

Abstract
Insect nephrocytes are highly endocytic scavenger cells that represent the only invertebrate model for the study of human kidney podocytes. Despite their importance, nephrocyte development is largely uncharacterised. This work tested whether the insect ortholog of mammalian Kidney Krüppel-Like Factor (Klf15), a transcription factor required for mammalian podocyte differentiation, was required for insect nephrocyte development. It was found that expression of Drosophila Klf15 (dKlf15, previously known as Bteb2) was restricted to the only two nephrocyte populations in Drosophila, the garland cells and pericardial nephrocytes. Loss of dKlf15 function led to attrition of both nephrocyte populations and sensitised larvae to the xenotoxin silver nitrate. Although pericardial nephrocytes in dKlf15 loss of function mutants were specified during embryogenesis, they failed to express the slit diaphragm gene sticks and stones and did not form slit diaphragms. Conditional silencing of dKlf15 in adults led to reduced surface expression of the endocytic receptor Amnionless and loss of in vivo scavenger function. Over-expression of dKlf15 increased nephrocyte numbers and rescued age-dependent decline in nephrocyte function. The data place dKlf15 upstream of sns and Amnionless in a nephrocyte-restricted differentiation pathway and suggest dKlf15 expression is both necessary and sufficient to sustain nephrocyte differentiation. These findings explain the physiological relevance of dKlf15 in Drosophila and imply that the role of KLF15 in human podocytes is evolutionarily conserved.
Introduction
Invertebrates, like mammals, have both phagocytic and non-phagocytic scavenger systems for the clearance from circulation of effete cells, foreign material and macromolecules [1–3]. In Drosophila the non-phagocytic system is comprised primarily of nephrocytes, highly endocytic cells that exists close to the heart (pericardial nephrocytes) and oesophagus (garland cells). Nephrocytes have long been regarded as analogous to mammalian reticuloendothelial cells [4], and recent studies indicate they express genes conserved in human renal podocytes (e.g. sticks and stones, an ortholog of mammalian Nphs1, which is critical for slit diaphragm formation) [5, 6] and renal proximal tubule cells (Amnionless and Cubulin, genes critical for protein reabsorption and endocytosis in invertebrates and mammals)[7, 8]. As such, nephrocytes represent an ancestral blood filtration system whose molecular genetics has been conserved, yet repositioned in different mammalian cell types. The molecular genetics of nephrocyte development and differentiation has not been studied in detail and no transcriptional networks have been identified that control the post-embryonic differentiation of nephrocytes. Identifying such networks in Drosophila is important because they may provide important insights into genetic pathways relevant to human physiology and stem cell differentiation.
The Krüppel-like family (KLF) of transcription factors mediate the development, differentiation and function of multiple cell types from invertebrates to mammals [9]. KLF genes encode for proteins with a highly-conserved C-terminal triple tandem repeat zinc finger DNA binding domain and a highly variable N-terminal region. The human ‘kidney KLF’ (KLF15) has a rodent ortholog that regulates cardiac and renal fibrosis, circadian nitrogen metabolism and cardiac potassium channel expression, cardiac hypertrophy, adipogenesis and renal podocyte differentiation [10–15]. An ortholog of KLF15 exists on the X-chromosome in Drosophila but this gene was originally regarded as an ortholog of mammalian Bteb2/Klf5, hence it was named Bteb2. However, this gene shares greater amino acid sequence similarity with the more recently identified mammalian Klf15 [16] (see S1 Fig), and we refer to the Drosophila gene as dKlf15 to reflect this. Embryonic and adult gene expression data indicate that dKlf15 has a highly restricted expression pattern limited to the embryonic garland cells [17] and the adult heart [18], consistent with a role for the gene in nephrocyte development or function (see S2 Fig for expression data).
In this work we use loss and gain of function studies to test the hypothesis that dKlf15 may play a cell specific role in nephrocyte differentiation. Loss of function studies identified dKlf15 as critical for the development of Drosophila’s only two populations of nephrocytes, the garland cells and the pericardial nephrocytes (see anatomical schematics in Fig 1A). The expression of the slit diaphragm gene sticks and stones as well as the endocytic receptor Amnionless were both dependent upon dKlf15 expression. Focussing attention on pericardial nephrocytes revealed that these cells were specified normally during embryogenesis but then died during larval development. Using a conditional silencing strategy it was established that adult nephrocyte function depended upon sustained dKlf15 gene expression. Over-expression of dKlf15 increased nephrocyte numbers and prevented their age-dependent dysfunction. These findings define the physiological relevance of dKlf15 in Drosophila and begin to annotate a genetic pathway driving and sustaining nephrocyte differentiation. The findings also suggest that the role of Klf15 in mammalian kidney podocytes is evolutionarily conserved and that Drosophila may be used to identify novel pathways relevant to the regulation and role of KLF15 in humans.
Fig 1. dKlf15 expression is restricted to nephrocytes.

(A’ & A”) Schematics showing location of the two nephrocyte populations in larval and adult Drosophila, the garland cells (GCs) and pericardial nephrocytes (PNs). Garland cells are at the interface between the paraventriculus (PV) and oesophagus (oe), whereas pericardial nephrocyte are either side of the heart tube (HT); A>P, anterior-posterior. (B’ & B”) Antisera raised to dKlf15 (but not the pre-immune sera, B”), locate to the nucleus of adult pericardial nephrocytes (arrow). (C) Two magnifications of the adult heart immunostained with anti-dKlf15 (1:10) and anti-Amnionless (1:100) antibodies. Arrows denote pericardial nephrocytes, the arrowhead indicates a cardiomyocyte nucleus. Scale bar = 20 μm. (D) The heart in a dissected L3 larva expressing the RedStinger fluorescent reporter driven by dKlf15-Gal4 and stained with wheat germ agglutinin which is taken up preferentially by pericardial nephrocytes (blue); HT = heart tube; scale bar = 40 μm. (E) L2 larva stained with anti-Amnionless and anti-dKlf15 showing localisation of dKlf15 to the nucleus of Amnionless-positive pericardial cells (arrow). (F’-F”“) dKlf15-Gal4 driven RedStinger expression in living larvae and a dissected adult. Fluorescence is seen in binucleate garland cells next to the paraventriculus (PV) in L3 larvae (arrows). Fluorescence was also detected in pericardial nephrocytes (asterisks) either side of the heart tube (HT) at L2, L3 and adult (Ad) stages. Scale bars = 50 μm.
