

Abstract
Fibrosis and defective muscle regeneration can hamper the functional recovery of the soft palate muscles after cleft palate repair. This causes persistent problems in speech, swallowing, and sucking. In vitro culture systems that allow the study of satellite cells (myogenic stem cells) from head muscles are crucial to develop new therapies based on tissue engineering to promote muscle regeneration after surgery. These systems will offer new perspectives for the treatment of cleft palate patients. A protocol for the isolation, culture and differentiation of satellite cells from head muscles is presented. The isolation is based on enzymatic digestion and trituration to release the satellite cells. In addition, this protocol comprises an innovative method using extracellular matrix gel coatings of millimeter size, which requires only low numbers of satellite cells for differentiation assays.
Keywords: Developmental Biology, Issue 101, Head muscles, levator veli palatini muscle, digastric muscle, masseter muscle, satellite cells, isolation primary cells, cleft palate, regenerative medicine, tissue engineering, stem cells, differentiation, myofibers
Introduction
About 1:500 to 1:1,000 newborns exhibit a cleft involving the lip and/or palate (CLP); thus this is the most common congenital malformation in humans1. The muscles of the soft palate are critical for the functioning of the soft palate during speech, swallowing, and sucking. If a cleft of the soft palate is present, these muscles are abnormally inserted into the posterior end of the palatal bone.
The soft palate moves up and down during speech, preventing air to escape through the nose. Children with a cleft in the palate do not have this control function resulting in a phenomenon known as velopharyngeal dysfunction2,3. Although the treatment protocols are variable, surgical repair of the soft palate takes place in early childhood (6-36 months of age)4. The abnormally inserted muscles of the soft palate can be surgically corrected5-7, however, velopharyngeal dysfunction persists in 7% to 30% of the patients2,3,8-10.
The ability of skeletal muscle to regenerate through the action of satellite cells (SCs) is well established11,12. Upon muscle injury, SCs are activated and migrate to the site of injury. They then proliferate, differentiate, and fuse to form new myofibers or repair damaged ones13. Quiescent SCs express the transcription factor Pax714,15, while their progeny, the proliferating myoblasts, additionally express the myogenic determination factor 1 (MyoD)16. Differentiating myoblasts start to express myogenin (MyoG)17. The terminal differentiation of myoblasts is marked by the formation of myofibers, and the expression of muscle-specific proteins such as myosin heavy chain (MyHC)16,18.
Recently, several strategies have been used in regenerative medicine to improve muscle regeneration of limb muscles19-23. Specific studies on branchiomeric head muscles are also important because it was recently demonstrated that they differ from other muscles in several aspects24. In contrast with limb muscles, it has been suggested that branchiomeric head muscles contain less SCs25, regenerate slower, and more fibrous connective tissue is formed after injury26 In addition, proliferating SCs from branchiomeric head muscles also express other transcription factors. For instance, Tcf21, a transcription factor for craniofacial muscle formation is strongly expressed in regenerating head muscles but hardly in regenerating limb muscles25. The muscles in the soft palate of CLP patients are usually smaller and less well-organized compared to normal palatal muscles27,28. Slow and fast fibers are both present in the soft palate muscles but the slow fibers are more abundant. In contrast, cleft muscles contain a higher proportion of fast fibers and also a reduced capillary supply compared with normal soft palate muscles29-31. Fast fibers are more prone to contraction-induced injury31-33. The accompanying poor capillary supply may also promote fibrosis34,35. All these aspects may contribute to the poor regeneration of soft palate muscles after surgical cleft closure36. In view of this, a protocol for the isolation and characterization of branchiomeric head muscle SCs is crucial. This provides the possibility to study SC biology of branchiomeric head muscles. In addition, new therapies based on tissue engineering can be developed to promote muscle regeneration after surgery in CLP and other conditions compromising the craniofacial area.
In general, SCs can be obtained after dissociation of muscle tissue14. Mincing, enzymatic digestion, and trituration are generally required to release SCs from their niche. SCs can be purified by pre-plating on uncoated dishes14,37,38, fractionation on Percoll39,40, or fluorescent- or magnetic cell sorting41-43. Here we present a new economic and rapid protocol for the isolation of satellite cells from branchiomeric head muscles of young adult rats. This protocol is based on a previous manuscript14 and specifically adapted for small tissue samples. The isolation of SCs from representative muscles originating from the 1st, 2nd, and 4th branchial arches are described. After isolation, low numbers of satellite cells are cultured on extracellular matrix gel spots of millimeter size to study their differentiation. This approach avoids the requirement for the expansion and passaging of SCs.
Protocol
All experiments described herein were approved by the local Board for Animal Experiments from the Radboud University Nijmegen in accordance with Dutch laws and regulations (RU-DEC 2013-205).
1. Extracellular Matrix Gel Spots
- Perform the following steps one day before the isolation:
- Thaw an aliquot extracellular matrix gel (100 µl) at 4 °C for at least 1.5 hr. Dilute 1:10 in Dulbecco's modified Eagle's medium; with 4,500 mg/L glucose, 4 mM L-glutamine, and 110 mg/ml sodium pyruvate (DMEM). Keep the extracellular matrix gel at 4 °C at all times. Note: Abrupt temperature changes will result in uneven coating and crystal formation.
- Keep the diluted extracellular matrix gel solution on ice for 15 min.
- Pre-chill a 20 µl micropipette for 10 min.
- Put 8-well chamber slides into a 100 mm Petri dish and transfer the dish onto a cold surface (e.g. a freezer pack) for 10 min.
- Use the pre-chilled micropipette to put a drop of 10 µl extracellular matrix gel in each well. Keep the Petri dish on the cold surface for at least another 7 min (Figure 1A).
- Completely remove the remaining extracellular matrix gel (Figure 1B), and dry the wells at 37 °C overnight.
