

Abstract
It was hypothesized that nitric oxide synthases (NOS) regulated SIRT1 expression and lead to a corresponding changes of contractile and metabolic properties in skeletal muscle. The purpose of the present study was to investigate the influence of long-term inhibition of nitric oxide synthases (NOS) on the fiber-type composition, metabolic regulators such as and silent information regulator of transcription 1 (SIRT1) and peroxisome proliferator-activated receptor γ coactivator-1α (PGC-1α), and components of mitochondrial biogenesis in the soleus and plantaris muscles of rats. Rats were assigned to two groups: control and NOS inhibitor (Nω-nitro-L-arginine methyl ester hydrochloride (L-NAME), ingested for 8 weeks in drinking water)-treated groups. The percentage of Type I fibers in the L-NAME group was significantly lower than that in the control group, and the percentage of Type IIA fibers was concomitantly higher in soleus muscle. In plantaris muscle, muscle fiber composition was not altered by L-NAME treatment. L-NAME treatment decreased the cytochrome C protein expression and activity of mitochondrial oxidative enzymes in the plantaris muscle but not in soleus muscle. NOS inhibition reduced the SIRT1 protein expression level in both the soleus and plantaris muscles, whereas it did not affect the PGC-1α protein expression. L-NAME treatment also reduced the glucose transporter 4 protein expression in both muscles. These results suggest that NOS plays a role in maintaining SIRT1 protein expression, muscle fiber composition and components of mitochondrial biogenesis in skeletal muscle.
Key points.
NOS inhibition by L-NAME treatment decreased the SIRT1 protein expression in skeletal muscle.
NOS inhibition induced the Type I to Type IIA fiber type transformation in soleus muscle.
NOS inhibition reduced the components of mitochondrial biogenesis and glucose metabolism in skeletal muscle.
Key words: Mitochondrial biogenesis, muscle fiber composition, nitric oxide, nitric oxide synthase, silent information regulator of transcription 1
Introduction
Skeletal muscle is a heterogeneous tissue, the contractile and metabolic properties of which vary considerably. The skeletal muscle fibers in rodents are generally categorized as slow-twitch Type I and fast-twitch Type II fibers. Type II fibers are further subclassified into Type IIA, IIX and IIB fibers. The Type I, IIA, IIX, and IIB fibers express myosin heavy chains (MHC) I, IIA, IIX, and IIB, respectively (Pereira Sant’Ana et al., 1997). The fiber composition and metabolic capacity of skeletal muscle vary greatly (Gollnick et al., 1972). The basis of this variation in the contractile and metabolic properties of skeletal muscle fibers has been an important focus of research for several decades.
Nitric oxide (NO) is an inorganic molecule that acts as a signaling molecule in numerous cellular processes. NO has frequently been studied in the fields of physiology, biochemistry, and biomedicine. NO is generated mainly from the amino acid, L-arginine, by NO synthases (NOS). There are three NOS isoforms: endothelial (eNOS), neuronal (nNOS), and inducible NOS (iNOS), all of which are expressed in skeletal muscle fibers (Stamler and Meissner, 2001). The eNOS and nNOS isoforms are constitutively expressed, and are regulated by Ca2+-calmodulin binding (Stamler and Meissner, 2001; Toledo and Augusto, 2012). In contrast, iNOS is expressed during the activation of defense mechanisms against inflammation and infection, and is not regulated by the Ca2+-calmodulin system (Ghafourifar and Nathan, 1994).
NO is associated with the regulation of contractile properties of skeletal muscle, such as the muscle fiber composition and the pattern of MHC isoform expression (Drenning et al., 2008; Martins et al., 2012; Smith et al., 2002). Treatment with the NOS inhibitor, Nω-nitro-L-arginine methyl ester hydrochloride (L-NAME), has been shown to prevent the chronic functional overload-induced increase in Type I MHC and the decrease in Type IIB MHC in rat plantaris muscle (Smith et al., 2002). In addition, L-NAME treatment partially blocks the chronic low-frequency stimulation-induced fast-to-slow MHC transformation in rat fast-twitch muscles (Martins et al., 2012). In vitro experiments have also indicated that L-NAME treatment prevents the calcium ionophore A23187-induced increase in Type I MHC mRNA expression in C2C12 myotubes (Nathan and Xie, 1994). Collectively, these data have led to speculation that NOS/NO signals mediate the fast-to-slow conversion induced by muscle use, such as contraction and weight bearing, and the increasing sarcoplasmic Ca2+ concentrations. However, whether NOS/NO signaling is also associated with the muscle fiber composition under basal sedentary conditions remains to be determined.
NO controls the metabolic properties in skeletal muscle cells. Balon and Nadler (1994) first postulated that NO has a role in modulating skeletal muscle metabolism. They demonstrated that incubation of the rat extensor digitorum longus muscle with the NOS inhibitor, NG-monomethyl-L-arginine monoacetate (L-NMMA), significantly decreased both the basal 2-deoxyglucose transport and NO efflux. NOS/NO signaling regulates glucose metabolism, including glucose uptake (Higaki et al., 2001; Merry et al., 2010; Ross et al., 2007) and glucose transporter 4 (GLUT4) expression (Lira et al., 2007). In addition, NO seems to promote mitochondrial biogenesis and fatty acid oxidation. Treatment with the NO donor, (Z)-1-[N-(2-aminoethyl)-N-(2-ammonioethyl)amino] diazen-1-ium-1,2-diolate (DETA-NO), enhanced the expression of transcriptional regulators controlling mitochondrial biogenesis and fatty acid oxidation, such as peroxisome proliferator-activated receptor γ coactivator-1α (PGC-1α), nuclear respiratory factor 1 (NRF1), and mitochondrial transcriptional factor A (Tfam), as well as mitochondrial components, in L6 muscle cells (Lira et al., 2010; Nisoli et al., 2004). Furthermore, eNOS-/- mice showed a decreased mitochondrial volume and reduced expression of proteins related to mitochondrial β-oxidation (Le Gouill et al., 2007; Nisoli et al., 2004). In contrast, the activity of citrate synthase (CS), a marker of the mitochondrial tricarboxylic aid cycle, in skeletal muscle, as well as that in brain, kidney, and liver, in mice lacking nNOS is higher than that in wild-type mice (Schild et al., 2006).
