

Abstract
Metronidazole has been used clinically for over 50 years as an antiparasitic and broad-spectrum antibacterial agent effective against anaerobic bacteria. However resistance to metronidazole in parasites and bacteria has been reported, and improved second-generation metronidazole analogues are needed. The copper catalysed Huigsen azide-alkyne 1,3-dipolar cycloaddition offers a way to efficiently assemble new libraries of metronidazole analogues. Several new metronidazole-triazole conjugates (Mtz-triazoles) have been identified with excellent broad spectrum antimicrobial and antiparasitic activity targeting Clostridium difficile, Entamoeba histolytica and Giardia lamblia. Cross resistance to metronidazole was observed against stable metronidazole resistant C. difficile and G. lamblia strains. However for the most potent Mtz-triazoles, the activity remained in a therapeutically relevant window.
Keywords: Nitroimidazole, Click chemistry, Antibiotic, Anaerobe
Abbreviations: MIC, minimum inhibition concentration; Mtz, metronidazole; MtzS, metronidazole sensitive; MtzR, metronidazole resistant
Graphical abstract
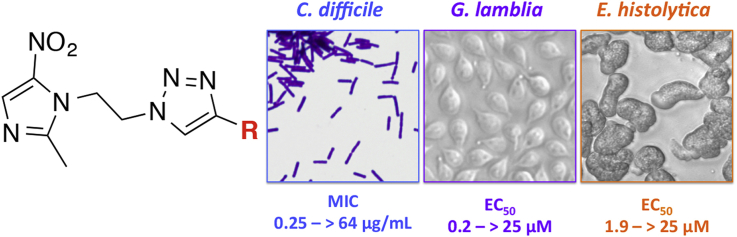
Highlights
-
•
A series of 20 metronidazole (Mtz)-triazole conjugates were synthesised.
-
•
Several Mtz-triazoles had increased potency against pathogenic anaerobes.
-
•
The compounds were not cytotoxic at 100 μM against mammalian cell lines.
-
•
Cross-resistance was observed against Mtz-resistant bacteria and parasites.
-
•
Therapeutically relevant activity was retained against Mtz-resistant G. lamblia.
1. Introduction
For over 50 years the 5-nitroimidazole antibiotic metronidazole 1 has been in clinical use as a broad-spectrum agent for treatment of Gram-positive and Gram-negative anaerobic bacterial infections as well as parasitic infections [1]. Reduction of the nitro group to the nitro radical anion by electron carriers in an anaerobic environment leads to decomposition to form toxic metabolites, which cause DNA damage and nonspecific macromolecular damage leading to cell death [2]. However, metronidazole resistance has been observed in both parasites [3,4] and anaerobic bacteria [5,6]. Over the last ten years metronidazole has also been extensively used to treat Clostridium difficile infection (CDI), an intestinal infection that causes life-threatening severe diarrhea, abdominal pain and fever [7]. The spores produced by C. difficile are resistant to heat and alcohol based disinfectants, complicating eradication and promoting hospital-based outbreaks. Metronidazole is one of three antibiotics routinely prescribed to treat the disease [8]. Metronidazole is effective against mild to moderate C. difficile infection, but, while not wide-spread, resistance has been observed in clinical isolates [9]. To this end, the development of next generation metronidazole analogues that can overcome resistance is therapeutically important.
Beena et al. previously reported the synthesis of metronidazole-triazole conjugates (Mtz-triazoles) with potent activity (reported as IC50 values 0.06–0.35 μg/mL) against the facultative anaerobic bacteria Staphylococcus aureus, Staphylococcus epidermis, Escherichia coli and Pseudomonas aeruginosa [10]. Metronidazole is generally not active against Gram-negative and Gram-positive facultative anaerobic bacteria, so the potent activities of Mtz-triazoles observed by Beena et al. are suggestive of a different mode of action to metronidazole. Given that the active 5-nitroimidazole core (“warhead”) in metronidazole is maintained in Mtz-triazoles, and that the hydroxyl group of metronidazole is amenable to modification [11] (Fig. 1), we reasoned that this class might also exhibit potent activity against the anaerobic bacteria and parasites for which metronidazole is used as a treatment. Therefore, we believed Mtz-triazoles warranted further investigation for both antimicrobial and antiparasitic activity under anaerobic conditions.
Fig. 1.

Metronidazole contains the 5-nitroimidazole warhead and a hydroxyl group amenable to modification.
Independently, during the course of this work, Miyamoto et al. [12] reported a similar approach to Mtz-triazoles, prepared by reaction of six different 5-nitroimidazole azides with a library of alkynes. The compounds were prepared for testing by dilution of the crude reaction mixtures (>85% purity by LCMS) with dimethyl sulfoxide. This methodology bypassed the bottleneck of compound purification and characterization and allowed for the rapid assessment of activity. The solutions of crude material were tested against the parasites Giardia lamblia and Trichomonas vaginalis, the microaerophilic bacteria Helicobacter pylori, the anaerobes C. difficile and Bacteroides fragilis and the facultative anaerobic bacteria E. coli. It is possible that the crude mixtures contained unreacted starting azide 3, which we have shown in this study retains activity, and may influence the reported crude compound potency. A key finding by Miyamoto was that Mtz-triazoles were often active against metronidazole resistant (MtzR) strains of T. vaginalis (47% of compounds) and G. lamblia (100% of compounds), but not against H. pylori frxA and rdxA double mutant (1.4% of compounds) [12]. From this result, we hypothesized that Mtz-triazoles may possess activity against metronidazole resistant C. difficile. More recently Beena et al. described the activity of Mtz-triazoles against the anaerobic protozoan Entamoeba histolytica [13]. They tested a library of 10 Mtz-triazoles and found 4a, 4h, 4p and 4q to have potent activity (IC50 = 0.008–2.36 μM) against E. histolytica HM1: IMSS [13].
