

Abstract
Gastric cancer (GC), one of the most common human cancers, can be classified into gastric or intestinal phenotype according to mucin expression. TP53 mutation, allelic deletion of the APC gene and nuclear staining of β-catenin are frequently detected in the intestinal phenotype of GC, whereas CDH1 gene mutation, microsatellite instability and DNA hypermethylation of MLH1 are common events in the gastric phenotype of GC. Our Serial Analysis of Gene Expression (SAGE) and Escherichia coli ampicillin secretion trap (CAST) analyses revealed that CDH17, REG4, OLFM4, HOXA10, DSC2, TSPAN8 and TM9SF3 are upregulated in GC and that CLDN18 is downregulated in GC. Expression of CDH17, REG4, HOXA10 and DSC2 and downregulation of CLDN18 are observed in the intestinal phenotype of GC. In contrast, OLFM4 is expressed in the gastric phenotype of GC. Expression of TSPAN8, TM9SF3 and HER2 are not associated with either gastric or intestinal phenotypes. Ectopic CDX2 expression plays a key function in the GC intestinal phenotype. MUC2, CDH17, REG4, DSC2 and ABCB1 are direct targets of CDX2. Importantly, these genes encode transmembrane/secretory proteins, indicating that the microenvironment as well as cancer cells are also different between gastric and intestinal phenotypes of GC.
Keywords: Escherichia coli ampicillin secretion trap, gastric cancer, mucin, phenotype, Serial Analysis of Gene Expression
Gastric cancer (GC), one of the most common human cancers, is a heterogeneous disease with different phenotypes and varying prognoses and responses to treatment. Therefore, subtype classification of GC is necessary for prognosis prediction and decisions regarding effective treatment. The methods for subtype classification include molecular analysis, immunohistochemistry and histologic analysis. Histologically, GC cases are classified into two major types, the differentiated and undifferentiated types, as described by Nakamura et al.,1 or the Lauren intestinal and diffuse types,2 based on glandular structure. Intestinal and diffuse GC types show distinct clinical characteristics,3 and type-specific genetic and epigenetic alterations have been identified.4–6 Although Lauren classification is important information in clinical practice, it is not critical for prognosis prediction or determining treatment. Therefore, there is an urgent need for new histologic classification for GC.
Gastric cancer can also be classified into gastric or intestinal phenotype according to mucin expression. Accumulating evidence has indicated that gastric/intestinal phenotypes of GC have distinct clinical characteristics and exhibit specific genetic and epigenetic changes.7,8 Here we focus on the clinical and molecular characteristics of the gastric and intestinal phenotypes of GC.
Definition of gastric and intestinal phenotypes of gastric cancer
Previously, gastric/intestinal classification was determined by H&E staining. Egashira et al.9 provided the initial histologic characteristics of gastric/intestinal phenotypes of GC. The gastric phenotype of GC consists of cuboidal or columnar cells with clear cytoplasm that are arranged side-by-side like foveolar epithelial cells or pyloric gland cells. Their nuclei are round and situated in the basal cytoplasm. Mucin droplets are found in the apical cytoplasm. In contrast, the intestinal phenotype of GC resembles colorectal cancer, and is mainly composed of columnar cells with eosinophilic cytoplasm and goblet cell differentiation. The intraluminal surface of tubules has a striated border and surface coat mucin.
After 2000, the gastric/intestinal phenotypes of GC were analyzed by immunohistochemistry using MUC5AC (or Human gastric mucin (HGM)) and MUC6 as markers for the gastric phenotype, and MUC2 and CD10 (or villin) as markers for the intestinal phenotype. Based on expression of these markers, GC cases are classified into four phenotypes: gastric or foveolar (G type), intestinal (I type), gastric and intestinal mixed (GI type), and neither gastric nor intestinal (N type).10,11 Several additional classifications have been proposed. Kabashima et al.12 classify GC cases into four phenotypes: complete intestinal, incomplete intestinal, gastric and unclassified. Egashira et al.9 classify GC cases into three phenotypes: G-type GC, I-type GC and GI-type GC. This indicates that there are several strategies for gastric/intestinal phenotype classification, although it remains unclear as to which definition is the best. However, at the least, these approaches demonstrate that immunohistochemical analysis is required for classification of gastric/intestinal phenotypes in addition to H&E staining.
Representative images of gastric/intestinal phenotype GC cases are shown in Figure1(a). We previously analyzed the phenotypes of 870 GC cases.13 However, a clear association between the gastric/intestinal phenotypes and histological classification of Japanese classification of gastric carcinoma (3rd GC English edition)14 was not observed (Fig.1b).
Figure 1.
