

Abstract
Purpose
L-carnitine suppresses inflammatory responses in human corneal epithelial cells (HCECs) exposed to hyperosmotic stress. In this study, we determined if L-carnitine induces this protective effect through suppression of reactive oxygen species (ROS)-induced oxidative damage in HCECs.
Methods
Primary HCECs were established from donor limbal explants. A hyperosmolarity dry-eye model was used in which HCECs are cultured in 450 mOsM medium with or without L-carnitine for up to 48 hours. Production of reactive oxygen species (ROS), oxidative damage markers, oxygenases and antioxidative enzymes were analyzed by 2′,7′-dichlorofluorescein diacetate (DCFDA) kit, semiquantitative PCR, immunofluorescence, and/or Western blotting.
Results
Reactive oxygen species production increased in HCECs upon substitution of the isotonic medium with the hypertonic medium. L-carnitine supplementation partially suppressed this response. Hyperosmolarity increased cytotoxic membrane lipid peroxidation levels; namely, malondialdehyde (MDA) and hydroxynonenal (HNE), as well as mitochondria DNA release along with an increase in 8-OHdG and aconitase-2. Interestingly, these oxidative markers were significantly decreased by coculture with L-carnitine. Hyperosmotic stress also increased the mRNA expression and/or protein production of heme oxygenase-1 (HMOX1) and cyclooxygenase-2 (COX2), but inhibited the levels of antioxidant enzymes, superoxide dismutase-1 (SOD1), glutathione peroxidase-1 (GPX1), and peroxiredoxin-4 (PRDX4). However, L-carnitine partially reversed this altered imbalance between oxygenases and antioxidant enzymes induced by hyperosmolarity.
Conclusions
Our findings demonstrate for the first time that L-carnitine protects HCECs from oxidative stress by lessening the declines in antioxidant enzymes and suppressing ROS production. Such suppression reduces membrane lipid oxidative damage markers and mitochondrial DNA damage.
Keywords: L-carnitine, oxidative injury, hyperosmolarity, human corneal epithelial cells, dry eye
This study reveals that L-carnitine protects human corneal epithelial cells from oxidative stress by suppressing reactive oxygen species production and restoring antioxidative enzymes, which in turns reduces the cell oxidative damage markers in membrane lipid and mitochondrial DNA.
Dry eye is a multifactorial disease of the tears and ocular surface with symptoms of discomfort, visual disturbance, and tear film instability, which is often accompanied by tear film hyperosmolarity, inflammation of corneal and conjunctival epithelial cells, as well as decrease of conjunctival goblet cells and mucin production.1–5 Tear film hyperosmolarity has been considered as a key factor that initiates the ocular surface inflammation in dry-eye patients, as well as in mouse models.2,6–8 Our previous studies have also shown increased expression and production of proinflammatory cytokines (TNF-α, IL-1β, IL-6), chemokine (IL-8), and matrix metalloproteinases (MMP-13, -3, -9) in primary cultured human corneal epithelial cells (HCECs) exposed to hyperosmotic media.9–11
Current dry-eye therapies include tear supplementation, tear retention, tear volume stimulation, biological tear substitutes, antiinflammatory therapy, essential fatty acids, and environmental strategies, which improve dry-eye symptoms.12 However, many of them are palliative rather than disease-modifying, which often does not provide adequate symptom relief or prevent disease progression.13 Studies on pathogenic role of hyperosmolarity led to development of new preventive and therapeutic approaches to treat the patients with dry-eye syndrome. Osmoprotectants may become potential candidates based on their role in protecting cells from the effects of hyperosmolarity.6,14,15
Osmoprotectants are small organic molecules that are used in many cell types to restore isotonic cell volume and stabilize protein function, allowing adaptation to hyperosmolarity.6,16,17 Osmoprotectants are known as “organic osmolytes” or “compatible solutes,” and their uptake is accompanied by a decreasing concentration of intracellular inorganic salts.18–20 Recently, osmoprotectants, L-carnitine, erythritol, and betaine, have been demonstrated to inhibit in HCECs and in a dry-eye mouse model hyperosmotic-induced increases in proinflammatory cytokines (TNF-α, IL-1β, IL-6, and IL-17) and chemokines (IL-8, CCL2, and CCL20).15,21 We further showed their suppressive effects on the expression, production, and activity of matrix metalloproteinases (MMP-13, MMP-2, MMP-9, MMP-3, and MMP-7) induced by hyperosmotic media in primary HCECs.22 We also observed that L-carnitine and erythritol protected HCECs from hyperosmotic stress via suppressive effect on activation of c-Jun N-terminal kinases (JNK) and p38MAP kinase.23 However, the mechanism of their protective effect on the ocular surface remains to be elucidated.
Oxidative stress caused by overproduced reactive oxygen species (ROS) has been recognized to be an important mechanism involved in ocular surface inflammation and dry-eye disease, such as ROS-induced NLRP3 inflammasome activation, JNK pathway, CD95/CD95L apoptotic signaling activation, and so on.24–28 Reactive oxygen species are generated under normal physiological conditions from the mitochondrial electron transport chain and other sources, and play an important role in activating cellular signaling for survival. However, high levels of ROS cause oxidative stress and cell injury, including at least three reactions, lipid peroxidation of membranes, intracellular oxidative modification of proteins, and oxidative damage to DNA. These oxidative reactions form adducts with lipids, protein, and DNA, and lead to decreased functions of intracellular organelles and further damage.29,30 It is not clear whether and how hyperosmolarity induces oxidative injury to ocular surface epithelium. Here, we determined the protective cell defense effects of L-carnitine against oxidative damage induced by hyperosmotic stress in HCECs using an in vitro culture model of dry-eye disease.
