

Abstract
Neutrophils, the most abundant of all white blood cells in the human circulation, play an important role in the host defense against invading microorganisms. In addition, neutrophils play a central role in the immune surveillance of tumor cells. They have the ability to recognize tumor cells and induce tumor cell death either through a cell contact-dependent mechanism involving hydrogen peroxide or through antibody-dependent cell-mediated cytotoxicity (ADCC). Neutrophils with anti-tumor activity can be isolated from peripheral blood of cancer patients and of tumor-bearing mice. These neutrophils are termed tumor-entrained neutrophils (TEN) to distinguish them from neutrophils of healthy subjects or naïve mice that show no significant tumor cytotoxic activity. Compared with other white blood cells, neutrophils show different buoyancy making it feasible to obtain a > 98% pure neutrophil population when subjected to a density gradient. However, in addition to the normal high-density neutrophil population (HDN), in cancer patients, in tumor-bearing mice, as well as under chronic inflammatory conditions, distinct low-density neutrophil populations (LDN) appear in the circulation. LDN co-purify with the mononuclear fraction and can be separated from mononuclear cells using either positive or negative selection strategies. Once the purity of the isolated neutrophils is determined by flow cytometry, they can be used for in vitro and in vivo functional assays. We describe techniques for monitoring the anti-tumor activity of neutrophils, their ability to migrate and to produce reactive oxygen species, as well as monitoring their phagocytic capacity ex vivo. We further describe techniques to label the neutrophils for in vivo tracking, and to determine their anti-metastatic capacity in vivo. All these techniques are essential for understanding how to obtain and characterize neutrophils with anti-tumor function.
Keywords: Immunology, Issue 100, Neutrophil isolation, tumor-entrained neutrophils, high-density neutrophils, low-density neutrophils, anti-tumor cytotoxicity, BrdU labeling, CFSE labeling, luciferase assay, neutrophil depletion, anti-metastatic activity, lung metastatic seeding assay, neutrophil adoptive transfer.
Introduction
Neutrophils were initially characterized as the innate immune cells which serve as first line defense against invading microorganisms. Today it is known that neutrophils have more far-reaching functions, being involved in mounting adaptive immune responses against foreign antigens1,2, regulating hematopoiesis3, angiogenesis4 and wound healing5. In addition, neutrophils may affect tumor growth and metastatic progression by virtue of their pro- and anti-tumor activities6,7. Neutrophils are characterized by a polymorphic segmented nucleus (hence termed polymorphonuclear (PMN) leukocytes) and contain at least three distinct subclasses of granules as well as secretory vesicles8 (Figure 1A-C).
Neutrophils possess high phagocytic capacity and high NADPH oxidase activity critical for microbial elimination, and secrete a wide range of chemokines important for attraction of additional neutrophils and other immune cells to the site of inflammation8,9. Neutrophils are characterized by the expression of a large amount of surface receptors including Toll-like receptors (TLRs), C-type Lectin Receptors (CLRs), complement receptor 3 (CD11b/CD18) and other adhesion molecules (e.g., L-selectin, LFA-1, VLA-4 and carcinoembryonic antigen-related cell adhesion molecule 3 (CEACAM3/CD66b)), chemokine receptors (e.g., CXCR1, CXCR2, CCR1, CCR2), chemoattractant receptors (e.g., PAFR, LTB4R and C5aR), cytokine receptors (e.g., G-CSFR, IL-1R, IL-4R, IL-12R, IL-18R, TNFR), formyl-peptide receptors (e.g., FPR1-3), and Fc receptors (e.g., CD16 (FcγRIII), CD32 (FcγRII), and CD64 (FcγRI)10. In mice, neutrophils are usually identified as CD11b+Ly6G+, whereas human neutrophils are identified using the CD11b, CD15, CD16 and CD66b leukocyte markers. It is also generally accepted to stain for the granule proteins myeloperoxidase (MPO) and neutrophil elastase (NE) for detection of neutrophils in tissues.
It is still unclear whether the diverse functions of neutrophils are mediated by the same cell or by distinct cell sub-populations. Accumulating data suggest for the presence of a heterogenic neutrophil population that exhibits a high degree of plasticity affected by pro-inflammatory stimuli and the microenvironment11,12. Fridlender et al.13 have grossly divided the neutrophils in cancer into two major sub-populations termed N1 with anti-tumor properties and N2 with pro-tumor properties. In cancer, as well as in chronic inflammation, there is an additional sub-population composed of granulocytic myeloid-derived suppressor cells (G-MDSCs) that suppress T cell responses14. G-MDSCs are considered to be immature myeloid cells characterized by a CD11b+Ly6ClowLy6Ghi phenotype in mice15, while having a CD15+/CD16low phenotype in human16. G-MDSCs express higher levels of arginase and myeloperoxidase, while lower levels of cytokines and chemokines than normal circulating neutrophils. They are less phagocytic and migratory, but produce higher levels of ROS15,17,18. In the present paper we will describe some basic methodologies for isolation and characterization of neutrophils with anti-tumor properties.
While neutrophils constitute the largest population of all white blood cells in the human circulation (45 - 70%; 1,800 - 6,000/μl), in mice, under normal conditions, they are rather sparse (10 - 15%; 300 - 500/μl). The neutrophil count increases steadily upon inflammation and occasionally in cancer, which represents a state of chronic inflammation7. Neutrophils develop from multipotent common myeloid precursor (CMP) cells in the bone marrow, through a differentiation process passing the stages of myeloblasts (MB), promyelocytes (PM), myelocytes (MC), metamyelocytes (MM) and band cells (BC)8. The mature, post-mitotic neutrophils may remain within the bone marrow for 4 - 7 days before they are released to the circulation8. Neutrophil turnover in the blood is usually rapid with an average half-life of 6 - 12 hrs, which may be prolonged under inflammatory conditions. Unstimulated neutrophils have limited anti-tumorigenic activity, a feature that can be acquired by exposing the naïve neutrophils to the chemokines IL-8 (CXCL2), CCL2, CCL5 and CXCL56,19 or artificially, by exposing them to the phorbol ester phorbol 12-myristate 13-acetate (PMA)6.
The short half-life of blood neutrophils together with the low number of neutrophils (~ 3 - 5 x 105) achieved from 1 ml blood of a naïve 6 - 8 week old mouse, have made it difficult to explore the function of circulating mouse neutrophil in vitro. To overcome this difficulty, other sources have been used. For instance, large numbers of neutrophils may be obtained from the bone marrow20 or the peritoneum following the induction of sterile inflammation (e.g., after intraperitoneal injection of thioglycollate broth or Zymosan A). It should be noted that neutrophils obtained from the peritoneal cavity do not exert any anti-tumorigenic activity (unpublished observation).
Granot et al.6 observed that BALB/c mice inoculated orthotopically with the mouse 4T1 breast carcinoma cell line develop neutrophilia which aggravates with tumor progression6 (Figure 2A), such that 20 - 40 million blood neutrophils can be easily isolated from 1 ml blood 3 - 4 weeks post-tumor inoculation. These neutrophils have acquired anti-tumor activities, and have accordingly been coined tumor-entrained neutrophils (TEN), in order to distinguish them from naïve neutrophils6 (Figure 2B). While high-density neutrophils (HDN, Figure 1A) are highly anti-tumorigenic, low-density neutrophils (LDN, Figure 1B) generated in the context of cancer are not21. Also, high-density neutrophils from the bone marrow and spleen of tumor-bearing mice have anti-tumor activity (unpublished data). It should be noted that with tumor progression the spleen becomes gradually enlarged (splenomegaly), with increasing amounts of neutrophils.
It should be noted that TEN are also generated in other models of cancer including both spontaneous (MMTV-PyMT and MMTV-Wnt1 mammary tumors and k-Ras driven lung tumors) and injected (AT-3 (MMTV-PyMT) and E0771 breast carcinoma cells, LLC Lewis lung carcinoma cells and B16-F10 melanoma cells). However, the extent of neutrophil mobilization in these tumor models is far less than of 4T1-inoculated mice, reaching 5 - 10 x 106 neutrophils in 1 ml blood after 3 weeks.
Protocol
Animals: 5-7 weeks old BALB/c mice are purchased from Harlan (Israel). All experiments involving animals were approved by the Hebrew University’s Institutional Animal Care and Use Committee (IACUC). Human samples: Collection of blood from cancer patients and healthy volunteers was approved by Hadassah Medical Center Institutional Review Board (IRB).
1. Induction of Neutrophils with Anti-tumor Properties in vivo Using a Breast Cancer Mouse Model.
NOTE: All steps should be performed using sterile solutions in a laminar airflow (LAF) Bio-Safety cabinet.
Seed 5 x 105 4T1 cells in 100 mm tissue culture plate in 10 ml of Dulbecco's modified Eagle medium (DMEM) containing 4.5 g/L D-glucose supplemented with 10% heat-inactivated fetal bovine serum (FBS), 2 mM D-glutamine, 1 mM sodium pyruvate, 100 U/ml penicillin G and 100 μg/ml streptomycin sulfate. Incubate the cells at 37 °C in a humidified incubator containing 5% CO2 for 3 to 4 days. Ensure that the cells are 50 - 100% confluent on the day of the experiment.
To detach the tumor cells from the tissue culture plate, aspirate the culture medium, wash the cells with 5 ml PBS, aspirate the PBS and add 3 ml of a 2.5 g/L trypsin solution. Incubate the cells 2 - 3 min with trypsin at 37 ºC.
Add 10 ml serum-containing medium to neutralize the trypsin and pipette up and down until all of the cells have been detached. Transfer the cells suspension to a 15 ml conical centrifuge tube.
Centrifuge the cells at 200 x g for 5 min at RT. Aspirate the medium leaving 200 μl medium above the cell pellet. Resuspend the cells in the residual volume and add 10 ml of PBS. Invert the tube 2 - 3 times to get a homogeneous cell suspension and take out 10 μl to count the cells in a hemocytometer. Use trypan blue to distinguish between live and dead cells.
Centrifuge the cell suspension for 5 min at 200 x g at RT. Aspirate the PBS and resuspend the cells in PBS at a final concentration of 2 x 106 cells/ml. Ensure that the single cell suspension has no clumping.
- Inject 1 x 106 4T1 cells or luciferase-expressing 4T1 cells in 50 μl PBS orthotopically into the left inguinal mammary fat pad of female BALB/c mice using a 0.3 ml syringe with a 30G x 8mm needle.
- Before injection, anesthetize the mice in an induction chamber receiving a slow flow rate of isoflurane (3 - 5%) in 100% oxygen.
- Lay the mouse on a sterile surgical pad and make sure the head is properly placed inside the isoflurane nose cone. Confirm proper anesthesia by pinching the paw. Use vet ointment on eyes to prevent dryness while under anesthesia. Shave the hair around the injection site and disinfect with 70% EtOH.
- Use a sterile set of surgical tools to make a small horizontal incision (5 mm) approximately half way between the inguinal and the abdominal nipples, expose the fat pad and inject 1 x 106 4T1 cells in 50 μl. Close the incisions with 9 mm clips that should be removed one week later. Injection of luciferase-expressing cells allows monitoring of tumor size and metastasis by bioluminescence imaging. NOTE: This minimally invasive procedure does not require any post-surgical treatment and the injected mice recover within 2 - 3 min.
- Monitor the mice until they regain consciousness, and make sure they regain full consciousness before joining the company of other mice.
After 3 - 4 weeks, when the primary tumor has reached a volume of 2 cm3, sacrifice the mice by a slow CO2 stream, and immediately after the last breath, obtain blood by cardiac puncture using a 25G x 5/8' needle connected to a 1 ml tuberculin syringe pretreated with heparin. Keep the mouth of the needle upward when inserting the syringe in a horizontal position through the diaphragm towards the heart, and slowly and gradually draw the blood avoiding excess pressure (Figure 3).
2. Neutrophil Isolation
- Isolation of cytotoxic neutrophils from blood of tumor-bearing mice.
- Dilute 1 ml blood drawn from a tumor-bearing mouse (see Protocol 1.11) in PBS containing 0.5% (w/v) bovine serum albumin (BSA) to a final volume of 6 ml.
- Fractionation of diluted blood on a freshly prepared discontinuous sucrose gradient:
- Add 3 ml of sterile-filtered sucrose 1.119 g/ml to the bottom of a 15 ml conical polypropylene centrifuge tube.
- Slowly and carefully, layer 3 ml of sterile-filtered sucrose 1.077 g/ml on top of the 1.119 g/ml layer. Thereafter, add slowly and carefully the 6 ml of diluted blood (Protocol 2.1.1) on top of the 1.077 g/ml layer (Figure 4A). It is recommended to hold the tube tilted and adding the different components by a slow, but continuous flow, keeping the mouth of the pipette towards the lower wall of the tube, such that no turbulence is formed.
- Centrifuge the tube containing the diluted blood on the sucrose gradient at 700 x g for 30 min at RT without brake.
- Carefully remove the tube from the centrifuge without causing any turbulence. Most of the erythrocytes will be at the bottom of the tube. High-density neutrophils (HDN) are found as a white-to-red ring at the interface between the 1.119 g/ml and the 1.077 g/ml layers (around the 3 ml mark), while the low-density leukocytes are found in a white ring at the interface between the 1.077g/ml layer and the BSA-containing PBS (around the 6 ml mark, see Figure 4B).
- Aspirate the PBS + 0.5% BSA until reaching 5 mm above the low-density cell layer. Pipette out the low-density cells by slow suction into a 1 ml tip through slowly swirling around the cells. Transfer the cells into 30 ml of PBS with 0.5% BSA.
- Aspirate the upper layer in the same gradient tube until reaching 5 mm above the high-density cell band. Pipette out the high-density cells, which are mostly high-density neutrophils, and transfer the cells into 30 ml of PBS containing 0.5% BSA.
- Centrifuge the cells at 400 x g for 10 min at RT.
- Aspirate the supernatant and lyse erythrocytes by resuspending the cells in 36 ml sterile HPLC grade water for 30 sec. Isotonicity should be restored by adding 9 ml of a 5x concentrated PBS supplemented with 2.5% (w/v) BSA.
- Centrifuge the cells at 400 x g for 10 min at RT.
- Aspirate the supernatant, and resuspend the cells in PBS-BSA. Count the number of neutrophils in a hemocytometer and use trypan blue to distinguish between live and dead cells.
- Centrifuge the cells at 400 x g for 10 min at RT and resuspend the neutrophils in desired incubation medium to final cell density. Use the neutrophils immediately.
- Then, after another centrifugation, resuspend the neutrophils in incubation medium to final cell density. Use the neutrophils immediately.
- Isolation of circulating neutrophils from cancer patients.
- Mix 10 ml of heparinized (20 U/ml) human blood with an equal volume of 3% Dextran T500 in saline and incubate for 30 min at RT. During this incubation the erythrocytes will sediment.
- Prepare a 50 ml conical polypropylene tube with 10 ml sucrose 1.077 g/ml and slowly layer the leukocyte-rich supernatant on top of the 1.077 g/ml sucrose layer (Figure 4C).
- Centrifuge at 400 x g for 30 min at RT without brake. The high-density neutrophils (HDN) will appear in the pellet. Low-density neutrophils (LDN) co-purify with monocytes and lymphocytes at the interface between the 1.077 g/ml sucrose layer and plasma (Figure 4D).
- Resuspend the neutrophils in 10 ml 0.2% NaCl for 30 sec to lyse the contaminating erythrocytes, and restore isotonicity by adding 10 ml of 1.6% NaCl and invert once.
- Centrifuge 5 min at 160 x g at RT, and wash three times in 20 ml Hanks' balanced salt solution. Centrifuge after each wash and aspirate the supernatant.
- Count the neutrophils and resuspend the cells in RPMI-1640 supplemented with 2% FBS at 2 x 106 neutrophils/ml or as desired.
3. Enrichment of Blood Neutrophils Using Magnetic Beads
- Positive selection
- Take 1 ml of blood from a mouse as described in Protocol 1.8, with a heparinized syringe.
- Centrifuge the blood in a 15 ml conical tube at 400 x g for 5 min at RT.
- Aspirate the supernatant or keep the blood plasma for further studies.
- Lyse the erythrocytes by resuspending the cells in 8 ml HPLC-grade water. After 30 sec restore isotonicity by adding 2 ml of 5x concentrated PBS containing 2.5% BSA. Count the cells.
- Centrifuge at 400 x g for 5 min at RT.
- Resuspend the cell pellet in 200 μl PBS containing 0.5% BSA and 2 mM EDTA per 108 cells.
- Add 50 μl of biotinylated anti-Ly6G antibody. Mix well and incubate for 10 min in the refrigerator (not on ice).
- Add 150 μl cold PBS containing 0.5% BSA and 2 mM EDTA.
- Vortex the anti-biotin coated magnetic microbead stock solution. Transfer 100 μl to the cell suspension. Mix well and incubate for 15 min in the refrigerator (not on ice).
- Wash cells by adding 10 ml PBS containing 0.5% BSA and 2 mM EDTA, and centrifuge at 400 x g for 10 min at RT.
- Aspirate supernatant completely, and resuspend in 500 μl cold PBS containing 0.5% BSA and 2 mM EDTA.
- Insert a magnetic separation column into a magnet holder that is attached to a magnetic stand, and rinse it with 500 μl cold PBS containing 0.5% BSA and 2 mM EDTA.
- Apply the cell suspension onto the column. The flow-through contains unlabeled LyG6-negative cells.
- Wash the column with 500 μl PBS containing 0.5% BSA and 2 mM EDTA. Additional unlabeled cells will be in the flow through.
- Repeat the washing step with another 500 μl PBS containing 0.5% BSA and 2 mM EDTA.
- Remove the column from the magnet and place it on a 15 ml collection tube. Add 1 ml of PBS containing 0.5% BSA and 2 mM EDTA, and flush out the magnetically labeled cells by firmly pushing the plunger into the column. The flow through contains Ly6G+ neutrophils.
- Negative selection
- Prepare blood leukocytes according to steps 3.1.1 to 3.1.5.
- Resuspend 1 x 108 cells in 1 ml PBS containing 0.5% BSA and 2 mM EDTA in a 5 ml polystyrene round-bottom tube.
- Add 50 μl normal rat serum.
- Add 50 μl of neutrophil enrichment cocktail (containing biotinylated antibodies specific for non-neutrophil white blood cells), mix well and incubate for 15 min in the refrigerator.
- Wash the cells by adding 4 ml PBS containing 0.5% BSA and 2 mM EDTA, and centrifuge 400 x g for 10 min.
- Discard the supernatant and resuspend the cells in 1 ml PBS containing 0.5% BSA and 2 mM EDTA.
- Add 50 μl of tetrameric antibody complexes directed against biotin and dextran. Mix well and incubate for 10 min in the refrigerator.
- Vortex well the tube containing dextran-coated magnetic beads before adding 150 μl to the cell suspension. Mix well and incubate 10 min in the refrigerator.
- Bring the cell suspension to a total volume of 2.5 ml by adding PBS containing 0.5% BSA and 2 mM EDTA. Mix gently to get a homogenous cell suspension.
- Insert the tube (without the cap) into the magnet, and let stand for 3 min.
- Invert the magnet with the tube in one continuous motion such that the unbound cells in the fluid will be transferred to a new tube. Leave the magnet and tube inverted for 2 - 3 sec, then return to upright position. The magnetically labeled unwanted cells will remain bound to the wall of the original tube, while the unbound neutrophils will be in the transferred fluid.
4. Cytological Staining of Neutrophils
Resuspend 1x105 neutrophils in 50 µl PBS and transfer the cell suspension to a thin-layer cell preparation adaptor such as a Cytospin. Centrifuge the adaptor at 150 x g for 5 min. Separate the pre-labeled glass slide from the adaptor.
Fix cells by dipping the glass slides in 70% ethanol for 2 min. Allow the preparations from the adaptor (see step 4.1) to dry at RT before staining. Dip the slides 5 - 6 times in distilled water.
Stain 1 - 2 min in Mayer's Hematoxylin solution. Rinse 1 min in tap water. Stain 10 sec in Eosin Y solution. Wash in Tap water. Dehydrate by rinsing the slides in increasing ethanol concentrations (70%, 96% and 100%). Let the glass slides air-dry shortly and inspect the slides under a light microscope. NOTE: Hematoxylin has a deep blue-purple color and stains nucleic acids, whereas eosin is pink and stains proteins nonspecifically. The cytoplasmic granules of neutrophils remain unstained by acidic or basic dyes, which is the origin for the name 'loving to be neutral'. Whereas basophilic granulocytes stain dark blue with hematoxylin and eosin and eosinophilc granulocytes bright red, neutrophils appear neutral pink (See Figure 1A). Mature neutrophils are characterized by a polymorphonuclear nucleus, which is in general large with 2 - 5 lobes 'segmented neutrophils' (Figure 1A and 1C). Immature neutrophils are characterized by a one-lobed curved or ring-shaped nucleus (Figure 1B).
5. Determination of the Purity of Neutrophils by Flow Cytometry.
Resuspend 1x106 cells in 100 μl of FACS buffer (PBS containing 0.5% BSA, 2 mM EDTA and 0.02% NaN3). For whole blood samples, hemolysis is required before staining (steps 3.1.1 to 3.1.5). Add 10 μl of FcR Blocking Reagent for 5 min.
Add 0.5 μg of fluorescent-labeled antibody with a specificity to Ly-6G for mouse neutrophils or to CD11b and CD66b for human neutrophils, mix well and incubate for 15 min at RT.
Adjust volume to 500 μl with PBS containing 0.5% BSA and 2 mM EDTA and analyze staining by flow cytometry.
6. Follow Neutrophil Gate in vivo
- In vivo BrdU labeling of neutrophils
- Inject 100 μl of a 10 mg/ml bromodeoxyuridine (BrdU) solution in sterile PBS intraperitoneally to tumor-bearing mice.
- Isolate blood neutrophils 48 hrs post-injection according to Protocol 2.1. Stain BrdU-labeled neutrophils using a BrdU flow kit.
- CFSE labeling of neutrophils
- Resuspend 107 neutrophils in 1 ml of pre-warmed PBS.
- Add 2 μl of a 5 mM CFSE stock solution to the suspended neutrophils to a final concentration of 10 μM. Mix well and incubate for 15 min at 37 °C. 5 mM CFSE is prepared by dissolving 2.8 mg of CFSE (5-(and 6-)-Carboxyfluorescein diacetate, succinimidyl ester) in 1 ml DMSO. Divide into 10 μl aliquots in sterile 200 μl tubes and store in the dark at -20 °C.
- Neutralize excess CFSE by adding an equal volume of RPMI-1640 containing 10% FBS. Incubate for 10 min at 37 °C. Centrifuge the neutrophils at 400 x g for 10 min at RT. Wash the neutrophils twice in 10 ml of RPMI-1640 containing 10% FBS.
- Centrifuge at 400 x g for 10 min and resuspend in an appropriate volume of PBS. The neutrophils (e.g., 1 x 107 cells) can now be injected intravenously or via cardiac injection to recipient mice.
7. In vitro Luciferase Assay to Monitor the Anti-tumor Activity of Isolated Neutrophils.
Cultivate luciferase-labeled tumor cells as described for 4T1 cells in Protocol 1.1-1.5, but resuspend the trypsin-dissociated cells in optimized reduced serum medium supplemented with 2% FBS. Adjust the cell density to 5 x 104 cells per ml.
Seed 5,000 luciferase-labeled tumor cells in 100 μl optimized reduced serum medium containing 0.5% FBS in each well of a white 96-flat-bottom tissue-culture well plate.
4 hrs after seeding the tumor cells, add 1 x 105 neutrophils in 50 μl optimized reduced serum medium containing 0.5% FBS, and incubate O/N. Prepare a neutrophil cell suspension with a density of 2 x 106 cells/ml. Control wells should get 50 μl medium without neutrophils. Do multiple repeats of each experimental setting.
On the following morning, gently aspirate the supernatant and wash each well with 200 μl PBS. Aspirate the PBS and add 50 μl of passive lysis buffer. For easily detaching cells (such as AT-3 cells), do not wash in PBS and add the cell culture lysis buffer immediately after aspirating the growth medium.
Cover the plate with aluminum foil and incubate it on an orbital shaker at 150 rpm for 20 min at RT.
Place the plate in a luminescence plate reader. Inject well-wise 50 μl luciferase assay solution, and read the chemiluminescence for 10 sec per well.
Calculate the % tumor lysis by the following formula: % tumor lysis = (1-[luminescence of samples with neutrophils]/[luminescence of samples in medium]) x 100%.
NOTE: To assess the contribution of neutrophils to metastatic seeding, neutrophils may be depleted as described in protocol 8.1. For effective depletion administer neutrophil-depleting antibodies starting on day 7 post-tumor engraftment.
8. Anti-metastatic Activity of Neutrophils in a Breast Cancer Mouse Model.
- In vivo depletion of neutrophils.
- Freshly prepare 12.5 μg of rat anti-Ly6G antibody (neutrophil depleting antibody) or of rat isotype control antibody (IgG2a, Κ) in saline at a final volume of 100 μl per mouse.
- Starting on day 3 post-tumor engraftment inject daily an intraperitoneal dose of 12.5 μg rat anti-Ly6G antibody (100 μl). Inject control mice with or 12.5 μg (100 μl) rat isotype control antibody (IgG2a, Κ).
- Starting on day 14, administer the antibodies twice daily, as the neutrophil production rate dramatically increases when the 4T1 tumor grows.
- Every other day, obtain a blood sample (2 - 3 drops) by nicking the lateral tail-vein. Collect the blood into an anti-coagulant containing tube (Heparin, Citrate or EDTA).
- Verify neutrophil depletion using flow cytometry as described in Protocol 5.
- Tumor Neutralization Test (Modified Winn Assay)
- Isolate neutrophils from tumor-bearing mice. Mix 1 x 106 tumor cells and 3 x 106 neutrophils in 50 μl saline (per mouse).
- Inject the cells subcutaneously to the flank of 6 - 8 weeks old naïve BALB/c mice. Shave the flank prior to tumor engraftment to allow accurate measurements of tumor size. Measure tumor size daily starting on day 5 post-engraftment.
- Neutrophil Adoptive Transfer
- Inject 2 x 104 luciferase-expressing tumor cells in 200 μl PBS to the tail vein.
- Neutrophil transfer should be performed 4 hrs following tumor cell injection. Hence, start the purification of HDN from tumor-bearing mice (protocol 2.1) approximately 2 hrs before their planned in vivo transfer. Resuspend neutrophils in PBS at a final concentration of 2.5 x 107 cells/ml.
- 4 hrs after introducing the tumor cells place the mice under a heat lamp for 5 min. Place the mice in a restrainer and inject 5 x 106 HDN (200 μl) via the tail vein. Control mice are injected with vehicle (PBS).
- Monitor the formation of lung metastases at various time points by using a bioluminescence in vivo imaging system or by immunohistochemistry.
- Lung metastatic seeding assay
- Inject 0.5 - 1 x 106 parental 4T1 cells orthotopically into the left inguinal mammary fat pad as described in Protocol 1.
- On day 10, resuspend GFP-expressing 4T1 cells in PBS at a final concentration of 5 x 105 cell/ml. Place the mice under a heat lamp for 5 min.
- Place the mice in a restrainer and inject 1x105 GFP-expressing 4T1 cells (200 μl) intravenously to 4T1 tumor-bearing mice or naïve mice.
- On the following day, euthanize the mice and perfuse the lungs with 20 ml PBS to remove remaining RBCs.
- Excise the lungs for analysis of GFP-positive cells by immunohistochemistry. NOTE: To assess the contribution of neutrophils to metastatic seeding, neutrophils may be depleted as described in protocol 8.1. For effective depletion administer neutrophil-depleting antibodies starting on day 7 post-tumor engraftment.
9. Suppression of T Cell Proliferation by Neutrophils from Tumor-bearing Mice.
Remove the spleen from a euthanized naïve BALB/c mouse and place in 10 ml PBS.
Place the spleen onto a 40 μm cell strainer that is fit on a petri dish filled with RPMI-1640. Using the plunger end of the syringe, mash the spleen through the cells strainer into the petri dish. Rinse the cell strainer with 5 ml RPMI. Discard the strainer.
Transfer the resuspended cells into a 50 ml conical tube and centrifuge 400 x g for 10 min.
Discard the supernatant, and lyse the erythrocytes by suspending the cells into 36 ml of pure water for 20 sec, and adjust to isotonicity by adding 4 ml of PBS concentrated x 10. Alternatively, lyse the erythrocytes by suspending the cells into 5 ml of erythrocyte lysing buffer (ACK), and incubate 5 min at RT. Neutralize the ACK by adding 10 ml of RPMI-1640 medium. Centrifuge at 400 x g for 10 min at RT. Resuspend the cells in 5 ml PBS and count the cells.
Resuspend the splenocytes in PBS to a final density of 4 × 107cells/2 ml in 15 ml tube. Add 2 ml of a 2.5 μM CFSE solution in PBS. Quickly invert the tube and incubate for 10 min at 37 °C with occasional mixing (every 2 min), protected from light.
Quench excess CFSE by adding 4 ml of pre-warmed FBS (100%) and incubate for 1 min at RT. Add 3 ml of PBS and centrifuge at 400 x g for 10 min.
Wash the cells with 30 ml PBS, and centrifuge at 400 x g for 10 min.
Filter the cells through a 40 μm cell strainer and wash again with PBS.
Resuspend the cells in RPMI-1640 medium supplemented with 10% FBS to a final density of 2 x 107 cells/ml.
Seed 2 x 106/well (200 μl) in a 24-well tissue-culture plate.
Stimulate the cells by adding 1 μg of purified Armenian hamster anti-mouse CD3ε antibody in 500 μl RPMI-1640 with 10% FBS.
Add 2 x 106 HDN or LDN in 300 μl RPMI-1640 with 10% FBS to the CFSE-labeled splenocytes, and incubate for 3 days at 37 °C. Wells without neutrophils should get 300 μl medium. Total volume in each well should be 1 ml.
Collect the cells and prepare them for flow cytometry. Resuspend the cells in 100 μl FACS buffer (Protocol 5), and add 10 μl of FcR Blocking Reagent. Incubate for 5 min at RT.
Add 1 μl of APC-conjugated anti-CD8α antibody and incubate for 15 min at RT.
Determine the CFSE fluorescence intensity on CD8+ T cells by flow cytometry (Figure 5). The CFSE intensity is halved upon each cell division. Thus, the number of cell divisions can be determined by the intensity of CFSE staining.
10. Neutrophil Migration Assay
Seed 5x105 4T1 cells in 7 ml optimized reduced serum medium supplemented with 0.5% FBS in a 25 cm2 tissue culture flask and incubate 24 hrs at 37 °C.
Transfer 800 μl of the supernatant to the bottom chamber of a migration plate with a pore size of 5 µm.
Resuspend 2 x 105 neutrophils in 400 μl of optimized reduced serum medium supplemented with 0.5% FBS. Apply the cell suspension to the top chamber and incubate for 2 hrs at 37 °C.
At the end of incubation, remove the top chamber and count the number of neutrophils that have migrated to the bottom chamber.
11. Monitoring Neutrophil Production of Reactive Oxygen Species (ROS).
Prepare 1.1 x 106 neutrophils/ml in Hank's balanced salt solution without phenol red. Plate 180 μl containing 2 x 105 neutrophils in each well of a white 96-flat-bottom well plate.
Place the plate in a luminescence plate reader. Add 20 μl of a 500 μM Luminol solution in PBS to each well to get a final concentration of 50 μM. Read the basal chemiluminescence for 1sec in a time course of 5 min with 10 sec intervals.
Add a stimulant (e.g., PMA at a concentration of 10 nM or 100 nM or fMLP at a concentration of 10 μM). Prepare a 10x concentrated solution of each agent in Hank's balanced salt solution without phenol red and add 22 μl to the respective wells. To control wells add 22 μl vehicle.
Measure the chemiluminescence in the plate reader. Do both a short (every 10 sec for 5 min) and a long (every minute for 1 hr) time course.
Representative Results
In a recent study we identified an anti-metastatic function for neutrophils6. Neutrophils from tumor-bearing mice acquire a cytotoxic phenotype and have the capacity to kill tumor cells6. This is in contrast to neutrophils from naïve mice that have no significant anti-tumor effect6. Several of the techniques described in the Protocol Section have been used for studying anti-tumor neutrophil function in vitro and in vivo6.
Tumor cytotoxic neutrophils can be obtained from tumor-bearing mice6. To achieve this aim, mice were orthotopically injected with 4T1 cells into the left inguinal mammary fat pad (Protocol 1). By day 21 post-tumor inoculation, the primary tumor reached a size of 1 - 2 cm3 (Figure 3 – the tumor is evident in the lower left abdomen). At this time, the mice were euthanized and 1 ml blood was drawn (per mouse) by cardiac puncture (Figure 3). In parallel, blood from a naïve, non-tumor-bearing mouse was drawn. HDN were then purified on a density gradient (Protocol 2.1 and Figure 4) to yield a highly pure (>98%) population of neutrophils. Neutrophil viability was determined by Trypan Blue staining (Protocol 2.1). The neutrophils were then resuspended in optimized reduced serum medium containing 0.5% FBS at 2 x 106 neutrophils/ml.
To test the extent of neutrophil cytotoxicity, we added 105 neutrophils (50 μl from a 2 x 106 neutrophils/ml stock solution) to luciferase expressing 4T1 target cells (5,000 cells/well in 100 μl) in a flat-bottom white 96-well plate (Protocol 7). Following an O/N incubation, the cells were washed in PBS and lysed in passive cell lysis buffer and the luciferase activity in each sample was tested to evaluate the extent of neutrophil cytotoxicity (Figure 2B) (Protocol 7). The luciferase activity correlates with the number of tumor cells surviving and must be compared to the luciferase activity in control wells where tumor cells are cultured alone. To calculate the extent of tumor cell killing we use the following formula: % tumor cell killing = (1 – [luminescence of samples with neutrophils] / [luminescence of control samples]) x 100%. The data is then presented as % of cells killed under different conditions where control cells cultured alone have a killing % = 0. Using these experimental procedures we found that while neutrophils purified from tumor-free mice show no cytotoxicity towards tumor cells (Figure 2B, tumor free), neutrophils purified from tumor-bearing mice show significant cytotoxicity (Figure 2B, tumor bearing).
To test the immune suppressive properties in LDN and HDN we used the T-cell proliferation assay (Protocol 9). We evaluated the number of CD8+ cells in untreated splenocytes and in splenocytes treated with a αCD3 antibody, which were cultured alone and cells that were cultured in the presence of LDN or in the presence of HDN (Figure 5A-D). Note the dramatic increase in CD8+CFSE+ cells following anti-CD3 stimulation (compare upper right panel in A and B) the dramatic inhibitory effect of suppressive LDN (compare upper right panel in B and C) and the lack of inhibitory effect of HDN (panel D). We also evaluated the extent of CFSE retention, as an indication for proliferation. In Figure 5E, the orange curve presents untreated CD8+ cells, the blue curve CD8+ cells, treated with αCD3 antibodies, the red curve CD8+ cells stimulated with αCD3 in the presence of LDN and the green curve represents CD8+ cells stimulated with αCD3 in the presence of HDN. Note the leftward shift in the αCD3 treated cells (blue curve) and the αCD3 treated cells cultured with HDN (green curve), which indicate CD8+ cell proliferation.
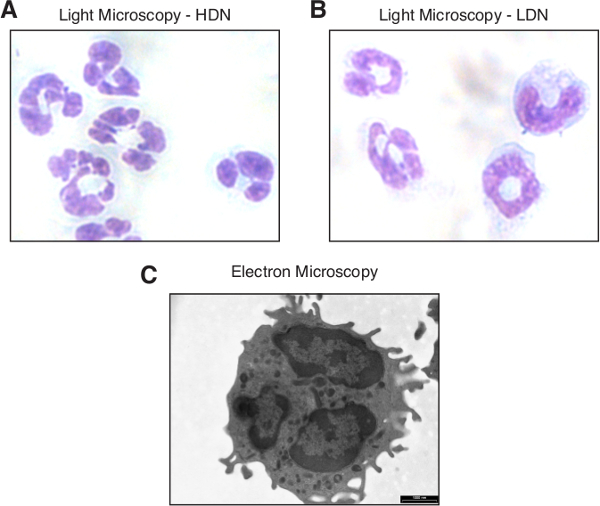 Figure 1. Neutrophil morphology. Light microscopy image of high-density (A) and low-density neutrophils (B) stained with Hematoxylin and Eosin (H&E) following thin-layer cell preparation. (C) A transmission electron microscopy (TEM) image of a high-density neutrophil. The bar represents 1,000 nm. Please click here to view a larger version of this figure.
Figure 1. Neutrophil morphology. Light microscopy image of high-density (A) and low-density neutrophils (B) stained with Hematoxylin and Eosin (H&E) following thin-layer cell preparation. (C) A transmission electron microscopy (TEM) image of a high-density neutrophil. The bar represents 1,000 nm. Please click here to view a larger version of this figure.
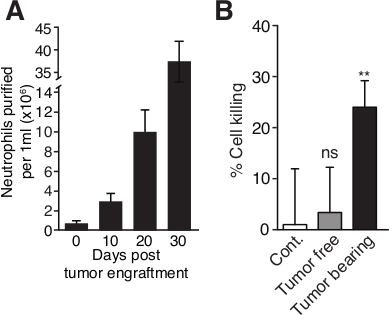 Figure 2. Blood neutrophil counts increase with tumor progression and neutrophils acquire a cytotoxic phenotype. (A) The circulating neutrophil number per 1 ml was counted by FACS at various days following tumor cell inoculation in BALB/c mice. The number ofcirculating CD11b+Ly6G+ neutrophil continuously increases with 4T1 tumor progression. (B) 4T1 breast carcinoma cells were co-cultured with high density neutrophils from either naïve mice (Tumor free) or 4T1 tumor-bearing mice (Tumor bearing) mice, or incubated in medium in the absence of neutrophils (Cont.) at 37 °C for 20 hrs. Neutrophils for tumor bearing mice, but not from tumor free mice, show significant cytotoxicity towards 4T1 tumor cells. Error bars represent ± S.E.M. ** p < 0.01 using a student’s t-test. Please click here to view a larger version of this figure.
Figure 2. Blood neutrophil counts increase with tumor progression and neutrophils acquire a cytotoxic phenotype. (A) The circulating neutrophil number per 1 ml was counted by FACS at various days following tumor cell inoculation in BALB/c mice. The number ofcirculating CD11b+Ly6G+ neutrophil continuously increases with 4T1 tumor progression. (B) 4T1 breast carcinoma cells were co-cultured with high density neutrophils from either naïve mice (Tumor free) or 4T1 tumor-bearing mice (Tumor bearing) mice, or incubated in medium in the absence of neutrophils (Cont.) at 37 °C for 20 hrs. Neutrophils for tumor bearing mice, but not from tumor free mice, show significant cytotoxicity towards 4T1 tumor cells. Error bars represent ± S.E.M. ** p < 0.01 using a student’s t-test. Please click here to view a larger version of this figure.
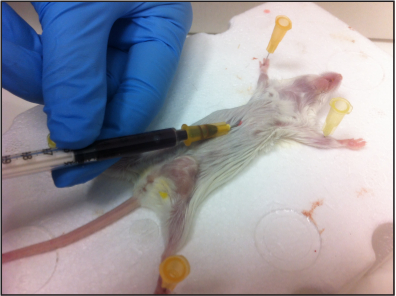 Figure 3. Collection of murine blood via cardiac puncture. The mouse was euthanized in an induction chamber under slow CO2 flow. Immediately after the mouse has taken its terminal breath, it is laid on its back and a 1 ml heparinized syringe is inserted at the base of the sternum until reaching the heart. Slowly pull on the plunger to aspirate the blood. Please click here to view a larger version of this figure.
Figure 3. Collection of murine blood via cardiac puncture. The mouse was euthanized in an induction chamber under slow CO2 flow. Immediately after the mouse has taken its terminal breath, it is laid on its back and a 1 ml heparinized syringe is inserted at the base of the sternum until reaching the heart. Slowly pull on the plunger to aspirate the blood. Please click here to view a larger version of this figure.
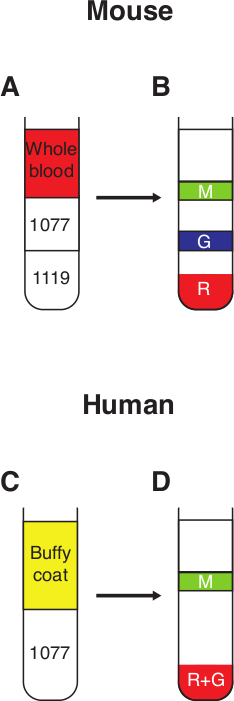 Figure 4. Neutrophil purification from whole blood. (A) 3 ml of 1.077 g/ml sucrose is carefully layered on top of 3 ml 1.119 g/ml sucrose to form a discontinuous gradient. Whole murine blood, diluted in PBS-BSA (0.5%) to a final volume of 6 ml, is then layered on top of the 1.077 g/ml sucrose. (B) Following a 30 min spin at 700 x g with no break, 3 distinct fractions can be observed; R – red blood cells in the pellet, G – the granulocytic fraction containing high-density neutrophils, M – mononuclear fraction containing mononuclear cells and low-density neutrophils. (C) Freshly drawn human blood is mixed with an equal volume of Dextran 500 (3%) and incubated at RT for 30 min. The top fraction containing the white blood cells (buffy coat) is then layered on top of 10 ml 1.077 g/ml sucrose. (D) Following a 30 min spin at 400 x g with no break, 2 distinct fractions may be observed; R+G – pellet containing red blood cells and high-density neutrophils, M – mononuclear fraction containing mononuclear cells and low-density neutrophils. Please click here to view a larger version of this figure.
Figure 4. Neutrophil purification from whole blood. (A) 3 ml of 1.077 g/ml sucrose is carefully layered on top of 3 ml 1.119 g/ml sucrose to form a discontinuous gradient. Whole murine blood, diluted in PBS-BSA (0.5%) to a final volume of 6 ml, is then layered on top of the 1.077 g/ml sucrose. (B) Following a 30 min spin at 700 x g with no break, 3 distinct fractions can be observed; R – red blood cells in the pellet, G – the granulocytic fraction containing high-density neutrophils, M – mononuclear fraction containing mononuclear cells and low-density neutrophils. (C) Freshly drawn human blood is mixed with an equal volume of Dextran 500 (3%) and incubated at RT for 30 min. The top fraction containing the white blood cells (buffy coat) is then layered on top of 10 ml 1.077 g/ml sucrose. (D) Following a 30 min spin at 400 x g with no break, 2 distinct fractions may be observed; R+G – pellet containing red blood cells and high-density neutrophils, M – mononuclear fraction containing mononuclear cells and low-density neutrophils. Please click here to view a larger version of this figure.
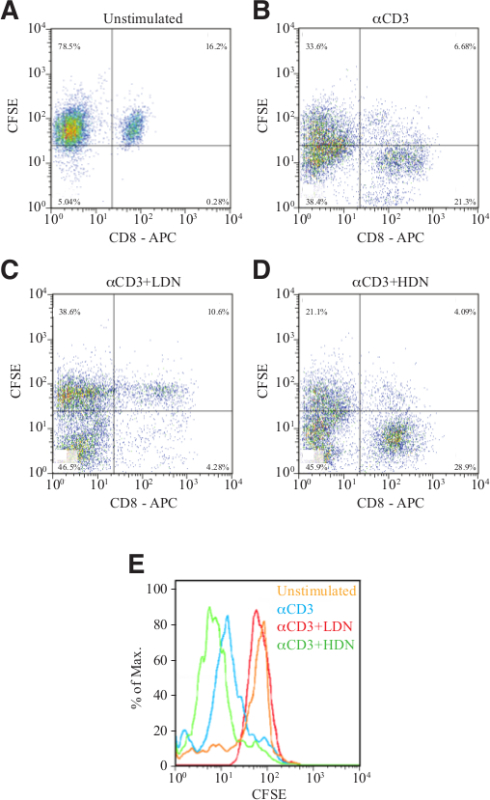 Figure 5. Suppression of T cell proliferation. Flow cytometry analyses showing the number of CFSE-labeled CD8+ cells cultured in the absence of stimulus (A), following stimulation with αCD3 antibody in the absence (B) or presence of LDN (C) or HDN (D). (E) Histogram presentation of CFSE intensity in CD8+ cells from panels A-D. Please click here to view a larger version of this figure.
Figure 5. Suppression of T cell proliferation. Flow cytometry analyses showing the number of CFSE-labeled CD8+ cells cultured in the absence of stimulus (A), following stimulation with αCD3 antibody in the absence (B) or presence of LDN (C) or HDN (D). (E) Histogram presentation of CFSE intensity in CD8+ cells from panels A-D. Please click here to view a larger version of this figure.
Discussion
Neutrophils are the most abundant of all white blood cells and are the first responders in cases of infection and inflammation. As such, they are highly sensitive to external cues and are easily activated. In addition, neutrophils have a very short half-life and a rapid turnover. Together, these characteristics raise several difficulties in working with neutrophils, such that unique experimental strategies are required. For example, there are several neutrophil purification strategies, each with its own pros and cons.
A critical step in working with neutrophils is their purification from whole blood. Neutrophils may be efficiently purified using either density gradients or antibody-based strategies (positive or negative selection). Our method of choice is the use of density gradients since it yields high numbers of highly purified neutrophils with minimal non-specific activation. However, as we show in a recent study21, with tumor progression neutrophils accumulate in high numbers in the mononuclear low-density fraction. Under these conditions the use of a density gradient provides a highly pure high-density neutrophil fraction, that does not represent the entire circulating neutrophil repertoire, and a low-density neutrophil fraction that is heavily contaminated with other mononuclear cells (lymphocytes and monocytes). Under these circumstances the method of choice is an antibody-based purification, preferably negative selection. The use of antibodies to purify neutrophils yields highly pure neutrophils and better represents the entire circulating neutrophil repertoire. However, we noticed that the longer the neutrophils are incubated with the antibodies, the chances for non-specific activation increase. We therefore suggest that for best results, antibody-based neutrophil purification should be performed as quickly as possible. Antibody-based neutrophil purification is also the method of choice when purifying neutrophil from tissues or tumors.
Regardless of the purification procedure chosen, the purity, viability and functional integrity must be rigorously evaluated. The purity of neutrophils can be determined by flow cytometry using antibodies that reacts with neutrophil surface markers. In mouse, Ly-6G is specific for neutrophils, which are characterized by a CD11b+Ly-6ClowLy-6G+F4/80– phenotype. Human neutrophils do not express a marker analogous to Ly-6G and are often characterized by the expression of CD11b, CD15, CD16, and CD66b. Since neutrophils have Fc receptors, these need to be blocked before immunostaining. Neutrophils can also be distinguished from the other white blood cells by having a higher SSC. Viability should be determined at the end of the purification process (trypan blue, protocol 2.1) and should be consistently greater than 98%. Functional integrity should be determined by purification of neutrophils from naïve mice. These neutrophils are not activated and provide non-cytotoxic control for tumor-entrained neutrophils in a co-culture setting with tumor cells (protocol 7).
The short half-life of blood neutrophils together with the low number of neutrophils (~ 3 - 5 x 105) achieved from 1 ml blood of a naïve 6 - 8 week old mouse, have made it difficult to explore mouse blood neutrophil function in vitro. Neutrophil numbers increase steadily in states of inflammation and occasionally in cancer, which represents a state of chronic inflammation7. Some researchers have tried to find alternative sources for neutrophils, such as the bone marrow20. A high number of mouse neutrophils may be obtained within 4 - 24 hr after an intraperitoneal injection of 1 ml of a 3% thioglycollate broth solution or 1 ml of a 1 mg/ml Zymosan A solution in saline. But these elicited neutrophils do not exert any anti-tumorigenic activity (unpublished observation).
Granot et al.6 observed that BALB/c mice inoculated orthotopically with the mouse 4T1 breast carcinoma develop neutrophilia which aggravates upon time (Figure 2A), such that 20 - 40 million blood neutrophils can be easily isolated from 1 ml blood 3 - 4 weeks post-tumor inoculation. These neutrophils have acquired anti-tumor activities, and have accordingly been termed tumor-entrained neutrophils (TEN), in order to distinguish them from naïve neutrophils (Figure 2B). While high-density neutrophils (HDN) are highly anti-tumorigenic, low-density neutrophils (LDN) generated in the context of cancer are not21. High-density neutrophils from bone marrow and spleen of tumor-bearing mice also have anti-tumor activity (unpublished data). It should be noted that with tumor progression the spleen becomes gradually enlarged (splenomegaly), with increasing amounts of neutrophils.
In order to track the fate of neutrophils following their adoptive transfer, these need to be labeled. Neutrophils can be labeled in vivo by injecting bromodeoxyuridine (BrdU) into tumor-bearing or naïve mice 2 days before isolation. BrdU is an analog of the DNA precursor thymidine which is incorporated into newly synthesized DNA in proliferating cells. In the case of neutrophils, BrdU will be incorporated into proliferating precursor cells that retain BrdU staining when differentiating into mature post-mitotic neutrophils. The incorporated BrdU can be stained using specific anti-BrdU fluorescent antibodies. The BrdU+ cells can then be analyzed by flow cytometry. Another approach is to label the isolated neutrophils with a cell tracker dye such as 5-carboxyfluorescein N-succinimidyl ester (CFSE). CFSE is an ester compound that can pass through viable cell membranes. It has an amino-reactive succinimidyl group which leads to the covalent binding of fluorescein to proteins and other amino groups in the cell and the cell surface. The CFSE-labeled cells can be analyzed by flow cytometry using the 488 nm argon laser. The two labeling techniques differ in the fact that BrdU labeling depends on proliferating precursor cells, and not all circulating neutrophils will be labeled, whereas CFSE will stain all neutrophils. Detection of CFSE is more straightforward than BrdU staining, however BrdU labeling is a good means to follow the immature to mature neutrophil conversion.
We have also described several methods to determine the anti-tumor and anti-metastatic function of neutrophils. These include neutrophil depletion, neutrophil adoptive transfer, tumor neutralization test and lung metastasis seeding assay. Each of these assays accomplishes a specific aspect of anti-tumor neutrophil functions. For instance, Granot et al.6 observed that upon depletion of neutrophils, the number of lung metastases in 4T1 tumor-bearing mice is increased, suggesting for an anti-metastatic role of neutrophils. Upon neutrophil adoptive transfer, the tumor cells are injected intravenously 4 hr before injection of purified HDN. The ability of the tumor cells to form lung and liver metastases are followed in a time-course study using an optical in vivo imaging system. Mice receiving HDN showed fewer metastatic foci than did control mice6. In the tumor neutralization test, the tumor cells are injected subcutaneously with or without HDN, the presence of HDN reduces tumor growth21. In the metastatic seeding assay, GFP-labeled tumor cells are injected intravenously into control or neutrophil-depleted pre-metastatic tumor-bearing mice, and the ability of the GFP-labeled cells to seed in the lung is determined. The lungs of pre-metastatic tumor-bearing mice are characterized by high neutrophil infiltration that prevents tumor cell seeding in the specific organ6. This translates to more metastatic foci in neutrophil-depleted mice compared with control tumor-bearing mice.
The protocols described focus on studying neutrophil function in the context of cancer and provide strategies to evaluate cancer-related neutrophil properties both in vitro and in vivo. However, the neutrophil purification strategies as well as some of the experimental procedures described may be used for studying neutrophil function in a wide range of experimental settings where neutrophil play a critical role (i.e., inflammation and infection).
Disclosures
The authors have nothing to disclose.
Acknowledgments
ZG is supported by grants from the I-CORE Program of The Israel Science Foundation (Grant No. 41/11), the Abisch-Frenkel Foundation, the Rosetrees Trust, the Israel Cancer Research Foundation (ICRF - Research Career Development Award) and the CONCERN foundation. ZGF is supported by grants from the Israel Cancer Research Foundation (ICRF - Research Career Development Award), Chief Scientist of the Israel Ministry of Health and the Israel Lung Association.
References
- Tangye SG, Brink R. A helping hand from neutrophils in T cell-independent antibody responses. Nat Immunol. 2012;13:111–113. doi: 10.1038/ni.2214. [DOI] [PubMed] [Google Scholar]
- Mantovani A, Cassatella MA, Costantini C, Jaillon S. Neutrophils in the activation and regulation of innate and adaptive immunity. Nat Rev Immunol. 2011;11:519–531. doi: 10.1038/nri3024. [DOI] [PubMed] [Google Scholar]
- Pruijt JF, et al. Neutrophils are indispensable for hematopoietic stem cell mobilization induced by interleukin-8 in mice. Proc Natl Acad Sci U S A. 2002;99:6228–6233. doi: 10.1073/pnas.092112999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tecchio C, Cassatella MA. Neutrophil-derived cytokines involved in physiological and pathological angiogenesis. Chem Immunol Allergy. 2014;99:123–137. doi: 10.1159/000353358. [DOI] [PubMed] [Google Scholar]
- Liu M, et al. Formylpeptide receptors mediate rapid neutrophil mobilization to accelerate wound healing. PloS one. 2014;9:e90613. doi: 10.1371/journal.pone.0090613. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Granot Z, et al. Tumor entrained neutrophils inhibit seeding in the premetastatic lung. Cancer Cell. 2011;20:300–314. doi: 10.1016/j.ccr.2011.08.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sionov RV, Fridlender ZG, Granot Z. The Multifaceted Roles Neutrophils Play in the Tumor Microenvironment. Cancer Microenviron. 2014. [DOI] [PMC free article] [PubMed]
- Borregaard N. Neutrophils, from marrow to microbes. Immunity. 2010;33:657–670. doi: 10.1016/j.immuni.2010.11.011. [DOI] [PubMed] [Google Scholar]
- Yamashiro S, et al. Phenotypic and functional change of cytokine-activated neutrophils: inflammatory neutrophils are heterogeneous and enhance adaptive immune responses. J Leukoc Biol. 2001;69:698–704. [PubMed] [Google Scholar]
- Futosi K, Fodor S, Mocsai A. Neutrophil cell surface receptors and their intracellular signal transduction pathways. Int Immunopharmacol. 2013;17:638–650. doi: 10.1016/j.intimp.2013.06.034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scapini P, Cassatella MA. Social networking of human neutrophils within the immune system. Blood. 2014;124:710–719. doi: 10.1182/blood-2014-03-453217. [DOI] [PubMed] [Google Scholar]
- Fridlender ZG, et al. Transcriptomic analysis comparing tumor-associated neutrophils with granulocytic myeloid-derived suppressor cells and normal neutrophils. PloS one. 2012;7:e31524. doi: 10.1371/journal.pone.0031524. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fridlender ZG, et al. Polarization of tumor-associated neutrophil phenotype by TGF-beta: 'N1' versus 'N2. TAN. Cancer Cell. 2009;16:183–194. doi: 10.1016/j.ccr.2009.06.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Talmadge JE, Gabrilovich DI. History of myeloid-derived suppressor cells. Nat Rev Cancer. 2013;13:739–752. doi: 10.1038/nrc3581. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Youn JI, Nagaraj S, Collazo M, Gabrilovich DI. Subsets of myeloid-derived suppressor cells in tumor-bearing mice. J Immunol. 2008;181:5791–5802. doi: 10.4049/jimmunol.181.8.5791. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Choi J, et al. CD15+/CD16low human granulocytes from terminal cancer patients: granulocytic myeloid-derived suppressor cells that have suppressive function. Tumour Biol. 2012;33:121–129. doi: 10.1007/s13277-011-0254-6. [DOI] [PubMed] [Google Scholar]
- Brandau S, et al. Myeloid-derived suppressor cells in the peripheral blood of cancer patients contain a subset of immature neutrophils with impaired migratory properties. J Leukoc Biol. 2011;89:311–317. doi: 10.1189/jlb.0310162. [DOI] [PubMed] [Google Scholar]
- Pillay J, Tak T, Kamp VM, Koenderman L. Immune suppression by neutrophils and granulocytic myeloid-derived suppressor cells: similarities and differences. Cell Mol Life Sci. 2013;70:3813–3827. doi: 10.1007/s00018-013-1286-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lopez-Lago MA, et al. Neutrophil chemokines secreted by tumor cells mount a lung antimetastatic response during renal cell carcinoma progression. Oncogene. 2013;32:1752–1760. doi: 10.1038/onc.2012.201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boxio R, Bossenmeyer-Pourie C, Steinckwich N, Dournon C, Nusse O. Mouse bone marrow contains large numbers of functionally competent neutrophils. J Leukoc Biol. 2004;75:604–611. doi: 10.1189/jlb.0703340. [DOI] [PubMed] [Google Scholar]
- Sagiv J, et al. Phenotypic Diversity and Plasticity in Circulating Neutrophil Subpopulations in Cancer. Cell Reports. 2015. [DOI] [PubMed]