We now report an expansion and advancement of these approaches with the synthesis of a small library of purified Mtz-triazoles 4a-t (>95% purity, Scheme 1), including ten novel derivatives (4b, 4c, 4f, 4g, 4i, 4j, 4l, 4o, 4s and 4t). We also explore the antimicrobial and antiparasitic properties of the set of Mtz-triazoles and the parent azide 3 (>95% purity) and determine their activity against additional microbial targets that have not previously been examined. We evaluated the activity of Mtz-triazoles against the parasites E. histolytica and G. lamblia, multiple pathogenic strains of the anaerobe C. difficile, the microaerophile H. pylori and the facultative anaerobic bacterial ESKAPE pathogens E. coli, methicillin resistant S. aureus (MRSA), Klebsiella pneumoniae, Acinetobacter baumanni and P. aeruginosa. The antibacterial activity was measured against a stable MtzR C. difficile strain (CD26A54_R), the parent C. difficile strain with elevated metronidazole minimum inhibitory concentration (MIC) (CD26A54_S) and MtzR H. pylori clinical isolates, while cytotoxicity of the compounds was evaluated against mammalian liver (HepG2) and kidney (HEK293) cell lines.
Scheme 1.

Synthesis of metronidazole-triazole conjugate library. i) MsCl, TEA, DCM, 2 °C to rt, 80 min, 85% yield ii) NaN3, DMF, 50 °C, 21 h, quant. yield iii) CuSO4, sodium ascorbate, MeOH, rt or 45 °C, 20 min–48 h, 9–97% iv) propargyl bromide (80% w/v in toluene), K2CO3, TBAB, rt, 3 days, 16%.
2. Materials and methods
2.1. Synthesis of Mtz-triazole library
Mtz-triazoles were synthesized from metronidazole via an azide intermediate by activation of the hydroxyl group with methanesulfonyl chloride (Scheme 1). Displacement of methanesulfonate 2 with sodium azide provided the desired azide 3 by nucleophilic substitution. The azide substituent was then reacted with a library of alkynes to give 4a-t by copper-catalysed Huisgen 1,3-dipolar cycloaddition with copper sulfate and sodium ascorbate in methanol, with heating to 45 °C and/or additional reagents added if monitoring indicated the reaction was incomplete. All alkynes utilized were commercially available with the exception of pyrazole alkyne 6. Pyrazole alkyne 6 was prepared by reacting pyrazole 5 with propargyl bromide in the presence of potassium carbonate and t-butylammonium bromide as a phase transfer catalyst (Scheme 1) [14]. All compounds were characterized by 1H and 13C NMR, LCMS and HRMS and detailed experimental procedures and characterization are provided in the supplementary information.
2.2. MIC assays
All compounds were tested for activity against a set of anaerobic C. difficile bacteria (630 ATCC BAA-1382, NAP1/027 M7404, NAP1/027 ATCC BAA-1803, VPI10463 ATCC 43255, CD26A54_S and CD26A54_R) and representative ESKAPE pathogens S. aureus (MRSA ATCC 43300), E. coli (ATCC 25922), K. pneumoniae (ATCC 700603), A. baumannii (ATCC 19606) and P. aeruginosa (ATCC 27853) using a standard broth microdilution assay. Selected compounds were tested for activity against H. pylori strains (26695, and clinical isolates MtzS 13/25, MtzR 98/285 and MtzR 13/61) using agar plate dilution, and for antimicrobial activity against twelve Gram-positive bacterial strains: Enterococcus faecalis (VanA clinical isolate), Enterococcus faecium (MDR VanA ATCC 51559), Streptococcus pneumoniae (MDR ATCC 700677), S. aureus (MRSA ATCC 43300, MRSA clinical isolate, MRSA/DRSA clinical isolate, GISA NARSA NRS 1, GISA NARSA NRS 17, VRSA NARSA VRS1, VRSA NARSA VRS4 and VRSA NARSA VRS10) using broth microdilution. C. difficile strains were grown at 37 °C in a COY anaerobic chamber (5% H2, 10% CO2, 85% N2). H. pylori strains were grown at 37 °C under a microaerobic atmosphere (5% O2, 10% CO2). All facultative anaerobic bacteria were grown at 37 °C with normal atmospheric oxygen levels. All experiments were performed in duplicate with metronidazole, vancomycin, linezolid or colistin as positive controls for relevant strains (see Table 1). Positive growth control of bacteria and DMSO as well as a negative control of only media were included for every plate. Full assay details are provided in the supplementary information.
Table 1.