Materials and Methods
Materials and Reagents
Cell culture dishes, plates, centrifuge tubes, and other plastic ware were purchased from BD Biosciences (Lincoln Park, NJ, USA); Dulbecco's modified Eagle's medium (DMEM), Ham F-12, amphotericin B, and gentamicin were from Invitrogen (Grand Island, NY, USA); fetal bovine serum (FBS) was from Hyclone (Logan, UT, USA); L-carnitine was from Sigma-Aldrich Corp. (St. Louis, MO, USA); RNeasy Plus Mini RNA extraction kit from Qiagen (Valencia, CA, USA); Ready-To-Go You-Prime First-Strand Beads were from GE Healthcare (Piscataway, NJ, USA); TaqMan gene expression assays and real-time PCR master mix were from Applied Biosystems (Foster City, CA, USA); 2′,7′-dichlorofluorescein diacetate (DCFDA)-Cellular Reactive Oxygen Species Detection Assay Kit, rabbit polyconal antibody against human malondialdehyde (MDA), 4–hydroxynonenal (HNE), aconitase-2, glutathione peroxidase-1 (GPX1), peroxiredoxin (PRDX) 4, and mouse monoclonal antibody against superoxide dismutase-1 (SOD1) were purchased from Abcam (Cambridge, MA, USA); rabbit polyclonal antibody against 8-hydroxy-2′-deoxyguanosine (8-OHdG), human heme oxygenase (HMOX)1, and cyclooxygenase (COX) 2 were from Santa Cruz Biotechnology (Santa Cruz, CA, USA); β-actin was from BioLegend (San Diego, CA, USA); and fluorescein Alexa-Fluor 488–conjugated secondary antibodies (donkey anti-goat IgG, donkey anti-rabbit, or goat anti-mouse IgG) were from Molecular Probes (Eugene, OR, USA; available in the public domain at http://www.invitrogen.com).
Primary Cultures of HCECs and In Vitro Hyperosmolarity Model
Human donor corneoscleral tissues (<72 hours after death) not suitable for clinical use, from donors aged 19 to 67 years, were obtained from the Lions Eye Bank of Texas (Houston, TX, USA). Primary HCECs were cultured in 12-well plates using explants from corneal limbal rims in a supplemented hormonal epidermal medium (SHEM) containing 5% FBS using our previous methods.31 Confluent primary corneal epithelial cultures in 14 to 18 days were switched to an equal volume (0.5 mL/well) of serum-free medium (SHEM without FBS) for 24 hours, and then treated for 4 or 24 hours with isosmolar (312 mOsM) and hyperosmolar media (450 mOsM), which was achieved by adding 69 mM sodium chloride (NaCl), with or without 1 hour prior incubation with 20 mM of L-carnitine. The osmolarity of the culture media was measured by a vapor pressure osmometer in the Body Fluid Chemistry Clinical Laboratory of the Methodist Hospital (Houston, TX, USA). The cells treated for 4 hours were lysed in RLT buffer from Qiagen RNeasy Plus Mini kit for RNA extraction. The cells treated for 24 hours were used for immunostaining or lysed in RIPA buffer for Western blot analysis.
RNA Extraction, Reverse Transcription, and Quantitative Real-Time PCR (RT-qPCR)
Total RNA was extracted with a RNeasy Plus Mini Kit according to the manufacturer's instructions, quantified with a spectrophotometer (NanoDrop ND-1000; Thermo Scientific, Wilmington, DE, USA), and stored at −80°C before use. The first strand cDNA was synthesized by RT from 1.0 μg total RNA using Ready-To-Go You-Prime First-Strand Beads, as previously described.9,32 Quantitative real-time PCR was performed in a Mx3005P QPCR System (Stratagene, La Jolla, CA, USA) with 20-μL reaction volume containing 5 μL cDNA, 1 μL gene expression assay, and 10 μL gene expression master mix (TaqMan; Applied Biosystems, Foster City, CA, USA). TaqMan gene expression assays used for this study were: glyceraldehyde 3-phosphate dehydrogenase (GAPDH; Hs99999905_m1), HMOX-1(Hs01110250_m1), and COX2 (Hs00153133_m1). The thermocycler parameters were 50°C for 2 minutes and 95°C for 10 minutes, followed by 40 cycles of 95°C for 15 seconds and 60°C for 1 minute. A nontemplate control was included to evaluate DNA contamination. The results were analyzed by the comparative threshold cycle (Ct) method and normalized by GAPDH as an internal control.33
Measurement of Cellular ROS Production
Cellular ROS production was measured using a DCFDA assay kit according to manufacturer's protocol. DCFDA, a cell-permeable fluorogenic dye, is deacetylated by cellular esterases to a nonfluorescent compound, and later oxidized by ROS into highly fluorescent 2′,7′-dichlorofluorescein (DCF), which measures hydroxyl, peroxyl, and other ROS activity within the cell. Human corneal epithelial cells were grown on the 96-well plates or 8-chamber slides. When the cells had reached confluence, they were washed twice with phosphate-buffered saline (PBS) and then incubated with 25 μM DCFDA in 1× buffer solution (provided in the kit) in 37°C incubator for 45 minutes. After washing twice with PBS, the cells were exposed to hyperosmotic media (350, 400, or 450 mOsM) obtained by supplementation of the isotonic 312 mOsM medium with NaCl (19, 44, or 69 mM, respectively) for different time periods (30–180 minutes). Cell images were taken under a fluorescence microscope. Cell fluorescence in 96-well plates was measured at 488-nm excitation and 525-nm emission using Tecan Infinite M200 Multimode Microplate Reader (Tecan US, Inc., Morrisville, NC, USA) after adding NaCl for 15 to 120 minutes. Relative changes of DCF fluorescence were expressed as the fold increase over untreated cells.