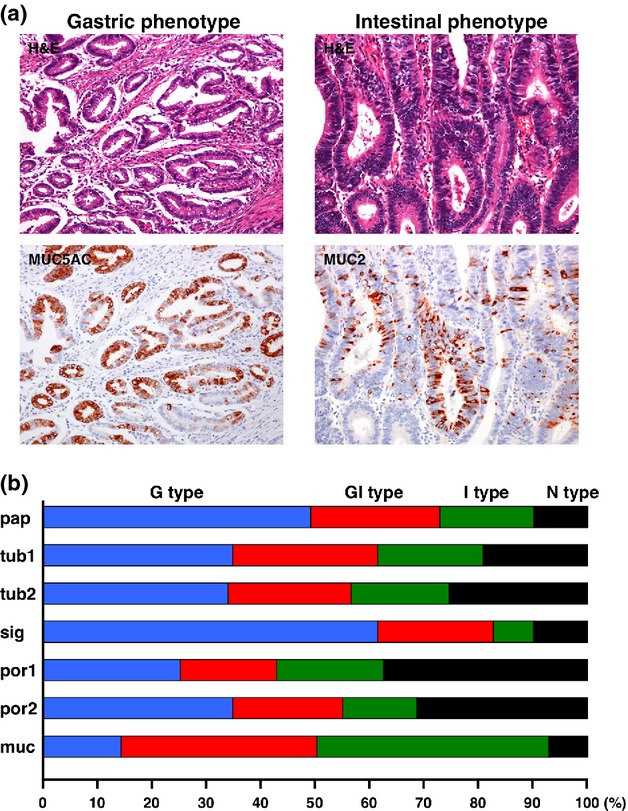
Representative images of gastric and intestinal phenotypes of gastric cancer (GC). (a) The gastric phenotype of GC shows cuboidal or columnar cells arranged side by side like foveolar epithelial cells. The nuclei are round and situated in the basal cytoplasm. Staining of MUC5AC is observed. The intestinal phenotype of GC resembles colorectal cancer, and is mainly composed of columnar cells with eosinophilic cytoplasm and goblet cell differentiation. MUC2 staining is detected. Original magnification, ×200. (b) Analysis of association between gastric/intestinal phenotypes and histological classification of Japanese classification of gastric carcinoma in 870 GC cases.13
Clinical characteristics of gastric cancer gastric/intestinal phenotypes
Substantial effort has been devoted to analyzing characteristics of gastric/intestinal phenotypes of GC. Although the data have been controversial and the characteristics of gastric/intestinal GC types are still ambiguous, several important points have been established. One important concept is that almost all intramucosal GC cases show a gastric phenotype, including gastric and intestinal mixed phenotype, whereas gastric phenotype is diminished according to GC progression. A previous report revealed that the incidence of GC showing gastric phenotype decreases as the tumor diameter increases.9 In submucosal GC, the frequency of gastric phenotype decreases.15 GC cases at early stages, independent of the histological type, mainly consist of gastric phenotype, and phenotypic shift from gastric to intestinal phenotype is clearly observed with progression of tumor stage.7 There is no clear correlation between phenotype and clinicopathologic characteristics, including sex, age, location of GC, or macroscopic features.15
Prognosis of gastric/intestinal phenotypes of GC has been investigated. However, a definitive conclusion has not been established. One possible explanation is that each study uses different definitions of gastric/intestinal phenotypes. To establish a clear association between prognosis and gastric/intestinal phenotypes, the gastric/intestinal phenotype definitions must be clarified.
Molecular characterization of gastric cancer gastric/intestinal phenotypes
To understand the gastric/intestinal phenotypes of GC at the molecular level, several genetic and epigenetic alterations have been investigated (Table1). TP53 mutation and allelic deletion of the APC gene are detected more frequently in the intestinal phenotype of GC.8,16 In contrast, CDH1 gene mutation is detected in differentiated type GC a showing gastric phenotype.17 Microsatellite instability (MSI) is detected more frequently in the gastric phenotype of GC.8,18 Alterations of TP73, including loss of heterozygosity and abnormal expression, play an important role in the genesis of the gastric phenotype of GC.19 Several epigenetic alterations have also been identified. DNA methylation of MLH1 gene frequently occurs in the gastric phenotype of GC,20 whereas MGMT gene is frequently methylated in the intestinal phenotype of GC.21
Table 1.
Summary of genetic/epigenetic/gene expression alterations in gastric and intestinal phenotypes of gastric cancer
| Function | Frequency (%) | References | ||
|---|---|---|---|---|
| Gastric phenotype | Intestinal phenotype | |||
| Transcription factor | SOX2 | CDX1, CDX2 | ||
| TP53 mutation | Tumor suppressor | 5 | 31 | 8 |
| APC deletion | Tumor suppressor | 0 | 38 | 16 |
| CDH1 mutation | Calcium-dependent cell adhesion protein | 21 | 0 | 17 |
| TP73 deletion | Apoptotic response to DNA damage | 80 | 0 | 19 |
| Microsatellite instability | – | 45 | 0 | 8 |
| MLH1 DNA methylation | DNA mismatch repair | 74 | 33 | 20 |
| MGMT DNA methylation | DNA repair | 46 | 82 | 21 |
| β-catenin nuclear staining | Canonical Wnt signaling pathway | 6 | 46 | 13 |
| AID expression | Single-stranded DNA-specific cytidine deaminase | 14 | 38 | 22 |
| EGFR expression | Receptor tyrosine kinase | 12 | 31 | 32 |
| Cytokeratin profile | – | CK7+/CK20− | CK7−/CK20+ | 13 |
| LI-cadherin expression | Calcium-dependent cell adhesion protein | 5 | 63 | 32 |
| Reg IV expression | Calcium-independent lectin | 1 | 77 | 11 |
| HoxA10 expression | Sequence-specific transcription factor | 25 | 44 | 54 |
| Desmocollin 2 expression | Component of intercellular junction | 28 | 45 | 61 |
| MDR1 expression | Energy-dependent efflux pump | 48 | 74 | 83 |
| Olfactomedin 4 expression | Unknown function | 73 | 44 | 46 |
| Claudin-18 down-regulation | Calcium-independent cell-adhesion | 44 | 74 | 70 |
AID, activation-induced cytidine deaminase; CK, cytokeratin; EGFR, epidermal growth factor receptor.