Silent information regulator of transcription 1 (SIRT1) is an oxidized form of nicotinamide adenine dinucleotide (NAD+)-dependent protein deacetylase, and plays an important role in a variety of pathways leading to metabolic regulation and adaptation through its interactions with, and deacetylation of, transcriptional regulators (Boutant and Cantó, 2014). In skeletal muscle cells, SIRT1 regulates mitochondrial biogenesis and fatty acid oxidation via deacetylation and activation of PGC-1α (Gerhart-Hines et al., 2007; Rodgers et al., 2005). SIRT1 protein expression seems to be regulated by NOS/NO signaling. A previous study demonstrated that SIRT1 protein expression was increased in white adipocytes by treatment with NO donors including DETA-NO and S-nitroso N-acetylpenicilamine (SNAP), and 8 Br-cGMP, which is an analog of guanosine 3',5'-cyclic monophosphate (cGMP, a downstream molecule of NOS/NO signaling) (Nisoli et al., 2005). In addition, caloric restriction-induced enhancement of SIRT1 protein expression, as well as mitochondrial biogenesis in white and brown adipose tissue, brain, liver, and heart, are diminished or prevented in eNOS-/- mice (Nisoli et al., 2005). It was speculated that NOS/NO signaling regulated SIRT1 expression, and that SIRT1 played a role in NOS/NO signaling-promoted mitochondrial biogenesis in these tissues. We hypothesized that chronic inhibition of NOS would decrease SIRT1 expression and lead to a corresponding reduction of mitochondrial biogenesis in skeletal muscle.
This study was designed to identify the role of NOS in contractile and metabolic profiles under basal sedentary conditions. In this study, we investigated the effects of long-term NOS inhibition on muscle fiber composition. We also assessed the effects of NOS inhibition on SIRT1 protein expression and metabolic modifications in skeletal muscle.
Methods
Animals and design
Male Wistar rats (aged 8 weeks and weighing 279–304 g; Kyudo, Tosu, Saga, Japan) were housed two per cage (42 × 25 × 20 cm deep) in a temperature- (22 ± 2°C) and humidity- (60 ± 5%) controlled room with a 12-h light (7:00 to 19:00) and 12-h dark (19:00 to 7:00) cycle. All experimental procedures were conducted strictly in accordance with the Nakamura Gakuen University guidelines for the Care and Use of Laboratory Animals and were approved by the University Animal Experiment Committee.
The rats were divided into control (n = 10) and non-selective NOS inhibitor L-NAME-treated (n = 10) groups. The rats in the L-NAME group were treated with L-NAME (Cayman Chemical Company, Ann Arbor, MI, USA; 1 mg/mL) in their drinking water for 8 weeks (the water was replaced daily). This dose of L-NAME was previously shown to mediate strongly inhibition NOS activity in skeletal muscle (Roberts et al., 1999; Wang et al., 2001). The drinking water was provided ad libitum in both groups. A preliminary experiment suggested that the L-NAME-treated rats showed reduced (approximately 20%) food intake compared with freely fed untreated rats; therefore, the rats in the control group were pair-fed to match the quantity of food intake in the L-NAME-treated group. On the day of sampling, the chow was withdrawn for 30 min. Subsequently, the chow was returned to the rats for 30 min. After an 8-h fast, the rats were anesthetized with pentobarbital sodium (60 mg/kg body weight intraperitoneally) and the soleus and plantaris muscles of both legs were dissected rapidly. The muscles of the right leg were used for western blotting analyses and enzyme assays, and those of the left leg were used for histochemistry studies. The muscle tissues of the right leg were immediately frozen in liquid nitrogen, and were then stored at -80°C until the analyses were performed. Concerning the muscles of the left leg, the complete cross-sectional segments were cut at the muscle belly. Each muscle piece was mounted on a specimen holder in Tissue-Tek OCT compound (Sakura Finetechnical Co, Tokyo, Japan), and was frozen in isopentane previously cooled to a viscous fluid with liquid nitrogen. These muscle pieces were also stored –80°C until the analyses were performed. The epididymal fat pads were also excised and weighed.
Muscle histochemistry
Transverse sections (7 μm) were cut from each muscle using a cryostat maintained at –20°C, and the sections were then mounted on a cover slip. Myosin adenosine triphosphatase (ATPase) staining was performed using the previously described procedures (Suwa et al., 2002; 2003). Consecutive serial sections were processed using three different pretreatments: preincubation at pH 4.3, 4.6 and 10.4. The muscle fibers were identified as Type I, IIC, IIA, IIX and IIB fibers on the basis of the myosin ATPase staining intensity as described previously (Suwa et al., 2003). Each section was photographed using an Axioskop 2 plus microscope (Carl Zeiss, Hallbergmoos, Germany) mounted with an Axiocam HRm CCD camera (Carl Zeiss). The fiber type was identified and counted, and then the muscle fiber composition was determined by evaluating all countable fibers in both muscles. The percentage of each fiber type was calculated as follows: % of each fiber type = (number of each fiber type/number of all fiber types) × 100.
Preparation of muscle samples for biochemical analyses
The frozen muscle samples were homogenized in ice-cold homogenizing buffer (1:10 wt/vol) (25 mM HEPES, 250 mM sucrose, 2 mM EDTA, 0.1% Triton X-100, and one tablet/50 mL Complete™ Protease Inhibitor Cocktail Tablets (Roche Diagnostics, Tokyo, Japan), pH 7.4. The homogenate was centrifuged at 15,000 ×g (4°C) for 25 min. The protein concentration of the supernatant was determined using a protein determination kit (Protein Assay II, 500-0006, Bio-Rad Laboratories, Richmond, CA, USA). This muscle homogenate was used for the western blotting analyses and enzymatic assays. The muscle protein homogenate was solubilized in sample loading buffer (50 mM Tris-HCl, pH 6.8, 2% sodium dodecyl sulfate (SDS), 10% glycerol, 5% β-mercaptoethanol and 0.005% bromophenol blue) for the western blotting analyses.
