

Abstract
Ischemia-reperfusion (I/R)-mediated intestinal mucosal injury is usually induced by oxygen-derived toxic free radicals from the xanthine oxidase system after reperfusion, but the detailed molecular mechanisms underlying glutamine protection is still unclear. This study aims to elucidate whether glutamine prevents damage to the intestinal mucosa after I/R in rats and to investigate signaling by the Nrf2/ARE pathway induced by GLN in a rat model. Our results revealed that Glutamine pretreatment reduced jejunum injury and microvascular hyper-permeability induced by I/R. MDA level significantly increased while the SOD and GSH-Px levels decreased in the I/R group compared to the sham group and the GLN-I/R group. Both the mRNA and protein levels of the Nrf2 and HO-1 were significantly elevated by GLN pretreatment when compared to the I/R group. GLN treatment also elevated Bcl-2 levels, and accordingly suppressed apoptotic damage in the jejunum cells shown by decreased cleaved caspase-3 level. Mechanistic investigation revealed that GLN treatment augmented binding of Nrf2 onto Bcl2 gene promoter. These results indicate that glutamine has protective effects on I/R in vivo by activating the Nrf2/ARE signaling pathway to inhibit ROS production and reduce intestinal apoptosis.
Keywords: Glutamine, intestinal ischemia-reperfusion injury, Nrf2/ARE signaling pathway, Bcl2
Introduction
Intestinal ischemia/reperfusion (I/R) injury secondary to splanchnic hypoperfusion is a common event after a variety of clinical conditions [1,2]. The intestine is vulnerable to hypoperfusion after injuries such as shock, neonatal necrotizing enterocolitis, intestinal transplant, and mesenteric ischemia [3]. Resuscitation allows intestinal blood flow to recover, resulting in ischemia/reperfusion. Previous studies suggest that the mechanism of I/R is complicated and that reactive oxygen species (ROS) attack, inflammation infiltration, Ca2+ overload, energy metabolism obstacles, and apoptosis may be involved [4-6]. These mechanisms are interconnected, thus ultimately leading to apoptosis and necrosis. Recently, a better understanding of the mechanism(s) of oxidative stress in I/R injury has been achieved. ROS activation, contributing to oxidative stress, is considered the key factor. When I/R occur, ROS activates various signaling pathways and results in inflammatory amplification, while other proteins may also be activated to produce a protective effect. For example, the transcription factor NF-E2-related factor-2 (Nrf2) regulates a major environmental and oxidative stress response. It is held in the cytoplasm by a cytoskeletal-associated specific inhibitory protein, the Kelch-like ECH associating protein 1 (Keap1) in normal quiescent cells [7-10]. Upon stimulation by oxidative stress, cysteine residues within the hinge region of Keap1 can be modified and cause a conformational change resulting in the loss of Nrf2 binding. The Nrf2 is then translocated into the nucleus, where it binds to the antioxidant response element (ARE) in the promoter region of a number of genes encoding anti-oxidative and phase 2 enzymes, such as heme oxygenase 1 (HO-1), superoxide dismutase (SOD), GSH-Px, and NAD(P)H, which can antagonize oxidative stress induced by ROS.
Glutamine is the primary metabolic fuel of small intestinal enterocytes, it constitutes 50% of the free amino acid pool in the body, and is an essential metabolic component for the synthesis of glutathione, because its metabolism by the intestinal epithelium generates substantial quantities of glutamate, a precursor for glutathione synthesis [11]. Enterocytes rely dominantly on glutamine as an essential metabolic precursor in nucleotide, glucose and amino sugar, and protein synthesis [12]. Although classified as “non-essential”, glutamine appears essential for the viability and growth of intestinal cells. Studies have established that glutamine reduced atrophy of the intestinal mucosa in rats on total parenteral nutrition (TPN). The potential mechanisms involved in I/R-mediated intestinal mucosal injury include oxygen-derived toxic free radicals, generated by the activation of the xanthine oxidase system after reperfusion [13]. Studies show that glutamine pre-supplementation improves survival after I/R in mice models [14]. Other experimental studies [15] demonstrate that intraluminal injection of GLN protects the mucosa and diminishes the accumulation of neutrophils in the lamina propria of the small bowel during I/R.
With the discovery that the Nrf2/ARE signaling pathway can reduce the tissue damage induced by ROS, and increasing number of studies have shown that modulating Nrf2/ARE signaling plays a protective role in splanchnic ischemia reperfusion injuries, including heart, brain, and renal injuries. However, no reports addresses whether glutamine protects against intestinal ischemia reperfusion by activating the Nrf2/ARE signaling pathway. The purpose of this study was to elucidate whether GLN prevents damage to the intestinal mucosa after II/R in rats and to investigate signaling by the Nrf2/ARE pathway induced by GLN.
Materials and methods
Animals
Male Wister rats weighing 250-300 g were provided by the Animal Research Center of Shandong University (Jinan, China). The rats were maintained in a 12:12-hour day-night rhythm at a constant temperature of 23°C and a relative humidity of 40-60%. The experiments were performed in adherence with the National Institutes of Health Guidelines for the Use of Laboratory Animals and were approved by the Binzhou Medical University Committee on Animal Care. All efforts were made to minimize the number of animals used and their suffering.
Surgical protocols and histologic evaluation
The rats were randomly assigned into three groups (10 rats per group): the sham group served as a normal control, and the ischemia/reperfusion group (I/R group) was the model control. These two groups received 1 g/kg of normal saline via metal tube gavage every day. The glutamine pretreatment group (GLN-I/R group) received 1 g/kg of glutamine every day for 7 days before surgery. The rats were anesthetized with an intraperitoneal injection of 10% chloral hydrate (3 ml/kg body weight). Rats in the sham group were subjected to a laparotomy and the superior mesenteric artery (SMA) was exposed but not occluded; rats in the I/R and GLN-I/R groups were subjected to a laparotomy and the SMA was occluded with a microvascular clamp for 30 min. The laparotomy incision was then closed, to be opened later for removal of the clamps after 30 min of ischemia [16]. After verification of blood supply to the intestine, the abdominal incision was closed. After 24 h of reperfusion, the incision was opened again, and a 25-cm intestinal segment was obtained from a point 10 cm distal to the ligament of Treiz for morphological and biochemical analysis; another 25-cm intestinal segment was collected and stored at -80°C for western blotting. Intestinal samples were embedded in paraffin and stained with hematoxylin and eosin (H&E staining). Three sections were prepared from each fixed tissue sample, and each slide was analyzed. A blinded observer performed the histologic analyses using the histologic scoring system described by Chiu et al [17]. All measurements were made in triplicate, and mean values were obtained.
Intestinal permeability
The permeability of the intestinal mucosa was assayed by measurement of D-lactate levels and endotoxin levels in serum. Briefly, standard sample (1800 μg/L) was diluted to 1200, 800, 400, 200, and 100 μg/L; then a standard curve was plotted based on the absorbance (OD) of a standard sample at 450 nm. The OD of samples was measured used to calculate the concentration of the sample’s D-lactic acid in rats via the standard curve. Serum was separated from the blood sample by centrifugation and stored at -80°C until analysis. A chromomeric end-point tachypleus amebocyte lysate (CE TAL) test was used to measure serum endotoxin levels. Platelet-rich serum was prepared from the blood by centrifugation at 3000 rpm for 15 min. The serum samples were thawed at room temperature for ~30 min before the assay. LAL (100 μl) was added to 100 μL of serum and incubated at 37°C for 5 min. In the next step, 100 ml chromogenic substrate solution was added and incubated at 37°C for another 10 min. To terminate the reaction, 100 ml HCl was added the OD was read at 545 nm. A standard curve from 0.1 to 1.0 EU/mL was used to evaluate the concentration of endotoxin.
Spectrophotometric measurement of T-SOD, MDA, and GSH-Px in serum
The enzymatic activity of these factors in samples was measured by colorimetric analyses using a spectrophotometer with relevant detection kits according to the instructions of the manufacturer. All measurements of SOD activity, MDA, and GSH-Px content in serum were performed in accordance with the technical manuals of the detection kits (Jianchen Biological Institute, China).
Detection of Nrf2, HO-1, caspase-3 and bcl-2 proteins by immunohistochemistry (IHC) analysis
IHC was performed using an SV hypersensitivity two-step kit (BOSTER, Wuhan, China). Briefly, slides were deparaffinized, and tissue sections were hydrated through xylene and a graded alcohol series. Then, slides were incubated in 3% H2O2 for 30 min to block the endogenous peroxidase activity and treated with citrate buffer (pH 6.0) at 95°C for antigen retrieval. The slides were incubated overnight with primary antibody (Nrf2, HO-1, c-Caspase-3 or bcl-2; 1:100 dilution) from ImmunoWay (USA) at 4°C, washed three times with PBS, and incubated with HRP-labeled secondary antibody for 30 min at 37°C. The slides were again rinsed three times for 5 min with PBS, and the sections were incubated with DAB. Finally, the slides were counterstained with haemotoxylin, mounted with DPX, and visualized under the microscope.
RNA isolation and analysis of Nrf2, HO-1 mRNA by real-time fluorescence quantitative PCR (RT-PCR)
Total RNA was extracted, and cDNA was synthesized according to the instructions of the manufacturer (TaKaRa, Japan). qPCR analysis was performed using a real-time PCR System (Rotor Gene 3000) with the following primers: Nrf2, F 5-ACACGGTCCACAGCTCATC-3, R 5-TGCCTCCAAGTATGTCAATA-3; HO-1, F 5-TCAGTCCCAAACGTCGCGGT-3, R 5-GCTGTGCAGGTGTTGAGCC-3; β-actin, F 5-GAAGTGTGACGTTGACATCCG-3, R 5-TGCTGATCCACATCTGCTGGA-3. Relative changes in expression were calculated using the 2-ΔΔCt method.
Immunoprecipitation (ChIP) assay
The ChIP assay was performed using a kit from Cell Signal Technology (CST, CA, USA). Briefly, intestinal tissues were lysed, and nuclei were pelleted and sheared using a sonicator with five 20-s pulses. Sheared chromatin was immunoprecipitated with 2 µg of anti-Nrf2 or control IgG antibody. The cross-links were reversed overnight at 65°C, and the chromatin was deprotonated with 20 µg/ml proteinase K. PCR amplification detected the Bcl-2 promoter region containing Nrf2/AREr3 binding sites were performed using the following primers was used for PCR: Bcl2-BS1, F 5-GTTCTTAAGCCCGATGTGGCAAC-3, R 5-GAGTAGTACCAATATGCTACCCTT-3; Bcl2-BS2, 5-ACCTTTCAGCATCACAGA-3, R 5-AATCACGCGGAACACTTG-3.
Statistical analyses
Statistical analyses were used to compare mean values from different groups by the homogeneity test. One-way analysis of variance (ANOVA) and Tukey’s multiple comparisons test were used to evaluate the significance of differences in other results by using Statistical Product and Service Solutions (SPSS) software package 17.0. P<0.05 was considered statistically significant.
Results
GLN reduces tissue injury in the jejunum after I/R
A sham operation was well-formed since villosity cell did not show any lysis or inflammatory process (Figure 1A). The I/R group displayed structural destruction of the villi (with only draft of one of them formed by inflammatory cells and necrotic material), hemorrhage and basal glandular ulceration (mainly for the intestinal mucosa villus epithelium), lamina propria disintegration, and some fluff top bleeding gland was significantly impaired (Figure 1B). The GLN group displayed expansion of the intestinal mucosa villi subcutaneous space but not obvious villus edema, generally normal glands, and lamina propria edema (Figure 1C). The 24 h reperfusion injury degree of comparison was: Sham group, 0.313±0.063 points; I/R group, 4.120±0.502 points; and GLN-I/R group: 2.461±0.372 points (Figure 1D). Comparing the I/R and the sham groups, the difference was statistically significant (P<0.001), as was the difference between the GLN-I/R group and the I/R group (P<0.05).
Figure 1.
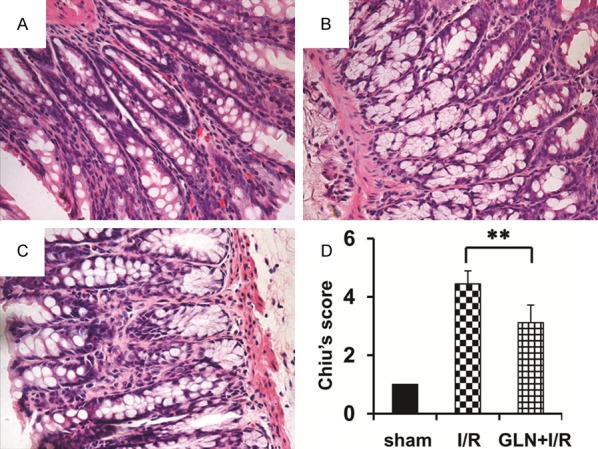
Histological characteristics of the of ileum segments in the I/R rat model. H&E staining showing normal histology of the ileum structure of rat in the sham group (A), showing the massive epithelial lifting, dilated capillaries, hemorrhage, and ulceration in the I/R group (B), and showing glutamine pretreatment results in significant structural improvement as evidenced by decreased edema and inflammatory cell infiltration in intestinal (C). Chiu’s score of comparison in these three groups (D). Magnification, 40×. **, P<0.01 vs. I/R group.
GLN protects rats against I/R-induced microvascular hyper-permeability
Glutamine pretreatment attenuated I/R-induced elevation of plasma endotoxin and D-lactic acid levels. These levels in the I/R group were significantly higher than in the Sham and GLN-I/R groups (both P<0.001). There was no significant difference between the sham group and the GLN-I/R groups (Figure 2), indicating glutamine successfully restore the microvascular hyper-permeability in ischemia/reperfusion process.
Figure 2.
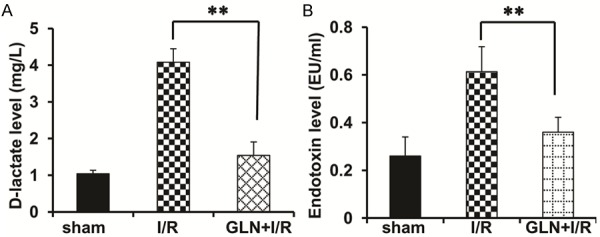
GLN protects rats against I/R-induced microvascular hyper-permeability. Shown as the D-lactic acid levels (A) and endotoxin levels (B) of the sham group, the I/R group and the GLN-I/R group. **P<0.01 vs. I/R group.
Effects of GLN in oxidative stress
Oxidative stress plays a significant role in the I/R and can be reflected by MDA, GSH-Px, and T-SOD levels. In this study, the serum concentrations of MDA and GSH-Px were significantly higher in the I/R group compared to the sham group (P<0.01, Figure 3A, 3B). By contrast, the MDA and GSH-Px concentrations in the GLN-I/R group were obviously lower than that in the I/R group (P<0.05, Figure 3A, 3B), suggesting that treatment with glutamine generated a low risk of lipid peroxidation. The increase in serum T-SOD activities after 30 min of ischemia and 24 h of reperfusion in rats pretreated with glutamine was significantly more pronounced than in I/R rats, thus confirming the anti-oxidative properties of glutamine in rats (P<0.01, Figure 3C).
Figure 3.
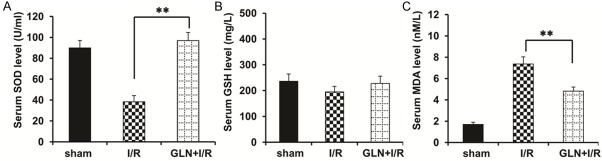
Effects of glutamine on oxidative stress in I/R rats. Shown as comparison of serum (A) T-SOD activities, (B) GSH-Px, (C) MDA levels and in the sham group, the I/R group and the GLN-I/R group. **P<0.01 vs. I/R group.
Effects of GLN on Nrf2 and HO-1 expressions
As shown in the Figure 4A, the expression of Nrf2 in the sham group showed light brown immunostaining in the cytoplasm and no staining in the nuclei. Significantly positive expression of Nrf2 in the cytoplasm and nuclei were observed in the I/R group. Compared to the I/R group, the positive rates of Nrf2 expression significantly increased in the GLN-I/R group. The expression of HO-1 in the sham group showed sparse brown immunostaining in the cytoplasm, but significant positive expression of HO-1 was observed in the cytoplasm of the I/R group . Compared to the I/R group, the positive rates of HO-1 expression increased significantly in the cytoplasm in the GLN-I/R group. The mRNA level and protein levels of the Nrf2 and Ho-1 were also validated by RT-PCR and western blot respectively, and the results were consistent with the IHC staining assay (Figure 4B, 4C).
Figure 4.
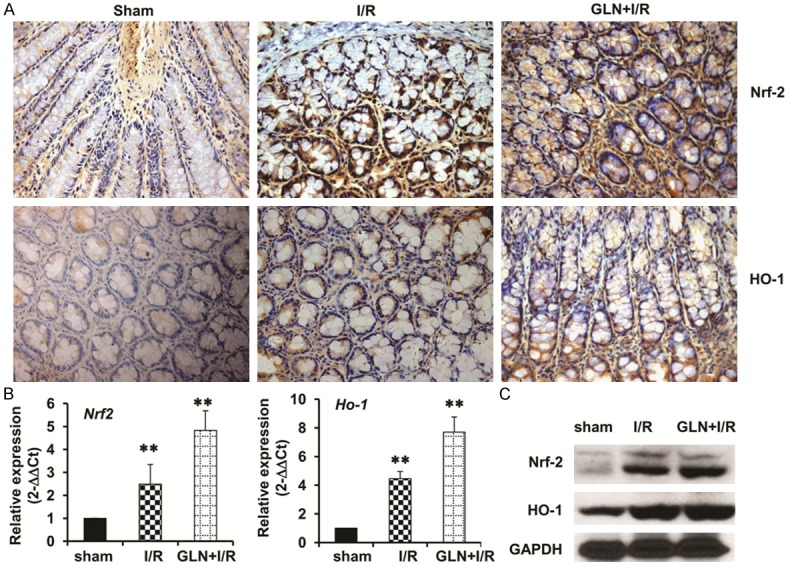
Effects of GLN on Nrf2 and HO-1 protein. (A) Immunohistochemical staining of intestinal Nrf2 expression (upper panel) and HO-1 expression (lower panel) in the sham group (left), the I/R group (middle), and the GLN-I/R group (right). (B) Real time PCR assay and (C) Western blotting assay showing the mRNA level and protein levels of Nrf2 and HO-1, respectively. GAPDH was used as the endogenous control. **, P<0.01. Magnification, 40×.
GLN promotes Bcl2 expression and inhibits intestinal apoptosis
Apoptosis is the major mechanism of cell death immediately following a short period of ischemia with ensuing reperfusion. In this study, intestinal apoptosis was evaluated by expressions of Bcl-2 and the cleaved caspase-3. The expression of Bcl-2 was elevated in the GLN-I/R group compared with the I/R group (Figure 5A upper panel, 5B). Compared with the I/R group, the cleaved caspase-3 protein significantly decreased in the GLN-I/R group (Figure 5A lower panel, 5C). These findings suggest that pretreatment with GLN can reduce intestinal apoptosis. Since GLN activates Nrf2/ARE signaling, we also evaluated whether GLN-induced Bcl2 expression was regualated by Nrf2 transcriptional factor in I/R process. We used immunoprecipitation assay to examine the binding of Nrf2 protein on two sites on Bcl2 promoter, and found that GLN treatment augmented the binding of Nrf2 on both sites (Figure 5D), indicating that GLN promotes Bcl2 expression through Nrf2/ARE signaling pathway.
Figure 5.
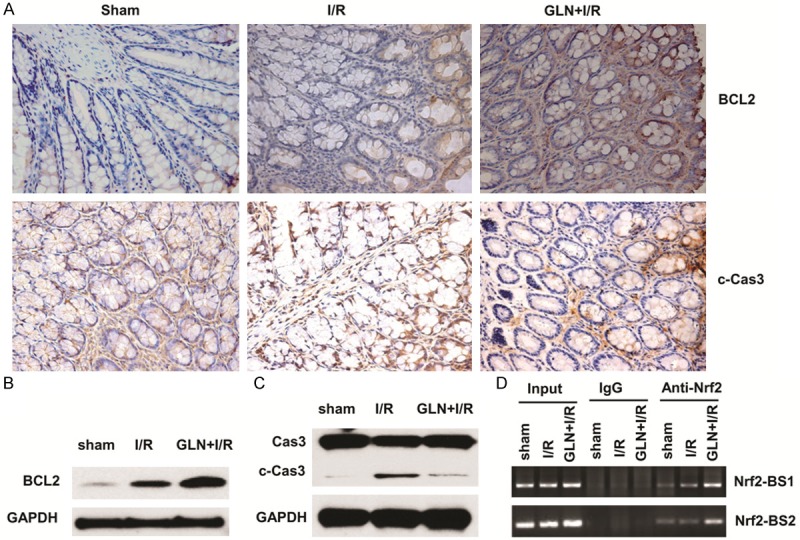
GLN inhibits intestinal cell apoptosis via upregulating Bcl2. (A) Immunohistochemical staining of intestinal Bcl2 expression (upper panel) and cleavage of Caspase 3 (lower panel) in the sham group (left), the I/R group (middle), and the GLN-I/R group (right). Western blotting assay showed the protein levels of Bcl2 (B), and cleavage of Caspase-3 in the sham group, the I/R group, and the GLN-I/R group (C), respectively. GAPDH was used as the endogenous control. (D) ChIP assay showing the augmentation binding of Nrf2/AREr3 onto two biding sites of Bcl2 promoter. Magnification, 40×.
Discussion
This study evaluated the protective effects of glutamine in a rat model of intestinal ischemia-reperfusion injury, and revealed that glutamine promotes Bcl2 expression through Nrf2/ARE signaling pathway as the molecular mechanism. To the best of our knowledge, our current study is the first one to disclose the function of glutamine against oxidative stress as well as the detailed molecular mechanism.
GLN is the most important and most available energy source in intestinal epithelial cells, lymphocytes, and macrophages [18-20], as well as a precursor of nucleotide synthesis and glutathione in the important antioxidant defense system. Luo et al show that GLN supplementation maintains gut glutathione levels during I/R [21]. Further, Wu and Ban’s studies demonstrate that GLN treatment has a protective effect during intestinal mucosal I/R injury, can reduce endotoxins elevation in I/R, and reduces bacterial translocation [22-24]. However, it is unclear whether GLN is protective for the intestinal mucosa against oxidative stress. Therefore, we established a rat I/R injury model to investigate the effects of GLN on I/R and its mechanism of action. Our results demonstrated that the GLN pretreated I/R rats had a greater villous height and a lower degree of intestinal permeability compared to the I/R rats. This suggests that GLN pretreatment plays an important role in the maintenance of the intestinal structure and barrier function in I/R-injured rats. Furthermore, we found that I/R leads to oxidative stress. Villi are rich in ROS-related enzymes, and I/R activates adenine-hypoxanthine metabolic pathways that produce abnormal increases in oxygen free radicals. Phospholipids are made of polyunsaturated fatty acids, which have a plurality of unsaturated bonds, and oxygen radicals have high affinity for these unsaturated bonds and can stimulate excessive lipid peroxidation chain reactions. This results in cell membrane lipid peroxidation, inhibition of mitochondrial activity, and increased membrane permeability of the cell and lysosome, leading to cell swelling/rupture. Our results indicate that GLN pretreatment inhibited lipid peroxidation, improved the body’s ability to scavenge oxygen free radicals, and thus, inhibited membrane disintegration. The potential protective mechanisms may include upregulation of endogenous antioxidant factors (i.e., SOD, Nrf2, and HO-1) and downregulation of MDA levels by the Nrf2/ARE signaling pathway.
In our I/R injury model, we also observed an interesting result in that GLN exerted potent anti-apoptotic effects, as inferred by the anti-apoptotic marker Bcl-2. Intestinal apoptosis (programmed cell death) is a rare event in healthy intestinal cells but is the early and predominant form of cell death in intestinal necrosis and has been linked to I/R [25]. The concentrations of Bcl-2 have an important role in the protection or acceleration of intestinal apoptosis after ischemia and/or reperfusion [26]. When the Bcl-2 levels in a cell decrease, the mPTP can be opened, resulting in a caspase cascade and ultimately leading to cell apoptosis. In contrast, increased Bcl-2 can inhibit apoptosis [27]. A recent publications has revealed that Nrf2/AREr3 signaling regulates Bcl2 gene expression by directly binding to the promoter regions of Bcl2 gene and prevents human hepatoblastoma cell apoptosis [28]. In our study, we also confirmed that GLN treatment increased Nrf2 level and promoted Nrf2/AREr3 binding to the promoter of Bcl2 gene, therefore promotes Bcl2 gene expression in the I/R process.
In conclusion, our results demonstrate that GLN pretreatment protects the rat intestine from morphologic and functional mucosal injury, as well as increases Nrf2 and HO-1 protein levels after I/R. The protective effect may be achieved by scavenging oxygen free radicals and inhibiting the ROS/pASK1/pJNK/Bcl-2/Caspase/Apoptosis lipid peroxidation. These results suggest that GLN would be clinically useful in the treatment of I/R injury.
Acknowledgements
This work was supported by the Shandong Province Natural Science Foundation of China (Grants ZR2010hl051). The authors gratefully acknowledge Cui-ping Mao for her technical assistance during the course of the study and Yan-yan Wang for her help in the surgery.
Disclosure of conflict of interest
None.
References
- 1.Lee MA, Yatani A, Sambol JT, Deitch EA. Role of gut-lymph factors in the induction of burn-induced and trauma-shock-induced acute heart failure. Int J Clin Exp Med. 2008;1:171–180. [PMC free article] [PubMed] [Google Scholar]
- 2.Hassoun HT, Kone BC, Mercer DW, Moody FG, Weisbrodt NW, Moore FA. Post-injury multiple organ failure: the role of the gut. Shock. 2001;15:1–10. doi: 10.1097/00024382-200115010-00001. [DOI] [PubMed] [Google Scholar]
- 3.Young CM, Kingma SD, Neu J. Ischemia-reperfusion and neonatal intestinal injury. J Pediatr. 2011;158:e25–28. doi: 10.1016/j.jpeds.2010.11.009. [DOI] [PubMed] [Google Scholar]
- 4.Fishman JE, Levy G, Alli V, Sheth S, Lu Q, Deitch EA. Oxidative modification of the intestinal mucus layer is a critical but unrecognized component of trauma hemorrhagic shock-induced gut barrier failure. Am J Physiol Gastrointest Liver Physiol. 2013;304:G57–63. doi: 10.1152/ajpgi.00170.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Yang Y, Li W, Sun Y, Han F, Hu CA, Wu Z. Amino acid deprivation disrupts barrier function and induces protective autophagy in intestinal porcine epithelial cells. Amino Acids. 2014 doi: 10.1007/s00726-014-1844-6. [Epub ahead of print] [DOI] [PubMed] [Google Scholar]
- 6.Bhattacharyya A, Chattopadhyay R, Mitra S, Crowe SE. Oxidative stress: an essential factor in the pathogenesis of gastrointestinal mucosal diseases. Physiol Rev. 2014;94:329–354. doi: 10.1152/physrev.00040.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Guo H, Li MJ, Liu QQ, Guo LL, Ma MM, Wang SX, Yu B, Hu LM. Danhong injection attenuates ischemia/reperfusion-induced brain damage which is associating with Nrf2 levels in vivo and in vitro. Neurochem Res. 2014;39:1817–1824. doi: 10.1007/s11064-014-1384-1. [DOI] [PubMed] [Google Scholar]
- 8.Han J, Wang M, Jing X, Shi H, Ren M, Lou H. (-)-Epigallocatechin gallate protects against cerebral ischemia-induced oxidative stress via Nrf2/ARE signaling. Neurochem Res. 2014;39:1292–1299. doi: 10.1007/s11064-014-1311-5. [DOI] [PubMed] [Google Scholar]
- 9.Li L, Dong H, Song E, Xu X, Liu L, Song Y. Nrf2/ARE pathway activation, HO-1 and NQO1 induction by polychlorinated biphenyl quinone is associated with reactive oxygen species and PI3K/AKT signaling. Chem Biol Interact. 2014;209:56–67. doi: 10.1016/j.cbi.2013.12.005. [DOI] [PubMed] [Google Scholar]
- 10.Satoh T, McKercher SR, Lipton SA. Reprint of: Nrf2/ARE-mediated antioxidant actions of pro-electrophilic drugs. Free Radic Biol Med. 2014;66:45–57. doi: 10.1016/j.freeradbiomed.2013.11.002. [DOI] [PubMed] [Google Scholar]
- 11.Alves WF, Aguiar EE, Guimaraes SB, da Silva Filho AR, Pinheiro PM, Soares Gdos S, de Vasconcelos PR. L-alanyl-glutamine preoperative infusion in patients with critical limb ischemia subjected to distal revascularization reduces tissue damage and protects from oxidative stress. Ann Vasc Surg. 2010;24:461–467. doi: 10.1016/j.avsg.2010.01.005. [DOI] [PubMed] [Google Scholar]
- 12.Fukatsu K, Ueno C, Hashiguchi Y, Hara E, Kinoshita M, Mochizuki H, Hiraide H. Glutamine infusion during ischemia is detrimental in a murine gut ischemia/reperfusion model. JPEN J Parenter Enteral Nutr. 2003;27:187–192. doi: 10.1177/0148607103027003187. discussion 192. [DOI] [PubMed] [Google Scholar]
- 13.Zhang Y, Lv Y, Liu YJ, Yang C, Hu HJ, Meng XE, Li MX, Pan SY. Hyperbaric oxygen therapy in rats attenuates ischemia-reperfusion testicular injury through blockade of oxidative stress, suppression of inflammation, and reduction of nitric oxide formation. Urology. 2013;82:489.e9–489.e15. doi: 10.1016/j.urology.2013.04.016. [DOI] [PubMed] [Google Scholar]
- 14.Ikeda S, Zarzaur BL, Johnson CD, Fukatsu K, Kudsk KA. Total parenteral nutrition supplementation with glutamine improves survival after gut ischemia/reperfusion. JPEN J Parenter Enteral Nutr. 2002;26:169–173. doi: 10.1177/0148607102026003169. [DOI] [PubMed] [Google Scholar]
- 15.Salomao AB, Aguilar-Nascimento JE, Percario S, Sano V, Marques NR, Dias CC. Intestinal intraluminal injection of glutamine increases trolox total equivalent antioxidant capacity (TEAC) in hepatic ischemia-reperfusion. Acta Cir Bras. 2006;21(Suppl 4):69–73. doi: 10.1590/s0102-86502006001000015. [DOI] [PubMed] [Google Scholar]
- 16.Lindestrom LM, Ekblad E. Structural and neuronal changes in rat ileum after ischemia with reperfusion. Dig Dis Sci. 2004;49:1212–1222. doi: 10.1023/b:ddas.0000037815.63547.08. [DOI] [PubMed] [Google Scholar]
- 17.Chiu CJ, McArdle AH, Brown R, Scott HJ, Gurd FN. Intestinal mucosal lesion in low-flow states. I. A morphological, hemodynamic, and metabolic reappraisal. Arch Surg. 1970;101:478–483. doi: 10.1001/archsurg.1970.01340280030009. [DOI] [PubMed] [Google Scholar]
- 18.Zuhl MN, Lanphere KR, Kravitz L, Mermier CM, Schneider S, Dokladny K, Moseley PL. Effects of oral glutamine supplementation on exercise-induced gastrointestinal permeability and tight junction protein expression. J Appl Physiol (1985) 2014;116:183–191. doi: 10.1152/japplphysiol.00646.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Han T, Li XL, Cai DL, Zhong Y, Geng SS. Effects of glutamine-supplemented enteral or parenteral nutrition on apoptosis of intestinal mucosal cells in rats with severe acute pancreatitis. Eur Rev Med Pharmacol Sci. 2013;17:1529–1535. [PubMed] [Google Scholar]
- 20.Kaufmann Y, Spring P, Klimberg VS. Oral glutamine prevents DMBA-induced mammary carcinogenesis via upregulation of glutathione production. Nutrition. 2008;24:462–469. doi: 10.1016/j.nut.2008.01.003. [DOI] [PubMed] [Google Scholar]
- 21.Luo M, Bazargan N, Griffith DP, Estivariz CF, Leader LM, Easley KA, Daignault NM, Hao L, Meddings JB, Galloway JR, Blumberg JB, Jones DP, Ziegler TR. Metabolic effects of enteral versus parenteral alanyl-glutamine dipeptide administration in critically ill patients receiving enteral feeding: a pilot study. Clin Nutr. 2008;27:297–306. doi: 10.1016/j.clnu.2007.12.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Wu GH, Wang H, Zhang YW, Wu ZH, Wu ZG. Glutamine supplemented parenteral nutrition prevents intestinal ischemia- reperfusion injury in rats. World J Gastroenterol. 2004;10:2592–2594. doi: 10.3748/wjg.v10.i17.2592. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Ban K, Sprunt JM, Martin S, Yang P, Kozar RA. Glutamine activates peroxisome proliferator-activated receptor-gamma in intestinal epithelial cells via 15-S-HETE and 13-OXO-ODE: a novel mechanism. Am J Physiol Gastrointest Liver Physiol. 2011;301:G547–554. doi: 10.1152/ajpgi.00174.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Ban K, Kozar RA. Enteral glutamine: a novel mediator of PPARgamma in the postischemic gut. J Leukoc Biol. 2008;84:595–599. doi: 10.1189/jlb.1107764. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Li JL, Wang QY, Luan HY, Kang ZC, Wang CB. Effects of L-carnitine against oxidative stress in human hepatocytes: involvement of peroxisome proliferator-activated receptor alpha. J Biomed Sci. 2012;19:32. doi: 10.1186/1423-0127-19-32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Ge M, Chi X, Zhang A, Luo G, Sun G, Xie H, Hei Z. Intestinal NF-E2-related factor-2 expression and antioxidant activity changes in rats undergoing orthotopic liver autotransplantation. Oncol Lett. 2013;6:1307–1312. doi: 10.3892/ol.2013.1576. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Ola MS, Nawaz M, Ahsan H. Role of Bcl-2 family proteins and caspases in the regulation of apoptosis. Mol Cell Biochem. 2011;351:41–58. doi: 10.1007/s11010-010-0709-x. [DOI] [PubMed] [Google Scholar]
- 28.Niture SK, Jaiswal AK. Nrf2 protein up-regulates antiapoptotic protein Bcl-2 and prevents cellular apoptosis. J Biol Chem. 2012;287:9873–9886. doi: 10.1074/jbc.M111.312694. [DOI] [PMC free article] [PubMed] [Google Scholar]