Immunofluorescent and Immunohistochemistry Staining
Human corneal epithelial cells on 8-chamber slides were fixed with freshly prepared 2% paraformaldehyde at 4°C for 10 minutes. Cells were permeabilized with 0.2% Triton X-100 in PBS at room temperature for 10 minutes. Indirect immunofluorescent and immunohistochemistry staining was performed with our previous methods.34,35 Primary antibody against human MDA, HNE, aconitase-2, 8-OHdG, HMOX-1, COX2, SOD1, GPX1, and PDRX4 were used. Alexa-Fluor 488 conjugated secondary antibodies were applied, and propidium iodide (PI) was used for nuclear counterstaining for immunofluorescence. The stained slides were photographed with Zeiss laser scanning confocal microscope (LSCM510META, Thornwood, NY, USA).
Western Blot Analysis
Western blot analysis was performed using a previously reported method.11 Equal amounts of protein measured by a BCA protein assay kit, were mixed with 6 × SDS reducing sample buffer and boiled for 10 minutes before loading. The proteins (50 μg/lane) were separated on an SDS-PAGE and transferred electronically to PVDF membranes. The membranes were blocked with 5% nonfat milk in Tris-Tween buffered saline (TTBS; mM Tris [pH 7.5], 0.9% NaCl, and 0.1% Tween-20) for 1 hour at room temperature and incubated with primary antibodies against MDA (1:200), HNE (1:200), aconitase-2 (1:200), HMOX-1 (1:200), COX2 (1:200), SOD1 (1:200), GPX1 (1:200), PDRX4 (1:200), or β-actin (1:1000) overnight at 4°C. After washing three times with Tris-buffered saline with 0.05% Tween 20 for 10 minutes each, the membranes were incubated with HRP conjugated goat anti-mouse IgG (1:1000) or goat anti-rabbit IgG (1:1000) for 1 hour at room temperature. The signals were detected with a chemiluminescence reagent (ECL; GE Healthcare), and the images were acquired by an imaging station (model 2000R; Eastman Kodak, Rochester, NY, USA).
Statistical Analysis
Student's t-test was used to compare differences between two groups. One-way ANOVA test was used to make comparisons among three or more groups, followed by Dunnett's post hoc test. P less than 0.05 was considered statistically significant.
Results
L-Carnitine Reduced Hyperosmolarity-Induced ROS Production
Reactive oxygen species generation started to increase at 60 minutes, reached a 2.08 ± 0.16-fold increase at 90 minutes, and 2.24 ± 0.09 at 2 hours after HCEC exposure to 450 mOsM medium. The increased production of ROS was partially but significantly suppressed to 1.79 ± 0.23-fold at 90 minutes, and 1.96 ± 0.21 at 2 hours (all P < 0.05) by prior incubation with 20 mM L-carnitine (Fig. 1A). Using fluorescence microscopy a significant increase of the DCF fluorescence intensity and DCF positive cells was observed in HCECs exposed to 450 mOsM medium for 90 minutes, compared with the normal control. However, the rise of DCF fluorescence was significantly less in the culture with hyperosmotic medium prior incubated with 20 mM L-carnitine (Fig. 1B).
Figure 1.
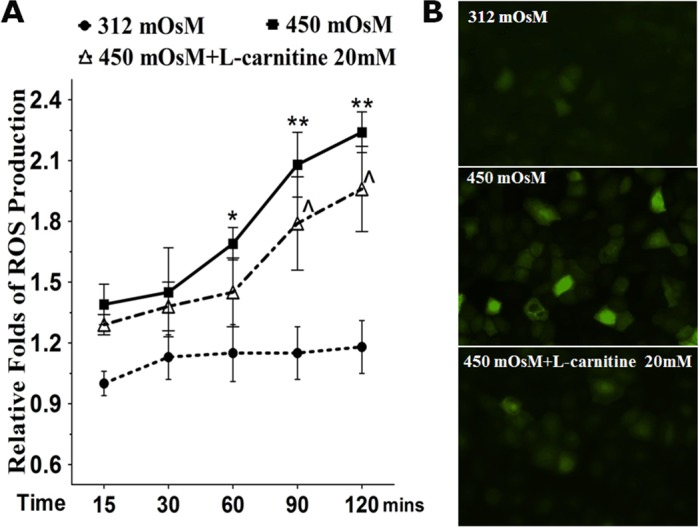
Production of ROS in primary HCECs exposed to hyperosmotic media without or with L-carnitine. (A) Time course graph displayed the relative fold of ROS production by HCECs in 312 or 450 mOsM medium with or without prior L-carnitine (20 mM) incubation. (B) Representative images showed ROS-DCF fluorescent-positive cells in HCECs exposed to media with 312 or 450 mOsM for 120 minutes. *P < 0.05, **P < 0.01, compared with 312 mOsM; ∧P < 0.05, as compared with 450 mOsM.
L-Carnitine Suppressed Lipid Peroxidation in HCECs Exposed to Hyperosmotic Media
Hypertonic media increased MDA and HNE production by approximately 3.79 ± 1.19 (P < 0.01) and 4.43 ± 0.83 (P < 0.01) fold, respectively, compared with normal controls (Figs. 2A, 2B). Immunohistochemical staining also showed increased stain in the cytoplasm and membrane of cells exposed to 450 mOsM media (Fig. 2C). Interestingly, 20 mM L-carnitine reduced MDA and HNE expression induced by hypertonic medium. Malondialdehyde and HNE declined by 58.32% and 58.47%, respectively, by Western blotting, and their immunoreactivity also declined.