Expression of cancer-associated genes has also been investigated by immunohistochemistry (Table1). Aberrant expression of activation-induced cytidine deaminase is common event in intestinal phenotype of GC.22 Studies have shown that the cytokeratin (CK) profile is different between GC and colorectal cancer. Colorectal cancer shows a CK7−/CK20+ expression pattern, whereas adenocarcinomas of foregut origin, including GC, demonstrate a CK7+/CK20− expression pattern.23 In our study, GC cases showing CK7−/CK20+ were frequently found in intestinal phenotype of GC, whereas GC cases showing CK7+/CK20− were commonly found in gastric phenotype of GC.13 Nuclear β-catenin staining was frequently found in the intestinal phenotype of GC. However, expression of MMP7, laminin γ2 or HER2 was not correlated with GC gastric or intestinal phenotypes.24 Together these observations indicate that in addition to histologic characteristics, genetic, epigenetic and gene expression alterations in the intestinal phenotype of GC are similar to those of colorectal cancer, while those of the gastric phenotype of GC are clearly different from those of colorectal cancer.
Identification of gastric cancer-associated genes by comprehensive gene expression analysis
To identify potential molecular markers for GC and to better understand the development of GC at the molecular level, comprehensive gene expression analysis is useful. Serial Analysis of Gene Expression (SAGE) is used to analyze 14-bp tags derived from defined positions of cDNA without a priori knowledge of the sequence of the genes expressed, and offers an unbiased, comprehensive gene expression profiling approach.25 Escherichia coli ampicillin secretion trap (CAST) is a signal sequence trap method to identify genes that encode transmembrane or secretory proteins.26 Schematic outline of the SAGE and CAST methods are shown in Figures S1 and S2. We performed SAGE and CAST on GC samples and identified several genes whose expression was altered in GC. Among them, CDH17, REG4, OLFM4, HOXA10, DSC2, TSPAN8 and TM9SF3 were upregulated in GC, and CLDN18 was downregulated in GC. Importantly, many of these genes are tightly associated with gastric/intestinal phenotypes of GC.
CDH17
Through SAGE analysis, CDH17 was found to be one of the upregulated genes in GC.27 CDH17 encodes the liver-intestinal (LI)-cadherin protein, a member of the cadherin family of cell adhesion molecules.28 LI-cadherin mediates homotypic Ca2+-dependent cell–cell adhesion in L cells.29 LI-cadherin is a structurally different cadherin that is specifically expressed in the liver and intestine of the rat.28 In contrast, human LI-cadherin is found in the intestinal epithelium but not in the liver. In the human intestinal mucosa, LI-cadherin is concentrated in the lateral domain of the plasma membrane.
Our immunohistochemical study detected LI-cadherin expression in 67% of GC tissue samples, and LI-cadherin expression was significantly more frequent in advanced stage GC than in early stage GC.30 A previous report showed that LI-cadherin is a marker of intestinal metaplasia of the stomach,31 suggesting LI-cadherin as a marker for the intestinal phenotype. Indeed, LI-cadherin expression is frequently found in the intestinal phenotype of GC.32 These results indicate that LI-cadherin is one of the key regulators for the intestinal phenotype of GC.
REG4
Through SAGE analysis, REG4 was found to be one of the upregulated genes in GC.27 REG4 is a member of the REG gene family and encodes Reg IV protein. REG4 was originally identified by high-throughput sequence analysis of an inflammatory bowel disease cDNA library.33 Reg IV is an activator of the epidermal growth factor receptor (EGFR) signaling pathway and increases expression of Bcl-2, Bcl-xl and survivin, which inhibit apoptosis.34 We previously reported that Reg IV inhibits 5-fluorouracil (5-FU)-induced apoptosis through EGFR activation in GC cells.35 It has been reported that Reg IV is upregulated in undifferentiated-type GC, and that the increased tumourigenic ability of ALDH1-positive cells depends on Reg IV.36 These findings support the notion that Reg IV protein participates in carcinogenesis.
In non-neoplastic human tissue, Reg IV expression is narrowly restricted.11 In non-neoplastic stomach, foveolar epithelial cells do not express Reg IV, whereas goblet cells of intestinal metaplasia and neuroendocrine cells at the base of intestinal metaplasia express Reg IV, suggesting Reg IV as a marker for the intestinal phenotype. Expression and localization of Reg IV in human cancers have been analyzed by immunohistochemistry.37–42 The immunohistochemistry reports show that Reg IV is overexpressed in adenocarcinoma cells that are positive for MUC2. Overexpression of Reg IV is also observed in neuroendocrine neoplasms. Intestinal carcinoid tumors, parathyroidal cell tumors, small-cell carcinomas of the lung, and Merkel cell carcinomas also overexpress Reg IV.11,43 In our study, Reg IV expression was detected in 29% of GC cases, and was frequently found in the intestinal phenotype of GC.11 Together these data indicate that Reg IV activates EGFR and plays an important role in the inhibition of apoptosis in the intestinal phenotype of GC.
