaCl, 1% NP40, 50 mM Tris/HCl, 1 mM ethylene glycol tetraacetic acid, 1 mM phenylmethanesulfonyl fluoride, 10 mM Na4P2O7, 10 mM NaF, 1 mM Na3VO5, 10 µg/ml leupeptin and 20 µg/ml aprotinin (Wuhan Boster Biological Technology Ltd.). A bicinchoninic acid assay (Pierce Biotechnology, Inc., Rockford, IL, USA) was used to measure the total protein concentration in the sample. Following the addition of the protein loading buffer (50 mM Tris/HCl, 10% glycerol, 0.02% bromophenol blue, 2% β-mercaptoethanol and 5% sodium dodecyl sulfate (SDS), pH 6.8) and denaturation for 5 min at 95°C, 12% SDS-polyacrylamide gel electrophoresis was used to separate 40 µg total protein from each sample, prior to the protein being transferred onto nitrocellulose membranes. The membranes were blocked in 5% non-fat milk for 1 h at 37°C and then incubated with the primary antibodies (active-caspase-3, Bik and β-actin) for 24 h at 4°C. Following incubation with the primary antibodies, the membranes were incubated with the horseradish peroxidase-conjugated secondary antibody for 1 h at room temperature in blocking buffer. Signal visualization was performed using enhanced chemiluminescence, and the data were quantified by scanning and analyzing the average volume density of the hybridization signals, corrected against β-actin, using Gel-Pro 3.0 image analysis software (Media Cybernetics, Silverspring, MD, USA).
Statistical analysis
Data are expressed as the mean ± standard error from at least three independent experiments. Statistical analysis of the data was performed with the Student's t-test for the comparison of two groups or one-way analysis of variance for multiple comparisons using SPSS 13.0 software (SPSS, Inc., Chicago, IL, USA). P<0.05 was considered to indicate a statistically significant difference.
Results
Bortezomib inhibits the proliferative activity of DU145 cells
The MTT assay results showed that the various concentrations of bortezomib had no significant inhibitory effect on the proliferation of DU145 cells within 12 h (P>0.05) (Table I). After 24 h, bortezomib inhibited the proliferation of the DU145 cells in a time- and dose-dependent fashion (P<0.05) (Table I). A bortezomib concentration of 1.6 µmol/l was selected to perform the subsequent experiments, as the half maximal inhibitory concentration of bortezomib against the DU145 cells.
Table I.
Effect of bortezomib on cell proliferative activity.
| Concentration of bortezomib (µmol/l) | Growth inhibition rate (%) | ||
|---|---|---|---|
| 12 h | 24 h | 48 h | |
| 0.2 | 3.26±0.82 | 4.17±1.64 | 5.13±0.16 |
| 0.4 | 5.32±2.12 | 9.65±1.50* | 11.48±1.23* |
| 0.8 | 6.59±2.43 | 21.31±1.51* | 27.88±2.46* |
| 1.6 | 7.29±4.30 | 53.13±8.84* | 58.66±8.14* |
| 3.2 | 6.98±5.13 | 61.36±7.24* | 72.17±8.72* |
| 6.4 | 8.63±5.81* | 66.25±9.15* | 76.57±9.06* |
Results are presented as the mean ± standard error.
P<0.05 vs. control group.
Bortezomib induces G1-phase cell cycle arrest in DU145 cells
The results from the flow cytometric analysis are shown in Fig. 1. After 24 h of exposure to 1.6 µmol/l bortezomib, flow cytometric analysis revealed a significant sub-G1 peak (apoptosis peak) in the cell cycle of the DU145 cells (Fig. 1B). In the control group, the proportion of cells in G0/G1, S and G2/M phase was 48.15±4.3, 23.42±3.83 and 28.37±4.0%, respectively (Fig. 1A), compared with 70.18±6.25, 12.60±1.72 and 17.28±6.25%, respectively, in the cells treated with 1.6 µmol/l bortezomib (Fig. 1B) (P<0.05).
Figure 1.