Gel electrophoresis and the western blotting analysis
The proteins (20 μg) present in these homogenates were separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) using 7.5% (SIRT1 and PGC-1α), 10% (GLUT4), and 15% (cytochrome C) resolving gels. The proteins separated by SDS-PAGE were then transferred electrophoretically to a polyvinylidene difluoride membrane. The membrane was incubated with a blocking buffer of casein solution (SP-5020; Vector Laboratories, Burlingame, CA, USA) for 1 h at room temperature. The membrane was incubated with the appropriate primary detection antibodies [affinity-purified rabbit polyclonal antibody to Sir2 (1:1,000 dilution; #07-131; Upstate Biotechnology, Lake Placid, NY, USA), PGC-1α (1:500 dilution; AB3242; Chemicon International, Temecula, CA, USA), or GLUT4 (1:8,000 dilution; AB1346; Chemicon International) or a mouse monoclonal antibody to cytochrome C (1:200 dilution; ANN0012, clone 7H8.2C12, Biosource, Camarillo, CA, USA)] overnight at 4°C. Subsequently, the membrane was incubated with the biotinylated anti-rabbit/mouse IgG (1:1,000 dilution; BA-1400; Vector Laboratories) secondary detection antibody for 30 min. The bands on the membrane were visualized by the avidin and biotinylated horseradish peroxidase macromolecular complex technique (PK-6100; Vector Laboratories). The band densities were determined using the NIH Image 1.62 software package (National Institute of Health, Bethesda, MD, USA).
Enzyme assay
Enzyme activities were measured spectrophotometrically. All enzymatic assays were carried out at 30°C for 30 min using saturating concentrations of substrates and cofactors.
CS activity was measured at 412 nm to detect the transfer of sulfhydryl groups to 5,5’-dithiobis(2-nitrobenzonic acid) (DTNB). The extinction coefficient for DTNB, which is a reference for CS activity, was 13.6.
The activity of malate dehydrogenase (MDH), β-hydroxyacyl-coenzyme A dehydrogenase (βHAD), hexokinase (HK), pyruvate kinase (PK), and lactate dehydrogenase (LDH) was measured at 340 nm by monitoring the production or consumption of the reduced form of nicotinamide adenine dinucleotide (NADH) or the reduced form of nicotinamide adenine dinucleotide phosphate (NADPH). The extinction coefficient for NAD(P)H, which is the reference for these enzyme activities, was 6.22. Further details of these methods have been described previously (Suwa et al., 2008).
Statistical analysis
All data are expressed as the means ± SE. The unpaired t-test was used to compare the data between the groups. A value of p < 0.05 was considered to indicate statistical significance.
Results
Body composition
Table 1 shows the body composition of the rats. The pre- and post-treatment body weights in the L-NAME group were identical to those in the control group. The weights of the soleus and plantaris muscles and the epididymal fat tissue in the L-NAME group were not significantly different from those in the control group.
Table 1.
Body composition of the rats. Data are means (±SE).
| Control (n = 10) |
L-NAME (n = 10) |
|
|---|---|---|
| Pre-treatment body weight, g | 287(2) | 291(2) |
| Post-treatment body weight, g | 471(9) | 492(14) |
| Soleus weight, mg | 168(5) | 168(7) |
| Plantaris weight, mg | 443(12) | 466(12) |
| Epididymal fat tissue weight, g | 7.31 (.32) | 7.82 (.73) |
Muscle fiber composition
In myosin ATPase (preincubation pH 4.3)-stained sections of soleus muscle (Figure 1a, i and ii), the proportion of lightly stained Type II fibers in the L-NAME group was apparently higher than that in the control group. As shown in Figure 1b, the percentage of Type I fibers in the L-NAME group was significantly lower than that in the control group (79.7 ± 3.4 and 92.6 ± 0.7%, respectively, P < 0.01). The percentage of Type IIA fibers in the L-NAME group was significantly higher than that in the control group (18.1 ± 3.2 and 6.2 ± 0.5%, respectively, P < 0.01). No Type IIX or IIB fibers were observed in the soleus muscle. In the plantaris muscle, no differences were observed between the groups in the composition of the any fiber types (Figure 1c).
Figure 1.
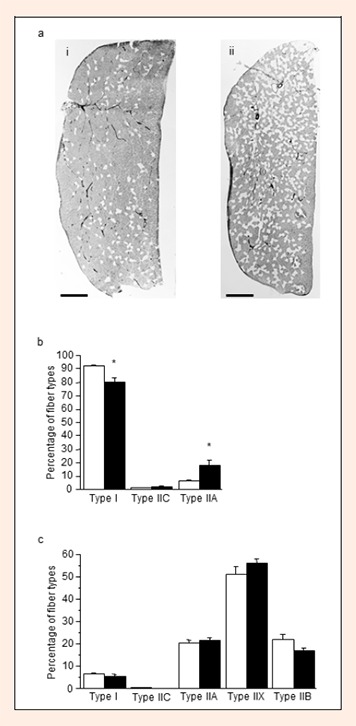
Muscle fiber composition. (a) Representative transverse sections of soleus muscles stained with myosin ATPase preincubated at pH 4.3 from the control (i) and L-NAME-treated (ii) groups. Dark and light fibers are Types I and II, respectively. Scale bar, 500 μm. b and c, Percentages of the fiber types of soleus (b) and plantaris (c) muscles in the control (open columns) and L-NAME-treated (filled columns) groups. Values represent means ± SE; n = six to nine muscles per group. * P < 0.05 vs. control group.