OLFM4
OLFM4 was identified as one of the upregulated genes in GC in SAGE analysis.27 OLFM4 encodes olfactomedin 4 protein (also known as hGC-1 or GW112) and was originally cloned from human myeloblasts.44 Although the precise function of olfactomedin 4 is unclear, a previous study revealed that olfactomedin 4 is a highly specific and robust marker for Lgr5-positive stem cells of the small intestine,45 suggesting that olfactomedin 4 plays an important role in stem cell function. In fact, olfactomedin 4 expression is detected in crypt base columnar cells, which are intestinal stem cells. Thus, antibody against olfactomedin 4 is useful to identify intestinal stem cells.46 Using anti-olfactomedin 4 antibody in APC(Min/+) mice, we found that dietary sulindac induces apoptosis to remove intestinal stem cells with nuclear or phosphorylated β-catenin.47 We also showed that canonical Wnt signals support homeostatic intestinal stem/progenitor cell proliferation.48 Two studies have investigated OLFM4 knockout (KO) mice. Liu et al.49 demonstrated that Helicobacter pylori colonization in the gastric mucosa of OLFM4 KO mice was significantly lower compared with wild-type mice, and reduced bacterial colonization was associated with enhanced infiltration of inflammatory cells in gastric mucosa. Schuijers et al.50 report that OLFM4 KO mice showed no phenotype.
In non-neoplastic stomach, foveolar epithelial cells do not express olfactomedin 4, whereas olfactomedin 4 is expressed in the basal crypt epithelium in the intestinal metaplasia of the stomach, suggesting that olfactomedin 4 may be a marker for the intestinal phenotype.46 In our immunohistochemical analysis,46 expression of olfactomedin 4 was detected in 56% of GC cases, and expression of olfactomedin 4 was frequently detected in well-differentiated adenocarcinomas. In well-differentiated adenocarcinomas, patients with olfactomedin 4-positive GC have a better survival rate than those with olfactomedin 4-negative GC. Expression of olfactomedin 4 was frequently observed in the gastric phenotype. Together this indicates that olfactomedin 4 plays an important role in the gastric phenotype of GC. Similar results were shown in colorectal and endometrioid adenocarcinoma.51–53
Both Reg IV and olfactomedin 4 are secreted proteins, and serum Reg IV and olfactomedin 4 serve as tumor markers for GC. The sensitivity and specificity of serum olfactomedin 4 combined with Reg IV for GC detection were 52% and 95%, respectively.46 These data suggest that serum olfactomedin 4 combined with Reg IV is likely to be suitable for screening of GC.
HOXA10
HOXA10, which encodes HoxA10 protein, was identified as one of the upregulated genes in GC in SAGE analysis.54 HOX genes are important regulators of embryonic morphogenesis and differentiation, and control normal development patterning along the anteroposterior axis.55 The homeodomain binds to sequence-specific DNA motifs and regulates the transcription of genes relevant to the formation of specific segmental architecture. Overexpression of HOXA10 has been detected in prostate, lung and ovarian cancer.56–58 Forced expression of HoxA10 has been reported to promote cell proliferation, suggesting that overexpression of HoxA10 may participate in the pathogenesis of cancer.56 In contrast, another study showed that HoxA10 induces expression of CDKN1, which encodes p21 protein,59 and downregulation of HoxA10 has been reported in endometrial cancer.60 Therefore, the significance of HoxA10 expression in human cancers is still unclear, and further investigation is required.
Our study by immunohistochemistry revealed that in non-neoplastic stomach, foveolar epithelial cells do not express HoxA10, whereas HoxA10 is expressed in the intestinal metaplasia of the stomach, suggesting that HoxA10 could be a marker for the intestinal phenotype.54 HoxA10 expression was detected in 30% of GC cases, and the prognosis of patients with positive HoxA10 expression was significantly better than those with negative HoxA10 expression. In addition, HoxA10 expression is frequently found in the intestinal phenotype of GC. Together this suggests that HoxA10 is a key factor in the intestinal phenotype of GC.
DSC2
Through CAST analysis, DSC2 was identified as one of the upregulated genes in GC.61 DSC2 encodes desmocollin 2 protein, one of the three known desmocollin proteins. In the mature organism, desmosomes are most abundant in areas subject to mechanical stress, including skin, heart and esophagus.62 Desmocollins are membrane-spanning glycoproteins that form desmosomes along with desmogleins and function as Ca2+-dependent cell adhesion molecules.63
In our study, immunohistochemical analysis showed weak or no staining of desmocollin 2 in the foveolar epithelium of the stomach, whereas desmocollin 2 expression was observed in the intestinal metaplasia. Expression of desmocollin 2 was detected in 28% of GC tissue samples, and was frequently found in the intestinal phenotype of GC.61
TSPAN8
Through CAST analysis, TSPAN8 was demonstrated to be one of the upregulated genes in GC.64 TSPAN8 encodes tetraspanin 8 protein and is a member of the tetraspanin family. Tetraspanin proteins cross the membrane four times and are involved in numerous biological processes.65 Tetraspanin proteins are components of exosomes, and exosomes containing rat Tspan8 have been shown to affect tumor cell migration, proliferation and tumor angiogenesis.66
Our immunohistochemical analysis revealed that 34% of GC cases were positive for tetraspanin 8, and microvessel density was higher in tetraspanin 8-positive GC cases compared with tetraspanin 8-negative GC cases.64 Furthermore, tetraspanin 8 expression was an independent prognostic classifier of patients with GC. Expression of tetraspanin 8 was not associated with the gastric or intestinal phenotype of GC, indicating that tetraspanin 8 plays a crucial role in both the gastric and intestinal phenotypes of GC.