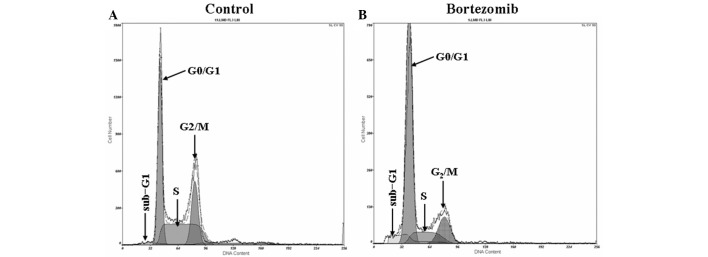
Effect of bortezomib on the cell cycle of DU145 cells. Following incubation in the presence or absence of 1.6 µmol/l bortezomib for 24 h, the cell cycle distribution of the DU145 cells was measured via flow cytometry and quantitative analysis. (A) Normal control group; (B) bortezomib-treated group.
Bortezomib triggers apoptotic morphological changes in DU145 cells
After 24 h of treatment with 1.6 µg/ml bortezomib, the adherent cells showed morphological changes of apoptosis, including condensed chromatin, nuclear condensation, nuclear marginalized, nuclear fragmentation and apoptotic body formation. These adherent cells then became crimpled, grew round and at last detached (Fig. 2). FCM showed that, at 24 h, the apoptosis rate of the 1.6 µmol/l bortezomib-induced DU145 cells reached 30.1%, which was significantly higher than that of the control group (Fig. 3A and B).
Figure 2.
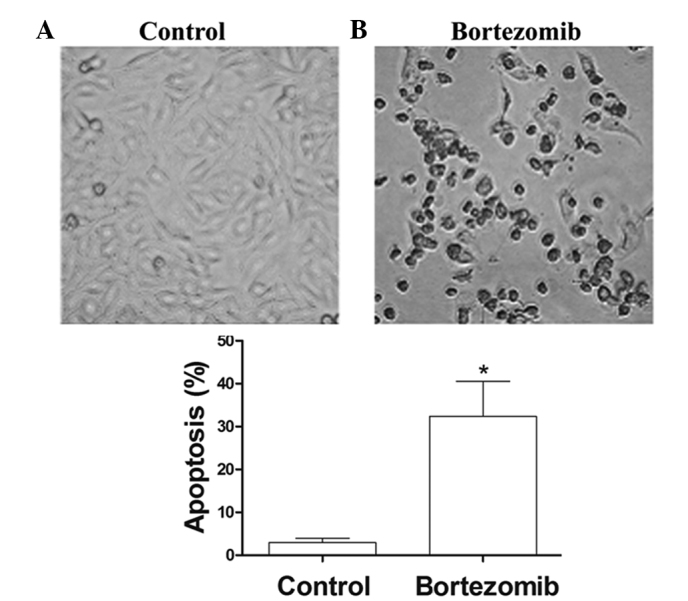
Effect of bortezomib on the morphology of DU145 cells observed under the light microscope (magnification, x400). Following incubation in the presence or absence of 1.6 µmol/l bortezomib for 24 h, the cells were observed using light microscopy and quantitatively analyzed. (A) Normal control group; (B) bortezomib-treated group. Results are presented as the mean ± standard error. *P<0.05 vs. the control group.
Figure 3.

Effect of bortezomib on the apoptosis of DU145 cells. Following incubation in the presence or absence of 1.6 µmol/l bortezomib for 24 h, the cells were assessed using (A and B) flow cytometry and (C) quantitative analysis. (A) Normal control group; (B) bortezomib-treated group. (C) In quantitative analysis, results are presented as the mean ± standard error. *P<0.05 vs. the control group. PI, propidium iodide; FITC, fluorescein isothiocyanate.
Bortezomib increases the levels of Bik and active-caspase-3 in DU145 cells
The DU145 cells were incubated in the presence or absence of 1.6 µmol/l bortezomib for 24 h. Western blot analysis showed that the levels of active-caspase-3 (Fig. 4A and B) and Bik (Fig. 4A and C) in the DU145 cells were significantly increased following treatment with bortezomib when compared with the levels in the control group (P<0.05).
Figure 4.

Detection of Bik and active-caspase-3 expression in DU145 cells. DU145 cells were incubated in the presence or absence of 1.6 µmol/l bortezomib for 24 h. Levels of (A and B) active-caspase-3 and (A and C) Bik were assessed using immunoblotting and quantitative analysis. Results are presented as the mean ± standard error. *P<0.05 vs. the control group. Bik, Bcl-2-interacting killer.