Expression of metabolic components
L-NAME treatment decreased the expression of cytochrome C protein in the plantaris muscle (–18%, P < 0.05), but not in the soleus muscle (Figure 2a). The expression levels of GLUT4 protein in the soleus and plantaris muscles decreased significantly with L-NAME treatment (Figure 2b, –14% and –22%, respectively, compared with the control, P < 0.005.
Figure 2.
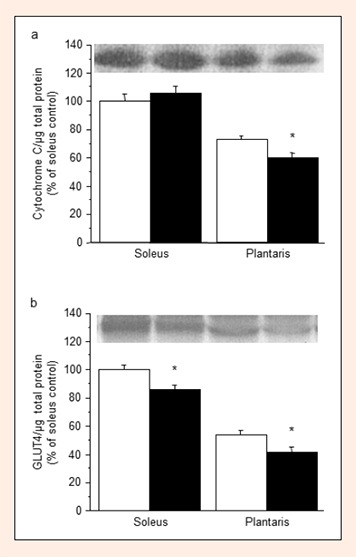
Expression levels of cytochrome C (a) and GLUT4 (b) proteins in the soleus and plantaris muscles in the control (open columns) and L-NAME-treated (filled columns) groups. Values represent means ± SE; n = 10 muscles per group. * P < 0.05 vs. control group.
Enzyme activities
Figure 3 shows the metabolic enzyme activities in the control and L-NAME groups. Concerning the mitochondrial oxidative enzymes, there was no difference between the groups in the activity of CS, a marker of the tricarboxylic acid cycle, in any of the muscles (Figure 3a). The activity of MDH, also a marker of the tricarboxylic acid cycle, of the plantaris muscle in the L-NAME group was significantly lower than that in the control group (–9%, P < 0.05); however, the MDH activity in the soleus muscle was not (Figure 3b). The activity of βHAD, a marker of β-oxidation, decreased with L-NAME treatment in the plantaris muscle (–18%, P < 0.01), but not in the soleus muscle (Figure 3c).
Figure 3.
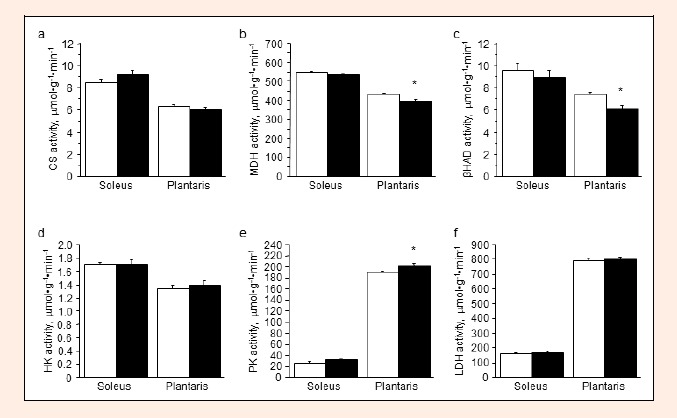
Metabolic enzyme activity in the soleus and plantaris muscles: CS (a), MDH (b), βHAD (c), HK (d), PK (e) and LDH (f) in the control (open columns) and L-NAME-treated (filled columns) groups. Values represent means ± SE; n = 10 muscles per group. * P < 0.05 vs. control group.
Concerning the enzymes associated with the glucose metabolism and glycolysis, the HK activity, which is an indicator of blood glucose mobilization, did not differ significantly between the groups in the muscles examined (Figure 3d). In the plantaris muscle, the activity of PK, a marker of glycolysis, was increased by L-NAME treatment (+6%, P <0.05), while that in the soleus muscle was not (Figure 3e). L-NAME treatment did not affect the activity of LDH, also a marker of glycolysis, in either muscle (Figure 3f).
Expression of metabolic regulators
The expression levels of SIRT1 protein in the soleus and plantaris muscles decreased significantly with L-NAME treatment (Figure 4a, –15% and –10%, respectively, compared with the control, P < 0.05). PGC-1α protein expression was not altered by L-NAME treatment in either muscle (Figure 4b).
Figure 4.
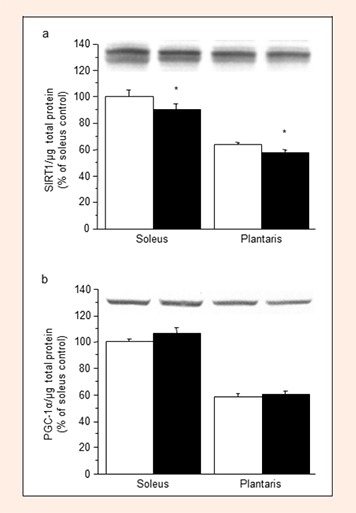
Expression levels of SIRT1 (a) and PGC-1α (b) proteins in the soleus and plantaris muscles in the control (open columns) and L-NAME-treated (filled columns) groups. Values represent means ± SE; n = 10 muscles per group. * P < 0.05 vs. control group.
Discussion
The current study demonstrates that treatment with the NOS inhibitor, L-NAME, for 8 weeks significantly increases the percentage of Type IIA fibers, with a concomitant decrease in the percentage of Type I fibers in the slow-twitch fiber-rich soleus muscle. These data suggest that NOS activity plays a role in sustaining the slow fiber phenotype of the soleus muscle under basal conditions. In contrast, L-NAME treatment does not affect the proportion of any of the fiber types in the fast-twitch fiber-rich plantaris muscle in rats. These results are consistent with previous reports that did not demonstrate any change in the proportion of the MHC isoforms in the plantaris muscle of the sham-operated leg following L-NAME treatment in rats (Sellman et al., 2006; Smith et al., 2002).