TM9SF3
TM9SF3 was identified as one of the upregulated genes in GC by CAST analysis.67 TM9SF3, which encodes transmembrane 9 superfamily member 3 protein, is a member of the TM9SF family. TM9SF proteins are characterized by a large noncytoplasmic domain and nine putative transmembrane domains.68 TM9SF proteins are required for adhesion and phagocytosis in innate immune responses; however, the biological functions of TM9SF proteins are largely unknown.69
We found that 50% of GC cases were positive for transmembrane 9 superfamily member 3 protein.67 Expression of transmembrane 9 superfamily member 3 protein is frequently detected in scirrhous-type GC and associated with poor prognosis. There was no association between expression of transmembrane 9 superfamily member 3 protein and the gastric or intestinal phenotype. Together these data suggest that transmembrane 9 superfamily member 3 protein is an ideal molecular target for treatment of scirrhous-type GC.
CLDN18
CLDN18 was one of the downregulated genes in GC identified by SAGE analysis.70 CLDN18, which encodes claudin-18 protein, is a member of the claudin family, and a component of tight junctions. The claudin family comprises 27 members, and all claudins are 20–27 kDa proteins with four transmembrane domains.71 CLDN18 has two variants in mice: variant 1 is expressed in the lung, whereas variant 2 is expressed in the stomach.72
In our immunohistochemistry analysis,70 in non-neoplastic stomach, foveolar epithelial cells expressed claudin-18 on the cell membrane, whereas claudin-18 was not expressed in the intestinal metaplasia of the stomach. This suggests that claudin-18 may be a marker for the gastric phenotype. Downregulation of claudin-18 was observed in 58% of GC cases and was correlated with poor survival. Downregulation of claudin-18 was frequently found in the intestinal phenotype of GC. Although the precise function of claudin-18 has not been described, the function of the tight junction is to maintain the luminal barrier, paracellular transport and signal transduction. Therefore, downregulation of claudin-18 and disruption of tight junctions can cause loss of cell polarity, resulting in an abnormal influx of growth factors, which can provide autocrine and paracrine stimulation to tumorigenic epithelial cells. Indeed, CLDN18 KO mice show paracellular H+ leakage, upregulation of interleukin-1β and atrophic gastritis.73 These results indicate that downregulation of claudin-18 participates in the pathogenesis of the intestinal phenotype of GC.
A recent study demonstrated an interchromosomal translocation between CLDN18 and ARHGAP26.74 ARHGAP26 is a GTPase-activating protein that facilitates conversion of RHO GTPases to the GDP state and has been implicated in enhancing cellular motility. However, the significance of interchromosomal translocation between CLDN18 and ARHGAP26 has not been analyzed.
IMAB362, a highly potent and tumor-cell selective therapeutic antibody, is a medicinal product directed against the tight junction molecule claudin-18 variant 2.75 A Phase II trial (NCT01630083), in which IMAB362 is combined with standard chemotherapy for first-line treatment of gastroesophageal cancer, is ongoing (NCT01630083).
It is important to note that these genes encode transmembrane/secretory proteins, suggesting that the microenvironment as well as cancer cells are different between gastric and intestinal phenotypes of GC. Although the precise functions of Reg IV and olfactomedin 4 are unclear, these two proteins are upregulated in inflammatory bowel disease.33,49 Therefore, inflammatory response may be different between gastric and intestinal phenotypes of GC.
Transcription factors of gastric/intestinal phenotypes of gastric cancer and their target genes
Several transcription factors that induce the gastric/intestinal phenotypes have been identified. In the intestinal phenotype of GC, ectopic CDX2 expression has a key function.7 In mammals, the CDX1 and CDX2 homeobox transcription factors play critical roles in intestinal development, differentiation and maintenance of the intestinal phenotype.76 CDX1 and CDX2 proteins show significant homology to the protein product of the Drosophila caudal gene, a key regulator of anterior–posterior regional identity. Ectopic expression of Cdx2 in the gastric mucosa of transgenic mice induced intestinal metaplasia.77 In contrast, SOX2 may be an important transcription factor of the gastric phenotype of GC. SOX2 induces expression of MUC5AC and pepsinogen A, both of which are markers for the gastric phenotype.78,79 Furthermore, SOX2 negatively regulates the CDX2 promoter by hampering the action of other transcription factors.80
To characterize the intestinal phenotype of GC, identification of CDX2 target genes is important. Among the genes aberrantly expressed in GC described above, we showed that CDH17, REG4 and DSC2 are direct targets of CDX2, and these genes are expressed in CDX2-positive GC cells.61,81,82 For further characterization of the intestinal phenotype of GC, we investigated CDX2-target genes, and found that ABCB1, which encodes multidrug resistance protein 1 (MDR1), is a direct target of CDX2.83 Immunohistochemical analysis detected MDR1 expression in CDX2-positive GC cells, and showed that MDR1-positive GC cases are frequently found in the intestinal phenotype of GC. As described above, CDX2 induces Reg IV expression. Reg IV inhibits 5-FU-induced apoptosis and MDR1 inhibits taxane-induced apoptosis. These data lead us to speculate that chemotherapy, including 5-FU-based or taxane-based chemotherapy, is not beneficial in patients with the intestinal phenotype of GC. For these patients, molecular-targeted therapies could be suitable.