Discussion
Prostate cancer occurs mainly in elderly male patients. The treatment of prostate cancer is closely dependent on clinical staging and mainly includes active surveillance, radical surgery, radiotherapy and endocrine therapy. Due to the high risk of bone metastases associated with prostate cancer, surgical treatment has significant limitations (10). The majority of patients with prostate cancer that are initially treated by androgen suppression medication experience relapse or refractory disease due to the evolution of the tumor from hormone-dependent to hormone-independent (11). Few treatments are effective against androgen-independent prostate cancer (12).
In the present study, different concentrations of bortezomib were used to treat the DU145 prostate cancer cell line. In the DU145 cells treated for >12 h, cell growth was significantly inhibited in a time- and dose-dependent manner. FCM demonstrated the occurrence of G0/G1-phase cell cycle arrest in the bortezomib-treated group of DU145 cells. The mechanisms responsible for this bortezomib-induced apoptosis may involve the inhibition of the 26S proteasome, an essential element in the degradation of intracellular proteins, including p53, NF-κB inhibitor IB and cyclin-dependent kinase inhibitors, and particularly the repression of the ability of NF-κB to transcribe its target genes involved in tumor growth, angiogenesis and metastasis, such as VEGF and IL-8 (13). Bortezomib therefore induces cell cycle arrest and apoptosis by shifting the balance between proapoptotic and antiapoptotic signals (14), accompanying the degradation of DNA in the target cells (15). The evolvement of androgen-independent tumors has been associated with several different mechanisms, such as the perturbation of mediators of prostate cancer cell growth and tumor suppression in the cell. The most important mediators are PTEN and HER2/neu, which act upstream of the PI3K/Akt signaling pathway; in turn, the critical proapoptotic signaling by Bad and p27 is deactivated and the prosurvival NF-κB signaling is activated (16). The presence of bortezomib may thus prevent the transformation from androgen-dependence to androgen-independence in prostate cancer cells.
To explore the mechanisms associated with the effect of bortezomib, the expression of the proapoptotic proteins Bik and active-caspase-3 was examined in the present study. Enhanced Bik protein expression can block the pathway of the antiapoptotic proteins B-cell lymphoma 2 (Bcl-2) and Bcl-XL, and therefore activate the mitochondrial pathway of apoptosis, which involves cytochrome c release, and lead to apoptosis (17). Proteasome inhibitors have been confirmed to induce the rapid aggregation of the Bik/NBK proapoptotic proteins in the DLD-l, LoVo, SW620 and HCTI116 human colon cancer cell lines and then promote apoptosis (7). Similarly, the Bik/NBK proteins are activated by bortezomib in DU145 prostate cancer cells. The caspase family plays an important role in the regulation of apoptosis. Caspase-3 has an indispensable function in the caspase cascade reaction and is a key enzyme and initiator of apoptosis (18). The inhibition of caspase-3 activity significantly blocks apoptosis in vitro and in vivo (19). In the present study it was found that the treatment of DU145 cells with bortezomib caused an increase in the levels of Bik and active-caspase-3, which was responsible for the bortezomib-induced apoptosis.
In conclusion, the present study has shown that bortezomib can inhibit prostate cancer cell growth and induce apoptosis. The underlying mechanism may be associated with the upregulation of Bik and active-caspase-3 expression. This finding suggests the potential promise of bortezomib in the treatment of prostate cancer.
Acknowledgements
This study was supported by a grant from National Natural Science Foundation (no. 81460067).