It can be speculated that the differences in the muscle fiber composition of the soleus and plantaris muscles following L-NAME treatment observed in this study are associated with differences in muscle activity and/or recruitment levels under sedentary conditions. The previous studies demonstrated that forced hyperactivity of the plantaris muscle, such as chronic overload and chronic low-frequency stimulation, induced fast-to-slow muscle fiber-type conversion, whereas L-NAME treatment significantly blocked such changes (Martins et al., 2012; Sellman et al., 2006; Smith et al., 2002). The soleus muscle works as a postural and anti-gravitational muscle. The activity of the slow soleus muscle, evaluated by the blood flow under basal sedentary conditions, is much higher than that of fast-twitch muscle (Laughlin and Armstrong, 1982). Collectively, these findings indicate that NOS activity promotes or maintains the slow muscle phenol-type, especially in a chronically active state.
A potential explanation for the observed changes in the muscle fiber composition in the soleus muscle following L-NAME treatment is that the nuclear factor of activated t-cells (NFAT) mediates such alterations. For example, NO promotes the transcriptional activation of NFAT in myotubes (Drenning et al., 2008). L-NAME treatment significantly blocks the electrical stimulation-induced dephosphorylation of NFAT in the extensor digitorum longus muscle in rats (Martins et al., 2012). Dephosphorylated NFAT translocates to the nucleus, and binds to specific sequences in the promoters of its target genes related to the slow muscle fiber-specific phenotype (Calabria et al., 2009; Meissner et al., 2007). It is, therefore, possible that L-NAME treatment induces transcriptional activation of NFAT followed by alteration in the fiber-type composition in the soleus muscle.
NOS/NO signaling can be associated with mitochondrial biogenesis in skeletal muscle cells. Previous studies using cultured muscle cells showed that treatment with NO donors, such as DETA-NO and SNAP, increases the expression of mitochondrial components and transcriptional regulators associated with mitochondrial biogenesis (Lira et al., 2010; McConell et al., 2010; Nisoli et al., 2004), thus suggesting that NO is a potent signaling molecule associated with mitochondrial biogenesis. In this study, we found that 8 weeks of chronic NOS inhibition by L-NAME treatment significantly decreased the expression of cytochrome C protein and the activity of MDH and βHAD in the fast-twitch fiber-rich plantaris muscle, but not in the slow-twitch fiber-rich soleus muscle. These results suggest that NOS is necessary for sustaining the basal level of mitochondrial biogenesis in the fast-twitch fibers in vivo. Although the mechanisms underlying such differences between fast- and slow-twitch fibers are unclear, one potential explanation is that the different expression pattern of NOS isoforms between fiber types is associated with the different results of metabolic changes following L-NAME treatment. nNOS and eNOS proteins are preferentially expressed in fast- and slow-twitch fibers, respectively(Lau et al., 2000; Punkt et al., 2006). Moreover, the two NOS isoforms appear to be differentially involved in mitochondrial biogenesis under basal conditions (Wadley et al., 2007). Based on these results, it is likely that the effects of nNOS inhibition are mainly observed in plantaris muscle while those of eNOS inhibition dominantly occur in soleus muscle after L-NAME treatment.
The current study provides the novel finding that the inhibition of NOS by L-NAME treatment for 8 weeks decreases SIRT1 protein expression in the soleus and plantaris muscles. These observations suggest that NOS regulates SIRT1 expression in skeletal muscle. Nisoli et al. (2005) also demonstrated that the SIRT1 protein expression is associated with NOS/NO in other tissues, evident by an increase of SIRT1 protein expression in white adipocytes following treatment with NO donors, such as DETA-NO and SNAP. Furthermore, a caloric restriction-induced increase of SIRT1 protein expression in the white and brown adipose tissues, brain, liver, and heart was diminished or prevented in eNOS-/- mice. Collectively, these findings provide compelling evidence that SIRT1 expression is partially controlled by NOS/NO signaling in skeletal muscle, as well as other tissues. In contrast, an in vitro study demonstrated that the treatment of primary human myotubes with DATA-NO for 4 days did not affect the SIRT1 expression (Civitarese et al., 2007). These discrepancies might be related to the differences in the study designs.
Both NOS/NO and SIRT1 have been proposed as regulators of mitochondrial biogenesis (Gerhart-Hines et al., 2007; Lira et al., 2010; McConell et al., 2010; Nisoli et al., 2004; Rodgers et al., 2005). The present study demonstrates that chronic inhibition of NOS reduces SIRT1 protein expression, with a concomitant decrease in the mitochondrial components in the plantaris muscle. In contrast, the inhibition of NOS reduces SIRT1 expression with no changes in mitochondrial biogenesis in soleus muscle. From these results, we can hypothesize that the SIR1 protein expression level is not crucial for the maintenance of mitochondrial biogenesis in soleus muscle. Mitochondrial biogenesis was previously associated with SIRT1 activity but not SIRT1 expression (Gurd et al., 2011), suggesting that SIRT1 activity is more important for the regulation of mitochondrial biogenesis than SIRT1 protein expression levels. It is likely that L-NAME treatment decreases SIRT1 expression with a coincidental decrease of SIRT1 activity in plantaris muscle, but does not affect SIRT1 activity in soleus muscle. Another possible explanation for the inconsistent results between SIRT1 expression and mitochondrial biogenesis is that unknown compensatory mechanisms sustain the basal level of mitochondrial biogenesis in soleus but not in plantaris muscle.
PGC-1α is a transcriptional coactivator that controls mitochondrial biogenesis in skeletal muscle (Benton et al., 2008; Lin et al., 2002; 2004). It has, therefore, been speculated that PGC-1α expression levels are associated with mitochondrial biogenesis in skeletal muscle. Previous studies suggest that PGC-1α expression is induced by NOS/NO signaling in skeletal muscle cells, as well as in other cells, with a concomitant increase in mitochondrial biogenesis (Lira et al., 2010; Nisoli et al., 2003; 2004; 2005; Wadley and McConell, 2007). We hypothesized that NOS inhibition reduces PGC-1α expression. However, this study demonstrated that L-NAME treatment did not affect the PGC-1α protein content in either the soleus or plantaris muscles, whereas it decreased the mitochondrial enzyme activity and cytochrome C protein content in the plantaris muscle. Presumably, redundant mechanisms maintain the PGC-1α level under such conditions of NOS/NO signal attenuation. However, PGC-1α is deacetylated and functionally activated by SIRT1 (Gerhart-Hines et al., 2007; Rodgers et al., 2005). Therefore, it can be postulated that reduced SIRT1 protein expression leads to downregulation of the deacetylation/activation levels of PGC-1α and a subsequent decrease in mitochondrial biogenesis, independent of the PGC-1α expression level in the plantaris muscle of L-NAME-treated rats.
The current study demonstrates that NOS inhibition by L-NAME treatment reduces GLUT4 protein expression in the soleus and plantaris muscles of rats. These results suggest that NOS/NO maintains the basal level of GLUT4 expression. A previous study demonstrated that treatment with a cGMP analog, 8 Br-cGMP, and a NO donor, SNAP, both promoted GLUT4 expression, leading to AMPK phosphorylation, in L6 myotubes (Lira et al., 2007). AMPK controls GLUT4 expression in skeletal muscle (Ojuka et al., 2000; Suwa et al., 2011; 2015). It is likely that NO plays a role in the regulation of GLUT4 expression through cGMP- and AMPK-dependent mechanisms. Interestingly, NO also regulates the translocation of GLUT4 to the plasma membrane and promotes glucose transport into skeletal muscle cells (McConell et al., 2012). These results indicate that NO controls glucose metabolism in skeletal muscle by promoting both the expression and translocation of GLUT4.
Conclusion
Eight weeks of treatment with the NOS inhibitor, L-NAME, in rats induced the slow-to-fast fiber-type transformation in the soleus muscle. In the plantaris muscle, NOS inhibition reduced the levels of mitochondrial components, such as oxidative enzyme activity and cytochrome C protein expression. L-NAME treatment reduced SIRT1 protein expression in both the soleus and plantaris muscles. NOS inhibition also decreased GLUT4 protein expression in both muscles. These results suggest that NOS/NO signaling plays a role in maintaining the basal levels of muscle fiber composition and components of mitochondrial biogenesis in skeletal muscle. Additionally, it can be speculated that such changes induced by L-NAME treatment are partially associated with reduced SIRT1 protein expression. It is likely that such roles of NOS/NO contribute muscle metabolic capacity and endurance exercise performance.
Acknowledgements
This work was supported by a Grant-in-Aid for Young Scientists from the Ministry of Education, Culture, Sports, Science and Technology of Japan (No. 20700524) to MS.
Biographies
Masataka SUWA
Employment
Tohoku Institute of Technology
Degree
PhD
Research interests
Exercise Physiology
E-mail: suwa-m@tohtech.ac.jp
Hiroshi NAKANO
Employment
Nakamura Gakuen University
Degree
PhD
Research interests
Exercise Physiology
E-mail: nakahi@nakamura-u.ac.jp
Zsolt RADAK
Employment
Semmelweis University
Degree
PhD
Research interests
Exercise Physiology
E-mail: radak@tf.hu
Shuzo KUMAGAI
Employment
Kyushu University
Degree
PhD
Research interests
Exercise Epidemiology and Physiology
E-mail: shuzo@ihs.kyushu-u.ac.jp
References
- Balon T.W., Nadler J.L. (1994) Nitric oxide release is present from incubated skeletal muscle preparations. Journal of Applied Physiology 77, 2519-2521. [DOI] [PubMed] [Google Scholar]
- Benton C.R., Nickerson J.G., Lally J., Han X.X., Holloway G.P., Glatz J.F., Luiken J.J., Graham T.E., Heikkila J.J., Bonen A. (2008) Modest PGC-1α overexpression in muscle in vivo is sufficient to increase insulin sensitivity and palmitate oxidation in subsarcolemmal, not intermyofibrillar, mitochondria. Journal of Biological Chemistry 283, 4228-4240. [DOI] [PubMed] [Google Scholar]
- Boutant M., Cantó C. (2014) SIRT1 metabolic actions: Integrating recent advances from mouse models. Molecular Metabolism 3, 5-18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Calabria E., Ciciliot S., Moretti I., Garcia M., Picard A., Dyar K.A., Pallafacchina G., Tothova J., Schiaffino S., Murgia M. (2009) NFAT isoforms control activity-dependent muscle fiber type specification. Proceedings of the National Academy of Sciences of the United States of America 106, 13335-13340. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Civitarese A.E., Carling S., Heilbronn L.K., Hulver M.H., Ukropcova B., Deutsch W.A., Smith S.R., Ravussin E., CALERIE Pennington Team (2007) Calorie restriction increases muscle mitochondrial biogenesis in healthy humans. PLoS Medicine 4, e76. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Drenning J.A., Lira V.A., Simmons C.G., Soltow Q.A., Sellman J.E., Criswell D.S. (2008) Nitric oxide facilitates NFAT-dependent transcription in mouse myotubes. American Journal of Physiology Cell Physiology 294, C1088-C1095. [DOI] [PubMed] [Google Scholar]
- Gerhart-Hines Z., Rodgers J.T., Bare O., Bare O., Lerin C., Kim S.H., Mostoslavsky R., Alt F.W., Wu Z., Puigserver P. (2007) Metabolic control of muscle mitochondrial function and fatty acid oxidation through SIRT1/PGC-1α. EMBO Journal 26, 1913-1923. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ghafourifar P., Cadenas E. (2005) Mitochondrial nitric oxide synthase. Trends in Pharmacological Sciences 26, 190-195. [DOI] [PubMed] [Google Scholar]
- Gollnick P.D., Armstrong R.B., Saubert C.W., IV, Piehl K., Saltin B. (1972) Enzyme activity and fiber composition in skeletal muscle of untrained and trained men. Journal of Applied Physiology 33, 312-319. [DOI] [PubMed] [Google Scholar]
- Gurd B.J., Yoshida Y., McFarlan J.T., Holloway G.P., Moyes C.D., Heigenhauser G.J., Spriet L., Bonen A. (2011) Nuclear SIRT1 activity, but not protein content, regulates mitochondrial biogenesis in rat and human skeletal muscle. American Journal of Physiology Regulatory Integrative and Comparative Physiology 301, R67-R75. [DOI] [PubMed] [Google Scholar]
- Higaki Y., Hirshman M.F., Fujii N., Goodyear L.J. (2001) Nitric oxide increases glucose uptake through a mechanism that is distinct from the insulin and contraction pathways in rat skeletal muscle. Diabetes 50, 241-247. [DOI] [PubMed] [Google Scholar]
- Lau K.S., Grange R.W., Isotani E., Sarelius I.H., Kamm K.E., Huang P.L., Stull J.T. (2000) nNOS and eNOS modulate cGMP formation and vascular response in contracting fast-twitch skeletal muscle. Physiological Genomics 2, 21-27. [DOI] [PubMed] [Google Scholar]
- Laughlin M.H., Armstrong R.B. (1982) Muscular blood flow distribution patterns as a function of running speed in rats. American Journal of Physiology 243, H296-H306. [DOI] [PubMed] [Google Scholar]
- Le Gouill E. Jimenez M. Binnert C. Jayet P.Y. Thalmann S. Nicod P. Scherrer U. Vollenweider P. (2007) Endothelial nitric oxide synthase (eNOS) knockout mice have defective mitochondrial β-oxidation. Diabetes 56, 2690-2696. [DOI] [PubMed] [Google Scholar]
- Lin J., Wu P.H., Tarr P.T., Lindenberg K.S., St-Pierre J., Zhang C.Y., Mootha V.K., Jäger S., Vianna C.R., Reznick R.M., Cui L., Manieri M., Donovan M.X., Wu Z., Cooper M.P., Fan M.C., Rohas L.M., Zavacki A.M., Cinti S., Shulman G.I., Lowell B.B., Krainc D., Spiegelman B.M. (2004) Defects in adaptive energy metabolism with CNS-linked hyperactivity in PGC-1α null mice. Cell 119, 121-135. [DOI] [PubMed] [Google Scholar]
- Lin J., Wu H., Tarr P.T., Zhang C.Y., Wu Z., Boss O., Michael L.F., Puigserver P., Isotani E., Olson E.N., Lowell B.B., Bassel-Duby R., Spiegelman B.M. (2002) Transcriptional co-activator PGC-1α drives the formation of slow-twitch muscle fibres. Nature 418, 797-801. [DOI] [PubMed] [Google Scholar]
- Lira V.A., Brown D.L., Lira A.K., Kavazis A.N., Soltow Q.A., Zeanah E.H., Criswell D.S. (2010) Nitric oxide and AMPK cooperatively regulate PGC-1 in skeletal muscle cells. Journal of Physiology 588, 3551-3566. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lira V.A., Soltow Q.A., Long J.H., Betters J.L., Sellman J.E., Criswell D.S. (2007) Nitric oxide increases GLUT4 expression and regulates AMPK signaling in skeletal muscle. American Journal of Physiology Endocrinology and Metabolism 293, E1062-E1068. [DOI] [PubMed] [Google Scholar]
- Martins K.J., St-Louis M., Murdoch G.K., MacLean I.M., McDonald P., Dixon W.T., Putman C.T., Michel R.N. (2012) Nitric oxide synthase inhibition prevents activity-induced calcineurin-NFATc1 signalling and fast-to-slow skeletal muscle fibre type conversions. Journal of Physiology 590, 1427-1442. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McConell G.K., Ng G.P., Phillips M., Ruan Z., Macaulay S.L., Wadley G.D. (2010) Central role of nitric oxide synthase in AICAR and caffeine-induced mitochondrial biogenesis in L6 myocytes. Journal of Applied Physiology 108, 589-595. [DOI] [PubMed] [Google Scholar]
- McConell G.K., Rattigan S., Lee-Young R.S., Wadley G.D., Merry T.L. (2012) Skeletal muscle nitric oxide signaling and exercise: a focus on glucose metabolism. American Journal of Physiology Endocrinology and Metabolism 303, E301-E307. [DOI] [PubMed] [Google Scholar]
- Meissner J.D., Umeda P.K., Chang K.C., Gros G., Scheibe R.J. (2007) Activation of the beta myosin heavy chain promoter by MEF-2D, MyoD, p300, and the calcineurin/NFATc1 pathway. Journal of Cell Physiology 211, 138-148. [DOI] [PubMed] [Google Scholar]
- Merry T.L., Lynch G.S., McConell G.K. (2010) Downstream mechanisms of nitric oxide-mediated skeletal muscle glucose uptake during contraction. American Journal of Physiology Regulatory Integrative and Comparative Physiology 299, R1656-R1665. [DOI] [PubMed] [Google Scholar]
- Nathan C., Xie Q.W. (1994) Nitric oxide synthases: roles, tolls, and controls. Cell 78, 915-918. [DOI] [PubMed] [Google Scholar]
- Nisoli E., Clementi E., Paolucci C., Cozzi V., Tonello C., Sciorati C., Bracale R., Valerio A., Francolini M., Moncada S., Carruba M.O. (2003) Mitochondrial biogenesis in mammals: the role of endogenous nitric oxide. Science 299, 896-899. [DOI] [PubMed] [Google Scholar]
- Nisoli E., Falcone S., Tonello C., Cozzi V., Palomba L., Fiorani M., Pisconti A., Brunelli S., Cardile A., Francolini M., Cantoni O., Carruba M.O., Moncada S., Clementi E. (2004) Mitochondrial biogenesis by NO yields functionally active mitochondria in mammals. Proceedings of the National Academy of Sciences of the United States of America 101, 16507-16512. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nisoli E., Tonello C., Cardile A., Cozzi V., Bracale R., Tedesco L., Falcone S., Valerio A., Cantoni O., Clementi E., Moncada S., Carruba M.O. (2005) Calorie restriction promotes mitochondrial biogenesis by inducing the expression of eNOS. Science 310, 314-317. [DOI] [PubMed] [Google Scholar]
- Ojuka E.O., Nolte L.A., Holloszy J.O. (2000) Increased expression of GLUT-4 and hexokinase in rat epitrochlearis muscles exposed to AICAR in vitro. Journal of Applied Physiology 88, 1072-1075. [DOI] [PubMed] [Google Scholar]
- Pereira Sant'Ana J.A., Ennion S., Sargeant A.J., Moorman A.F., Goldspink G. (1997) Comparison of the molecular, antigenic and ATPase determinants of fast myosin heavy chains in rat and human: a single-fibre study. Pflügers Archiv 435, 151-153. [DOI] [PubMed] [Google Scholar]
- Punkt K., Fritzsche M., Stockmar C., Hepp P., Josten C., Wellner M., Schering S., Buchwalow I.B. (2006) Nitric oxide synthase in human skeletal muscles related to defined fibre types. Histochemistry and Cell Biology 125, 567-573. [DOI] [PubMed] [Google Scholar]
- Roberts C.K., Barnard R.J., Jasman A., Balon T.W. (1999) Acute exercise increases nitric oxide synthase activity in skeletal muscle. American Journal of Physiology 277, E390-E394. [DOI] [PubMed] [Google Scholar]
- Rodgers J.T., Lerin C., Haas W., Gygi S.P., Spiegelman B.M., Puigserver P. (2005) Nutrient control of glucose homeostasis through a complex of PGC-1α and SIRT1. Nature 434, 113-118. [DOI] [PubMed] [Google Scholar]
- Ross R.M., Wadley G.D., Clark M.G., Rattigan S., McConell G.K. (2007) Local nitric oxide synthase inhibition reduces skeletal muscle glucose uptake but not capillary blood flow during in situ muscle contraction in rats. Diabetes 56, 2885-2892. [DOI] [PubMed] [Google Scholar]
- Schild L., Jaroscakova I., Lendeckel U., Wolf G., Keilhoff G. (2006) Neuronal nitric oxide synthase controls enzyme activity pattern of mitochondria and lipid metabolism. FASEB Journal 20, 145-147. [DOI] [PubMed] [Google Scholar]
- Sellman J.E., DeRuisseau K.C., Betters J.L., Lira V.A., Soltow Q.A., Selsby J.T., Criswell D.S. (2006) In vivo inhibition of nitric oxide synthase impairs upregulation of contractile protein mRNA in overloaded plantaris muscle. Journal of Applied Physiology 100, 258-265. [DOI] [PubMed] [Google Scholar]
- Smith L.W., Smith J.D., Criswell D.S. (2002) Involvement of nitric oxide synthase in skeletal muscle adaptation to chronic overload. Journal of Applied Physiology 92, 2005-2011. [DOI] [PubMed] [Google Scholar]
- Stamler J.S., Meissner G. (2001) Physiology of nitric oxide in skeletal muscle. Physiological Reviews 81, 209-237. [DOI] [PubMed] [Google Scholar]
- Suwa M., Kumagai S., Higaki Y., Nakamura T., Katsuta S. (2002) Dietary obesity-resistance and muscle oxidative enzyme activities of the fast-twitch fibre dominant rat. International Journal of Obesity and Related Metabolic Disorders 26, 830-837. [DOI] [PubMed] [Google Scholar]
- Suwa M., Nakano H., Higaki Y., Nakamura T., Katsuta S., Kumagai S. (2003) Increased wheel-running activity in the genetically skeletal muscle fast-twitch fiber-dominant rats. Journal of Applied Physiology 94, 185-192. [DOI] [PubMed] [Google Scholar]
- Suwa M., Nakano H., Radak Z., Kumagai S. (2008) Endurance exercise increases the SIRT1 and peroxisome proliferator-activated receptor γ coactivator-1α protein expressions in rat skeletal muscle. Metabolism Clinical and Experimental 57, 986-998. [DOI] [PubMed] [Google Scholar]
- Suwa M., Nakano H., Radak Z., Kumagai S. (2011) Short-term adenosine monophosphate-activated protein kinase activator 5-aminoimidazole-4-carboxamide-1-β-D-ribofuranoside treatment increases the sirtuin 1 protein expression in skeletal muscle. Metabolism Clinical and Experimental 60, 394-403. [DOI] [PubMed] [Google Scholar]
- Suwa M., Nakano H., Radak Z., Kumagai S. (2015) A comparison of chronic AICAR treatment-induced metabolic adaptations in red and white muscles of rats. Journal of Physiological Sciences 65, 121-130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Toledo J.C. Jr. and Augusto O. (2012) Connecting the chemical and biological properties of nitric oxide. Chemical Research in Toxicology 25, 975-989. [DOI] [PubMed] [Google Scholar]
- Wadley G.D., Choate J., McConell G.K. (2007) NOS isoform-specific regulation of basal but not exercise-induced mitochondrial biogenesis in mouse skeletal muscle. Journal of Physiology 585, 253-262. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wadley G.D., McConell G.K. (2007) Effect of nitric oxide synthase inhibition on mitochondrial biogenesis in rat skeletal muscle. Journal of Applied Physiology 102, 314-320. [DOI] [PubMed] [Google Scholar]
- Wang M.X., Murrell D.F., Szabo C., Warren R.F., Sarris M., Murrell G.A. (2001) Nitric oxide in skeletal muscle: inhibition of nitric oxide synthase inhibits walking speed in rats. Nitric Oxide 5, 219-232. [DOI] [PubMed] [Google Scholar]