Conclusions
Molecular alterations of GC are summarized in Figure2. Expressions of transmembrane/secretory proteins in cancers are ideal diagnostic biomarkers. Moreover, if these molecules are involved in the neoplastic process, the molecules are not just biomarkers but may also be therapeutic targets. Here we described clinical and molecular characteristics of the gastric/intestinal phenotypes of GC. Expression of transmembrane/secretory proteins, including LI-cadherin, Reg IV, olfactomedin 4, desmocollin 2 and claudin-18, is different between gastric and intestinal phenotypes of GC. These transmembrane/secretory proteins are extracellular proteins, indicating that the microenvironment as well as cancer cells are different between the gastric and intestinal phenotypes. About 10 years ago, oncogenic driver mutations have emerged as major treatment targets for molecular therapies in a variety of cancers. Whole genome or exon sequencing in GC has been performed, and mutation of RHOA gene in undifferentiated-type GC has been reported.84 According to the COSMIC website (http://cancer.sanger.ac.uk),85 the most frequently mutated gene is TP53 (32%), and the second most frequently mutated gene is ARID1A (14%).86 Frequencies of other gene mutations are approximately 10% or below 10%. Although the associations between mutation of these genes and gastric/intestinal phenotypes are unclear, driver gene mutation is a rare event, and it is difficult to plan an effective treatment according to driver gene mutations. In contrast, the Cancer Genome Atlas Network has reported that GC can be classified into four distinct molecular subtypes: GC positive for Epstein–Barr virus; microsatellite unstable GC; genomically stable GC; and GC with chromosomal instability.74 As described above, MSI is detected more frequently in the gastric phenotype of GC.8 GC positive for Epstein–Barr virus are also frequently found in the gastric phenotype of GC.87 However, the mucin phenotypes of genomically stable GC and GC with chromosomal instability remains unclear. Classification of these subtypes may be used to provide personalized medicine.
Figure 2.

Molecular alterations of gastric cancer (GC). This graphic overview depicts the specific alterations in differentiated/undifferentiated GC or intestinal/gastric phenotypes of GC.
Acknowledgments
This work was supported, in part, by Grants-in-Aid for Cancer Research from the Ministry of Education, Science, Sports and Culture of Japan and from the Ministry of Health and Welfare of Japan.
Disclosure statement
The authors have no conflict of interest to declare.
Supporting Information
Fig. S1. Schematic outline of the Serial Analysis of Gene Expression (SAGE) method.
Fig. S2. Schematic outline of the Escherichia coli ampicillin secretion trap (CAST) method.
References
- Nakamura K, Sugano H, Takagi K. Carcinoma of the stomach in incipient phase: its histogenesis and histological appearances. Gann. 1968;59:251–8. [PubMed] [Google Scholar]
- Lauren P. The two histological main types of gastric carcinoma. Diffuse and so-called intestinal type carcinoma: an attempt at histological classification. Acta Pathol Microbiol Scand. 1965;64:31–49. doi: 10.1111/apm.1965.64.1.31. [DOI] [PubMed] [Google Scholar]
- Vauhkonen M, Vauhkonen H, Sipponen P. Pathology and molecular biology of gastric cancer. Best Pract Res Clin Gastroenterol. 2006;20:651–74. doi: 10.1016/j.bpg.2006.03.016. [DOI] [PubMed] [Google Scholar]
- Oue N, Oshimo Y, Nakayama H, et al. DNA methylation of multiple genes in gastric carcinoma: association with histological type and CpG island methylator phenotype. Cancer Sci. 2003;94:901–5. doi: 10.1111/j.1349-7006.2003.tb01373.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yasui W, Oue N, Ito R, Kuraoka K, Nakayama H. Search for new biomarkers of gastric cancer through serial analysis of gene expression and its clinical implications. Cancer Sci. 2004;95:385–92. doi: 10.1111/j.1349-7006.2004.tb03220.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yasui W, Sentani K, Sakamoto N, Anami K, Naito Y, Oue N. Molecular pathology of gastric cancer: research and practice. Pathol Res Pract. 2011;207:608–12. doi: 10.1016/j.prp.2011.09.006. [DOI] [PubMed] [Google Scholar]