2. Dissection of Head Muscles (Masseter, Digastric, and Levator Veli Palatini)
Before dissection, prepare 50 ml of phosphate-buffered saline (PBS) supplemented with 2% Penicillin-Streptomycin (P/S). Keep on ice.
After euthanasia of one young adult rat (9 weeks) with CO2/O2, decapitate the head and remove the skin from the head. Transfer the head to ice-cold PBS supplemented with 2% P/S in a 50 ml tube.
- Masseter muscle (derived from the 1st branchial arch)
- Place the head with one side up on a silicone pad and fix with hypodermic needles (Figure 2A).
- Identify the parotid gland and the facial nerve (Figure 2A). Expose the deep fascia covering the gland. Cut the fascia and remove the gland using dissection scissors. Identify the external auditory canal. Trace the facial nerve from the stylomastoid foramen and carefully remove the temporal, zygomatic, and buccal branches with a scalpel blade No. 15.
- Free the superficial head of the masseter muscle by removing the fascia. Identify both superficial and deep heads of the masseter muscle. Trace the superficial head until its thick tendinous aponeurosis inserted in the zygomatic process of the maxilla.
- Separate the tendon from its origin at the zygomatic process with a straight forceps. Cut it with a scalpel blade No. 15 or dissection scissors and carefully life it (Figure 2B).
- Dissect the superficial head of the masseter until its insertion at the angle and inferior half of the lateral surface of the ramus of the mandible with a scalpel blade No. 15 (Figure 2C). Now, completely remove the muscle.
- Posterior belly of the digastric muscle (derived from the 2nd branchial arch)
- Place the head in a supine position on the silicone pad and fix with hypodermic needles (Figure 3A).
- Remove the subcutaneous fat overlying both sublingual and submandibular glands. Next, remove the superficial fascia and glands using dissection scissors. Expose the digastric muscle (anterior and posterior belly).
- Hold the anterior tendon of the posterior belly with a straight forceps, cut it, and dissect it carefully until its origin in the tympanic bulla (Figure 3B). Do the same at the contralateral side.
- Levator veli palatini muscle (derived from the 4th branchial arch)
- After dissection of the posterior belly of the digastric muscle, localize the stylohyoid muscle, pull it laterally, and carefully remove it (Figure 4A).
- Localize the tendon of the levator veli palatini that inserts at the tympanic bulla (Figure 4A). Dissect it carefully and cut it on both sides.
- Look for the trachea and the esophagus that runs behind it. Lift the esophagus, and expose the pharynx and the larynx.
- Localize and dissect the area of the superior pharyngeal constrictor muscle. Identify the levator veli palatini and cut it at both sides (Figure 4B). Note: Directly after dissection, carefully remove tendon and connective tissue from each muscle under the stereo microscope. Submerge all specimens quickly in ethanol 70%, and transfer them to ice-cold PBS supplemented 2% P/S in a 15 ml tube.
3. Isolation of Satellite Cells
- Perform the following preparation steps for SC isolation from 3 groups of muscles:
- Prepare 7.5 ml of 0.1% pronase in DMEM. Filter the solution through a 0.22 µm filter. Pre-warm the solution at 37 °C in a water bath for 10 min before isolation.
- Prepare 35 ml of DMEM supplemented with 10% Horse Serum (HS) and 1% P/S. Also pre-warm at 37 °C in a water bath.
- Prepare 15 ml culture medium which consists of DMEM supplemented with 20% fetal bovine serum (FBS), 10% HS, 1% P/S and 1% chicken embryo extract (CEE). Pre-warm at 37 °C in a water bath.
- Pre-coat six plastic pipettes (10 ml) with HS and dry for at least 10 min before use.
In the culture hood, transfer each muscle into a well of a 6-well plate. Using the dissection scissors, cut the muscle in small pieces of about 2 mm. Be careful not to mince the tissue too much.
Carefully add 2.5 ml of 0.1% pronase solution to each well and incubate at 37 °C for 60 min. Gently shake the plate after 20, 40, and 60 min. Note: The exact duration of the incubation depends on factors like age and strain of the animals.
Monitor under the microscope. Check the muscle fragments and stop the enzymatic digestion when the fibers bundles get a loosened appearance (Figure 5).
Add 2.5 ml of DMEM supplemented with 10% HS and 1% P/S. Transfer to a 15 ml tube and centrifuge the tubes at 400 x g for 5 min. Discard the supernatant by decantation.
Add 5 ml DMEM supplemented with 10% HS and 1% P/S. Pipette the solution up and down with a 10 ml plastic pipette (trituration) for at least 20 times to homogenize the tissue.
Centrifuge the tubes at 200 x g for 4 min. Collect the supernatant and transfer into a 15 ml tube.
Add 5 ml DMEM supplemented with 10% HS and 1% P/S. Pipette again with a 10 ml plastic pipette until the tissue fragments passes easily through the pipette.
Centrifuge the tubes at 200 x g for 4 min and collect the supernatant in a 15 ml tube.
Put a cell strainer (40 µm) onto a 50 ml tube and transfer the supernatant containing the dissociated cells onto the filter. Wash with 1 ml DMEM for maximal cell recovery.
Centrifuge the tubes at 1,000 x g for 10 min and discard the supernatant with a pipet.
Resuspend the pellet in 300 µl culture medium and count the cells in a hemocytometer.
4. Differentiation of Satellite Cells on Extracellular Matrix Gel Spots
Dilute the cell suspension to obtain 1.5 x 103 cells in 10 µl of culture medium.
Secure the covers of the chambers slides with tape and mark the spots with a black marker on the bottom side of the object glass.
Using a micropipette, put a drop of 10 µl cell suspension onto the extracellular matrix gel spot. Check under the microscope whether the drop of cell suspension has been placed correctly on the spot. Incubate for six hours at 37 °C.