Figure 2.

Oxidative biomarkers for cell membrane lipid peroxidation. (A, B) Western blot showed that MDA and HNE protein levels increased in primary HCECs exposed to hyperosmotic media, but decreased by cocultured with 20 mM of L-carnitine. **P < 0.01, compared with 312 mOsM; ∧P < 0.05, as compared with 450 mOsM. (C) Representative images showed the immunohistochemical staining of MDA and HNE in primary HCECs exposed to hyperosmotic media without or with L-carnitine.
L-Carnitine Prevented Oxidative DNA Damage Induced by Hyperosmolarity
The oxidized DNA product, 8-OHdG increased based on more intense brown staining localized to the cytoplasmic and nuclear compartment. However, prior incubation with L-carnitine partially reduced the intensity of 8-OHdG staining in both nuclear and cytoplasmic locations, the latter indicated reduced mitochondrial oxidative injury (Fig. 3A).
Figure 3.

Oxidative biomarker for DNA damage. (A) Representative images showed 8-OHdG immunohistochemical staining in primary HCECs exposed to hyperosmotic media without or with L-carnitine. (B, C) Western blot showed aconitase-2 protein levels in HCECs exposed to hyperosmotic media without or with L-carnitine pretreatment. **P < 0.01, compared with 312 mOsM; ∧∧P < 0.01, as compared with 450 mOsM. (D) Representative images showed the aconitase-2 immunohistochemical staining in primary HCECs exposed to hyperosmotic media without or with L-carnitine.
L-Carnitine Reduced Rises in Aconitase-2 Induced by Hyperosmolarity
Aconitase-2 protein expression increased markedly by a 5.20 ± 1.15-fold (P < 0.01) relative to that in iso-osmolar medium, but it decreased approximately 47% to 2.79 ± 1.37-fold (P < 0.01) by L-carnitine prior treatment (Figs. 3B, 3C). This finding was confirmed by immunohistochemical staining that showed the increased cytoplasmic staining by 450 mOsM, which was significantly decreased with prior incubation with 20 mM L-carnitine (Fig. 3D).
L-Carnitine Suppressed Production of Oxidative-Induced Oxygenases HMOX-1 and COX2
Figure 4A shows that HMOX1 and COX2 gene and protein expression rose along with immunofluorescence in hyperosmotic medium. The increases were 3.42 ± 0.82- (P < 0.05) and 10.58 ± 2.30-fold (P < 0.01), respectively. On the other hand, during exposure to 20 mM L-carnitine the increases declined by 40% to 2.04 ± 0.53- (P < 0.05) and 70% to 3.10 ± 0.48-fold (P < 0.05), respectively, from those in the absence of L-carnitine. HMOX-1 and COX2 immunoreactivity located mainly in the cytoplasm was significantly greater in the hyperosmotic medium than in the isotonic medium (Fig. 4B). However, the level of immunoreactivity in hyperosmotic medium was suppressed by 20 mM L-carnitine pretreatment. The increases in hyperosmolar medium correspond to increases in HMOX1 and COX2 protein expression of 5.77 and 10.86, respectively. However, a decrease of 42% to 3.32 ± 0.96- and 67% to 3.54 ± 1.06-fold was observed, respectively, in HCECs with L-carnitine pretreatment (Figs. 4C, 4D).
Figure 4.

The mRNA and protein levels of oxygenases HMOX-1 and COX2 in HCECs. (A) The mRNA expression levels of HMOX-1 and COX2 in HCECs exposed to normal or hyperosmolar media for 4 hours without or with prior L-carnitine (20 mM) incubation, as evaluated by RT-qPCR. (B) Protein levels of HMOX-1 and COX2 in HCECs exposed to normal or hyperosmolar media for 24 hours without or with prior L-carnitine (20 mM) incubation, as evaluated by immunofluorescence. (C, D) Protein production of HMOX-1 and COX2 in HCECs with same treatment as in (B) by Western blotting. *P < 0.05, **P < 0.01, compared with 312 mOsM; ∧P < 0.05, ∧∧P < 0.01, as compared with 450 mOsM.
L-Carnitine Rescued Production of Antioxidative Enzymes SOD1, GPX1, and PRDX4 Suppressed by Hyperosmolarity
Hyperosmotic stress reduced protein levels of antioxidative enzymes SOD1, GPX1, and PRDX4 to levels that were 0.42, 0.62, and 0.18, respectively, of those in isotonic medium. Interestingly, L-carnitine pretreatment blunted declines in the levels of these three antioxidative enzymes. They only fell to levels that were 0.86, 0.88, and 0.83 of those in isotonic medium (Figs. 5A, 5B). Immunocytochemical staining results further confirmed the protective effect of L-carnitine on declines in SOD1, GPX1, and PRDX4 levels in cells exposed to hyperosmotic medium (Fig. 5C).
Figure 5.

The production of antioxidative enzymes SOD1, GPX1, and PDRX4 in HCECs. (A, B) Western blot analysis showed the protein levels of SOD1, GPX1, and PDRX4 reduced in HCECs exposed to hyperosmolar media for 24 hours compared with normal HCECs, but rescued significantly by L-carnitine. *P < 0.05, **P < 0.01, compared with 312 mOsM; ∧P < 0.05, ∧∧P < 0.01, as compared with 450 mOsM. (C) Representative images showed immunohistochemical staining for SOD1, GPX1, and PDRX4 in HCECs with same treatment as above in (A).