Results
dKlf15 is a nephrocyte-restricted transcription factor
High throughput expression data indicated that dKlf15 expression is limited to embryonic garland cells as well as the adult heart [17, 18]. To verify this, antisera was raised to a peptide corresponding to a region in the N-terminal of dKlf15 protein and used to stain the adult heart. Antibodies to dKlf15 (but not pre-immune sera) localised to the nucleus of pericardial nephrocytes that co-expressed the nephrocyte marker Amnionless in adults (Fig 1B and 1C) and L2 larvae (Fig 1E). The antibody did not stain the cardiomyocytes (arrowhead Fig 1C”); nor the fat body, ovary or oenocytes (see S3 Fig). A fluorescent reporter protein (RedStinger; dsRed tagged with a nuclear-localisation signal) driven by a 2131bp region of the putative dKlf15 enhancer was examined in larval and adult flies. Expression of the reporter was limited to pericardial cells in larvae (seen in dissected preparations as large cells either side of the heart tube that preferentially sequester wheat-germ agglutinin (Fig 1D). No expression of the reporter was seen in L1 pericardial nephrocytes, however expression of the reporter was seen in pericardial cells at the L2 stage (see S4 Fig), concomitant with the differentiation of pericardial cells after the L1 stage (see below) and immune-detection of dKlf15 in Amnionless positive pericardial cells (Fig 1E). The reporter was also expressed at L3 and adult stages, as well as in garland cells of L3 larvae (Fig 1F). Expression was restricted to the pericardial nephrocytes at abdominal sections A5-A8. No other cells were seen to express the reporter in adult or larval stages. These findings corroborate the high throughput data and are consistent with dKlf15 expression being restricted to garland cells and pericardial nephrocytes, the only populations of nephrocytes in Drosophila.
dKlf15 is critical for nephrocyte differentiation
To establish if dKlf15 was relevant to nephrocyte biology a line of flies was sourced which had a large insertion within the dKlf15 locus (Fig 2A). These flies were homozygous viable despite having no detectable dKlf15 expression (Fig 2B). Analysis of all dKlf15 NN mutants was performed on offspring from these homozygous mutant parents. Upon dissection of larvae and adults it was evident that binucleate garland cells and pericardial nephrocytes were missing (Fig 2C & 2D). This allele was therefore referred to as dKlf15 NN, to denote ‘No Nephrocytes’. Silencing dKlf15 with dorothy-Gal4 [19] or Hand-Gal4 [20], led to the absence of nephrocytes in adults (Fig 2E). Female dKlf15 +/NN heterozygous flies had a normal number of nephrocytes (Fig 2F), however females transheterozygous for the dKlf15 NN mutation and a chromosomal deficiency spanning the dKlf15 locus (Df(1)ED6727; i.e. they had no wild type copy of dKlf15) had no nephrocytes (Fig 2F).
Fig 2. dKlf15 is critical for nephrocyte development.
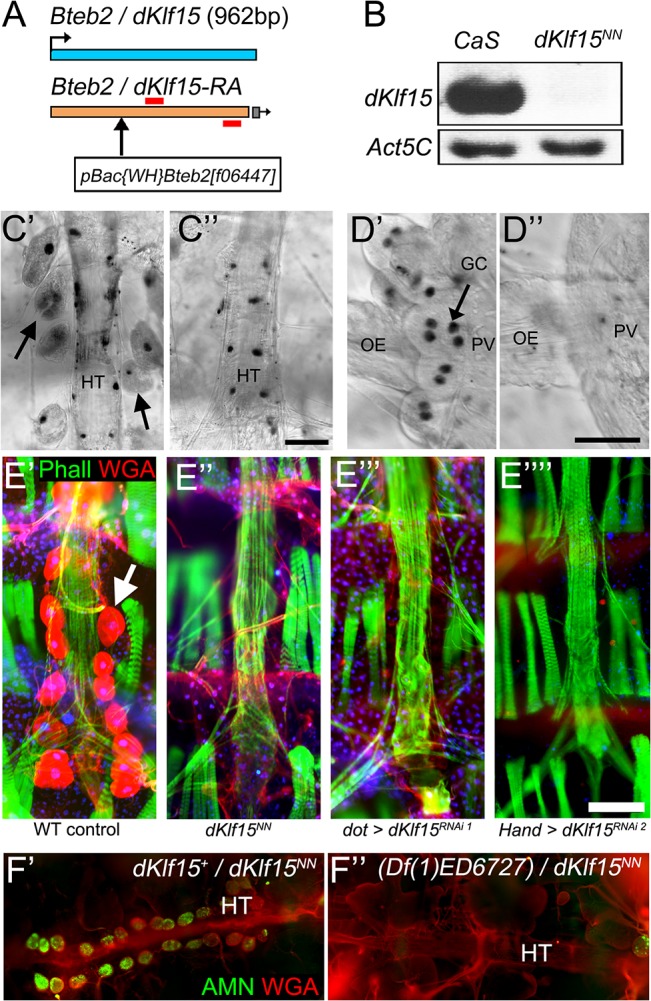
(A) Schematic of the dKlf15 gene. An allele of dKlf15, originally called Bteb2 f06447, and now renamed dKlf15 NN, contains a piggyBac insertion (white box) within the coding region. The red bars define the primer sequence used to detect dKlf15 expression. (B) dKlf15 expression in adult heart from control (CaS) and mutant (dKlf15 NN) flies. (C) Phase contrast micrographs of the heart from wild type (C’) and dKlf15 NN mutant females (C”) overlaid with fluorescence images of a Hoechst-stained adult heart; nuclei are re-coloured black against a white background. Arrows indicate pericardial nephrocytes, HT = heart tube; scale bar = 50 μm. (D) Micrographs of Hoechst-stained larval oesophagus (OE) at the point it meets the proventriculus (PV) in wild type and dKlf15 NN mutants (D’ % D”, respectively); GC = garland cells (arrow); scale bar = 25 μm. (E) Adult heart from wild type (E’), dKlf15 NN mutants (E”) or flies where dKlf15 had been silenced using either dot-Gal4 or Hand-Gal4 (E”‘ & E”“, respectively), stained with wheat-germ agglutinin (red) and phalloidinFITC (green); arrow indicates pericardial nephrocytes; scale bar = 100 μm. (F) Adult heart stained with antibodies to Amnionless (green) and phalloidin (red); hearts were from female flies heterozygous for the mutant dKlf15 NN allele and a wild type copy of the allele (F’) or heterozygous for the dKlf15 NN mutant allele and a deletion spanning the dKlf15 locus (F”; (Df(1)ED6727)); HT = heart tube.
Adult male flies hemizygous for the dKlf15 NN mutation (dKlf15 is on the X-chromosome) had no nephrocytes (Fig 3A & 3B) and this was rescued by a genomic duplication of the wild type dKlf15 locus but not duplications spanning regions either side of the dKlf15 locus (Fig 3A & 3B). These findings established that dKlf15 is critical for nephrocyte development in Drosophila.
Fig 3. Rescue of the dKlf15 NN mutation.
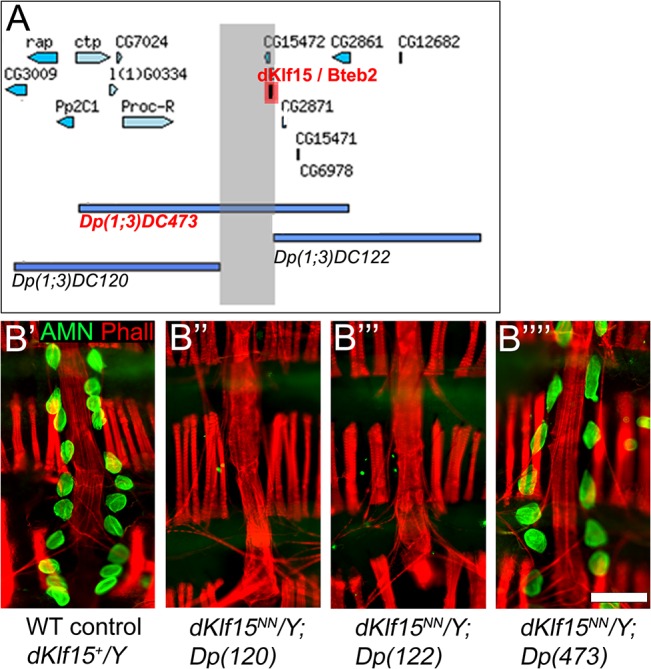
(A) Schematic showing genomic duplications. (B) Adult hearts stained with anti-Amnionless antibodies (green) and phalloidin (red). Male flies were hemizygous for the mutant dKlf15 allele (B’); or hemizygous for the mutant allele and a genomic duplication on the third chromosome corresponding to either side of the dKlf15 locus (Dp(120) and Dp(122); B” and B”‘, respectively) or covering the dKlf15 locus (Dp(473); B”“); scale bar = 100 μm.
Sensitivity to silver nitrate and lifespan of dKlf15 NN mutants
It is known that nephrocytes scavenge ingested silver nitrate and that disruption of nephrocyte function increases mortality of larvae fed silver nitrate [5, 21]. It was therefore hypothesised that loss of dKlf15 expression may reduce the survivorship of larvae fed silver nitrate. When larvae were fed 0.01% silver nitrate, there was increased mortality of dKlf15 NN mutants relative to wild-type controls (P<0.01; Fig 4A). There was no difference between genotypes when provided with a normal diet and larvae of both genotypes died when provided with a higher dose (0.05%) of silver nitrate (Fig 4A). Strikingly, the lifespan of adult control (w 1118) and dKlf15 NN mutant flies was identical (Fig 4B), even when provided with silver nitrate, despite this xenotoxin shortening the lifespan of both genotypes. These findings indicate that dKlf15 mitigates the toxicity of silver nitrate in larvae but not adult flies and that nephrocytes are dispensable for normal survival under control conditions.
Fig 4. Survival of dKlf15 mutants.
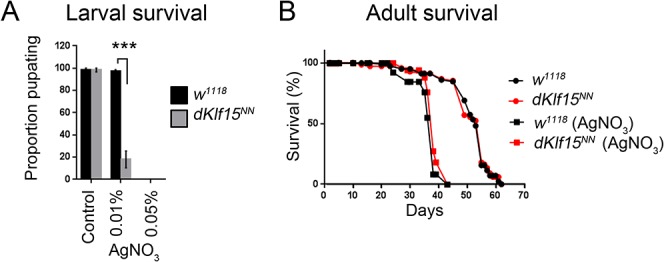
(A) Survival of larvae on yeast containing different amounts of silver nitrate (AgNO3); ***P<0.001; n = 3 independent trials starting with 20 eggs per trial. (B) Survival of adult males on control (circles) or AgNO3 diet (squares); n = 85–120 flies per genotype (housed as 15 flies per vial). Genotype had no effect on lifespan or survival on diet containing silver nitrate (P>0.05). Silver nitrate was toxic to both genotypes (P<0.001, relative to flies on the control diet).
dKlf15 is required for the differentiation of post-embryonic pericardial nephrocytes
To establish when nephrocyte development was affected by the loss of dKlf15, the embryonic heart was stained with antibodies to Tinman, Even-skipped and Odd-skipped, three proteins expressed by cells of the cardiac lineage destined to become pericardial nephrocytes [22, 23]. The number and anatomical location of pericardial nephrocytes in dKlf15 NN mutants at embryonic stage 16 were normal (Fig 5A and 5C). This suggested that the heart was specified normally in mutants, that there is normally no maternal dKlf15 contribution and that loss of dKlf15 function only becomes manifest at a post-embryonic stage of pericardial nephrocyte development. For ease of identification during larval development, control (w 1118) and dKlf15 NN mutants were crossed into a Hand-GFP background to mark cardiomyocytes and pericardial nephrocytes with green fluorescent protein [24]. At the L1 stage the phenotype of the heart in dKlf15 NN mutants was indistinguishable from that in controls. After L1, the pericardial nephrocytes in controls grew considerably in size, whereas this failed to occur in dKlf15 NN mutants and by the L3 stage, very few pericardial nephrocytes remained in the mutant (Fig 5B and 5D). dKlf15 protein was detected in wild type L3 pericardial nephrocytes but not dKlf15 NN mutants (S5 Fig), confirming that the mutant phenotype was due to loss of dKlf15 function.
Fig 5. dKlf15 regulates the post-embryonic differentiation of pericardial nephrocytes.

(A) Stage 16 embryos stained with antibodies to Odd-skipped (Odd) or Even-skipped (Eve) and Tinman (Tin) and β3 tubulin. (B) Wild type (dKlf15 +) and dKlf15 NN on a Hand-GFP background to mark cardiomyocytes and pericardial nephrocytes (arrows). (C) Number of Even-skipped, Odd-skipped and Tinman positive cells in stage 16 embryos; n = 8–12 embryos per genotype. (D) Number and size of nephrocytes in larvae at different stages. ***P<0.001; n = 8–14 larvae per genotype; (note, nephrocyte were too infrequent to quantify at L3 stage). (E) Ultrastructure of nephrocytes from L3 stage wild type (dKlf15 +/+) and mutant dKlf15 NN larvae. Arrows indicate slit diaphragms. Scale bars = 5 μm (upper panels); = 1 μm (lower panels). (F) Hand-GFP wild type and dKlf15 NN lines expressing a sticks and stones reporter (red). Nephrocytes in L1 (arrows); nephrocytes in L3 (asterisks); grey line defines the heart. (G) Nuclear morphology of Hand-GFP-positive cells co-stained with wheat germ agglutinin (red), scale bar = 25 μm. (H) Micrographs show the Hand-GFP fluorescence signal of the ‘wing hearts’ (arrows) seen through the cuticle of the scutellum of pupa.
The filtration slit is a key functional structure in nephrocytes and disruption of the genes which form it leads to nephrocyte death [5]. Transmission electron microscopy was therefore used to examine the ultrastructure of control and dKlf15 NN mutant nephrocytes. Whereas filtration slits could be readily identified at the surface of pericardial nephrocytes in controls, no such structures were seen in dKlf15 NN mutants (Fig 5E). Although pericardial nephrocytes were scarce in L3 larvae, those remaining often had a distinctive ‘poly-cellular’ phenotype. The absence of filtration slits prompted an examination of sticks and stones expression, a gene required for slit formation [5, 6]. dKlf15 NN mutant and control Hand-GFP lines were therefore crossed with flies expressing fluorescently tagged sticks and stones (sns). In controls there was no sns signal in L1 pericardial cells but strong expression in L2s (S6 Fig) and L3s (Fig 5F). This signal was only seen in the pericardial cells around regions A5-A8, the PNs destined to become the adult pericardial nephrocytes. In stark contrast, no sns expression was ever seen in any pericardial cells in the dKlf15 NN mutants, however sns expression was still seen in non-heart cells (S6 Fig), indicating that dKlf15 NN mutants specifically lacked sns expression in the pericardial nephrocytes. The ‘poly-cellular’ phenotype of residual pericardial nephrocytes was also evident in these experiments (asterisks in Fig 5F). Some Hand-GFP positive cells in the mutants at the L3 stage exhibited morphology consistent with cell death (compare upper and lower panels in Fig 5G), however by this stage there were very few nephrocytes remaining. In contrast, the pericardial cells located along the aorta and which form the scutellar pulsatile organs (‘wing hearts’ [25]), appeared unaffected by the dKlf15 NN mutation (Fig 5H). These findings suggest that dKlf15 regulates the post-embryonic maturation of a subset of pericardial cells destined to become the adult pericardial nephrocytes and that loss of dKlf15 function leads to the absence of filtration slits, a situation leading to loss of nephrocyte viability. Hence, the lack of pericardial nephrocytes in adult dKlf15 NN mutants is due to nephrocyte attrition during late larval development.
Adult nephrocytes’ endocytic scavenger function relies on sustained dKlf15 expression
To establish if dKlf15 expression mediated nephrocyte function in adult flies, dKlf15 was silenced in adults using the TARGET system driven by Hand-Gal4. This system, which exploits a temperature sensitive repressor of Gal4, allows for the temporal and tissue dependent switch-on and off of genes using the bipartite Gal4-UAS system [26]. Flies were raised at the non-permissive temperature (18°C, no RNAi expression) and then either maintained as adults at this temperature or placed at the permissive temperature (29°C) to allow RNAi expression. When dKlf15 was silenced in adults, immuno-detectable Amnionless was significantly reduced on the surface of the nephrocytes (Fig 6A–6C). Amnionless is known to mediate nephrocyte endocytosis [7] and it was predicted that this would impact on nephrocyte function. Accordingly it was found that the nephrocytes’ endocytic function as tested in dissected preparations of the adult heart (Fig 6D and 6E and also S7 Fig) and ability to scavenge silver nitrate in vivo (Fig 6F–6H) were significantly reduced by dKlf15 knock-down. Hence, the differentiated state and functionality of adult nephrocytes is dependent upon sustained dKlf15 expression.
Fig 6. Adult pericardial nephrocyte function is dependent on dKlf15.

(A & B) dKlf15 was conditionally silenced in adult nephrocytes using Hand-Target (Target) to drive UAS-dKlf15 RNAi. Flies were reared at 18°C until eclosion then either maintained at 18°C (A’, B’; no knock-down) or placed at 29°C (A”, B”; RNAi permitted) for 4 days. Adult hearts were then stained with wheat germ agglutinin (red) and antibodies to the nephrocyte marker Amnionless (green). Scale bars = 100 μm. (C) Quantification of Amnionless fluorescence signal. ***P<0.001; n = 8 hearts per genotype. (D) Pericardial nephrocytes in semi-intact heart preparations after incubation with fluorescently tagged 10 kDa dextran (green) for 5 or 30 minutes. Nephrocytes of all genotypes associated with dextran when adults were kept at the non-permissive temperature of 18°C (D’), whereas at 29°C, the permissive temperature for silencing, the dKlf15 RNAi line could no longer associate with dextran (D”). (E) Quantification of fluorescence after incubation for 0, 5 or 30 minutes with fluorescently-tagged. *P<0.05, ***P<0.001compared to control (Target genotype), ns = not significantly different from control; n = 12–16 nephrocytes from 4 individual flies per genotype, per time-point. (F) Control (Target flies outcrossed to w 1118, (F’)) and Target > dKlf15 RNAi (F”) flies reared at 18°C then transferred to 29°C for 4 days and fed silver nitrate for one week. Upper and middle panels show the pericardial nephrocytes in control flies containing silver (brown pigment, arrows) but not when dKlf15 had been silenced (arrowheads). Scale bar = 100μm and 50μm. Lower panels show that pericardial nephrocytes of both genotypes could still be identified with wheat-germ agglutinin (red). Scale bar = 20μm. (G &H) Quantification of nephrocytes and the percentage of nephrocytes containing silver nitrate. ns = not significantly different from the control, ***P<0.001; n = 8 flies per genotype.
Over-expression of dKlf15 increases nephrocyte numbers and rescues age-related decline in nephrocyte function
During these functional studies it became apparent that nephrocyte function declined as flies aged. Given that dKlf15 was required for adult nephrocyte function it was hypothesised that dKlf15 over-expression might rescue the age-dependent nephrocyte dysfunction. Over-expression of dKlf15 driven by Dorothy-Gal4 led to an increase in nephrocyte numbers due to the accumulation of small nephrocytes close to the conical chamber (Fig 7). Over-expression of dKlf15, despite leading to the production of the population of smaller nephrocyte, did not affect the expression of phenotypic markers (Amnionless) nor the function of the nephrocytes in young (1-week-old) flies (Fig 7A and 7B). All nephrocytes in all genotypes were capable of binding dextran when flies were one week of age (Fig 7C), however there was a significant reduction in the proportion binding dextran at seven weeks (reduced to 56% and 62% of nephrocytes in wild type and control flies, respectively P<0.001; Fig 7D). In stark contrast to nephrocytes in wild type and control flies, there was no such reduction in the proportion of nephrocytes binding dextran in 7-week-old flies over-expressing dKlf15 (Fig 7D).
Fig 7. dKlf15 overexpression increases the number of functional nephrocytes in adults and prevents age-dependent nephrocyte dysfunction.
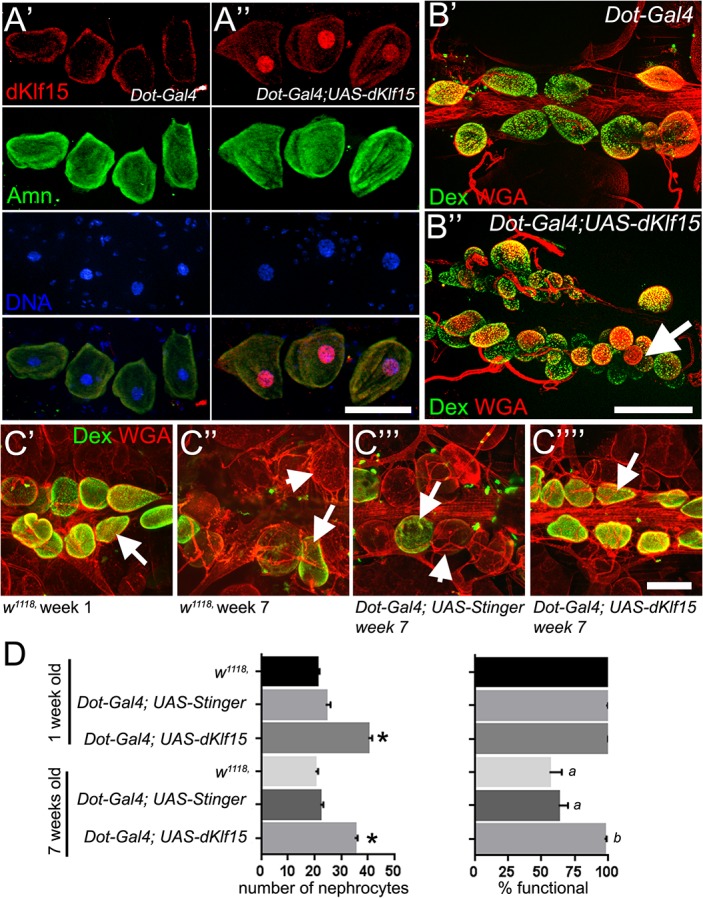
(A) Pericardial nephrocytes from control (progeny of Dot-Gal4 outcrossed to w 1118; A’) and dKlf15 over-expressing flies (Dot-Gal4; UAS-dKlf15; A”) stained with anti-dKlf15 (1:100) and anti-Amnionless antibodies (1:100). Scale bar = 50 μm. (B’) Pericardial nephrocytes from control (progeny of Dot-Gal4 outcrossed to w 1118; B’) and dKlf15 over-expressing flies (Dot-Gal4; UAS-dKlf15; B”) incubated with 10 kDa dextran and wheat germ agglutinin (to label cell surfaces). Arrow indicates a group of smaller nephrocytes. Scale bar = 80 μm. (C’-C”“) Nephrocytes from one and seven-week-old flies, incubated with 10 kDa dextran (green) and wheat germ agglutinin (red). Arrows indicate nephrocytes that have accumulated dextran; arrowheads identify nephrocytes that have not accumulated dextran. (D) Quantification of nephrocyte numbers and the proportion which were functional as determined by their ability to bind dextran. *P<0.001 compared to wild type and control; a P<0.001 compared to one-week-old wild type and control; b P<0.001 compared to seven-week-old wild type and control; n = 8–12 flies per genotype / per time-point.
Discussion
Multicellular organisms contain circulating fluids which must be filtered to remove toxins, effete cells and endogenous macromolecules. Whether this is mammalian blood or invertebrate hemolymph, elaborate filtration systems have evolved which maintain organ homeostasis. In insects the principal filtration cell is the nephrocyte. Nephrocytes have long been regarded as functionally analogous to highly endocytic scavenger cells within the mammalian reticuloendothelial system [4, 27, 28]. Recent reports support this, indicating that there is significant genetic overlap between insect nephrocytes and podocytes, a highly specialised cell-type crucial for glomerular filtration in the human kidney [5, 6, 29, 30]. Nephrocytes also express Amnionless and Cubilin, which form a receptor complex required for nephrocyte filtration function and which is expressed by highly endocytic mammalian cell types, including kidney proximal tubule cells [7]. Hence, the similarities between invertebrate and vertebrate filtration systems appear to share a common genetic basis. The current work reinforces this view by demonstrating that the insect ortholog of Kidney Krüppel-like factor (Klf15), a gene required for human renal podocyte differentiation, acts upstream of Amnionless and the slit diaphragm gene sticks and stones, to drive and maintain the differentiation of Drosophila nephrocytes.
In contrast to the restricted pattern of dKlf15 expression in Drosophila, mammalian Klf15 is expressed in a variety of tissues, suggesting diversification of the ortholog’s role during evolution. Although transcriptional activity of dKlf15 has been demonstrated in two independent, in vitro studies [31, 32] the specific genomic targets in insect nephrocytes remain undefined. Human KLF15 is known to directly activate NPHS1 and NPHS2 [13], which encode Nephrin and Podocin, proteins critical for podocyte slit diaphragm formation. Given that dKlf15 is restricted to nephrocytes, the only insect cell type known to form filtration slits, it is plausible the role of mammalian Klf15 in podocyte biology has been evolutionarily conserved and that dKlf15 may directly activate sns, the Drosophila ortholog of human NPHS1. This hypothesis is consistent with the observed loss of sns expression and lack of slit diaphragms in the pericardial nephrocytes of dKlf15 NN mutants. Garland cells are known to develop as mononuclear cells which then fuse to form binucleate, mature nephrocytes. This fusion process is dependent on sns and modulation of fusion can lead to multi-nucleate garland cells [6, 30]. Overexpression of neph genes as well as hyperactivation of Src64B, a kinase that regulates the slit-diaphragm protein encoded by Dumbfounded, can lead to multinucleate garland cells through agglutination, a phenotype reminiscent of the poly-nucleate morphology seen in dKlf15 NN mutants [29, 30]. How the loss of dKlf15 function leads to a phenotype similar to that seen in gain-of-function experiments is unclear.
Expression of a fluorescent reporter gene (RedStinger) driven by the presumed cis acting enhancer element of dKlf15 was limited to garland cells and pericardial nephrocytes. This corroborated high throughput datasets and anti-dKlf15 immuno-staining results, suggesting the enhancer element used to drive RedStinger provided faithful representation of the dKlf15 expression pattern. Attempts to perform in situs on larval tissue were unsuccessful, reflecting the possibly that dKlf15 expression is too low to be detected, so it is not known with certainty when dKlf15 expression is switched-on in pericardial nephrocytes. However, the timing of nephrocyte attrition and RedStinger reporter gene expression, from the L2 stage onwards, would strongly suggest that dKlf15 expression is initiated in pericardial nephrocytes soon after the L1 stage. No attempt to study the timing of dKlf15 expression in embryonic garland cells was made but high throughput data indicates dKlf15 is expressed in garland cells after embryonic stage 10 [17]. Given that dKlf15 was required for the differentiation and sustained function of pericardial nephrocytes it will be interesting to examine how dKlf15 affects the development and function of the garland cells.
Garland cells and pericardial nephrocytes develop in different anatomical locations, at different times, suggesting that at least two independent signalling pathways initiate dKlf15 expression during development. In addition, one of these pathways or possibly a third pathway regulates the sustained expression of dKlf15 throughout adulthood. In mammals, Klf15 expression is positively regulated by retinoic acid [13] and glucocorticoids [33]. Retinoid signalling affects eye development and tissue regeneration during larval development in Drosophila [34, 35]. Insects do not synthesise glucocorticoids, however analogous endocrine systems do exists in the form of the steroid hormone ecdysone, a hormone critical to many aspects of insect development. It is therefore plausible that dKlf15 gene expression is modulated by endocrine signalling pathways similar to those in mammals. It will be important to elaborate the pathway(s) controlling dKlf15 expression in order to identify both upstream regulators of Klf15 expression and the gene’s downstream effectors as these may provide insights into human KLF15 function.
It is well established that mutations affecting nephrocyte function reduce survivorship of Drosophila larvae that ingest silver nitrate [5, 21]. Given that pericardial nephrocyte attrition occurred during larval development in dKlf15 NN mutants, the observed sensitivity to silver nitrate was expected. In contrast, survivorship of adult mutants given silver nitrate was not different to that of controls fed the same diet, despite both genotypes showing sensitivity to the xenotoxin. This argues that nephrocytes are the primary system for mitigating xenotoxin insults in larvae but that other cells compensate for the loss in adults. It has been reported that the ablation of post-embryonic pericardial nephrocytes shortens adult lifespan [21], but this was not recapitulated in the present study. Because lifespan is influenced by genetic background [36], genetic differences in the present studies were minimised by extensive backcrossing. Whether this was also done in the post-embryonic nephrocyte ablation model is unclear but it may explain the discrepancy between the studies.
Pericardial nephrocytes are thought to modulate cardiac function [37], and preliminary studies suggest heart function is disrupted by dKlf15 loss of function. This is an important area to follow-up using this model because mechanisms affecting cardiac function in Drosophila can be of direct relevance to human cardiovascular physiology [38]. It is also known that nephrocytes modulate the immune response by clearing circulating Necrotic protein [33]. Beyond this observation, it is not known to what extent insect nephrocytes control circulating endogenous macromolecules. Given the intractability of vertebrate models with which to study the clearance and turnover of circulating proteins by scavenger cells within the reticuloendothelial system, the fly and genetic ablation of nephrocyte using dKlf15-loss of function, offers an attractive alternative for such studies.
In summary dKlf15, is defined as a nephrocyte-restricted transcription factor critically required for the differentiation and sustained function of insect nephrocytes. As nephrocytes are important model of human renal podocytes the model has great potential to guide and support research into podocytopathies [39], an important class of kidney pathologies relevant to human cardiovascular health.
Materials and Methods
Strains used in this study
The Canton Special (CaS), w 1118 (used as wild-type strains in this study), Bteb2 f06447 (CG2932, FBgn0025679; called dKlf15 NN in this study), Dorothy-Gal4 (Dot-Gal4, originally described in [19], UAS-mCherry (TRiP control line), Tub-Gal80 ts, Dp(1;3)122, Dp(1;3)120, Dp(1;3)473 and Df(1)ED6727 lines were all from the Bloomington Stock Centre (Bloomington, USA, IL). The HandC-Gal4 (Hand-Gal4) line was described in [20]). The Hand-GFP line was from Dr. Zhe Han (described in [24]). The two RNAi lines for knocking-down Bteb2 / dKlf15 were from the Vienna Drosophila RNAi stock Centre (with a targeting hairpin inserted into the second chromosome, VDRC) and Bloomington (Bteb2 JF02420, a Harvard TRiP line with a Bteb2 targeting hairpin inserted into the third chromosome). All genetic combinations were generated by standard crosses. The sns-mCherryNLS line was kindly provided by Susan Abmayr and is described in [40]. Generation of TARGET flies (Hand-Gal4; Tub-Gal80 ts) was achieved by standard crossing techniques [26].
Husbandry, propagation of flies, survivorship on silver nitrate and determination of lifespan
Flies were propagated routinely on a standard cornmeal-yeast diet at 25°C under a 12hr:12hr light-dark schedule. For TARGET experiments flies were reared at 18°C and then transferred to 29°C within 1–5 days of eclosing. Flies remained at 29°C for one to two weeks. Prior to analysis of heart function flies were transferred to 25°C for 24 hours. To reduce the effect of genetic background the dKlf15 NN mutant was backcrossed onto the w 1118 for >20 generations. Fur establishing survivorship of larvae on silver nitrate, 20 eggs from control or dKlf15 NN mutants were placed onto agar with yeast paste containing different concentrations of silver nitrate. Survivorship was calculated as the percentage of adults eclosing. For the determination of lifespan, male or female flies were collected within 1–3 days of eclosion and housed in vials with free access to food (cornmeal-molasses diet) in groups of 15. Flies were tipped to fresh food twice a week and the number of dead flies recorded until all flies were dead.
Generation of antibodies to dKlf15 and dAmnionless
Antisera were prepared by GenoSphere Biotechnologies (Paris, France). The immunogen chosen for dKlf15 was the peptide CPPDLSDWEQRLLDN, whereas the peptide for dAmnionless was DPRLWRWRHLGLRLR. Antisera to the peptides were generated in rabbits (dKlf15) and guinea pigs (dAmnionless). Antisera were affinity purified to yield the total IgG fraction. Antibodies were verified for specificity by ELISA using the peptide as target antigen. Pre-incubating antibodies with a 10-fold excess of peptide blocked all immunoreactivity in adult flies. Anti-dKlf15 antisera was used at 1:10 to detect protein in wild-type flies and 1:100 in flies over-expressing dKlf15. Anti-Amnionless antisera were used at 1:100.
Generation of transgenic dKlf15-Gal4 flies
To generate the dKlf15-Gal4 line, a 2131bp fragment of DNA containing sequence upstream of the predicted 5’ translation initiation site of the dKlf15 gene was cloned into pPTGAL4 and transgenic flies generated by a commercial company (BestGene Inc, CA, USA).
rtPCR
PCR was performed as described previously [41]. Primers for detecting dKlf15 gene expression were as follows: (left) GGAGGAGAGCAACAGCAATC; (right) CATCGTGTCCTTGGAATGTG. This amplified a 538bp amplicon, between base pairs 420 and 957 of the 962bp dKlf15 transcript. In the dKlf15 NN mutant this region is downstream of the P-element insertion (inserted at the 311st bp; see red bars in Fig 2A). The dKlf15 gene has no introns, so genomic DNA was reduced by DNase treatment. No amplicons were detected in cDNA prepared from whole wild-type adult flies, ovaries or larvae, so isolated hearts were used to prepare cDNA to establish expression in adult flies.
Imaging the larval and adult heart
Larvae were anaesthetised with Flynap (Carolina Biological Supply Company, Burlington, NC, USA) for 30 minutes, placed dorsal side up, on a microscope slide and then cover-slipped so as to partially flatten but not burst the larvae. Adult were dissected and hearts stained as described previously [41]. For some experiments, vital dyes were used to identify functional nephrocytes or test their endocytic function (wheat germ agglutinin at 1 μg/mL for 15 minutes or 50 μg/mL 10kDa fluorescently labelled dextran for 0–30 minutes). Semi-intact preparations were then washed three times, fixed for 20 minutes with 1% formaldehyde and co-stained with antibodies (and then the relevant secondary antibodies) or Hoechst to visualise DNA and then imaged.
Immunostaining in embryos
Eggs were collected from agar plates every 24h. Embryos were dechorionized in 50% bleach and fixed as described before [42]. Primary antibodies used were: rabbit anti-Tin (1:800) [43], rabbit anti-Odd (1:200) [44], mouse anti-Eve (1:5)(Developmental Studies Hybridoma Bank) and guinea pig anti-β3-Tubulin (1:1.000)[45]. Confocal stacks were acquired using a Zeiss LSM 5 Pascal and pictures were processed using ImageJ. Cells were counted in confocal stacks of stained embryos of both genotypes at stage 17. To test the statistical significance of the acquired numbers an unpaired, two-tailed student’s t-Test was used.
Tissue preparation and TEM imaging
Pericardial nephrocytes in third instar larvae were analysed by transmission electron microscopy (TEM) in semi-dissected animals. Therefore, larvae were collected, dissected in cold PBS and fixed in 3.7% formaldehyde for 1h at room temperature. Further fixation and preparation for TEM imaging was essentially done as described in [46]. Samples were analysed using a 60kV Zeiss 902 electron microscope and images were processed using ImageJ and Photoshop CS.
Analysis of endocytosis using 10kDa dextran488
Adult flies were anaesthetised by brief exposure to CO2 and then dissected as for heart analysis (see above). The semi-intact preparation was then incubated in a solution of Alexa488 fluorescently labelled 10kDa dextran (made with Hanks basic saline solution (HBSS, Sigma, Dorset UK) containing 2mM CaCl2), at 25°C for 0–30 minutes. Endocytosis was then inhibited by placing the preparation on ice, washing with ice-cold HBSS and fixing samples for 15 minutes with 4% formaldehyde in HBSS. Some samples were co-incubated with wheat-germ agglutinin (WGA) conjugated to the Alexa594 fluorochrome.
Epifluorescence and confocal microscopy and image analysis
Fluoresce microscopy of flies was performed with a Zeiss Axioskop MOT II microscope (Carl Zeiss, Welwyn Garden City, UK) illuminated with mercury or halogen light sources for fluorescence or differential interference contrast (DIC) imaging using a 40x objective. Images were captured with an ORCA-ER CCD camera (Hamamatsu Photonics KK, Japan; Welwyn Garden City, UK) coupled to Openlab 4.1 (Improvision, Coventry, UK). Confocal images were collected with a Zeiss LSM 780 coupled to Zen imaging software. Images were coloured, contrast enhanced and overlaid using Photoshop CS3. All micrographs from an experiment were collected using the same microscope settings and image alterations. For quantification of fluorescence, micrographs were opened in ImageJ and converted to 8-bit greyscale format. The line tool was then used to draw across a region of interest and the maximum pixel intensity recorded.
Statistics
When more than two genotypes or treatments were used in an experiment one-way ANOVA was used to test the hypothesis that genotype may have affected heart function and post hoc test (Tukey’s HSD) was used to establish P values between control and the different genotypes. If only two genotypes or treatments were used an unpaired two-tailed student’s t-Test was applied to establish the P-value. For endocytosis experiments a 2-way ANOVA was used to examine if there was an interaction between the fluorescence signal, time and genotype.
Supporting Information
(DOCX)
(DOCX)
(DOCX)
(DOCX)
(DOCX)
(DOCX)
(DOCX)
Acknowledgments
We thank the TRiP at Harvard Medical School (NIH/NIGMS R01-GM084947) for providing transgenic RNAi fly stocks used in this study. PSH acknowledges support of the BHF CoRE award (RE/08/001). PSH is especially grateful to Dr. Georg Vogler and Ms. Hanah Catan (Sanford Burnham Medical Research Institute) for their comments on, and assistance with, experiments and the technical assistance of Dr. Richard Marley and Ms. Ashleigh Cheyne. PSH would like to dedicate this manuscript to the memory of Professor Tony Harmar.
Data Availability
All relevant data are within the paper and its Supporting Information files.
Funding Statement
PSH is supported by a British Heart Foundation Intermediate Basic Science Fellowship (FS/13/17/29905). This work was also supported by a British Heart Foundation Centre of Research Excellence (CoRE) award to the University of Edinburgh and by a subsistence grant from the Scottish Universities Life Science Alliance (SULSA) to PSH. KO is supported by American Heart Association award 0835243N. KO is supported by AHA grant 14GRNT20490239. RB is supported by NIH grants R01 HL054732, P01 HL098053, P01 AG033561. A.P. received support from the DFG (SFB944) and the DAAD/IPID program. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Sorensen KK, McCourt P, Berg T, Crossley C, Le Couteur D, Wake K, et al. The scavenger endothelial cell: a new player in homeostasis and immunity. Am J Physiol Regul Integr Comp Physiol. 2012;303(12):R1217–30. 10.1152/ajpregu.00686.2011 . [DOI] [PubMed] [Google Scholar]
- 2. Das D, Aradhya R, Ashoka D, Inamdar M. Macromolecular uptake in Drosophila pericardial cells requires rudhira function. Experimental cell research. 2008;314(8):1804–10. Epub 2008/03/22. 10.1016/j.yexcr.2008.02.009 . [DOI] [PubMed] [Google Scholar]
- 3. Soukup SF, Culi J, Gubb D. Uptake of the necrotic serpin in Drosophila melanogaster via the lipophorin receptor-1. PLoS genetics. 2009;5(6):e1000532 Epub 2009/06/27. 10.1371/journal.pgen.1000532 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Wigglesworth VB. The pericardial cells of insects: analogue of the reticuloendothelial system. Journal of the Reticuloendothelial Society. 1970;7(2):208–16. Epub 1970/02/01. . [PubMed] [Google Scholar]
- 5. Weavers H, Prieto-Sanchez S, Grawe F, Garcia-Lopez A, Artero R, Wilsch-Brauninger M, et al. The insect nephrocyte is a podocyte-like cell with a filtration slit diaphragm. Nature. 2009;457(7227):322–6. Epub 2008/10/31. 10.1038/nature07526 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Zhuang S, Shao H, Guo F, Trimble R, Pearce E, Abmayr SM. Sns and Kirre, the Drosophila orthologs of Nephrin and Neph1, direct adhesion, fusion and formation of a slit diaphragm-like structure in insect nephrocytes. Development. 2009;136(14):2335–44. Epub 2009/06/12. 10.1242/dev.031609 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Zhang F, Zhao Y, Chao Y, Muir K, Han Z. Cubilin and amnionless mediate protein reabsorption in Drosophila nephrocytes. Journal of the American Society of Nephrology: JASN. 2013;24(2):209–16. 10.1681/ASN.2012080795 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. He Q, Madsen M, Kilkenney A, Gregory B, Christensen EI, Vorum H, et al. Amnionless function is required for cubilin brush-border expression and intrinsic factor-cobalamin (vitamin B12) absorption in vivo. Blood. 2005;106(4):1447–53. 10.1182/blood-2005-03-1197 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. McConnell BB, Yang VW. Mammalian Kruppel-like factors in health and diseases. Physiological reviews. 2010;90(4):1337–81. Epub 2010/10/21. 10.1152/physrev.00058.2009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Jeyaraj D, Haldar SM, Wan X, McCauley MD, Ripperger JA, Hu K, et al. Circadian rhythms govern cardiac repolarization and arrhythmogenesis. Nature. 2012;483(7387):96–9. Epub 2012/03/01. 10.1038/nature10852 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Jeyaraj D, Scheer FA, Ripperger JA, Haldar SM, Lu Y, Prosdocimo DA, et al. Klf15 orchestrates circadian nitrogen homeostasis. Cell Metab. 2012;15(3):311–23. Epub 2012/03/13. 10.1016/j.cmet.2012.01.020 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Gao X, Huang L, Grosjean F, Esposito V, Wu J, Fu L, et al. Low-protein diet supplemented with ketoacids reduces the severity of renal disease in 5/6 nephrectomized rats: a role for KLF15. Kidney international. 2011;79(9):987–96. Epub 2011/01/21. 10.1038/ki.2010.539 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Mallipattu SK, Liu R, Zheng F, Narla G, Ma'ayan A, Dikman S, et al. Kruppel-Like factor 15 (KLF15) is a key regulator of podocyte differentiation. J Biol Chem. 2012. Epub 2012/04/12. 10.1074/jbc.M112.345983 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Fisch S, Gray S, Heymans S, Haldar SM, Wang B, Pfister O, et al. Kruppel-like factor 15 is a regulator of cardiomyocyte hypertrophy. Proc Natl Acad Sci U S A. 2007;104(17):7074–9. Epub 2007/04/18. 10.1073/pnas.0701981104 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Wang B, Haldar SM, Lu Y, Ibrahim OA, Fisch S, Gray S, et al. The Kruppel-like factor KLF15 inhibits connective tissue growth factor (CTGF) expression in cardiac fibroblasts. J Mol Cell Cardiol. 2008;45(2):193–7. Epub 2008/07/01. 10.1016/j.yjmcc.2008.05.005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Vilella AJ, Severin J, Ureta-Vidal A, Heng L, Durbin R, Birney E. EnsemblCompara GeneTrees: Complete, duplication-aware phylogenetic trees in vertebrates. Genome research. 2009;19(2):327–35. Epub 2008/11/26. 10.1101/gr.073585.107 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Tomancak P, Berman BP, Beaton A, Weiszmann R, Kwan E, Hartenstein V, et al. Global analysis of patterns of gene expression during Drosophila embryogenesis. Genome Biol. 2007;8(7):R145 Epub 2007/07/25. 10.1186/gb-2007-8-7-r145 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Chintapalli VR, Wang J, Dow JA. Using FlyAtlas to identify better Drosophila melanogaster models of human disease. Nat Genet. 2007;39(6):715–20. Epub 2007/05/31. . [DOI] [PubMed] [Google Scholar]
- 19. Kimbrell DA, Hice C, Bolduc C, Kleinhesselink K, Beckingham K. The Dorothy enhancer has Tinman binding sites and drives hopscotch-induced tumor formation. Genesis. 2002;34(1–2):23–8. Epub 2002/09/27. 10.1002/gene.10134 . [DOI] [PubMed] [Google Scholar]
- 20. Sellin J, Albrecht S, Kolsch V, Paululat A. Dynamics of heart differentiation, visualized utilizing heart enhancer elements of the Drosophila melanogaster bHLH transcription factor Hand. Gene expression patterns: GEP. 2006;6(4):360–75. Epub 2006/02/04. 10.1016/j.modgep.2005.09.012 . [DOI] [PubMed] [Google Scholar]
- 21. Das D, Aradhya R, Ashoka D, Inamdar M. Post-embryonic pericardial cells of Drosophila are required for overcoming toxic stress but not for cardiac function or adult development. Cell and tissue research. 2008;331(2):565–70. Epub 2007/11/08. 10.1007/s00441-007-0518-z . [DOI] [PubMed] [Google Scholar]
- 22. Han Z, Bodmer R. Myogenic cells fates are antagonized by Notch only in asymmetric lineages of the Drosophila heart, with or without cell division. Development. 2003;130(13):3039–51. Epub 2003/05/21. . [DOI] [PubMed] [Google Scholar]
- 23. Alvarez AD, Shi W, Wilson BA, Skeath JB. pannier and pointedP2 act sequentially to regulate Drosophila heart development. Development. 2003;130(13):3015–26. Epub 2003/05/21. . [DOI] [PubMed] [Google Scholar]
- 24. Han Z, Yi P, Li X, Olson EN. Hand, an evolutionarily conserved bHLH transcription factor required for Drosophila cardiogenesis and hematopoiesis. Development. 2006;133(6):1175–82. Epub 2006/02/10. 10.1242/dev.02285 . [DOI] [PubMed] [Google Scholar]
- 25. Togel M, Pass G, Paululat A. The Drosophila wing hearts originate from pericardial cells and are essential for wing maturation. Dev Biol. 2008;318(1):29–37. Epub 2008/04/24. 10.1016/j.ydbio.2008.02.043 . [DOI] [PubMed] [Google Scholar]
- 26. McGuire SE, Mao Z, Davis RL. Spatiotemporal gene expression targeting with the TARGET and gene-switch systems in Drosophila. Sci STKE. 2004;2004(220):pl6. Epub 2004/02/19. . [DOI] [PubMed] [Google Scholar]
- 27. Aggarwal SK, King RC. The ultrastructure of the wreath cells of Drosophila melanogaster larvae. Protoplasma. 1967;63(4):343–52. Epub 1967/01/01. . [DOI] [PubMed] [Google Scholar]
- 28. Crossley AC. The ultrastructure and function of pericardial cells and other nephrocytes in an insect: Calliphora erythrocephala. Tissue & cell. 1972;4(3):529–60. . [DOI] [PubMed] [Google Scholar]
- 29. Tutor AS, Prieto-Sanchez S, Ruiz-Gomez M. Src64B phosphorylates Dumbfounded and regulates slit diaphragm dynamics: Drosophila as a model to study nephropathies. Development. 2014;141(2):367–76. 10.1242/dev.099408 . [DOI] [PubMed] [Google Scholar]
- 30. Helmstadter M, Luthy K, Godel M, Simons M, Ashish, Nihalani D, et al. Functional study of mammalian Neph proteins in Drosophila melanogaster. PLoS One. 2012;7(7):e40300 10.1371/journal.pone.0040300 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Brown JL, Grau DJ, DeVido SK, Kassis JA. An Sp1/KLF binding site is important for the activity of a Polycomb group response element from the Drosophila engrailed gene. Nucleic acids research. 2005;33(16):5181–9. Epub 2005/09/13. 10.1093/nar/gki827 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Zhu LJ, Christensen RG, Kazemian M, Hull CJ, Enuameh MS, Basciotta MD, et al. FlyFactorSurvey: a database of Drosophila transcription factor binding specificities determined using the bacterial one-hybrid system. Nucleic acids research. 2011;39(Database issue):D111–7. Epub 2010/11/26. 10.1093/nar/gkq858 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Sasse SK, Mailloux CM, Barczak AJ, Wang Q, Altonsy MO, Jain MK, et al. The glucocorticoid receptor and KLF15 regulate gene expression dynamics and integrate signals through feed-forward circuitry. Mol Cell Biol. 2013;33(11):2104–15. 10.1128/MCB.01474-12 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Arshavsky VY. Vision: the retinoid cycle in Drosophila. Curr Biol. 2010;20(3):R96–8. 10.1016/j.cub.2009.12.039 . [DOI] [PubMed] [Google Scholar]
- 35. Halme A, Cheng M, Hariharan IK. Retinoids regulate a developmental checkpoint for tissue regeneration in Drosophila. Curr Biol. 2010;20(5):458–63. 10.1016/j.cub.2010.01.038 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Burnett C, Valentini S, Cabreiro F, Goss M, Somogyvari M, Piper MD, et al. Absence of effects of Sir2 overexpression on lifespan in C. elegans and Drosophila. Nature. 2011;477(7365):482–5. 10.1038/nature10296 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Lim HY, Wang W, Chen J, Ocorr K, Bodmer R. ROS Regulate Cardiac Function via a Distinct Paracrine Mechanism. Cell reports. 2014. 10.1016/j.celrep.2014.02.029 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Neely GG, Kuba K, Cammarato A, Isobe K, Amann S, Zhang L, et al. A global in vivo Drosophila RNAi screen identifies NOT3 as a conserved regulator of heart function. Cell. 2010;141(1):142–53. Epub 2010/04/08. 10.1016/j.cell.2010.02.023 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Grahammer F, Schell C, Huber TB. The podocyte slit diaphragm—from a thin grey line to a complex signalling hub. Nat Rev Nephrol. 2013;9(10):587–98. 10.1038/nrneph.2013.169 . [DOI] [PubMed] [Google Scholar]
- 40. Haralalka S, Shelton C, Cartwright HN, Katzfey E, Janzen E, Abmayr SM. Asymmetric Mbc, active Rac1 and F-actin foci in the fusion-competent myoblasts during myoblast fusion in Drosophila. Development. 2011;138(8):1551–62. 10.1242/dev.057653 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Catterson JH, Heck MM, Hartley PS. Fermitins, the orthologs of mammalian Kindlins, regulate the development of a functional cardiac syncytium in Drosophila melanogaster. PloS one. 2013;8(5):e62958 10.1371/journal.pone.0062958 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Sellin J, Drechsler M, Nguyen HT, Paululat A. Antagonistic function of Lmd and Zfh1 fine tunes cell fate decisions in the Twi and Tin positive mesoderm of Drosophila melanogaster. Dev Biol. 2009;326(2):444–55. 10.1016/j.ydbio.2008.10.041 . [DOI] [PubMed] [Google Scholar]
- 43. Zaffran S, Reim I, Qian L, Lo PC, Bodmer R, Frasch M. Cardioblast-intrinsic Tinman activity controls proper diversification and differentiation of myocardial cells in Drosophila. Development. 2006;133(20):4073–83. 10.1242/dev.02586 . [DOI] [PubMed] [Google Scholar]
- 44. Ward EJ, Coulter DE. odd-skipped is expressed in multiple tissues during Drosophila embryogenesis. Mech Dev. 2000;96(2):233–6. . [DOI] [PubMed] [Google Scholar]
- 45. Leiss D, Hinz U, Gasch A, Mertz R, Renkawitz-Pohl R. Beta 3 tubulin expression characterizes the differentiating mesodermal germ layer during Drosophila embryogenesis. Development. 1988;104(4):525–31. . [DOI] [PubMed] [Google Scholar]
- 46. Lehmacher C, Abeln B, Paululat A. The ultrastructure of Drosophila heart cells. Arthropod Struct Dev. 2012;41(5):459–74. 10.1016/j.asd.2012.02.002 . [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
(DOCX)
(DOCX)
(DOCX)
(DOCX)
(DOCX)
(DOCX)
(DOCX)
Data Availability Statement
All relevant data are within the paper and its Supporting Information files.