- Tatematsu M, Tsukamoto T, Inada K. Stem cells and gastric cancer: role of gastric and intestinal mixed intestinal metaplasia. Cancer Sci. 2003;94:135–41. doi: 10.1111/j.1349-7006.2003.tb01409.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Endoh Y, Sakata K, Tamura G, et al. Cellular phenotypes of differentiated-type adenocarcinomas and precancerous lesions of the stomach are dependent on the genetic pathways. J Pathol. 2000;191:257–63. doi: 10.1002/1096-9896(2000)9999:9999<::AID-PATH631>3.0.CO;2-2. [DOI] [PubMed] [Google Scholar]
- Egashira Y, Shimoda T, Ikegami M. Mucin histochemical analysis of minute gastric differentiated adenocarcinoma. Pathol Int. 1999;49:55–61. doi: 10.1046/j.1440-1827.1999.00824.x. [DOI] [PubMed] [Google Scholar]
- Mizoshita T, Tsukamoto T, Nakanishi H, et al. Expression of Cdx2 and the phenotype of advanced gastric cancers: relationship with prognosis. J Cancer Res Clin Oncol. 2003;129:727–34. doi: 10.1007/s00432-003-0499-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oue N, Mitani Y, Aung PP, et al. Expression and localization of Reg IV in human neoplastic and non-neoplastic tissues: Reg IV expression is associated with intestinal and neuroendocrine differentiation in gastric adenocarcinoma. J Pathol. 2005;207:185–98. doi: 10.1002/path.1827. [DOI] [PubMed] [Google Scholar]
- Kabashima A, Yao T, Sugimachi K, Tsuneyoshi M. Gastric or intestinal phenotypic expression in the carcinomas and background mucosa of multiple early gastric carcinomas. Histopathology. 2000;37:513–22. doi: 10.1046/j.1365-2559.2000.01008.x. [DOI] [PubMed] [Google Scholar]
- Takami H, Sentani K, Matsuda M, Oue N, Sakamoto N, Yasui W. Cytokeratin expression profiling in gastric carcinoma: clinicopathologic significance and comparison with tumor-associated molecules. Pathobiology. 2012;79:154–61. doi: 10.1159/000335694. [DOI] [PubMed] [Google Scholar]
- Japanese Gastric Cancer Association. Japanese classification of gastric carcinoma: 3rd English edition. Gastric Cancer. 2011;14:101–12. doi: 10.1007/s10120-011-0041-5. [DOI] [PubMed] [Google Scholar]
- Nakamura T, Yao T, Kabashima A, Nishiyama K, Maehara Y, Tsuneyoshi M. Loss of phenotypic expression is related to tumour progression in early gastric differentiated adenocarcinoma. Histopathology. 2005;47:357–67. doi: 10.1111/j.1365-2559.2005.02242.x. [DOI] [PubMed] [Google Scholar]
- Wu LB, Kushima R, Borchard F, Molsberger G, Hattori T. Intramucosal carcinomas of the stomach: phenotypic expression and loss of heterozygosity at microsatellites linked to the APC gene. Pathol Res Pract. 1998;194:405–11. doi: 10.1016/S0344-0338(98)80031-9. [DOI] [PubMed] [Google Scholar]
- Endoh Y, Tamura G, Watanabe H, Ajioka Y, Motoyama T. The common 18-base pair deletion at codons 418–423 of the E-cadherin gene in differentiated-type adenocarcinomas and intramucosal precancerous lesions of the stomach with the features of gastric foveolar epithelium. J Pathol. 1999;189:201–6. doi: 10.1002/(SICI)1096-9896(199910)189:2<201::AID-PATH409>3.0.CO;2-A. [DOI] [PubMed] [Google Scholar]
- Shibata N, Watari J, Fujiya M, Tanno S, Saitoh Y, Kohgo Y. Cell kinetics and genetic instabilities in differentiated type early gastric cancers with different mucin phenotype. Hum Pathol. 2003;34:32–40. doi: 10.1053/hupa.2003.2. [DOI] [PubMed] [Google Scholar]
- Yokozaki H, Shitara Y, Fujimoto J, Hiyama T, Yasui W, Tahara E. Alterations of p73 preferentially occur in gastric adenocarcinomas with foveolar epithelial phenotype. Int J Cancer. 1999;83:192–6. doi: 10.1002/(sici)1097-0215(19991008)83:2<192::aid-ijc8>3.0.co;2-e. [DOI] [PubMed] [Google Scholar]
- Endoh Y, Tamura G, Ajioka Y, Watanabe H, Motoyama T. Frequent hypermethylation of the hMLH1 gene promoter in differentiated-type tumors of the stomach with the gastric foveolar phenotype. Am J Pathol. 2000;157:717–22. doi: 10.1016/S0002-9440(10)64584-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Motoshita J, Oue N, Nakayama H, et al. DNA methylation profiles of differentiated-type gastric carcinomas with distinct mucin phenotypes. Cancer Sci. 2005;96:474–9. doi: 10.1111/j.1349-7006.2005.00074.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goto A, Hirahashi M, Osada M, et al. Aberrant activation-induced cytidine deaminase expression is associated with mucosal intestinalization in the early stage of gastric cancer. Virchows Arch. 2011;458:717–24. doi: 10.1007/s00428-011-1086-x. [DOI] [PubMed] [Google Scholar]