References
- 1.Jemal A, Bray F, Center MM, Ferlay J, Ward E, Forman D. Global cancer statistics. CA Cancer J Clin. 2011;61:69–90. doi: 10.3322/caac.20107. [DOI] [PubMed] [Google Scholar]
- 2.Jemal A, Siegel R, Xu J, Ward E. Cancer statistics, 2010. CA Cancer J Clin. 2010;60:277–300. doi: 10.3322/caac.20073. [DOI] [PubMed] [Google Scholar]
- 3.Bray F, Sankila R, Ferlay J, Parkin DM. Estimates of cancer incidence and mortality in Europe in 1995. Eur J Cancer. 2002;38:99–166. doi: 10.1016/S0959-8049(01)00350-1. [DOI] [PubMed] [Google Scholar]
- 4.Mateos MV, Hernandez JM, Hernandez MT, et al. Bortezomib plus melphalan and prednisone in elderly untreated patients with multiple myeloma: Results of a multicenter phase 1/2 study. Blood. 2006;108:2165–2172. doi: 10.1182/blood-2006-04-019778. [DOI] [PubMed] [Google Scholar]
- 5.Ryan DP, O'Neil BH, Supko JG, et al. A Phase I study of Bortezomib plus irinotecan in patients with advanced solid tumors. Cancer. 2006;107:2688–2697. doi: 10.1002/cncr.22280. [DOI] [PubMed] [Google Scholar]
- 6.Voorhees PM, Orlowski RZ. The proteasome and proteasome inhibitors in cancer therapy. Annu Rev Pharmacol Toxicol. 2006;46:189–213. doi: 10.1146/annurev.pharmtox.46.120604.141300. [DOI] [PubMed] [Google Scholar]
- 7.Zhu H, Zhang L, Dong F, et al. Bik/NBK accumulation correlates with apoptosis-induction by Bortezomib (PS-341, Velcade) and other proteasome inhibitors. Oncogene. 2005;24:4993–4999. doi: 10.1038/sj.onc.1208683. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Tsapakidis K, Vlachostergios PJ, Voutsadakis IA, et al. Bortezomib reverses the proliferative and antiapoptotic effect of neuropeptides on prostate cancer cells. Int J Urol. 2012;19:565–574. doi: 10.1111/j.1442-2042.2012.02967.x. [DOI] [PubMed] [Google Scholar]
- 9.Morris MJ, Kelly WK, Slovin S, et al. A phase II trial of Bortezomib and prednisone for castration resistant metastatic prostate cancer. J Urol. 2007;178:2378–2383. doi: 10.1016/j.juro.2007.08.015. [DOI] [PubMed] [Google Scholar]
- 10.Jemal A, Siegel R, Ward E, Murray T, Xu J, Thun MJ. Cancer statistics, 2007. CA Cancer J Clin. 2007;57:43–66. doi: 10.3322/canjclin.57.1.43. [DOI] [PubMed] [Google Scholar]
- 11.Ulmer JB, Wahren B, Liu MA. DNA vaccines: Recent technological and clinical advances. Discov Med. 2006;6:109–112. [PubMed] [Google Scholar]
- 12.Nasu Y, Kumon H. Prostate cancer gene therapy. Nihon Rinsho. 2005;63:485–490. (In Japanese) [PubMed] [Google Scholar]
- 13.Chen Z, Hagler J, Palombella VJ, et al. Signal-induced site-specific phosphorylation targets I kappa B alpha to the ubiquitin-proteasome pathway. Genes Dev. 1995;9:1586–1597. doi: 10.1101/gad.9.13.1586. [DOI] [PubMed] [Google Scholar]
- 14.Adams J. The development of proteasome inhibitors as anticancer drugs. Cancer Cell. 2004;5:417–421. doi: 10.1016/S1535-6108(04)00120-5. [DOI] [PubMed] [Google Scholar]
- 15.Fan XM, Wong BC, Wang WP, et al. Inhibition of proteasome function induced apoptosis in gastric cancer. Int J Cancer. 2001;93:481–488. doi: 10.1002/ijc.1373. [DOI] [PubMed] [Google Scholar]
- 16.Papandreou CN, Logothetis CJ. Bortezomib as a potential treatment for prostate cancer. Cancer Res. 2004;64:5036–5043. doi: 10.1158/0008-5472.CAN-03-2707. [DOI] [PubMed] [Google Scholar]
- 17.Huang DC, Strasser A. BH3-only proteins - essential initiators of apoptotic cell death. Cell. 2000;103:839–842. doi: 10.1016/S0092-8674(00)00187-2. [DOI] [PubMed] [Google Scholar]
- 18.Kumar S. Caspase function in programmed cell death. Cell Death Differ. 2007;14:32–43. doi: 10.1038/sj.cdd.4402060. [DOI] [PubMed] [Google Scholar]
- 19.de Oca J, Azuara D, Sanchez-Santos R, et al. Caspase-3 activity, response to chemotherapy and clinical outcome in patients with colon cancer. Int J Colorectal Dis. 2008;23:21–27. doi: 10.1007/s00384-007-0362-3. [DOI] [PubMed] [Google Scholar]