Discussion
L-carnitine as an organic osmoprotectant19,20 has been proven to have protective roles against production of proinflammatory mediators21 and apoptosis16 in primary HCECs exposed to hyperosmotic media, as well as in dry-eye patients36,37 and animal models.15 However, the antioxidative effects of L-carnitine are not well understood in an in vitro dry-eye model. Here, comprehensive results from the present study show that L-carnitine is capable of reducing the impact of hyperosmolarity on HCEC function induced by increases in ROS levels. The diminution of rises in ROS caused less cell oxidative damage, a potential mechanism underlying rises in proinflammatory cytokine expression in dry-eye disease and in an in vitro model of this condition.
L-Carnitine Protected HCECs Against Overproduced ROS and Cell Oxidative Injuries in Lipid Peroxidation, Intracellular Protein Modification, and Oxidative DNA Damage
As a by-product of mitochondrial respiration, ROS can cause cell injury if over-produced. An imbalance between free radical-generating and scavenging results in oxidative stress, which is associated with the cell injury in many pathological conditions leading to corneal inflammation and dry eye. In the present study, we showed that hyperosmolarity induced excessive production of ROS, which caused cell oxidative damage; however, L-carnitine significantly suppressed ROS production, and in turn reduced oxidative damage in membrane lipid peroxidation, intracellular protein modification, and DNA oxidative damage.
Malondialdehyde and 4-HNE are major end-products of oxidation of polyunsaturated fatty acids, and their levels are frequently used as indicators of lipid peroxidation and oxidative stress. Malondialdehyde forms Schiff-base adducts with lysine residues and cross-links proteins in vitro; HNE also reacts with lysines, primarily via a Michael addition reaction.38,39 Malondialdehyde is a highly toxic molecule, and its interaction with DNA and proteins has often been referred to as potentially mutagenic and atherogenic.38 Hydroxynonenal is a toxic messenger of oxygen free radicals and undergoes reactions with proteins, peptides, phospholipids, and nucleic acids, which has cytotoxic, mutagenic, and genotoxic signal effects. They both have been identified as the most cytotoxic breakdown products generated from lipid peroxidation.40–42 The levels of HNE in tear and conjunctiva were observed to be significantly higher in Sjögren syndrome (SS) compared with healthy controls.43,44 In the present study, L-carnitine was found to significantly suppress the ROS production as evaluated by DCFDA assay (Fig. 1). L-carnitine also has a protective role by reducing MDA and HNE production as evaluated by Western blotting and immunohistochemical staining (Fig. 2).
Oxidative stress of mitochondria has been associated with many pathological processes influencing on cell death and survival such as apoptosis, autophagy, and plasticity. Aconitase activity and 8-OHdG levels are major markers of mitochondrial DNA damage. Our results showed hyperosmolarity increased production of both aconitase-2 and 8-OHdG in HCECs, as evaluated by immunohistochemical staining and Western blot analysis (Fig. 3). Interestingly, L-carnitine significantly suppressed these two markers at the protein level, indicating that L-carnitine protected the corneal epithelial cells from oxidative modification and DNA damage in mitochondria.
L-Carnitine Restored Balance Between Oxygenases and Antioxidative Enzymes Induced by Hyperosmolarity
HMOX1 and COX2 are two major oxygenases highly induced by a variety of agents causing oxidative stress. These molecules increase with inflammation and oxidative stress in dry eye.45 However, these oxygenases have not been well investigated in dry-eye models or patients. In our previous study, proinflammatory cytokines and chemokines, such as TNF-α, IL-1β, and IL-8, were found to be reduced by L-carnitine pretreatment in HCECs exposed to hyperosmotic media. This study further showed that the mRNA expression and protein production of HMOX1 and COX2 were markedly induced by hyperosmolarity, but significantly suppressed by L-carnitine (Fig. 4). The suppressive effect of L-carnitine on HMOX1 and COX2 not only protects HCECs from oxidative stress, but also serves as a mechanism by which it protects cells from inflammatory response to hyperosmotic stress.
Antioxidant enzymes form the first line of defense in organisms against free radicals and toxic reactants by metabolizing them to innocuous byproducts.46 Antioxidants are suggested as potential indirect markers of oxidative stress in ocular surface diseases such as dry eye. Oxidative stress has been speculated to cause antioxidant consumption that results in a decline in antioxidant level,47 Three major enzymes, SOD1, GPX1, and PRDX4, were investigated here. SODs, including cytosolic copper/zinc Cu, Zn-SOD (SOD1), mitochondrial Mn-SOD (SOD2), and extracellular SOD3, are antioxidant enzymes that catalyze the dismutation of superoxide into oxygen and hydrogen peroxide.48 GPX is an enzyme family with peroxidase activity whose main biological role is to protect the organism from oxidative damage by reducing lipid hydroperoxides to their corresponding alcohols and to reduce free hydrogen peroxide to water.49,50 PRDX is a ubiquitous family of antioxidant enzymes that uses reversibly oxidized catalytic cysteine residues to reduce peroxides, thus controlling peroxide levels and thereby mediate signal transduction in mammalian cells.51,52 The activities of SOD isoenzymes have been observed in tears, cornea, and the other parts of the human eye.48 GPX1 was found to be expressed in corneal and lens epithelia, and retina in human adult eye, and inhibits cellular inflammatory responses.50 However, few studies have investigated the role of these antioxidant enzymes in dry eye disease so far.
Our findings shown in Figure 5 revealed that hyperosmolarity interrupted the antioxidant defense system by reducing the production of antioxidant enzymes, such as SOD1, GPX1, and PRDX4, in HCECs exposed to hyperosmotic stimulation; however, L-carnitine largely rescued or restored the protein levels of these antioxidant enzymes. These data suggest that SOD1, GPX1, and PRDX4 may play an important antioxidative role during the oxidative stress induced by hyperosmolarity.
Oxidative stress biomarkers and antioxidants of corneal epithelial cells are believed to mirror the intrinsic oxidant/antioxidant balance of the ocular surface. Our data suggest that the oxidative cell damage may be caused by imbalance between the increased oxygenases HMOX1 and COX2 and suppressed antioxidant enzymes SOD1, GPX1, and PRDX4 in HCECs exposed to hyperosmotic media. Interestingly, our results showed that L-carnitine reversed the hyperosmolarity-induced courses by reducing two oxygenases and increasing three antioxidant enzymes. Taken together, our findings indicate that L-carnitine provides antioxidant protection by reversing an imbalance between the oxygenases and antioxidant enzymes.
In conclusion, our findings for the first time comprehensively revealed that L-carnitine protects HCECs from oxidative stress by suppressing ROS production, reducing the increased production of oxygenases HMOX1 and COX2, and restoring the inhibited protein levels of antioxidant enzymes SOD1, GPX1, and PRDX4, which in turns largely reduces the cell oxidative damage markers of lipid peroxidation of membranes (MDA and HNE), oxidative modification of proteins (aconitase 2), as well as oxidative damage to DNA (8-OHdG).
Follow-up studies are necessary to examine the clinical effect of L-carnitine application on ROS production or the consequences of its presence on the ocular surface.
Acknowledgments
The authors thank Terry Coursey, PhD, for his kind assistance with English editing of this manuscript.
Supported in part by National Institutes of Health Grants EY023598 (DQL), EY011915 (SCP), and Core Grant for Vision Research EY002520 (Bethesda, DC, USA); Allergan, Inc. (Irvine, CA, USA); Eye Bank Association of America (DQL; Washington, DC, USA); an unrestricted grant from Research to Prevent Blindness (New York, NY, USA); and the Oshman Foundation and the William Stamps Farish Fund (Houston, TX, USA).
Disclosure: X. Hua, None; R. Deng, None; J. Li, None; W. Chi, None; Z. Su, None; J. Lin, None; S.C. Pflugfelder, Allergan (F); D.-Q. Li, Allergan (F)
References
- 1. Brignole F,, Pisella PJ,, Goldschild M,, De Saint JM,, Goguel A,, Baudouin C. Flow cytometric analysis of inflammatory markers in conjunctival epithelial cells of patients with dry eyes. Invest Ophthalmol Vis Sci. 2000; 41: 1356–1363. [PubMed] [Google Scholar]
- 2. Julio G,, Lluch S,, Pujol P,, Merindano MD. Effects of tear hyperosmolarity on conjunctival cells in mild to moderate dry eye. Ophthalmic Physiol Opt. 2012; 32: 317–323. [DOI] [PubMed] [Google Scholar]
- 3. Stern ME,, Beuerman RW,, Fox RI,, Gao J,, Mircheff AK,, Pflugfelder SC. The pathology of dry eye: the interaction between the ocular surface and lacrimal glands. Cornea. 1998; 17: 584–589. [DOI] [PubMed] [Google Scholar]
- 4. Pflugfelder SC,, Solomon A,, Stern ME. The diagnosis and management of dry eye: a twenty-five-year review. Cornea. 2000; 19: 644–649. [DOI] [PubMed] [Google Scholar]
- 5. The definition and classification of dry eye disease: report of the Definition and Classification Subcommittee of the International Dry Eye WorkShop (2007). Ocul Surf. 2007; 5: 75–92. [DOI] [PubMed] [Google Scholar]
- 6. Baudouin C,, Aragona P,, Messmer EM,, et al. Role of hyperosmolarity in the pathogenesis and management of dry eye disease: proceedings of the OCEAN group meeting. Ocul Surf. 2013; 11: 246–258. [DOI] [PubMed] [Google Scholar]
- 7. Luo L,, Li D-Q,, Corrales RM,, Pflugfelder SC. Hyperosmolar saline is a proinflammatory stress on the mouse ocular surface. Eye Contact Lens. 2005; 31: 186–193. [DOI] [PubMed] [Google Scholar]
- 8. Liu H,, Begley C,, Chen M,, et al. A link between tear instability and hyperosmolarity in dry eye. Invest Ophthalmol Vis Sci. 2009; 50: 3671–3679. [DOI] [PubMed] [Google Scholar]
- 9. Luo L,, Li D-Q,, Doshi A,, Farley W,, Corrales RM,, Pflugfelder SC. Experimental dry eye stimulates production of inflammatory cytokines and MMP-9 and activates MAPK signaling pathways on the ocular surface. Invest Ophthalmol Vis Sci. 2004; 45: 4293–4301. [DOI] [PubMed] [Google Scholar]
- 10. Li D-Q,, Chen Z,, Song XJ,, Luo L,, Pflugfelder SC. Stimulation of matrix metalloproteinases by hyperosmolarity via a JNK pathway in human corneal epithelial cells. Invest Ophthalmol Vis Sci. 2004; 45: 4302–4311. [DOI] [PubMed] [Google Scholar]
- 11. Li D-Q,, Luo L,, Chen Z,, Kim HS,, Song XJ,, Pflugfelder SC. JNK, and ERK MAP kinases mediate induction of IL-1beta, TNF-alpha and IL-8 following hyperosmolar stress in human limbal epithelial cells. Exp Eye Res. 2006; 82: 588–596. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Pflugfelder SC,, Geerling G,, Kinoshita S,, et al. Management and therapy of dry eye disease: Report of the Management and Therapy Subcommittee of the International Dry Eye WorkShop (2007). Ocul Surf. 2007; 5: 163–178. [DOI] [PubMed] [Google Scholar]
- 13. Pflugfelder SC. Prevalence, burden, and pharmacoeconomics of dry eye disease. Am J Manag Care. 2008; 14: S102–S106. [PubMed] [Google Scholar]
- 14. Simmons PA,, Vehige JG. Clinical performance of a mid-viscosity artificial tear for dry eye treatment. Cornea. 2007; 26: 294–302. [DOI] [PubMed] [Google Scholar]
- 15. Chen W,, Zhang X,, Li J,, et al. Efficacy of osmoprotectants on prevention and treatment of murine dry eye. Invest Ophthalmol Vis Sci. 2013; 54: 6287–6297. [DOI] [PubMed] [Google Scholar]
- 16. Khandekar N,, Willcox MD,, Shih S,, Simmons P,, Vehige J,, Garrett Q. Decrease in hyperosmotic stress-induced corneal epithelial cell apoptosis by L-carnitine. Mol Vis. 2013; 19: 1945–1956. [PMC free article] [PubMed] [Google Scholar]
- 17. Garrett Q,, Khandekar N,, Shih S,, et al. Betaine stabilizes cell volume and protects against apoptosis in human corneal epithelial cells under hyperosmotic stress. Exp Eye Res. 2013; 108: 33–41. [DOI] [PubMed] [Google Scholar]
- 18. Yancey PH. Organic osmolytes as compatible, metabolic and counteracting cytoprotectants in high osmolarity and other stresses. J Exp Biol. 2005; 208: 2819–2830. [DOI] [PubMed] [Google Scholar]
- 19. Garrett Q,, Xu S,, Simmons PA,, Vehige J,, Flanagan JL,, Willcox MD. Expression and localization of carnitine/organic cation transporter OCTN1 and OCTN2 in ocular epithelium. Invest Ophthalmol Vis Sci. 2008; 49: 4844–4849. [DOI] [PubMed] [Google Scholar]
- 20. Xu S,, Flanagan JL,, Simmons PA,, Vehige J,, Willcox MD,, Garrett Q. Transport of L-carnitine in human corneal and conjunctival epithelial cells. Mol Vis. 2010; 16: 1823–1831. [PMC free article] [PubMed] [Google Scholar]
- 21. Hua X,, Su Z,, Deng R,, Lin J,, Li DQ,, Pflugfelder SC. Effects of l-carnitine, erythritol and betaine on pro-inflammatory markers in primary human corneal epithelial cells exposed to hyperosmotic stress. Curr Eye Res. 2014; 1–11. [DOI] [PMC free article] [PubMed]
- 22. Deng R,, Su Z,, Hua X,, Zhang Z,, Li D-Q,, Pflugfelder SC. Osmoprotectants suppress the production and activity of matrix metalloproteinases induced by hyperosmolarity in primary human corneal epithelial cells. Mol Vis. 2014; 20: 1243–1252. [PMC free article] [PubMed] [Google Scholar]
- 23. Corrales RM,, Luo L,, Chang EY,, Pflugfelder SC. Effects of osmoprotectants on hyperosmolar stress in cultured human corneal epithelial cells. Cornea. 2008; 27: 574–579. [DOI] [PubMed] [Google Scholar]
- 24. Pescosolido N,, Imperatrice B,, Karavitis P. The aging eye and the role of L-carnitine and its derivatives. Drugs R D. 2008; 9 (suppl 1): 3–14. [DOI] [PubMed] [Google Scholar]
- 25. Uchino Y,, Kawakita T,, Miyazawa M,, et al. Oxidative stress induced inflammation initiates functional decline of tear production. PLoS One. 2012; 7: e45805. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Zheng Q,, Ren Y,, Reinach PS,, et al. Reactive oxygen species activated NLRP3 inflammasomes prime environment-induced murine dry eye. Exp Eye Res. 2014; 125: 1–8. [DOI] [PubMed] [Google Scholar]
- 27. Zheng Q,, Ren Y,, Reinach PS,, et al. Reactive oxygen species activated NLRP3 inflammasomes initiate inflammation in hyperosmolarity stressed human corneal epithelial cells and environment-induced dry eye patients. Exp Eye Res. 2015; 134: 133–140. [DOI] [PubMed] [Google Scholar]
- 28. Chen Y,, Li M,, Li B,, Wang W,, Lin A,, Sheng M. Effect of reactive oxygen species generation in rabbit corneal epithelial cells on inflammatory and apoptotic signaling pathways in the presence of high osmotic pressure. PLoS One. 2013; 8: e72900. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Brambilla G,, Sciaba L,, Faggin P,, et al. Cytotoxicity, DNA fragmentation and sister-chromatid exchange in Chinese hamster ovary cells exposed to the lipid peroxidation product 4-hydroxynonenal and homologous aldehydes. Mutat Res. 1986; 171: 169–176. [DOI] [PubMed] [Google Scholar]
- 30. Esterbauer H,, Schaur RJ,, Zollner H. Chemistry and biochemistry of 4-hydroxynonenal, malonaldehyde and related aldehydes. Free Radic Biol Med. 1991; 11: 81–128. [DOI] [PubMed] [Google Scholar]
- 31. Kim HS,, Jun S,, de Paiva CS,, Chen Z,, Pflugfelder SC,, Li D-Q. Phenotypic characterization of human corneal epithelial cells expanded ex vivo from limbal explant and single cell cultures. Exp Eye Res. 2004; 79: 41–49. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Yoon KC,, de Paiva CS,, Qi H,, et al. Expression of Th-1 chemokines and chemokine receptors on the ocular surface of C57BL/6 mice: effects of desiccating stress. Invest Ophthalmol Vis Sci. 2007; 48: 2561–2569. [DOI] [PubMed] [Google Scholar]
- 33. Ma P,, Bian F,, Wang Z,, et al. Human corneal epithelium-derived thymic stromal lymphopoietin links the innate and adaptive immune responses via TLRs and Th2 cytokines. Invest Ophthalmol Vis Sci. 2009; 50: 2702–2709. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Chen Z,, de Paiva CS,, Luo L,, Kretzer FL,, Pflugfelder SC,, Li D-Q, Characterization of putative stem cell phenotype in human limbal epithelia. Stem Cells. 2004; 22: 355–366. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Li D-Q,, Tseng SC. Three patterns of cytokine expression potentially involved in epithelial-fibroblast interactions of human ocular surface. J Cell Physiol. 1995; 163: 61–79. [DOI] [PubMed] [Google Scholar]
- 36. Flanagan JL,, Simmons PA,, Vehige J,, Willcox MD,, Garrett Q. Role of carnitine in disease. Nutr Metab (Lond). 2010; 7: 30. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Pescosolido N,, Imperatrice B,, Koverech A,, Messano M. L-carnitine and short chain ester in tears from patients with dry eye. Optom Vis Sci. 2009; 86: E132–E138. [DOI] [PubMed] [Google Scholar]
- 38. Del RD,, Stewart AJ,, Pellegrini N. A review of recent studies on malondialdehyde as toxic molecule and biological marker of oxidative stress. Nutr Metab Cardiovasc Dis. 2005; 15: 316–328. [DOI] [PubMed] [Google Scholar]
- 39. Waldeck AR,, Stocker R. Radical-initiated lipid peroxidation in low density lipoproteins: insights obtained from kinetic modeling. Chem Res Toxicol. 1996; 9: 954–964. [DOI] [PubMed] [Google Scholar]
- 40. Esterbauer H,, Schaur RJ,, Zollner H. Chemistry and biochemistry of 4-hydroxynonenal malonaldehyde and related aldehydes. Free Radic Biol Med. 1991; 11: 81–128. [DOI] [PubMed] [Google Scholar]
- 41. Eckl PM,, Ortner A,, Esterbauer H. Genotoxic properties of 4-hydroxyalkenals and analogous aldehydes. Mutat Res. 1993; 290: 183–192. [DOI] [PubMed] [Google Scholar]
- 42. Uchida K. 4-Hydroxy-2-nonenal: a product and mediator of oxidative stress. Prog Lipid Res. 2003; 42: 318–343. [DOI] [PubMed] [Google Scholar]
- 43. Wakamatsu TH,, Dogru M,, Matsumoto Y,, et al. Evaluation of lipid oxidative stress status in Sjogren syndrome patients. Invest Ophthalmol Vis Sci. 2013; 54: 201–210. [DOI] [PubMed] [Google Scholar]
- 44. Andrade AS,, Salomon TB,, Behling CS,, et al. Alpha-lipoic acid restores tear production in an animal model of dry eye. Exp Eye Res. 2014; 120: 1–9. [DOI] [PubMed] [Google Scholar]
- 45. Higuchi A,, Inoue H,, Kawakita T,, Ogishima T,, Tsubota K. Selenium compound protects corneal epithelium against oxidative stress. PLoS One. 2012; 7: e45612. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Valentine JS,, Doucette PA,, Zittin PS. Copper-zinc superoxide dismutase and amyotrophic lateral sclerosis. Annu Rev Biochem. 2005; 74: 563–593. [DOI] [PubMed] [Google Scholar]
- 47. Polidori MC,, Stahl W,, Eichler O,, Niestroj I,, Sies H. Profiles of antioxidants in human plasma. Free Radic Biol Med. 2001; 30: 456–462. [DOI] [PubMed] [Google Scholar]
- 48. Behndig A,, Svensson B,, Marklund SL,, Karlsson K. Superoxide dismutase isoenzymes in the human eye. Invest Ophthalmol Vis Sci. 1998; 39: 471–475. [PubMed] [Google Scholar]
- 49. Zhang Y,, Zhang L,, Sun D,, Li Z,, Wang L,, Liu P. Genetic polymorphisms of superoxide dismutases, catalase, and glutathione peroxidase in age-related cataract. Mol Vis. 2011; 17: 2325–2332. [PMC free article] [PubMed] [Google Scholar]
- 50. Chang YC,, Chang WC,, Hung KH,, et al. The generation of induced pluripotent stem cells for macular degeneration as a drug screening platform: identification of curcumin as a protective agent for retinal pigment epithelial cells against oxidative stress. Front Aging Neurosci. 2014; 6: 191. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. Yamada S,, Ding Y,, Sasaguri Y. Peroxiredoxin 4: critical roles in inflammatory diseases. J UOEH. 2012; 34: 27–39. [DOI] [PubMed] [Google Scholar]
- 52. Veal EA,, Tomalin LE,, Morgan BA,, Day AM. The fission yeast Schizosaccharomyces pombe as a model to understand how peroxiredoxins influence cell responses to hydrogen peroxide. Biochem Soc Trans. 2014; 42: 909–916. [DOI] [PubMed] [Google Scholar]