- Park SY, Kim HS, Hong EK, Kim WH. Expression of cytokeratins 7 and 20 in primary carcinomas of the stomach and colorectum and their value in the differential diagnosis of metastatic carcinomas to the ovary. Hum Pathol. 2002;33:1078–85. doi: 10.1053/hupa.2002.129422. [DOI] [PubMed] [Google Scholar]
- Sentani K, Matsuda M, Oue N, et al. Clinicopathological significance of MMP-7, laminin gamma2 and EGFR expression at the invasive front of gastric carcinoma. Gastric Cancer. 2014;17:412–22. doi: 10.1007/s10120-013-0302-6. [DOI] [PubMed] [Google Scholar]
- Velculescu VE, Zhang L, Vogelstein B, Kinzler KW. Serial analysis of gene expression. Science. 1995;270:484–7. doi: 10.1126/science.270.5235.484. [DOI] [PubMed] [Google Scholar]
- Ferguson DA, Muenster MR, Zang Q, et al. Selective identification of secreted and transmembrane breast cancer markers using Escherichia coli ampicillin secretion trap. Cancer Res. 2005;65:8209–17. doi: 10.1158/0008-5472.CAN-04-3726. [DOI] [PubMed] [Google Scholar]
- Oue N, Hamai Y, Mitani Y, et al. Gene expression profile of gastric carcinoma: identification of genes and tags potentially involved in invasion, metastasis, and carcinogenesis by serial analysis of gene expression. Cancer Res. 2004;64:2397–405. doi: 10.1158/0008-5472.can-03-3514. [DOI] [PubMed] [Google Scholar]
- Berndorff D, Gessner R, Kreft B, et al. Liver-intestine cadherin: molecular cloning and characterization of a novel Ca(2+)-dependent cell adhesion molecule expressed in liver and intestine. J Cell Biol. 1994;125:1353–69. doi: 10.1083/jcb.125.6.1353. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kreft B, Berndorff D, Bottinger A, et al. LI-cadherin-mediated cell–cell adhesion does not require cytoplasmic interactions. J Cell Biol. 1997;136:1109–21. doi: 10.1083/jcb.136.5.1109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ito R, Oue N, Yoshida K, et al. Clinicopathological significant and prognostic influence of cadherin-17 expression in gastric cancer. Virchows Arch. 2005;447:717–22. doi: 10.1007/s00428-005-0015-2. [DOI] [PubMed] [Google Scholar]
- Grotzinger C, Kneifel J, Patschan D, et al. LI-cadherin: a marker of gastric metaplasia and neoplasia. Gut. 2001;49:73–81. doi: 10.1136/gut.49.1.73. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sakamoto N, Oue N, Sentani K, et al. Liver-intestine cadherin induction by epidermal growth factor receptor is associated with intestinal differentiation of gastric cancer. Cancer Sci. 2012;103:1744–50. doi: 10.1111/j.1349-7006.2012.02353.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hartupee JC, Zhang H, Bonaldo MF, Soares MB, Dieckgraefe BK. Isolation and characterization of a cDNA encoding a novel member of the human regenerating protein family: Reg IV. Biochim Biophys Acta. 2001;1518:287–93. doi: 10.1016/s0167-4781(00)00284-0. [DOI] [PubMed] [Google Scholar]
- Bishnupuri KS, Luo Q, Murmu N, Houchen CW, Anant S, Dieckgraefe BK. Reg IV activates the epidermal growth factor receptor/Akt/AP-1 signaling pathway in colon adenocarcinomas. Gastroenterology. 2006;130:137–49. doi: 10.1053/j.gastro.2005.10.001. [DOI] [PubMed] [Google Scholar]
- Mitani Y, Oue N, Matsumura S, et al. Reg IV is a serum biomarker for gastric cancer patients and predicts response to 5-fluorouracil-based chemotherapy. Oncogene. 2007;26:4383–93. doi: 10.1038/sj.onc.1210215. [DOI] [PubMed] [Google Scholar]
- Katsuno Y, Ehata S, Yashiro M, Yanagihara K, Hirakawa K, Miyazono K. Coordinated expression of REG4 and aldehyde dehydrogenase 1 regulating tumourigenic capacity of diffuse-type gastric carcinoma-initiating cells is inhibited by TGF-β. J Pathol. 2012;228:391–404. doi: 10.1002/path.4020. [DOI] [PubMed] [Google Scholar]
- Takehara A, Eguchi H, Ohigashi H, et al. Novel tumor marker REG4 detected in serum of patients with resectable pancreatic cancer and feasibility for antibody therapy targeting REG4. Cancer Sci. 2006;97:1191–7. doi: 10.1111/j.1349-7006.2006.00297.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nakata K, Nagai E, Ohuchida K, et al. REG4 is associated with carcinogenesis in the ‘intestinal’ pathway of intraductal papillary mucinous neoplasms. Mod Pathol. 2009;22:460–8. doi: 10.1038/modpathol.2008.205. [DOI] [PubMed] [Google Scholar]
- Oue N, Kuniyasu H, Noguchi T, et al. Serum concentration of Reg IV in patients with colorectal cancer: overexpression and high serum levels of Reg IV are associated with liver metastasis. Oncology. 2007;72:371–80. doi: 10.1159/000113147. [DOI] [PubMed] [Google Scholar]
- Ohara S, