

Abstract
Flavonoids, such as fisetin (3,7,3′,4′-tetrahydroxyflavone), are plant secondary metabolites. It has been reported that fisetin is able to perform numerous pharmacological roles including anti-inflammatory, anti-microbial, and anti-cancer activities; however, the exact anti-inflammatory mechanism of fisetin is not understood. In this study, the pharmacological action modes of fisetin in lipopolysaccharide (LPS)-stimulated macrophage-like cells were elucidated by using immunoblotting analysis, kinase assays, and an overexpression strategy. Fisetin diminished the release of nitric oxide (NO) and reduced the mRNA levels of inducible NO synthase (iNOS), tumor necrosis factor (TNF)-α, and cyclooxygenase (COX)-2 in LPS-stimulated RAW264.7 cells without displaying cytotoxicity. This compound also blocked the nuclear translocation of p65/nuclear factor (NF)-κB. In agreement, the upstream phosphorylation events for NF-κB activation, composed of Src, Syk, and IκBα, were also reduced by fisetin. The phospho-Src level, triggered by overexpression of wild-type Src, was also inhibited by fisetin. Therefore, these results strongly suggest that fisetin can be considered a bioactive immunomodulatory compound with anti-inflammatory properties through suppression of Src and Syk activities.
Keywords: Fisetin, Anti-inflammatory effect, NF-κB, Src, Syk
INTRODUCTION
Inflammation is one of the common biological reactions that protect our body from infection of bacteria, virus, and fungi. When inflammation occurs, the body responds by activating many different types of immune cells to clear out pathogens. Macrophages, the most common inflammatory cells, play a critical role in removing infectious materials and activate other immune cells. For these processes, activated macrophages produce pro-inflammatory cytokines and inflammatory mediators such as nitric oxide (NO) (Yang et al., 2014a; Soler Palacios et al., 2015). New production of these molecules is regulated at the transcriptional level by controlling nuclear translocation and DNA binding activity of inflammation-regulatory transcription factors such as nuclear factor (NF-κB) and activator protein (AP-1). By the action of these proteins, mRNA levels of inflammatory genes, such as inducible NO synthase (iNOS), cyclooxygenase (COX)-2, and tumor necrosis factor (TNF)-α can be dramatically increased (Van Den Berg and Bresnihan, 1999; Vilahur and Badimon, 2014).
Flavonoids are plant pigments that possess various physiological effects including antioxidant, antiviral, anti-cancer, anti-bacterial and anti-inflammatory activities (Rice-Evans et al., 1996; Jang et al., 2005; Khan et al., 2013; Jeong et al., 2014b). Fisetin (Fig. 1) is one of the polyphenolic flavonoids, which are widely found in many fruits and vegetables such as strawberry, mango, and onion (Khan et al., 2013; Pal et al., 2015). It has also been reported that fisetin displays anti-oxidative, anti-inflammatory, and anti-proliferative activities (Khan et al., 2013). In particular, several groups have suggested the anti-inflammatory role of fisetin occurs via suppression of NF-κB and AP-1 activation (Kim et al., 2012), however, how fisetin blocks these pathways is not fully elucidated in terms of identifying direct target enzymes. In this study, therefore, we sought to identify the molecular targets of fisetin involved in the negative regulation of lipopolysaccharide (LPS)-stimulated macrophages.
Fig. 1.
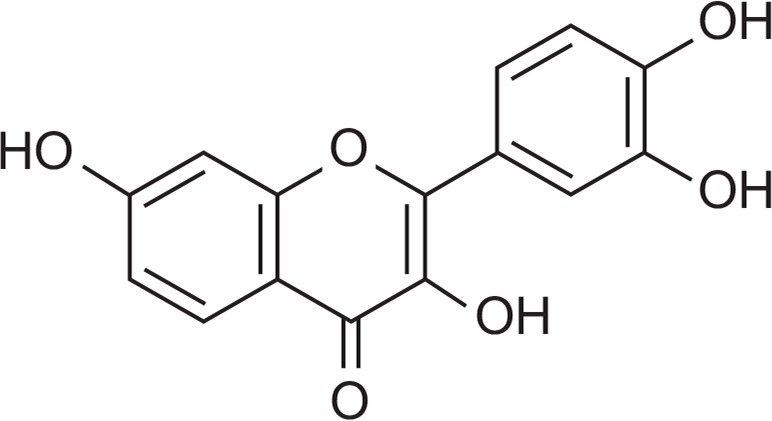
Chemical structure of fisetin.