Activity of Mtz-triazoles against C. difficile, ESKAPE bacteria, E. histolytica and G. lamblia.
| Compound | MIC (μg/mL) |
EC50(μM) |
||||||||
|---|---|---|---|---|---|---|---|---|---|---|
|
C. difficilea |
ESKAPE bacteriah |
E. histolytica |
G. lamblia |
|||||||
| 630b |
VPI 10463e |
NAP1/027c |
NAP1/027d |
NAP1/027f |
NAP1/027g |
HM1:IMSS |
WB |
|||
| MtzS | MtzS | MtzS | MtzS | MtzS | MtzR | MtzS | MtzS | |||
| 1 | metronidazole | 0.5 | 0.5 | 0.5 | 0.5 | 1 | 4 | >32 | 5 | 7.9 |
| 3 | azide | 1 | 0.5 | 0.5 | 0.5 | 2 | 8 | >32 | 2.1 | 1.5 |
| 4a | phenyl | 0.5 | 0.25-0.5 | 0.5 | 0.5 | 1 | 2 | >32 | 4.2 | 0.25 |
| 4b | p-OMe-phenyl | 0.5 | 0.25-0.5 | 0.5 | 0.5 | 1 | 4 | >32 | 4.8 | 0.27 |
| 4c | p-Cl-phenyl | 0.5 | 0.25 | 0.25 | 0.25 | 2 | 16 | >32 | 3.6 | 0.20 |
| 4d | p,m-Cl2-phenyl | 0.5 | 0.5-1 | 1 | 1 | 1 | 8-16 | >32 | 5.2 | 0.39 |
| 4e | p-Me-phenyl | 0.5 | 0.25 | 0.5 | 0.5 | 1 | 2 | >32 | 3.7 | 0.18 |
| 4f | methyl ester | 1 | 1 | 1 | 1 | 2 | 4 | >32 | >25 | 3.0 |
| 4g | pyrazole | 1 | 1 | 1 | 1 | 2 | 4 | >32 | >25 | 4.9 |
| 4h | pyridine | 0.5 | 0.5 | 0.5 | 0.5 | 1 | 4 | >32 | 4.6 | 0.74 |
| 4i | 5-pyrimidine | 1 | 1 | 0.5-1 | 1 | 1 | 4 | >32 | >25 | 3.3 |
| 4j | 2-pyrimidine | 2 | 2 | 2 | 2 | 2-4 | 4 | >32 | >25 | 1.7 |
| 4k | benzyl | 0.5 | 0.25-0.5 | 0.5 | 0.5 | 1 | 4 | >32 | 1.9 | 0.70 |
| 4l | CHOH-phenyl | 1 | 0.5 | 1 | 0.5-1 | 2 | 4 | >32 | >25 | 4.4 |
| 4m | CH2NMe-benzyl | 2 | 1-2 | 1 | 2 | 2 | 8 | >32 | 4.6 | 1.6 |
| 4n | thiophene | 0.5 | 0.25-0.5 | 0.5 | 0.5 | 1 | 2 | >32 | 3.8 | 0.37 |
| 4o | thiomorpholine | >64 | >64 | >64 | >64 | >64 | >64 | >32 | >25 | >25 |
| 4p | CH2CH2OH | 8 | 8 | 16 | 8 | 8 | 16 | >32 | >25 | >25 |
| 4q | CH2OH | 16 | 16 | 16-32 | 16 | 16-32 | 16 | >32 | >25 | >25 |
| 4r | CH2CH2COOH | >64 | >64 | >64 | >64 | >64 | >64 | >32 | >25 | >25 |
| 4s | COOH | 64 | 64 | 64 | 64 | >64 | >64 | >32 | >25 | >25 |
| 4t | CH2NH2 | >64 | >64 | >64 | >64 | >64 | >64 | >32 | >25 | >25 |
MIC results determined against CD26A54_S and CD26A54_R after 48 h growth, all other C. difficile strain MIC results determined at 24 h. See Supplementary Table 1 for MIC at 24 h for CD26A54_S and CD26A54_R and at 48 h for ATCC BAA-1803. C. difficile MICs are the median of at least n = 4, except for CD26A54_R where n = 8. ESKAPE pathogen MICs were performed in a single concentration screen with n = 3. EC50 results n = 3.bATCC BAA-1382.cATCC 43255.dM7404.eATCC BAA-1803.fCD26A54_S.gCD26A54_R.hS. aureus MRSA (ATCC 43300), E. coli (ATCC 25922), K. pneumoniae (ATCC 700603), A. baumannii (ATCC 19606) and P. aeruginosa (ATCC 27853).
2.3. Antiparasitic assays
Compounds were tested for antiparasitic activity against E. histolytica (HM1:1MSS strain) and G. lamblia (WB, BRIS/87/HEPU/713 (713) [15], BRIS/83/HEPU/106 (106) [16] and the metronidazole resistant syngeneic line 713-M3 [15,16]) using an ATP-bioluminescence based screen for cell growth and survival [17,18]. Assay plates were inoculated with trophozoites (5 × 103 parasites/well) and incubated in the GasPak™ EZ Anaerobe Gas Generating Pouch Systems (VWR, West Chester, PA) to maintain anaerobic conditions throughout the incubation period. The assays were performed in triplicate using the CellTiter-Glo Luminescent Cell Viability Assay [17]. Metronidazole was used as a positive control.
2.4. Cytotoxicity
Compounds were tested for cytotoxicity against mammalian HepG2 and HEK293 cell lines, as detailed in the supplementary information.
3. Results and discussion
3.1. Design of metronidazole-triazole conjugate library
The Mtz-triazole library was designed to contain a variety of structural groups with some compounds identical to those reported by Beena et al. (4a, 4h, 4p and 4q) [10]. Initial results against C. difficile showed that hydrophobic 4a (R = phenyl) maintained activity relative to metronidazole 1 while more hydrophilic 4p (R = hydroxyethyl) and 4q (R = hydroxymethyl) lost activity. Therefore the subsequent library was biased towards exploring variance of the aromatic ring substitutions, extension of the position of the aromatic ring relative to the triazole core and replacement of the phenyl group with various heterocycles, with several of the latter selected due to their rating in terms of medicinal chemistry ‘developability’ [19]. Amine 4t, and acids 4r and 4s were included to explore the structure activity relationships of non-aromatic ionisable groups. Compounds 4a, 4d, 4e, 4h, 4k, 4m, 4n, 4q and 4r were reported by Miyamoto [12], but 4a, 4h and 4q were not tested against C. difficile. Compounds 4b, 4c, 4f, 4g, 4i, 4j, 4l, 4o, 4s and 4t are described for the first time.
3.2. Antimicrobial and antiparasitic activity of metronidazole-triazole conjugates
The antimicrobial and antiparasitic spectrum of action of Mtz-triazoles was assessed against the anaerobic bacteria C. difficile and the anaerobic parasites E. histolytica and G. lamblia (Table 1). Variations to the phenyl group of 4a were well tolerated and activity of 4b-e was maintained against C. difficile, E. histolytica and G. lamblia (Table 1). Phenyl derivatives 4a-e were several fold more active against G. lamblia than metronidazole. Pyridine 4h and thiophene 4n heterocycles also possessed broad-spectrum activity against C. difficile, E. histolytica and G. lamblia. However, we found that 4h was not as potent compared to metronidazole in the ATP-bioluminescence parasite assay in contrast to the eosin-stain method used by Beena et al. [13] The pyrazole 4g and pyrimidine 4i-j maintained activity against C. difficile within one to two 2-fold dilutions of metronidazole. Compounds 4g and 4i-j were still active against G. lamblia compared to metronidazole, but were several fold less potent than the phenyl derivatives 4a-e. The pyrazole 4g and pyrimidines 4i-j did not inhibit E. histolytica at 25 μM, showing that the R group can be used as a handle to tune the selectivity of this class towards different organisms. Compound 4k (benzyl) and 4m (CH2NMe-benzyl) maintained broad-spectrum activity but 4l (CHOH-phenyl) was inactive against E. histolytica at 25 μM.
Several compounds were inactive, or weakly active, against all three microorganisms at the highest concentration tested. These included the non-aromatic thiomorpholine dioxide 4o, amine 4t and the carboxylic acids 4r and 4s. The methyl hydroxyl 4q and ethyl hydroxyl 4p were inactive against E. histolytica and G. lamblia and weakly potent against C. difficile, continuing the trend of reduced activity with more polar substituents.
We also demonstrate that the parent azide 3 possesses potent activity against C. difficile, E. histolytica and G. lamblia. Therefore, when assessing combinatorial-like libraries of crude material for biological activity, the activity of any unreacted parent compounds is an important consideration, particularly when the compound warhead is maintained.
The MICs of 1, 3 and 4a-t were >32 μg/mL against the representative ESKAPE pathogens (MRSA, E. coli, K. pneumoniae, A. baumannii and P. aeruginosa) tested under aerobic conditions. In addition, the MIC values of a subset of compounds (metronidazole 1, azide 3, 4a (phenyl) and 4q (CH2OH)) were all >64 μg/mL against a panel of 8 additional drug resistant S. aureus strains, vancomycin resistant E. faecalis, vancomycin resistant E. faecium and multidrug resistant S. pneumoniae. This lack of activity is contrary to Beena's report, but consistent with the reported inactivity of 378 Mtz-triazoles against E. coli in Miyamoto's study, and the inactivity of metronidazole against facultative anaerobic bacteria.
3.3. Activity of metronidazole-triazole conjugates against C. difficile strain panel
The activity of the Mtz-triazoles did not vary significantly against multiple strains of C. difficile, including two major pathogenic strains of NAP1/027 and a VPI10463 strain associated with epidemics (Table 1). The MICs of individual compounds typically remained within one 2-fold dilution against the four strains of C. difficile tested. This is important, as viable drug candidates must possess appropriate broad strain coverage. A review of antibiotic drug candidates in development found that activity against multiple strains of C. difficile could vary substantially [20]. This can be explained since C. difficile has a highly mobile, mosaic genome and there is wide strain diversity between isolates [21,22]. Isolates can be divided across five main phylogenetic clades, multiple ribotypes and toxinotypes [23]. Recently, genomic epidemiology studies examining and tracking C. difficile outbreaks in Europe found that a high proportion of strains causing infection were not related to prior infectious strains [22]. Therefore, there is a large pool of genetically diverse strains in the community and selection and spread of intrinsically resistant strains could occur. However, the broad-strain activity of Mtz-triazoles against C. difficile in this study encourages further development of this class.
3.4. Activity of metronidazole-triazole conjugates against metronidazole resistant C. difficile
While metronidazole resistance has been reported in the clinic [9,24], performing antimicrobial susceptibility testing against resistant C. difficile isolates is problematic since the resistance phenotype is unstable and often reported to be lost on freeze thaw cycles or on passaging of the isolates [5]. However, Lynch et al. have reported a stable metronidazole resistant (MtzR) C. difficile clone [25]. The activity of Mtz-triazoles was assessed against this stable MtzR strain of C. difficile (CD26A54_R) and the parent metronidazole sensitive (MtzS) (CD26A54_S) strain, which lost the metronidazole resistance on freeze thawing, but still retained a slightly elevated MIC value to metronidazole compared to the control MtzS NAP1/027 strain. The MIC to metronidazole in BHIS broth at 48 h was lower than the metronidazole MIC determined by E-test on BAKHS (4 μg/mL vs 48 μg/mL), similar to values reported by Chong et al. (8 μg/mL vs > 32 μg/mL) [26].
The resistance to metronidazole became more apparent on incubation in broth for 48 h (MIC = 2 μg/mL after 24 h incubation vs MIC = 4 μg/mL after 48 h) (Supplementary Table 1). In contrast, MICs against the control MtzS NAP1/027 strain ATCC 1803 did not change significantly between 24 h and 48 h (Supplementary Table 1). Although several compounds (4a, 4e and 4n) were active against MtzR C. difficile at 1 dilution lower than metronidazole (MIC = 2 μg/mL vs 4 μg/mL), none of the Mtz-triazoles were active against MtzR C. difficile at levels comparable to the activity of metronidazole against MtzS strains (MIC = 0.5 μg/mL). Reduced activity against the parent C. difficile strain CD26A54_S with intermediary metronidazole susceptibility was also observed.
3.5. Activity of metronidazole-triazole conjugates against additional G. lamblia strains including metronidazole resistant G. lamblia
Selected compounds (4a-f, 4h, 4k-n) were tested against two additional strains of MtzS G. lamblia (106 and 713) and one MtzR strain (713-M3) derived from the parent MtzS strain 713 (Table 2). The Mtz-triazoles displayed similar activity against the additional MtzS strains (106 and 713) compared to G. lamblia strain WB. Encouragingly, the Mtz-triazoles were all more potent than metronidazole against MtzR G. lamblia 713-M3. However, with the exception of 4d, all of the Mtz-triazoles lost activity against the MtzR G. lamblia 713-M3 when compared to the parent MtzS strain 713, similar to the loss of activity against MtzR C. difficile. Mtz-triazoles of scaffold 4 were also all observed by Miyamoto et al. to lose activity against MtzR G. lamblia 713-M3 and 106-2ID10. However, while activity diminished against the MtzR 713M strain, the Mtz-triazoles were often still much more potent than metronidazole itself, meaning that the EC50 remained within a therapeutically meaningful window.
Table 2.
EC50 of selected Mtz-triazoles against G. lamblia strains. The EC50 values for G. lamblia WB strain are shown from Table 3 for comparison.
| Compound | EC50 (μM) |
||||
|---|---|---|---|---|---|
|
G. lamblia | |||||
| WB |
106 |
713 |
713M |
||
| MtzS | MtzS | MtzS | MtzR | ||
| 1 | metronidazole | 7.9 | 2.8 | 2.3 | 17 |
| 4a | phenyl | 0.25 | 0.28 | 0.16 | 2.3 |
| 4b | p-OMe-phenyl | 0.27 | 0.64 | 0.64 | 1.0 |
| 4c | p-Cl-phenyl | 0.20 | 0.89 | 1.2 | 3.5 |
| 4d | p,m-Cl2-phenyl | 0.39 | 1.1 | 2.5 | 1.2 |
| 4e | p-Me-phenyl | 0.18 | 0.29 | 0.29 | 1.0 |
| 4f | methyl ester | 3.0 | 2.0 | 1.8 | 3.1 |
| 4h | pyridine | 0.74 | 0.38 | 0.28 | 0.95 |
| 4k | benzyl | 0.70 | 0.51 | 0.32 | 1.1 |
| 4l | CHOH-phenyl | 4.4 | 2.2 | 2.4 | 3.0 |
| 4m | CH2NMe-benzyl | 1.6 | 1.1 | 0.71 | 2.2 |
| 4n | thiophene | 0.37 | 0.34 | 0.18 | 0.9 |
The activities of purified Mtz-triazoles (>95% purity) against G. lamblia strains 106, 713 and 713M were compared with the literature activities [12] determined with crude reaction mixtures (Supplementary Table 3). The activities were generally in close agreement, supporting the methodology used by Miyamoto [12], although a 3.7–8 fold difference was observed for Mtz-triazoles 4d, 4e and 4m against 1 strain of G. lamblia.
3.6. Activity of metronidazole-triazole conjugates against metronidazole resistant H. pylori
Selected compounds (4f-h, 4m and 4s) were tested against a panel of H. pylori strains using the CLSI agar dilution susceptibility method (Table 3) [27]. These strains included the reference strain 26695 and three clinical isolates 13/25 (metronidazole E-test MIC = 2 μg/mL), 98/285 (E-test MIC = 24 μg/mL) and 13/61 (E-test MIC = 256 μg/mL) with a range of resistance levels to metronidazole determined by the E-test. The susceptibilities of the strains to metronidazole determined by the agar dilution method differed from the susceptibilities obtained from the E-test as has been described previously [28]. The methyl ester 4f was weakly active against H. pylori, while the carboxylic acid 4s was inactive against all H. pylori strains as observed with E. histolytica and G. lamblia as well. Compounds 4g, 4h and 4m were more active than metronidazole against the MtzS strains (26695 and clinical isolate 13/25) and 1–2 dilutions more active than metronidazole against the MtzR strains (clinical isolates 98/285 and 13/61) but they could not completely overcome metronidazole resistance, similar to the loss of activity against metronidazole resistant C. difficile.
Table 3.
MIC of selected Mtz-triazoles against H. pylori strains.
| Compound | MIC (μg/mL) |
||||
|---|---|---|---|---|---|
|
H. pylori | |||||
| 26695 |
13/25a |
98/285a |
13/61a |
||
| MtzS | MtzS | MtzR | MtzR | ||
| 1 | metronidazole | 8 | 4-8 | 64 | 32-64 |
| 4f | methyl ester | 32-64 | 32-64 | 64->64 | 64->64 |
| 4g | pyrazole | 2-4 | 2 | 16-32 | 16-32 |
| 4h | pyridine | 2 | 1 | 32 | 16 |
| 4m | CH2NMe-benzyl | 4 | 2 | 16->64 | 16 |
| 4s | COOH | >64 | >64 | >64 | >64 |
clinical isolate.
3.7. Cytotoxicity
All compounds, including metronidazole 1, azide 3, and 4a-t showed no cytotoxicity against HepG2 and HEK293 cells at concentrations up to 100 μM (CC50). This is consistent with the findings by Miyamoto et al. which did not observe cytotoxicity against HeLa cells at 50 μM [12].
3.8. Relationship of compound activity with compound properties
In order to understand the relationship between compound properties and activity, compound properties including logP (or logD at pH = 7.4), molecular weight, topological polar surface area (tPSA), and predicted solubility (logS) were examined for correlation with activity against MtzS strains of C. difficile, E. histolytica and G. lamblia, with the activity expressed as pMIC (-log10MIC) or pEC50 (-log10EC50)) (Supplementary Table 4). Inactive compounds were included in the analysis at the highest concentration of compound tested. A correlation can be observed between the logD values and the activity against G. lamblia (with R2 (linear regression) = 0.84, Supplementary Fig. 1) (Fig. 2). The logD of the most potent compounds (4a-e, 4h, 4k and 4n) against G. lamblia is between 1.5 and 3.0, with activity decreasing with lower logD. In contrast, only a mild correlation could be detected between logD and the activity against C. difficile (R2 = 0.68, Supplementary Fig. 1) and no correlation for activity against E. histolytica (R2 = 0.48, Supplementary Fig. 1). This might explain why the compounds with aromatic phenyl, thiophene and pyridine groups were more potent against G. lamblia than E. histolytica and C. difficile. Increasing the hydrophobicity (logD) of compounds to improve potency is generally considered undesirable in drug development due to the tendency for increased metabolism [29] and increased promiscuity [30]. However, in this case the antibiotic metronidazole has a low logD to begin with, such that the relatively higher logD of the compounds explored in this study does not place the compounds in an undesirable chemical space. On the other hand, the site of infection of G. lamblia is the small intestine, where low (<1) or high (>3) logD values [29], associated with reduced systemic uptake, might be beneficial for efficacy. However, metronidazole is 100% oral bioavailable [31] and so the importance of bioavailability for in vivo efficacy against G. lamblia infections is unclear.
Fig. 2.

Correlation of activity against G. lamblia, E. histolytica and C. difficile with logD. Compounds with activity against parasites (EC50 < 25 μM) and C . difficile (MIC ≤ 8 μg/mL) have pEC50 or pMIC > 5.0.
4. Conclusion
Mtz-triazoles were synthesized and assessed for activity against the anaerobe C. difficile, microaerophile H. pylori, the parasites E. histolytica and G. lamblia as well as facultative anaerobic Gram-positive and Gram-negative bacteria. While no activity was observed against the facultative anaerobic bacteria, nine compounds were identified with potent broad spectrum activity against anaerobic organisms, while having no cytotoxicity against mammalian cell lines. Hydrophobic R groups such as the benzyl 4k, varyingly substituted phenyl derivatives 4a-e and heterocyclic R groups such as pyridine 4h and thiophene 4n were well tolerated and favored broad spectrum anaerobic activity. In contrast, polar R groups including pyrimidines 4i-j, carboxylic acids 4p and 4r and methyl amine 4t either resulted in loss of broad spectrum activity or were inactive against all organisms tested.
While active Mtz-triazoles displayed a narrow MIC range against multiple strains of C. difficile and G. lamblia, there was metronidazole cross resistance against the stable MtzR C. difficile strain CD26A54_R, MtzR G. lamblia strain 713M and clinical isolates of MtzR H. pylori. Mtz-triazoles 4a-t generally lost activity against MtzR C. difficile, MtzR G. lamblia and MtzR H. pylori. However, since the Mtz-triazoles were often much more potent than metronidazole against G. lamblia, the EC50 remained within a therapeutically meaningful window. It is possible that surveying a more diverse chemical space of R group substituents or using alternative 5-nitroimidazole scaffolds could provide access to Mtz-triazoles that are even more potent and therefore active against MtzR strains of C. difficile and G. lamblia. Future work will focus on identifying such compounds and explore their in vivo efficacy against anaerobic pathogens.
Author contribution
Planned experiments: AMJ, TK, AD, LE, MAB, MAC.
Performed experiments: AMJ, AD, CYT, JH, GK, AG, YM, SR, AK, JZ.
Wrote paper: AMJ, TK, LE, MAB, MAC.
Conflict of interest
The authors declare no competing financial interest.
Acknowledgement
We would like to acknowledge Prof. Dena Lyras and Dr. Kate Mackin for training in C. difficile microbiology and for providing C. difficile M7404. We thank Prof. George Golding for kind provision of C. difficile CD26A54_S and CD26A54_R. We thank Prof. David Patterson for kind provision of Gram positive clinical isolates S. aureus (MRSA clinical isolate, MRSA DRSA clinical isolate), Enterococcus faecalis (VanA Clinical Isolate). The following isolates were obtained through the Network of Antimicrobial Resistance in Staphylococcus aureus (NARSA) program supported under NIAID/NIH Contract #HHSN272200700055C: GISA NARSA NRS 1, GISA NARSA NRS 17, VRSA NARSA VRS1, VRSA NARSA VRS4 and VRSA NARSA VRS10. This research was supported financially by a UQ CIEF grant, NHMRC grants APP631632, APP1026922 and AF511105, and NIH grants AI114671, DE020607 and DK080506. AMJ is supported by an Australian Postgraduate Award (APA) and QLD Government Smart Futures PhD scholarship, TK, SR, AK and MAB by a Wellcome Trust Seeding Drug Discovery Award (094977/Z/10/Z), and MAC by an NHMRC Principal Research Fellowship (APP1059354). CYT and his Helicobacter Research team are supported by the Vice-Chancellor of the University of Western Australia and NHMRC Sir McFarlane Burnett Fellowship grant (No. 572723).
Footnotes
Supplementary data related to this article can be found at http://dx.doi.org/10.1016/j.ejmech.2015.06.019.
Appendix A. Supplementary data
The following is the supplementary data related to this article:
References
- 1.Freeman C.D., Klutman N.E., Lamp K.C. Metronidazole. A therapeutic review and update. Drugs. 1997;54:679–708. doi: 10.2165/00003495-199754050-00003. [DOI] [PubMed] [Google Scholar]
- 2.Muller J., Schildknecht P., Muller N. Metabolism of nitro drugs metronidazole and nitazoxanide in Giardia lamblia: characterization of a novel nitroreductase (GlNR2) J. Antimicrob. Chemother. 2013;68:1781–1789. doi: 10.1093/jac/dkt106. [DOI] [PubMed] [Google Scholar]
- 3.Upcroft J.A., Dunn L.A., Wright J.M., Benakli K., Upcroft P., Vanelle P. 5-Nitroimidazole drugs effective against metronidazole-resistant Trichomonas vaginalis and Giardia duodenalis. Antimicrob. Agents Chemother. 2005;50:344–347. doi: 10.1128/AAC.50.1.344-347.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Meri T., Jokiranta T.S., Suhonen L., Meri S. Resistance of Trichomonas vaginalis to metronidazole: report of the first three cases from Finland and optimization of in vitro susceptibility testing under various oxygen concentrations. J. Clin. Microbiol. 2000;38:763–767. doi: 10.1128/jcm.38.2.763-767.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Peláez T., Cercenado E., Alcalá L., Marín M., Martín-López A., Martínez-Alarcón J. Metronidazole resistance in Clostridium difficile is heterogeneous. J. Clin. Microbiol. 2008;46:3028–3032. doi: 10.1128/JCM.00524-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Kaakoush N.O., Asencio C., Megraud F., Mendz G.L. A redox basis for metronidazole resistance in Helicobacter pylori. Antimicrob. Agents Chemother. 2009;53:1884–1891. doi: 10.1128/AAC.01449-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Antibiotic Resistance Threats in the United States. U.S. Department of Health and Human Services, Centers for Disease Control and Prevention; 2013. http://www.cdc.gov/drugresistance/threat-report-2013/ 2013. [Google Scholar]
- 8.Venugopal A.A., Johnson S. Current state of Clostridium difficile treatment options. Clin. Infect. Dis. 2012;55(Suppl. 2):S71–S76. doi: 10.1093/cid/cis355. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Baines S.D., O'Connor R., Freeman J., Fawley W.N., Harmanus C., Mastrantonio P. Emergence of reduced susceptibility to metronidazole in Clostridium difficile. J. Antimicrob. Chemother. 2008;62:1046–1052. doi: 10.1093/jac/dkn313. [DOI] [PubMed] [Google Scholar]
- 10.Beena N.K., Rohilla R.K., Roy N., Rawat D.S. Synthesis and antibacterial activity evaluation of metronidazole-triazole conjugates. Bioorg. Med. Chem. Lett. 2009;19:1396–1398. doi: 10.1016/j.bmcl.2009.01.037. [DOI] [PubMed] [Google Scholar]
- 11.Abu S.E.A.M., Saadeh H.A.M., Mosleh I.M.I., Al A.M.T.A, Mubarak M.S. 2009. Metronidazole derivatives as antiparasitic agents. EP2085394 A3. [Google Scholar]
- 12.Miyamoto Y., Kalisiak J., Korthals K., Lauwaet T., Cheung D.Y., Lozano R. Expanded therapeutic potential in activity space of next-generation 5-nitroimidazole antimicrobials with broad structural diversity. Proc. Natl. Acad. Sci. U. S. A. 2013;110:17564–17569. doi: 10.1073/pnas.1302664110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Negi B., Raj K.K., Siddiqui S.M., Ramachandran D., Azam A., Rawat D.S. In vitro antiamoebic activity evaluation and docking studies of metronidazole-triazole hybrids. ChemMedChem. 2014;9:2439–2444. doi: 10.1002/cmdc.201402240. [DOI] [PubMed] [Google Scholar]
- 14.Díez-Barra E., La Hoz A.D., Sánchez-Migallón A., Tejeda J. Synthesis of N-alkylpyrazoles by phase transfer catalysis without solvent. Synth. Commun. 1990;20:2849–2853. [Google Scholar]
- 15.Townson S.M., Laqua H., Upcroft P., Boreham P.F., Upcroft J.A. Induction of metronidazole and furazolidone resistance in Giardia. Trans. R. Soc. Trop. Med. Hyg. 1992;86:521–522. doi: 10.1016/0035-9203(92)90095-t. [DOI] [PubMed] [Google Scholar]
- 16.Boreham P.F.L., Phillips R.E., Shepherd R.W. Altered uptake of metronidazole in vitro by stocks of Giardia intestinalis with different drug sensitivities. Trans. R. Soc. Trop. Med. Hyg. 1988;82:104–106. doi: 10.1016/0035-9203(88)90278-7. [DOI] [PubMed] [Google Scholar]
- 17.Debnath A., Parsonage D., Andrade R.M., He C., Cobo E.R., Hirata K. A high-throughput drug screen for Entamoeba histolytica identifies a new lead and target. Nat. Med. 2012;18:956–960. doi: 10.1038/nm.2758. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Tejman-Yarden N., Miyamoto Y., Leitsch D., Santini J., Debnath A., Gut J. A reprofiled drug, auranofin, is effective against metronidazole-resistant Giardia lamblia. Antimicrob. Agents Chemother. 2013;57:2029–2035. doi: 10.1128/AAC.01675-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Ritchie T.J., Macdonald S.J.F., Peace S., Pickett S.D., Luscombe C.N. The developability of heteroaromatic and heteroaliphatic rings – do some have a better pedigree as potential drug molecules than others? Med. Chem. Commun. 2012;3:1062. [Google Scholar]
- 20.Jarrad A.M., Karoli T., Blaskovich M.A.T., Lyras D., Cooper M.A. Clostridium difficile drug pipeline: challenges in discovery and development of new agents. J. Med. Chem. 2015 doi: 10.1021/jm5016846. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Sebaihia M., Wren B.W., Mullany P., Fairweather N.F., Minton N., Stabler R. The multidrug-resistant human pathogen Clostridium difficile has a highly mobile, mosaic genome. Nat. Genet. 2006;38:779–786. doi: 10.1038/ng1830. [DOI] [PubMed] [Google Scholar]
- 22.Eyre D.W., Cule M.L., Wilson D.J., Griffiths D., Vaughan A., O'Connor L. Diverse sources of C. difficile infection identified on whole-genome sequencing. N. Engl. J. Med. 2013;369:1195–1205. doi: 10.1056/NEJMoa1216064. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Dingle K.E., Griffiths D., Didelot X., Evans J., Vaughan A., Kachrimanidou M. Clinical Clostridium difficile: clonality and pathogenicity locus diversity. PLoS One. 2011;6:e19993. doi: 10.1371/journal.pone.0019993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Freeman J., Bauer M.P., Baines S.D., Corver J., Fawley W.N., Goorhuis B. The changing epidemiology of Clostridium difficile infections. Clin. Microbiol. Rev. 2010;23:529–549. doi: 10.1128/CMR.00082-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Lynch T., Chong P., Zhang J., Hizon R., Du T., Graham M.R. Characterization of a stable, metronidazole-resistant Clostridium difficile clinical isolate. PLoS One. 2013;8:e53757. doi: 10.1371/journal.pone.0053757. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Chong P.M., Lynch T., McCorrister S., Kibsey P., Miller M., Gravel D. Proteomic analysis of a NAP1 Clostridium difficile clinical isolate resistant to metronidazole. PLoS One. 2014;9:e82622. doi: 10.1371/journal.pone.0082622. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Methods for Antimicrobial Dilution and Disk Susceptibility Testing of Infrequently Isolated or Fastidious Bacteria; Approved Guideline- Second Addition. 45 ed. Clinical and Laboratory Standards Institute; Pennsylvania: 2013. [Google Scholar]
- 28.Glupczynski Y., Broutet N., Cantagrel A., Andersen L.P., Alarcon T., López-Brea M. Comparison of the E test and agar dilution method for antimicrobial susceptibility testing of Helicobacter pylori. Eur. J. Clin. Microbiol. Infect. Dis. 2002;21:549–552. doi: 10.1007/s10096-002-0757-6. [DOI] [PubMed] [Google Scholar]
- 29.Van de Waterbeemd H., Lennernas H., Artursson P. Chapter 2. High-throughput measurement of log D and pKa. In: Comer J.E.A., editor. Drug Bioavailability. Wiley-VCH Verlag GmbH & Co. KGaA; Weinheim, FRG: 2003. pp. 21–45. [Google Scholar]
- 30.Waring M.J. Lipophilicity in drug discovery. Expert Opin. Drug Discov. 2010;5:235–248. doi: 10.1517/17460441003605098. [DOI] [PubMed] [Google Scholar]
- 31.Jensen J.C., Gugler R. Single- and multiple-dose metronidazole kinetics. Clin. Pharmacol. Ther. 1983;34:481–487. doi: 10.1038/clpt.1983.201. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.