Carefully add 400 µl of culture medium (DMEM supplemented with 20% FBS, 10% HS, 1% P/S and 1% CEE) and incubate for three days at 37 °C. Note: At this point, freshly isolated SC are subjected to massive trauma (enzymatic digest and harsh trituration) and they need to recover. Do not disturb the cells during the first three days 37. Next, the culture medium can be changed depending on the type of experiment. The extracellular matrix gel spots can be seeded with a high cell density (1.5-2.5 x 103/20 µl) for differentiation assay. Culture medium (DMEM supplemented with 20% FBS, 10% HS, 1% P/S and 1% chicken embryo extract) can be replaced every third day.
- Alternatively, if expansion and passing is desired follow the next steps:
- Thaw an aliquot extracellular matrix gel (500 µl) at 4 °C for at least 1.5 hr. Dilute 1:10 in DMEM and follow the recommendations in point 1.1.1.
- Pre-chill a 10 ml pipette for 10 min at 4 °C.
- Transfer three T75 flasks onto a cold surface (e.g. a freezer pack) for 10 min.
- Use the pre-chilled pipette to put 1 ml extracellular matrix gel into each flask. Check that the surface is covered completely. Keep the flasks on the cold surface for at least another 7 min (Figure 1A).
- Completely remove the remaining extracellular matrix gel with a 10 ml pipette, and dry the wells at 37 °C for 1 hr.
- After counting, resuspend the freshly isolated SCs in 10 ml of culture medium (DMEM supplemented with 20% FBS, 10% HS, 1% P/S and 1% chicken embryo extract) and seed in the pre-coated T75 flasks.
- After three days, change the medium (and every third day) until 80% confluence is reached. For passaging, wash the T75 flasks three times with PBS. Next add 1 ml 0.25% trypsin solution and incubate for three min at 37 °C. Resuspend in 9 ml of culture medium (DMEM supplemented with 20% FBS, 10% HS, 1% P/S and 1% chicken embryo extract) and centrifuge at 200 x g for 5 min. Discard the supernatant. After counting, resuspend 1 x 106 cells in 1,000 µl of culture medium and freeze the cells.
Representative Results
Using this protocol, the masseter muscle (one side) yields 0.8-1 x 106 cells, the digastric muscle (posterior belly) yields 1.5-2 x 105 cells, and levator veli palatini muscle yields 1-1.5 x 105 cells. Cell yields depend on the muscle type, strain, and age of the animal. For comparison between the three muscle groups, freshly isolated SCs were seeded at the same cell density (1.5 x 103/10 µl). Directly after isolation, more than 90% of the freshly isolated cells express Pax7 (Figure 6).
Day 4, 7 and 10 cultures were stained with antibodies against Pax7, MyoD, MyoG and MyHC immunostaining. Five arbitrary fields were counted per culture using a 20X objective. At day 4 Pax 7 and Myo D is expressed in all muscle groups (Figures 6 and 7 and 8), however the progeny of SatCs from the masseter and digastric muscles start expressing myogenin earlier than the levator veli palatini muscle (Figure 9). At day 10, the expression of MyoG is strongly reduced in all groups (Figure 9). A few days after seeding on the extracellular matrix gel spots, the proliferating cells begin to fuse and form multi-nucleated myotubes, which express myosin heavy chain. Small myotubes are clearly visible at day 7 (Figure 10). At day 10, twitching of the myotubes can be observed (Video 1).
 Figure 1: Extracellular matrix gel spots in a chamber slide. (A) For easy manipulation, place the 8-well chamber slide into a 100 mm Petri dish. Pipet 10 µl extracellular matrix gel in each chamber and put it on a cold surface (7 min). (B) Chamber slide after the excess extracellular matrix gel is removed.
Figure 1: Extracellular matrix gel spots in a chamber slide. (A) For easy manipulation, place the 8-well chamber slide into a 100 mm Petri dish. Pipet 10 µl extracellular matrix gel in each chamber and put it on a cold surface (7 min). (B) Chamber slide after the excess extracellular matrix gel is removed.
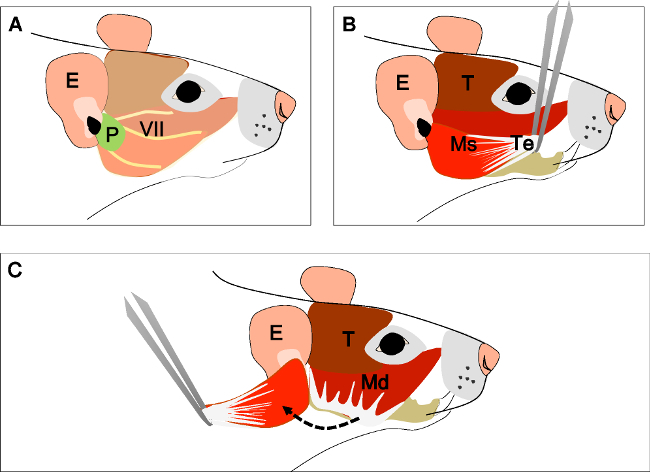 Figure 2: Dissection of the masseter muscle. (A) Head of the animal in a lateral view. Ear (E), Parotid gland (P) and facial nerve (VII). (B) Tendinous aponeurosis (Te) of the superficial head of the masseter muscle (Ms) and temporal muscle (T). Separate the tendon from its insertion with a forceps. (C) Carefully dissect the muscle until its insertion at the ramus of the mandible. E: ear, P: parotid gland, VII: facial nerve, T: Temporal muscle, Ms: superficial head of the masseter muscle, Te: tendon, Mp: deep head of the masseter muscle.
Figure 2: Dissection of the masseter muscle. (A) Head of the animal in a lateral view. Ear (E), Parotid gland (P) and facial nerve (VII). (B) Tendinous aponeurosis (Te) of the superficial head of the masseter muscle (Ms) and temporal muscle (T). Separate the tendon from its insertion with a forceps. (C) Carefully dissect the muscle until its insertion at the ramus of the mandible. E: ear, P: parotid gland, VII: facial nerve, T: Temporal muscle, Ms: superficial head of the masseter muscle, Te: tendon, Mp: deep head of the masseter muscle.
 Figure 3: Dissection of the posterior belly of the digastric muscle. (A) Head of the animal in a supine position. Localize the submandibular gland (Sg), masseter muscle (M), facial nerve (VII) and sternocleidomastoid muscle (SCM). Remove the submandibular gland. (B) Localize the digastric muscle anterior (AD) and posterior belly (PD). With a straight forceps, take the anterior tendon of the posterior belly, cut it and dissect it carefully until its origin in the tympanic bulla (ty). E: ear, Sg: submandibular gland, VII: facial nerve, M: masseter muscle, SMC: sternocleidomastoid muscle, AD: anterior belly digastric muscle, PD: posterior belly digastric muscle, Ty: Tympanic bulla.
Figure 3: Dissection of the posterior belly of the digastric muscle. (A) Head of the animal in a supine position. Localize the submandibular gland (Sg), masseter muscle (M), facial nerve (VII) and sternocleidomastoid muscle (SCM). Remove the submandibular gland. (B) Localize the digastric muscle anterior (AD) and posterior belly (PD). With a straight forceps, take the anterior tendon of the posterior belly, cut it and dissect it carefully until its origin in the tympanic bulla (ty). E: ear, Sg: submandibular gland, VII: facial nerve, M: masseter muscle, SMC: sternocleidomastoid muscle, AD: anterior belly digastric muscle, PD: posterior belly digastric muscle, Ty: Tympanic bulla.
 Figure 4: Dissection of the levator veli palatini muscle. (A) General view after dissection of the digastric muscle (posterior belly). Stylohyoid muscle (St) and tendon of the levator veli palatini can be localized. Note the trachea (T) and esophagus (Es) running behind it. (B) After lifting the trachea and the esophagus the pharynx (P) is exposed. The levator veli palatini that runs laterally towards the soft palate is now visible. The arrow indicates the dissected superior pharyngeal constrictor muscle; note the levator veli palatini muscles at both sides. E: ear, St: stylohyoid muscle, VII: facial nerve, M: masseter muscle, AD: anterior belly digastric muscle, PD: posterior belly digastric muscle, T: trachea, Es: esophagus, P: Pharynx, *levator veli palatini muscle.
Figure 4: Dissection of the levator veli palatini muscle. (A) General view after dissection of the digastric muscle (posterior belly). Stylohyoid muscle (St) and tendon of the levator veli palatini can be localized. Note the trachea (T) and esophagus (Es) running behind it. (B) After lifting the trachea and the esophagus the pharynx (P) is exposed. The levator veli palatini that runs laterally towards the soft palate is now visible. The arrow indicates the dissected superior pharyngeal constrictor muscle; note the levator veli palatini muscles at both sides. E: ear, St: stylohyoid muscle, VII: facial nerve, M: masseter muscle, AD: anterior belly digastric muscle, PD: posterior belly digastric muscle, T: trachea, Es: esophagus, P: Pharynx, *levator veli palatini muscle.
 Figure 5: Appearance of the muscle tissue (A) before and (B) after enzymatic digestion with pronase. Note that muscle bundles appear to be loosened after enzymatic digestion.
Figure 5: Appearance of the muscle tissue (A) before and (B) after enzymatic digestion with pronase. Note that muscle bundles appear to be loosened after enzymatic digestion.
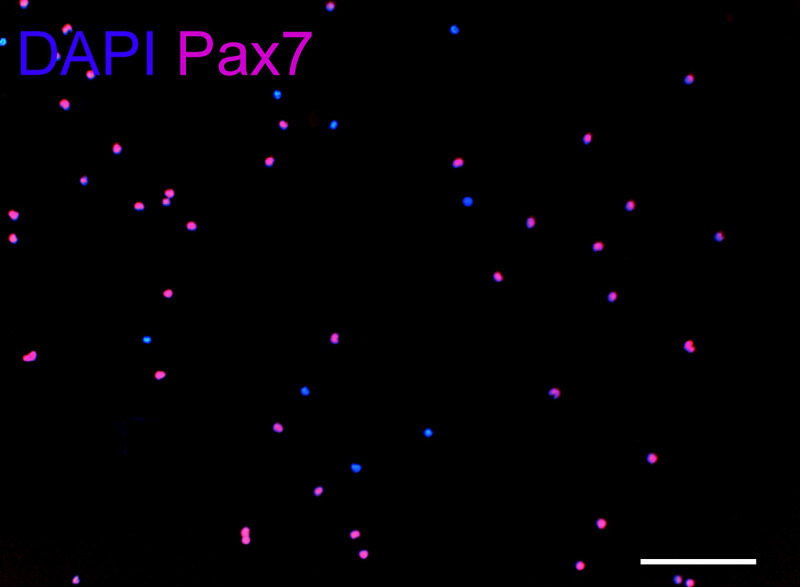 Figure 6: Pax 7 immunostaining. Freshly isolated SCs, applied to extracellular matrix gel at the end of isolation (about 6 hours after initial tissue digestion). Five arbitrary fields were counted using a 10X objective with an average of 210 cells per field. Approximately 90% of the cells are Pax 7 positive. DAPI: blue, Pax7: red. Scale bar, 100 µm.
Figure 6: Pax 7 immunostaining. Freshly isolated SCs, applied to extracellular matrix gel at the end of isolation (about 6 hours after initial tissue digestion). Five arbitrary fields were counted using a 10X objective with an average of 210 cells per field. Approximately 90% of the cells are Pax 7 positive. DAPI: blue, Pax7: red. Scale bar, 100 µm.
 Figure 7: Pax 7, MyoD immunostaining. Day 4, 7 and 10 cultures were stained with antibodies against Pax7, and MyoD immunostaining. (A–C) and (D–F) Representative photomicrographs of day 4 and 7 cultures from the masseter muscle. (G and H) The number of Pax7+ and MyoD+ nuclei per microscopic field was counted and expressed as a percentage of the total number of nuclei (DAPI). DAPI: blue, Pax7: red, and MyoD: green. Scales bar, 100 µm. Please click here to view a larger version of this figure.
Figure 7: Pax 7, MyoD immunostaining. Day 4, 7 and 10 cultures were stained with antibodies against Pax7, and MyoD immunostaining. (A–C) and (D–F) Representative photomicrographs of day 4 and 7 cultures from the masseter muscle. (G and H) The number of Pax7+ and MyoD+ nuclei per microscopic field was counted and expressed as a percentage of the total number of nuclei (DAPI). DAPI: blue, Pax7: red, and MyoD: green. Scales bar, 100 µm. Please click here to view a larger version of this figure.
 Figure 8: Distribution of Pax7±/MyoD± in cultures from mononucleated cells in cultures from masseter, digastric and levator veli palatine muscle. (A–C) Day 4, 7 and 10 cultures were stained with antibodies against Pax7, and MyoD immunostaining. The total number of cells is based on of the total number of nuclei (DAPI). (D) Data quantification of Pax7±/MyoD± cells. Please click here to view a larger version of this figure.
Figure 8: Distribution of Pax7±/MyoD± in cultures from mononucleated cells in cultures from masseter, digastric and levator veli palatine muscle. (A–C) Day 4, 7 and 10 cultures were stained with antibodies against Pax7, and MyoD immunostaining. The total number of cells is based on of the total number of nuclei (DAPI). (D) Data quantification of Pax7±/MyoD± cells. Please click here to view a larger version of this figure.
 Figure 9: Myogenin immunostaining. Day 4, 7 and 10 cultures were stained with antibodies against Myogenin. (A–D) Representative photomicrographs of day 4 and 7 cultures from the levator veli palatine muscle. (E) The number of MyoG+ nuclei per microscopic field was counted and expressed as a percentage of the total number of nuclei (DAPI). (F) Data quantification of MyoG+ cells. DAPI: blue, Myogenin: green. Scales bar, 100 µm. Please click here to view a larger version of this figure.
Figure 9: Myogenin immunostaining. Day 4, 7 and 10 cultures were stained with antibodies against Myogenin. (A–D) Representative photomicrographs of day 4 and 7 cultures from the levator veli palatine muscle. (E) The number of MyoG+ nuclei per microscopic field was counted and expressed as a percentage of the total number of nuclei (DAPI). (F) Data quantification of MyoG+ cells. DAPI: blue, Myogenin: green. Scales bar, 100 µm. Please click here to view a larger version of this figure.
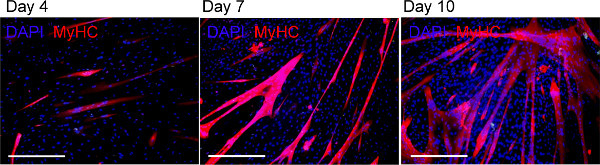 Figure 10: Myosin Heavy Chain immunostaining. Day 4, 7 and 10 cultures were stained with antibodies against myosin heavy chain (MyHC). Representative photomicrographs of day 4, 7 and 10 cultures from the digastric (DIG) muscle. At day 7, small myotubes are present while at day 10 long and well-organized myotubes are evident. Scales bar, 200 µm. Please click here to view a larger version of this figure.
Figure 10: Myosin Heavy Chain immunostaining. Day 4, 7 and 10 cultures were stained with antibodies against myosin heavy chain (MyHC). Representative photomicrographs of day 4, 7 and 10 cultures from the digastric (DIG) muscle. At day 7, small myotubes are present while at day 10 long and well-organized myotubes are evident. Scales bar, 200 µm. Please click here to view a larger version of this figure.
Video 1: Myotube twitching. Examples of two representative fields with twitching myotubes are shown for day 10 cultures from digastric muscle. Please click here to view this video.
Discussion
SCs from different branchiomeric head muscles were isolated from one 9-week-old Wistar rat and cultured directly on extracellular matrix gel spots without prior expansion and passaging. After isolation, the cells were counted and seeded at the same cell density. For the parallel isolation of three different muscles, this method takes about 4 hr. To avoid culture contamination, a critical step is the rapid washing in alcohol 70% after dissection of the muscles.
During SC isolation it is important to cut the muscle tissue into small pieces (about 2 mm) but avoid too much mincing as this will result in a small cell yield because of cell damage. Also, the duration of the enzymatic digestion must be checked carefully under the microscope to avoid further damage. The aim of the digestion is to dissociate the myofibers. Since more than 90% of the isolated cells express Pax7, no further purification is required (Figures 6-8). This avoids extra purification steps in other methods such as pre-plating on uncoated dishes14,37,38, fractionation on Percoll39,40, or fluorescent- or magnetic cell sorting41,43. For trituration it is essential to induce shear between the tissue fragments and the opening of the pipette tip as this allows the mechanical release of the SCs. If the trituration with a 10 ml pipette (inside diameter tip: 1 mm) is difficult, a 5 ml (inside diameter tip: 2 mm) pipette can be used first. Alternatively, glass Pasteur pipettes can be cut at the desired diameter and be used. This method is simple, efficient and allows the simultaneous isolation of SC from different muscle samples.
The culture plates for SCs can also be coated with gelatin or collagen, but our previous studies show that extracellular matrix gel is far better for the maintenance of the myogenic potential than collagen38. The extracellular matrix gel spots of millimeter size (10 µl/Ø 2 mm or 20 µl/Ø 4 mm) allows the study of proliferation and differentiation of SCs with limited numbers of cells. For the differentiation assay about 8 to 20 times fewer cells are required compared to a 24-well plate (Ø 15.6 mm), and about 80 to 200 times fewer compared to 35 mm Petri dishes (Ø 35 mm)14,38.
Since extracellular matrix gel is expensive, this method is also more cost-efficient. In addition, the chamber slides can be replaced by plastic cover slips to further reduce the costs. For the preparation of the extracellular matrix gel spots overnight drying of the chamber slides is essential. As the extracellular matrix gel spots are transparent, it is necessary to mark the spots at the bottom side using back lighting. The chambers slides are fixed in a Petri dish for easy manipulation. Further cell culture expansion is not necessary, which offers the possibility to study the SCs of smaller muscles or small muscle samples. Alternatively, e.g. for PCR or muscle constructs if more cells are needed, the freshly isolated SCs can first be expanded in T75 flasks as indicated above.
SCs isolated using this protocol are not suitable for further purification with flow cytometry immediately after isolation. The digestion with pronase causes extensive digestion of the surface antigens14. Horse serum and fetal bovine serum that are used for cell culture must first be properly characterized before isolation, as different lot numbers differentially affected myoblasts proliferation and differentiation.
In recent years, there is a growing interest in the muscles derived from the branchial arches and the head mesoderm (e.g. the extraocular muscles)24. It has been clearly demonstrated that head and limb muscles possess highly different properties. Masseter muscle from old animals seems to retain their regenerative capacity in comparison with limb muscles25,26. SCs from the extraocular muscles possess a robust proliferation and differentiation capacity comparable to SCs from head muscles, and show a larger engraftment potential than limb muscle SCs24.
The fiber type distribution and myosin composition varies among muscle groups and also between species. Muscles originating from the first branchial arch in humans contain both slow and fast fibers (subtypes IIA and IIX), neonatal myosins and myosins typical for developing cardiac muscle. In rodents these muscles contain about 95% fast fibers myosin IIA and IIb)44-46. Studies on avian muscles show that SCs from different muscle fiber types vary in differentiation capacity. SCs from fast fibers only differentiate into fast muscle fibers, while SCs from slow fibers can differentiate into both fiber types47. In addition, the percentage of SCs in fast muscle fibers is lower than in slow muscle fibers48,49. This indicates that the fiber type distribution must be taken into account for studies on muscles in the craniofacial area. Similar to cleft palate muscles, the LVP in rodents contains almost exclusively fast fibers50. For that reason, SCs from the LVP are suitable for pre-clinical studies in the field of cleft palate.
This protocol offers new possibilities to study SCs derived from branchiomeric head muscles or other smaller muscles or smaller muscles samples. This will facilitate the development of new therapies to improve the regeneration of muscles in the maxillofacial area in conditions such as cleft palate but also in other conditions affecting smaller muscles.
Disclosures
The authors have no conflicting interests to disclose.
Acknowledgments
This study was funded by a Mosaic grant (017.009.009) from The Netherlands Organization for Scientific Research (NWO) and a Start grant (S-13-167C) for young investigators from the AO Foundation. Z.Y.R is supported by the National Institutes of Health (grant # AG021566, AG035377, NS090051).
References
- Gritli-Linde A. Molecular control of secondary palate development. Developmental Biology. 2007;301:309–326. doi: 10.1016/j.ydbio.2006.07.042. [DOI] [PubMed] [Google Scholar]
- Marrinan EM, LaBrie RA, Mulliken JB. Velopharyngeal function in nonsyndromic cleft palate: relevance of surgical technique, age at repair, and cleft type. The Cleft Palate-Craniofacial Journal. 1998;35:95–100. doi: 10.1597/1545-1569_1998_035_0095_vfincp_2.3.co_2. [DOI] [PubMed] [Google Scholar]
- Morris HL. Velopharyngeal competence and primary cleft palate surgery, 1960-1971: a critical review. The Cleft Palate Journal. 1973;10:62–71. [PubMed] [Google Scholar]
- Mossey PA, Little J, Munger RG, Dixon MJ, Shaw WC. Cleft lip and palate. Lancet. 2009;374:1773–1785. doi: 10.1016/S0140-6736(09)60695-4. [DOI] [PubMed] [Google Scholar]
- Boorman JG, Sommerlad BC. Musculus uvulae and levator palati: their anatomical and functional relationship in velopharyngeal closure. British Journal of Plastic Surgery. 1985;38:333–338. doi: 10.1016/0007-1226(85)90237-1. [DOI] [PubMed] [Google Scholar]
- Bae YC, Kim JH, Lee J, Hwang SM, Kim SS. Comparative study of the extent of palatal lengthening by different methods. Annals of Plastic Surgery. 2002;48:359–362. doi: 10.1097/00000637-200204000-00004. [DOI] [PubMed] [Google Scholar]
- Braithwaite F, Maurice DG. The importance of the levator palati muscle in cleft palate closure. British Journal of Plastic Surgery. 1968;21:60–62. doi: 10.1016/s0007-1226(68)80087-6. [DOI] [PubMed] [Google Scholar]
- Inman DS, Thomas P, Hodgkinson PD, Reid CA. Oro-nasal fistula development and velopharyngeal insufficiency following primary cleft palate surgery--an audit of 148 children born between 1985 and 1997. British Journal of Plastic Surgery. 2005;58:1051–1054. doi: 10.1016/j.bjps.2005.05.019. [DOI] [PubMed] [Google Scholar]
- Phua YS, de Chalain T. Incidence of oronasal fistulae and velopharyngeal insufficiency after cleft palate repair: an audit of 211 children born between 1990 and 2004. The Cleft Palate-Craniofacial Journal. 1990;45:172–178. doi: 10.1597/06-205.1. [DOI] [PubMed] [Google Scholar]
- Kirschner RE, et al. Cleft-palate repair by modified Furlow double-opposing Z-plasty: the Children's Hospital of Philadelphia experience. Plastic and Reconstructive Surgery. 1999;104:1998–2010. doi: 10.1097/00006534-199912000-00009. [DOI] [PubMed] [Google Scholar]
- Mauro A. Satellite cell of skeletal muscle fibers. The Journal of Biophysical and Biochemical Cytology. 1961;9:493–495. doi: 10.1083/jcb.9.2.493. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yablonka-Reuveni Z. The skeletal muscle satellite cell: still young and fascinating at 50. The Journal of Histochemistry and Cytochemistry. 2011;59:1041–1059. doi: 10.1369/0022155411426780. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ten Broek RW, Grefte S, Von den Hoff JW. Regulatory factors and cell populations involved in skeletal muscle regeneration. Journal of Cellular Physiology. 2010;224:7–16. doi: 10.1002/jcp.22127. [DOI] [PubMed] [Google Scholar]
- Danoviz ME, Yablonka-Reuveni Z. Skeletal muscle satellite cells: background and methods for isolation and analysis in a primary culture system. Methods in Molecular Biology. 2012;798:21–52. doi: 10.1007/978-1-61779-343-1_2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Seale P, et al. Pax7 is required for the specification of myogenic satellite cells. Cell. 2000;102:777–786. doi: 10.1016/s0092-8674(00)00066-0. [DOI] [PubMed] [Google Scholar]
- Yablonka-Reuveni Z, et al. The transition from proliferation to differentiation is delayed in satellite cells from mice lacking MyoD. Developmental Biology. 1999;210:440–455. doi: 10.1006/dbio.1999.9284. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zammit PS, Partridge TA, Yablonka-Reuveni Z. The skeletal muscle satellite cell: the stem cell that came in from the cold. The Journal of Histochemistry And Cytochemistry. 2006;54:1177–1191. doi: 10.1369/jhc.6R6995.2006. [DOI] [PubMed] [Google Scholar]
- Andres V, Walsh K. Myogenin expression, cell cycle withdrawal, and phenotypic differentiation are temporally separable events that precede cell fusion upon myogenesis. The Journal of Cell Biology. 1996;132:657–666. doi: 10.1083/jcb.132.4.657. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fukushima K, et al. The use of an antifibrosis agent to improve muscle recovery after laceration. The American Journal of Sports Medicine. 2001;29:394–402. doi: 10.1177/03635465010290040201. [DOI] [PubMed] [Google Scholar]
- Grefte S, Kuijpers-Jagtman AM, Torensma R, Von den Hoff JW. Skeletal muscle fibrosis: the effect of stromal-derived factor-1α-loaded collagen scaffolds. Regenerative Medicine. 2010;5:737–747. doi: 10.2217/rme.10.69. [DOI] [PubMed] [Google Scholar]
- Jackson WM, Nesti LJ, Tuan RS. Potential therapeutic applications of muscle-derived mesenchymal stem and progenitor cells. Expert Opinion on Biological Therapy. 2010;10:505–517. doi: 10.1517/14712591003610606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sato K, et al. Improvement of muscle healing through enhancement of muscle regeneration and prevention of fibrosis. Muscle, & Nerve. 2003;28:365–372. doi: 10.1002/mus.10436. [DOI] [PubMed] [Google Scholar]
- Tatsumi R, Anderson JE, Nevoret CJ, Halevy O, Allen RE. HGF/SF is present in normal adult skeletal muscle and is capable of activating satellite cells. Developmental Biology. 1998;194:114–128. doi: 10.1006/dbio.1997.8803. [DOI] [PubMed] [Google Scholar]
- Stuelsatz P, et al. Extraocular muscle satellite cells are high performance myo-engines retaining efficient regenerative capacity in dystrophin deficiency. Developmental biology. 2014. [DOI] [PMC free article] [PubMed]
- Ono Y, Boldrin L, Knopp P, Morgan JE, Zammit PS. Muscle satellite cells are a functionally heterogeneous population in both somite-derived and branchiomeric muscles. Developmental Biology. 2010;337:29–41. doi: 10.1016/j.ydbio.2009.10.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pavlath GK, et al. Heterogeneity among muscle precursor cells in adult skeletal muscles with differing regenerative capacities. Developmental Dynamics. 1998;212:495–508. doi: 10.1002/(SICI)1097-0177(199808)212:4<495::AID-AJA3>3.0.CO;2-C. [DOI] [PubMed] [Google Scholar]
- Koo SH, Cunningham MC, Arabshahi B, Gruss JS, Grant JH., 3rd The transforming growth factor-beta 3 knock-out mouse: an animal model for cleft palate. Plastic and Reconstructive Surgery. 2001;108:938–948. doi: 10.1097/00006534-200109150-00018. [DOI] [PubMed] [Google Scholar]
- Fara M, Brousilova M. Experiences with early closure of velum and later closure of hard palate. Plastic and Reconstructive Surgery. 1969;44:134–141. [PubMed] [Google Scholar]
- Lindman R, Paulin G, Stal PS. Morphological characterization of the levator veli palatini muscle in children born with cleft palates. The Cleft Palate-Craniofacial Journal. 2001;38:438–448. doi: 10.1597/1545-1569_2001_038_0438_mcotlv_2.0.co_2. [DOI] [PubMed] [Google Scholar]
- Hanes MC, et al. Contractile properties of single permeabilized muscle fibers from congenital cleft palates and normal palates of Spanish goats. Plastic and Reconstructive Surgery. 2007;119:1685–1694. doi: 10.1097/01.prs.0000258832.84261.37. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rader EP, et al. Contraction-induced injury to single permeabilized muscle fibers from normal and congenitally-clefted goat palates. The Cleft Palate-Craniofacial Journal. 2007;44:216–222. doi: 10.1597/06-036.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rader EP, et al. Effect of cleft palate repair on the susceptibility to contraction-induced injury of single permeabilized muscle fibers from congenitally-clefted goat palates. The Cleft Palate-Craniofacial Journal. 2008;45:113–120. doi: 10.1597/06-171.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Macpherson PC, Dennis RG, Faulkner JA. Sarcomere dynamics and contraction-induced injury to maximally activated single muscle fibres from soleus muscles of rats. The Journal of Physiology. 1997;500(Pt 2):523–533. doi: 10.1113/jphysiol.1997.sp022038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koch KH, Grzonka MA, Koch J. The pathology of the velopharyngeal musculature in cleft palates) Annals of Anatomy. 1999;181:123–126. doi: 10.1016/s0940-9602(99)80117-4. [DOI] [PubMed] [Google Scholar]
- Fara M, Dvorak J. Abnormal anatomy of the muscles of palatopharyngeal closure in cleft palates: anatomical and surgical considerations based on the autopsies of 18 unoperated cleft palates. Plastic and Reconstructive Surgery. 1970;46:488–497. [PubMed] [Google Scholar]
- Carvajal Monroy PL, Grefte S, Kuijpers-Jagtman AM, Wagener FA, Von den Hoff JW. Strategies to Improve Regeneration of the Soft Palate Muscles After Cleft Palate Repair. Tissue Engineering. Part B, Reviews. 2012. [DOI] [PMC free article] [PubMed]
- Grefte S, Kuijpers MA, Kuijpers-Jagtman AM, Torensma R, Von den Hoff JW. Myogenic capacity of muscle progenitor cells from head and limb muscles. European Journal of Oral Sciences. 2012;120:38–45. doi: 10.1111/j.1600-0722.2011.00920.x. [DOI] [PubMed] [Google Scholar]
- Grefte S, Vullinghs S, Kuijpers-Jagtman AM, Torensma R, Von den Hoff JW. Matrigel but not collagen I, maintains the differentiation capacity of muscle derived cells in vitro. Biomedical Materials. 2012;7:055004. doi: 10.1088/1748-6041/7/5/055004. [DOI] [PubMed] [Google Scholar]
- Kastner S, Elias MC, Rivera AJ, Yablonka-Reuveni Z. Gene expression patterns of the fibroblast growth factors and their receptors during myogenesis of rat satellite cells. The Journal of Histochemistry and Cytochemistry. 2000;48:1079–1096. doi: 10.1177/002215540004800805. [DOI] [PubMed] [Google Scholar]
- Yablonka-Reuveni Z, Quinn LS, Nameroff M. Isolation and clonal analysis of satellite cells from chicken pectoralis muscle. Developmental Biology. 1987;119:252–259. doi: 10.1016/0012-1606(87)90226-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sherwood RI, et al. Isolation of adult mouse myogenic progenitors: functional heterogeneity of cells within and engrafting skeletal muscle. Cell. 2004;119:543–554. doi: 10.1016/j.cell.2004.10.021. [DOI] [PubMed] [Google Scholar]
- Gilbert PM, et al. Substrate elasticity regulates skeletal muscle stem cell self-renewal in culture. Science. 2010;329:1078–1081. doi: 10.1126/science.1191035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Motohashi N, Asakura Y, Asakura A. Isolation culture, and transplantation of muscle satellite cells. Journal of Visualized Experiments. 2014. [DOI] [PMC free article] [PubMed]
- Sciote JJ, Horton MJ, Rowlerson AM, Link J. Specialized cranial muscles: how different are they from limb and abdominal muscles. Cells, Tissues, Organs. 2003;174:73–86. doi: 10.1159/000070576. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rowlerson A, Mascarello F, Veggetti A, Carpene E. The fibre-type composition of the first branchial arch muscles in Carnivora and Primates. Journal of Muscle Research and Cell Motility. 1983;4:443–472. doi: 10.1007/BF00711949. [DOI] [PubMed] [Google Scholar]
- Muller J, et al. Comparative evolution of muscular dystrophy in diaphragm, gastrocnemius and masseter muscles from old male mdx mice. Journal of Muscle Research and Cell Motility. 2001;22:133–139. doi: 10.1023/a:1010305801236. [DOI] [PubMed] [Google Scholar]
- Feldman JL, Stockdale FE. Skeletal muscle satellite cell diversity: satellite cells form fibers of different types in cell culture. Developmental Biology. 1991;143:320–334. doi: 10.1016/0012-1606(91)90083-f. [DOI] [PubMed] [Google Scholar]
- Schmalbruch H, Hellhammer U. The number of nuclei in adult rat muscles with special reference to satellite cells. The Anatomical Record. 1977;189:169–175. doi: 10.1002/ar.1091890204. [DOI] [PubMed] [Google Scholar]
- Gibson MC, Schultz E. The distribution of satellite cells and their relationship to specific fiber types in soleus and extensor digitorum longus muscles. The Anatomical Record. 1982;202:329–337. doi: 10.1002/ar.1092020305. [DOI] [PubMed] [Google Scholar]
- Carvajal Monroy PL, et al. A rat model for muscle regeneration in the soft palate. PloS One. 2013;8:e59193. doi: 10.1371/journal.pone.0059193. [DOI] [PMC free article] [PubMed] [Google Scholar]