MATERIALS AND METHODS
Materials
Fisetin (purity: >98%), polyethylenimine (PEI), 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (a tetrazole) (MTT), and LPS (E. coli 0111:B4) were purchased from Sigma Chemical Co. (St. Louis, MO, USA). Fetal bovine serum (FBS) and RPMI1640 were obtained from GIBCO (Grand Island, NY, USA). Dulbecco’s modified Eagle’s medium (DMEM) was obtained from Thermo Fisher Scientific Inc. (Waltham, MA, USA). RAW264.7 (a mouse macrophage-like cell line) and HEK293 (a human embryonic kidney cell line) cells were purchased from ATCC (Rockville, MD, USA). All other chemicals used in this study were of analytical grade from Sigma Chemical Company. Phospho-specific or total-protein antibodies recognizing p65, p50, inhibitor of κBα (IκBα), Src, spleen tyrosine kinase (Syk), lamin A/C and β-actin were obtained from Cell Signaling Technology (Beverly, MA, USA).
Expression vectors and DNA transfection
Wild-type Src (Src-WT) was used as reported previously (Yang et al., 2008). All constructs were confirmed by automated DNA sequencing. Overexpression experiments were performed with HEK293 cells (1×106 cells/mL) by transfection with Src-WT using the PEI method in 12-well plates as reported previously (Shen et al., 2011; Song et al., 2012). The cells were utilized for the experiments 24 h post-transfection. Fisetin was added to cells 24 h before termination.
Cell culture and drug preparation
RAW264.7 cells were cultured in RPMI1640 with 10% heat-inactivated FBS and 1% penicillin/streptomycin at 37°C in 5% CO2. HEK293 cells were maintained in DMEM media supplemented with 5% heat-inactivated FBS and 1% penicillin/streptomycin at 37°C in 5% CO2. The stock solutions of fisetin for the in vitro experiments were prepared in dimethylsulfoxide (DMSO).
Determination of NO production
After pre-incubation of RAW264.7 cells (1×106 cells/mL) for 18 h, fisetin was added to the cells for 30 min. After that, the cells were treated with LPS (1 μg/mL) for 24 h. The effect of fisetin (0 to 30 μM) on the production of NO was determined by analyzing NO levels using Griess reagents (Kim et al., 2013a; Youn et al., 2013).
Cell viability test
The cytotoxic effects of fisetin (0 to 30 μM) were then evaluated using a conventional MTT assay as previously reported (Pauwels et al., 1988; Yayeh et al., 2012; Oh et al., 2013). For the final 3 h of culture, 10 μl of MTT solution (10 mg/mL in phosphate-buffered saline, pH 7.4) were added to each well. Reactions were stopped by the addition of 15% sodium dodecyl sulfate (SDS) into each well, solubilizing the formazan. The absorbance at 570 nm (OD570–630) was measured using a Spectramax 250 microplate reader (BioTex, Bad Friedrich-shall, Germany).
mRNA analysis using semi-quantitative and quantitative reverse transcriptase-polymerase chain reactions
In order to determine cytokine mRNA expression levels, total RNA was isolated from LPS-treated RAW264.7 cells using TRIzol Reagent, according to the manufacturer’s instructions. Total RNA was stored at −70°C until use. Semi-quantitative RT reactions were conducted as previously reported (Lee et al., 2009). Quantification of mRNA was performed by real-time reverse transcriptase polymerase chain reaction (RT-PCR) with SYBR Premix Ex Taq according to the manufacturer’s instructions (Takara, Shiga, Japan) using a real-time thermal cycler (Bio-Rad, Hercules, CA, USA) as reported previously (Kang et al., 2013). Semi-quantitative RT-PCR was conducted as previously reported with minor modifications (Sohn et al., 2013). All of the primers (Bioneer, Daejeon, Korea) used are listed in Table 1.
Table 1.
PCR primers used in this study
| Name | Sequence (5′ to 3′) | |
|---|---|---|
| iNOS | F | GGAGCCTTTAGACCTCAACAGA |
| R | TGAACGAGGAGGGTGGTG | |
| TNF-α | F | TGCCTATGTCTCAGCCTCTTC |
| R | GAGGCCATTTGGGAACTTCT | |
| COX-2 | F | GGGAGTCTGGAACATTGTGAA |
| R | GCACATTGTAAGTAGGTGGACTGT | |
| GAPDH | F | CAATGAATACGGCTACAGCAAC |
| R | AGGGAGATGCTCAGTGTTGG |
Preparation of cell lysates and nuclear fractions for immunoblotting
RAW264.7 or HEK293 cells (5×106 cells/mL) were washed three times in cold phosphate buffered saline (PBS) and lysed in lysis buffer as reported previously (Kim et al., 2014b). Nuclear lysates were prepared using a three-step procedure (Byeon et al., 2008). After treatment, the cells were collected with a rubber policeman, washed with 1×PBS, and lysed in 500 μl of lysis buffer on ice for 4 min. During the second step, the pellet (the nuclear fraction) was washed once with wash buffer without Nonidet P-40. During the final step, the nuclei were resuspended in an extraction buffer consisting of the lysis buffer plus 500 mM KCl and 10% glycerol. The nuclei/extraction buffer mixture was frozen at −80°C then thawed on ice and centrifuged at 14,000 rpm for 5 min. The supernatant was collected as the nuclear extract. Whole-cell or nuclear lysates were then analyzed by a conventional immunoblotting method (Yang et al., 2014b). The total and phosphorylated levels of p65, p50, IκBα, Src, Syk, HA, lamin A/C, and β-actin were visualized using an ECL system (Amersham, Little Chalfont, Buckinghamshire, UK) as reported previously (Lee et al., 2012).
In vitro kinase assay with purified enzymes
In order to evaluate the inhibition of the kinase activities of Src or Syk using purified enzymes, the kinase profiler service from Millipore (Billerica, MA, USA) was used. Purified Src or Syk (human) (1–5 mU) were incubated with the reaction buffer in a final reaction volume of 25 μl. The reaction was initiated by the addition of Mg-ATP. After incubation for 40 min at room temperature, the reaction was stopped by the addition of 5 mL of a 3% phosphoric acid solution. Ten microliters of the reaction were then spotted onto a P30 Filtermat that was washed three times for 5 min in 75 mM phosphoric acid and once in methanol prior to drying and scintillation counting.
Statistical analyses
All of the data presented in this paper are expressed as means ± SD. For statistical comparisons, results were analyzed using either ANOVA/Scheffe’s post-hoc test or the Kruskal-Wallis/Mann-Whitney test. A p-value<0.05 was considered to be statistically significant. All statistical tests were carried out using the computer program, SPSS (SPSS Inc., Chicago, IL, USA).
RESULTS
Effect of fisetin on inflammatory responses
First, to confirm the function of fisetin as an inflammatory-response suppressor in macrophage cells, we tested its capability to suppress NO production in LPS-treated RAW264.7 cells. As expected, production of NO was dose-dependently decreased by fisetin in RAW264.7 cells stimulated by LPS (Fig. 2A). It was found that there was no cytotoxic activity of fisetin at its effective anti-inflammatory concentrations (Fig. 2B).
Fig. 2.

The effect of fisetin on the production of NO and cell viability in LPS-stimulated RAW 264.7 cells. (A) RAW264.7 cells (1×106 cells/mL) were treated with LPS (1 μg/mL) in the presence or absence of fisetin (0 to 30 μM) for 24 h. The supernatants were then collected, and the NO concentration in the supernatants was determined using the Griess assay. (B) RAW264.7 cells (1×106 cells/mL) were treated with fisetin for 24 h, and cell viability was evaluated using the MTT assay. All data are expressed as the mean ± SD of experiments, which were performed with six samples. **p<0.01 compared to normal or control groups.
Effect of fisetin on transcriptional activation in LPS-treated RAW264.7 cells
To check whether the anti-inflammatory effect of fisetin occurs at the transcriptional level, we determined the mRNA expression levels of inflammatory genes such as inducible nitric oxide synthase (iNOS), cyclooxygenase-2 (COX-2), and tumor necrosis factor-α (TNF-α). As expected, the expression of inflammatory mediator genes was suppressed in cells treated with either 20 or 30 μM of fisetin according to both semi-quantitative (Fig. 3A) and real-time RT-PCR (Fig. 3B) analyses. We also examined transcription factor levels in nuclear extracts by immunoblotting analysis and found that fisetin treatment was capable of inhibiting the nuclear translocation of p65/NF-κB at 60 min but not of p50/NF-κB (Fig. 3C).
Fig. 3.

The effect of fisetin on transcriptional activation in LPS-stimulated RAW264.7 cells. (A and B) The mRNA levels of iNOS, COX-2, and TNF-α expressed in LPS (1 μg/mL)-treated RAW264.7 cells in the presence or absence of fisetin (20 and 30 μM) were measured by RT-PCR (A) or real-time PCR (B). (C) The nuclear levels of p65 and p50 in RAW264.7 cells treated with LPS (1 μg/mL) in the presence or absence of fisetin (30 VM) were analyzed by immunoblotting analysis. Data (B) are expressed as the mean ± SD of experiments, which were performed with six samples. **p<0.01 compared to normal or control groups.
Effect of fisetin on the upstream signaling of NF-κB activation
Many studies have demonstrated that the NF-κB pathway is regulated by intracellular signaling cascades (Byeon et al., 2012; Yi et al., 2014). These signaling cascades include Src, Syk, and IκBα. To identify fisetin-targeted molecules in this pathway, we confirmed the expression levels of these molecules by immunoblotting analysis. Expectedly, the phospho-IκBα level in fisetin-treated cells was clearly decreased at 5, 30, and 60 min (Fig. 4A). Also, the autophosphorylation levels of Src and Syk were strikingly suppressed by fisetin at 2 min, without decreasing total levels of Src and Syk (Fig. 4B).
Fig. 4.

The effect of fisetin on the upstream signaling cascade of NF-κB. (A and B) RAW264.7 cells were incubated with LPS (1 μg/mL) in the presence or absence of fisetin (30 μM) for 30 min. The total phospho-protein levels of Src, Syk, and IκBα from whole lysates were determined by immunoblotting analysis.
Inhibition of Src and Syk kinase activity and overexpression of Src by fisetin
In order to confirm inhibitory activity of Src/Syk phosphorylation, we checked whether these enzymes can directly block the kinase activity of Src and Syk. As expected, the kinase activities of purified Src and Syk were completely inhibited by fisetin (30 μM). This result indicates that fisetin can act as a direct inhibitor of these enzymes (Fig. 5A). To further validate fisetin-mediated suppression of Src activity, we employed an overexpression strategy using HA-Src. We found that over-expressed Src increased the phospho-Src level and fisetin suppressed the phosphorylation of Src in a dose-dependent manner (Fig. 5B).
Fig. 5.

The effect of fisetin on the activity of Src and Syk. (A) Inhibitory activity of fisetin (30 μM) on the kinase activity of purified Src or Syk was examined by kinase assay. (B) Inhibitory activity of fisetin (20 and 30 μM) on the phosphorylation of overexpressed Src was determined by immunoblotting analysis. (C) RAW264.7 cells (1×106 cells/mL) were treated with LPS (1 μg/mL) in the presence or absence of PP2 (10 and 20 μM) for 24 h. The supernatants were then collected, and the NO concentration in the supernatants was determined using the Griess assay. Data (A and C) are expressed as the mean ± SD of experiments, which were performed with six samples. *p<0.05 and **p<0.01 compared to normal or control groups.
DISCUSSION
It was reported that fisetin exhibits anti-inflammatory, anti-oxidative, and anti-proliferative activities (Pal et al., 2015). In our study, this compound was revealed to significantly inhibit NO production in RAW264.7 cells without affecting their cell viability (Fig. 2A, 2B). In addition, mRNA expression levels of pro-inflammatory genes were decreased by fisetin (Fig. 3A, 3B). Additionally, the translocation of transcription factor p65, a subunit of NF-κB, into the nucleus was remarkably inhibited at 60 min (Fig. 3C), implying that NF-κB upstream signaling pathway may be targeted by fisetin treatment. Indeed, the phosphorylation of NF-κB regulatory upstream protein (IκBα), a key event for p65 translocation (Courtois and Gilmore, 2006), was decreased by fisetin (Fig. 4A). Similarly, it was also reported that fisentin exterts downregulation of interleukin-6 and TNF-α in LPS-activated RAW264.7 cells via suppression of NF-κB, although exact target protein was not fully identified (Kim et al., 2012). Previous papers and our present results led us to hypothesize that early enzymes, activated within 5 min under LPS stimulation, could be regulated by fisetin.
Previously, we determined that Src and Syk are major NF-κB regulatory protein tyrosine kinases (Byeon et al., 2012; Yi et al., 2014). In parallel, inhibitory compounds, such as carnosic acid and quercetin, and plant-derived extracts from Evodia lepta, Artemisia asiatica, and Rhodomyrtus tomentosa, which show anti-inflammatory activities, were found to suppress Src and Syk (Oh et al., 2012; Endale et al., 2013; Jeong et al., 2013; Yoon et al., 2013). These enzymes’ active forms and pro-inflammatory cytokines were increased in response to LPS stimulation at early time points (Lee et al., 2009). In particular, activated Src and Syk are known to be involved in regulation of IκBα-phosphorylation (Lee et al., 2009). Therefore, we also tested whether fisetin is able to suppress the phosphorylation of Src and Syk, a hallmark to determine activated forms of Src and Syk (Byeon et al., 2012; Yi et al., 2014). As expected, the phosphorylation of Src and Syk at 2 min was strongly suppressed by fisetin (Fig. 4B). Since the phosphorylation of these enzymes is generated by their catalytic enzyme activities (Byeon et al., 2012; Yi et al., 2014), it is supposed that fisetin can directly diminish the enzyme activity of Src and Syk. To prove this possibility, we determined the kinase activity by a conventional enzyme assay with purified Src and Syk as reported previously (Kim et al., 2013b; Jeong et al., 2014b). As Fig. 5A shows, the kinase activity of Src was strongly reduced by fisetin (30 μM), while Syk was suppressed up to 52%, indicating that Src is the more preferred target of fisetin. Validation of this was also performed by overexpression work with HA-Src-expressing construct and by exploration of NO inhibitory activity under the treatment of PP2, a Src inhibitor (Byeon et al., 2013). In fact, it was confirmed that transfection of Src cDNA increased the phosphorylation level of Src (Byeon et al., 2013; Dung et al., 2014). Similarly, fisetin treatment dose-dependently inhibited the phosphorylation of Src raised by Src overexpression (Fig. 5B). PP2 also strongly blocked the release of NO in LPS-treated RAW264.7 cells (Fig. 5C) as reported previously (Yang et al., 2014a). Therefore, these results, and other previously reported results, strongly support that Src could be a major direct target of fisetin in its anti-inflammatory actions.
Our data, demonstrating fisetin’s inhibitory potency and its anti-inflammatory mechanisms, strongly suggest that it can be developed as an anti-inflammatory drug. However, development of a single compound from natural plants as an anti-inflammatory drug is difficult due to high production cost and low purification yield. Although the amount of this compound is very low, it has been demonstrated that fisetin is one of the major anti-inflammatory components in various plants, such as Muntingia calabura, Toxicodendron vernicifluum, and Rhus verniciflua, with in vitro and in vivo anti-inflammatory activity (Park et al., 2013; Kim et al., 2015). A recent trend in the pharmaceutical industry is discovering strategies to develop the natural plant itself as the new drug. Indeed, STILLENTM from Dong-A Pharmaceutical Co. in Korea, was developed from Artemisiae argyi Folium 14 years ago. The activity of this compound against gastric ulcer and gastritis is promising, even in the extract (Jeong et al., 2014a). The active components of this extract are flavonoids eupatilin, jaceosidin, and luteolin (Seo et al., 2003). Considering these, it is also proposed that fisetin-containing plants can be chosen as anti-inflammatory candidate herbal medicines. Numerous reports have proved that fisetin-containing Rhus verniciflua is orally effective against 2,4-dinitrofluorobenzene-induced allergic contact dermatitis and carrageenan-induced edema (Jung et al., 2011; Park et al., 2013; Kim et al., 2014a). A chemical synthetic approach for mass production of fisetin and its derivatives was also reported previously (Chiruta et al., 2012). Using this method, it is expected that fisetin can be prepared in a low-cost way. Although synthetic fisetin can be applied in industrial fields, extract formulas seem to be easier to develop and display more potent efficacy due to a mixture form with other active ingredients. Therefore, we will continue screening fisetin-rich plants by HPLC analysis and will prove their anti-inflammatory efficacy by using in vitro and in vivo experimental models.
In summary, we have demonstrated that fisetin can suppress IκBα phosphorylation, p65 translocation, inflammatory gene expression, and NO production by direct blockade of Src and Syk kinase activities as summarized in Fig. 6. Because fisetin shows potent anti-inflammatory activity, we suggest that fisetin-rich plants from edible sources could be applied to the development of therapeutic remedies to cure various inflammatory symptoms. Therefore, relevant works will be further proved in the future projects.
Fig. 6.

Putative inhibitory pathway of fisetin in LPS-activated inflammatory events.
Acknowledgments
This work was carried out with the support of the Cooperative Research Program for Agriculture Science & Technology Development (Project no. PJ009241), Rural Development Administration, Korea.
REFERENCES
- Byeon SE, Lee YG, Kim BH, Shen T, Lee SY, Park HJ, Park SC, Rhee MH, Cho JY. Surfactin blocks NO production in lipopolysaccharide-activated macrophages by inhibiting NF-kappaB activation. J Microbiol Biotechnol. 2008;18:1984–1989. [PubMed] [Google Scholar]
- Byeon SE, Yi YS, Oh J, Yoo BC, Hong S, Cho JY. The role of Src kinase in macrophage-mediated inflammatory responses. Mediators Inflamm. 2012;2012:512926. doi: 10.1155/2012/512926. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Byeon SE, Yu T, Yang Y, Lee YG, Kim JH, Oh J, Jeong HY, Hong S, Yoo BC, Cho WJ, Hong S, Cho JY. Hydroquinone regulates hemeoxygenase-1 expression via modulation of Src kinase activity through thiolation of cysteine residues. Free Radic Biol Med. 2013;57:105–118. doi: 10.1016/j.freeradbiomed.2012.12.013. [DOI] [PubMed] [Google Scholar]
- Chiruta C, Schubert D, Dargusch R, Maher P. Chemical modification of the multitarget neuroprotective compound fisetin. J Med Chem. 2012;55:378–389. doi: 10.1021/jm2012563. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Courtois G, Gilmore TD. Mutations in the NF-kappaB signaling pathway: implications for human disease. Oncogene. 2006;25:6831–6843. doi: 10.1038/sj.onc.1209939. [DOI] [PubMed] [Google Scholar]
- Dung TT, Kim SC, Yoo BC, Sung GH, Yang WS, Kim HG, Park JG, Rhee MH, Park KW, Yoon K, Lee Y, Hong S, Kim JH, Cho JY. (5-Hydroxy-4-oxo-4H-pyran-2-yl)methyl 6-hydroxynaphthalene-2-carboxylate, a kojic acid derivative, inhibits inflammatory mediator production via the suppression of Syk/Src and NF-kappaB activation. Int Immunopharmacol. 2014;20:37–45. doi: 10.1016/j.intimp.2014.02.019. [DOI] [PubMed] [Google Scholar]
- Endale M, Park SC, Kim S, Kim SH, Yang Y, Cho JY, Rhee MH. Quercetin disrupts tyrosine-phosphorylated phosphatidylinositol 3-kinase and myeloid differentiation factor-88 association, and inhibits MAPK/AP-1 and IKK/NF-kappaB-induced inflammatory mediators production in RAW 264.7 cells. Immunobiology. 2013;218:1452–1467. doi: 10.1016/j.imbio.2013.04.019. [DOI] [PubMed] [Google Scholar]
- Jang H-S, Kook S-H, Son Y-O, Kim J-G, Jeon Y-M, Jang Y-S, Choi K-C, Kim J, Han S-K, Lee K-Y. Flavonoids purified from Rhus verniciflua Stokes actively inhibit cell growth and induce apoptosis in human osteosarcoma cells. Biochim. Biophys. Acta. 2005;1726:309–316. doi: 10.1016/j.bbagen.2005.08.010. [DOI] [PubMed] [Google Scholar]
- Jeong D, Yang WS, Yang Y, Nam G, Kim JH, Yoon DH, Noh HJ, Lee S, Kim TW, Sung GH, Cho JY. In vitro and in vivo anti-inflammatory effect of Rhodomyrtus tomentosa methanol extract. J Ethnopharmacol. 2013;146:205–213. doi: 10.1016/j.jep.2012.12.034. [DOI] [PubMed] [Google Scholar]
- Jeong D, Yi YS, Sung GH, Yang WS, Park JG, Yoon K, Yoon DH, Song C, Lee Y, Rhee MH, Kim TW, Kim JH, Cho JY. Anti-inflammatory activities and mechanisms of Artemisia asiatica ethanol extract. J Ethnopharmacol. 2014a;152:487–496. doi: 10.1016/j.jep.2014.01.030. [DOI] [PubMed] [Google Scholar]
- Jeong HY, Sung G-H, Kim JH, Yoon JY, Yang Y, Park JG, Kim SH, Yi Y-S, Yang WS, Yoon DH. Syk and Src are major pharmacological targets of a Cerbera manghas methanol extract with kaempferol-based anti-inflammatory activity. J Ethnopharmacol. 2014b;151:960–969. doi: 10.1016/j.jep.2013.12.009. [DOI] [PubMed] [Google Scholar]
- Jung CH, Kim JH, Kim JH, Chung JH, Choi HS, Seo JB, Shin YC, Kim SH, Ko SG. Anti-inflammatory effect of Rhus verniviflua Stokes by suppression of iNOS-mediated Akt and ERK pathways: in-vitro and in-vivo studies. J Pharm Pharmacol. 2011;63:679–687. doi: 10.1111/j.2042-7158.2011.01251.x. [DOI] [PubMed] [Google Scholar]
- Kang GJ, Han SC, Ock JW, Kang HK, Yoo ES. Anti-inflammatory effect of quercetagetin, an active component of immature Citrus unshiu, in HaCaT human keratinocytes. Biomol Ther. 2013;21:138–145. doi: 10.4062/biomolther.2013.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Khan N, Syed DN, Ahmad N, Mukhtar H. Fisetin: a dietary antioxidant for health promotion. Antioxid Redox Signal. 2013;19:151–162. doi: 10.1089/ars.2012.4901. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim DH, Chung JH, Yoon JS, Ha YM, Bae S, Lee EK, Jung KJ, Kim MS, Kim YJ, Kim MK, Chung HY. Ginsenoside Rd inhibits the expressions of iNOS and COX-2 by suppressing NF-kappaB in LPS-stimulated RAW264.7 cells and mouse liver. J Ginseng Res. 2013a;37:54–63. doi: 10.5142/jgr.2013.37.54. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim JH, Lee YG, Yoo S, Oh J, Jeong D, Song WK, Yoo BC, Rhee MH, Park J, Cha SH, Hong S, Cho JY. Involvement of Src and the actin cytoskeleton in the anti-tumorigenic action of adenosine dialdehyde. Biochem Pharmacol. 2013b;85:1042–1056. doi: 10.1016/j.bcp.2013.01.012. [DOI] [PubMed] [Google Scholar]
- Kim JH, Shin YC, Ko SG. Integrating traditional medicine into modern inflammatory diseases care: multitargeting by Rhus verniciflua Stokes. Mediators Inflamm. 2014a;2014:154561. doi: 10.1155/2014/154561. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim KH, Moon E, Choi SU, Pang C, Kim SY, Lee KR. Identification of cytotoxic and anti-inflammatory constituents from the bark of Toxicodendron vernicifluum (Stokes) F.A. Barkley. J Ethnopharmacol. 2015;162:231–237. doi: 10.1016/j.jep.2014.12.071. [DOI] [PubMed] [Google Scholar]
- Kim MY, Yoo BC, Cho JY. Ginsenoside-Rp1-induced apolipoprotein A-1 expression in the LoVo human colon cancer cell line. J Ginseng Res. 2014b;38:251–255. doi: 10.1016/j.jgr.2014.06.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim S-C, Kang S-H, Jeong S-J, Kim S-H, Ko HS, Kim S-H. Inhibition of c-Jun N-terminal kinase and nuclear factor κ B pathways mediates fisetin-exerted anti-inflammatory activity in lipopolysccharide-treated RAW264.7 cells. Immunopharmacol Immunotoxicol. 2012;34:645–650. doi: 10.3109/08923973.2011.648270. [DOI] [PubMed] [Google Scholar]
- Lee JA, Lee MY, Shin IS, Seo CS, Ha H, Shin HK. Anti-inflammatory effects of Amomum compactum on RAW 264.7 cells via induction of heme oxygenase-1. Arch Pharm Res. 2012;35:739–746. doi: 10.1007/s12272-012-0419-x. [DOI] [PubMed] [Google Scholar]
- Lee YG, Chain BM, Cho JY. Distinct role of spleen tyrosine kinase in the early phosphorylation of inhibitor of kappaB alpha via activation of the phosphoinositide-3-kinase and Akt pathways. Int J Biochem Cell Biol. 2009;41:811–821. doi: 10.1016/j.biocel.2008.08.011. [DOI] [PubMed] [Google Scholar]
- Oh CT, Park JI, Jung YR, Joo YA, Shin DH, Cho HJ, Ahn SM, Lim YH, Park CK, Hwang JS. Inhibitory effect of Korean red ginseng on melanocyte proliferation and its possible implication in GM-CSF mediated signaling. J Ginseng Res. 2013;37:389–400. doi: 10.5142/jgr.2013.37.389. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oh J, Yu T, Choi SJ, Yang Y, Baek HS, An SA, Kwon LK, Kim J, Rho HS, Shin SS, Choi WS, Hong S, Cho JY. Syk/Src pathway-targeted inhibition of skin inflammatory responses by carnosic acid. Mediators Inflamm. 2012;2012:781375. doi: 10.1155/2012/781375. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pal HC, Athar M, Elmets CA, Afaq F. Fisetin inhibits UVB-induced cutaneous inflammation and activation of PI3K/AKT/NFκB signaling pathways in SKH-1 hairless mice. Photochem Photobiol. 2015;91:225–234. doi: 10.1111/php.12337. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Park DK, Lee YG, Park HJ. Extract of Rhus verniciflua bark suppresses 2,4-dinitrofluorobenzene-induced allergic contact dermatitis. Evid Based Complement Alternat Med. 2013;2013:879696. doi: 10.1155/2013/879696. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pauwels R, Balzarini J, Baba M, Snoeck R, Schols D, Herdewijn P, Desmyter J, De Clercq E. Rapid and automated tetrazolium-based colorimetric assay for the detection of anti-HIV compounds. J. Virol. Methods. 1988;20:309–321. doi: 10.1016/0166-0934(88)90134-6. [DOI] [PubMed] [Google Scholar]
- Rice-Evans CA, Miller NJ, Paganga G. Structure-anti-oxidant activity relationships of flavonoids and phenolic acids. Free Radic Biol Med. 1996;20:933–956. doi: 10.1016/0891-5849(95)02227-9. [DOI] [PubMed] [Google Scholar]
- Seo JM, Kang HM, Son KH, Kim JH, Lee CW, Kim HM, Chang SI, Kwon BM. Antitumor activity of flavones isolated from Artemisia argyi. Planta Med. 2003;69:218–222. doi: 10.1055/s-2003-38486. [DOI] [PubMed] [Google Scholar]
- Shen T, Lee J, Park MH, Lee YG, Rho HS, Kwak YS, Rhee MH, Park YC, Cho JY. Ginsenoside Rp1, a ginsenoside derivative, blocks promoter activation of iNOS and COX-2 genes by suppression of an IKKbeta-mediated NF-kB pathway in HEK293 cells. J Ginseng Res. 2011;35:200–208. doi: 10.5142/jgr.2011.35.2.200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sohn SH, Kim SK, Kim YO, Kim HD, Shin YS, Yang SO, Kim SY, Lee SW. A comparison of antioxidant activity of Korean white and red ginsengs on H2O2-induced oxidative stress in HepG2 hepatoma cells. J Ginseng Res. 2013;37:442–450. doi: 10.5142/jgr.2013.37.442. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Soler Palacios B, Estrada-Capetillo L, Izquierdo E, Criado G, Nieto C, Municio C, González-Alvaro I, Sánchez-Mateos P, Pablos JL, Corbí AL. Macrophages from the synovium of active rheumatoid arthritis exhibit an activin A-dependent pro-inflammatory profile. J Pathol. 2015;235:515–526. doi: 10.1002/path.4466. [DOI] [PubMed] [Google Scholar]
- Song SB, Tung NH, Quang TH, Ngan NT, Kim KE, Kim YH. Inhibition of TNF-alpha-mediated NF-kappaB transcriptional activity in HepG2 cells by dammarane-type saponins from Panax ginseng leaves. J Ginseng Res. 2012;36:146–152. doi: 10.5142/jgr.2012.36.2.146. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van den Berg WB, Bresnihan B. Pathogenesis of joint damage in rheumatoid arthritis: evidence of a dominant role for interleukin-1. Baillieres Best Pract Res Clin Rheumatol. 1999;13:577–597. doi: 10.1053/berh.1999.0047. [DOI] [PubMed] [Google Scholar]
- Vilahur G, Badimon L. Ischemia/reperfusion activates myocardial innate immune response: the key role of the toll-like receptor. Front Physiol. 2014;5:496. doi: 10.3389/fphys.2014.00496. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang K-J, Shin S, Piao L, Shin E, Li Y, Park KA, Byun HS, Won M, Hong J, Kweon GR. Regulation of 3-phosphoinositide-dependent protein kinase-1 (PDK1) by Src involves tyrosine phosphorylation of PDK1 and Src homology 2 domain binding. J Biol Chem. 2008;283:1480–1491. doi: 10.1074/jbc.M706361200. [DOI] [PubMed] [Google Scholar]
- Yang WS, Jeong D, Yi YS, Lee BH, Kim TW, Htwe KM, Kim YD, Yoon KD, Hong S, Lee WS, Cho JY. Myrsine seguinii ethanolic extract and its active component quercetin inhibit macrophage activation and peritonitis induced by LPS by targeting to Syk/Src/IRAK-1. J Ethnopharmacol. 2014a;151:1165–1174. doi: 10.1016/j.jep.2013.12.033. [DOI] [PubMed] [Google Scholar]
- Yang Y, Yang WS, Yu T, Yi YS, Park JG, Jeong D, Kim JH, Oh JS, Yoon K, Kim JH, Cho JY. Novel anti-inflammatory function of NSC95397 by the suppression of multiple kinases. Biochem Pharmacol. 2014b;88:201–215. doi: 10.1016/j.bcp.2014.01.022. [DOI] [PubMed] [Google Scholar]
- Yayeh T, Jung KH, Jeong HY, Park JH, Song YB, Kwak YS, Kang HS, Cho JY, Oh JW, Kim SK, Rhee MH. Korean red ginseng saponin fraction downregulates proinflammatory mediators in LPS stimulated RAW264.7 cells and protects mice against endotoxic shock. J Ginseng Res. 2012;36:263–269. doi: 10.5142/jgr.2012.36.3.263. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yi YS, Son YJ, Ryou C, Sung GH, Kim JH, Cho JY. Functional roles of Syk in macrophage-mediated inflammatory responses. Mediators Inflamm. 2014;2014:270302. doi: 10.1155/2014/270302. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yoon JY, Jeong HY, Kim SH, Kim HG, Nam G, Kim JP, Yoon DH, Hwang H, Kimc TW, Hong S, Cho JY. Methanol extract of Evodia lepta displays Syk/Src-targeted anti-inflammatory activity. J Ethnopharmacol. 2013;148:999–1007. doi: 10.1016/j.jep.2013.05.030. [DOI] [PubMed] [Google Scholar]
- Youn CK, Park SJ, Lee MY, Cha MJ, Kim OH, You HJ, Chang IY, Yoon SP, Jeon YJ. Silibinin inhibits LPS-induced macrophage activation by blocking p38 MAPK in RAW 264.7 Cells. Biomol Ther. 2013;21:258–263. doi: 10.4062/biomolther.2013.044. [DOI] [PMC free article] [PubMed] [Google Scholar]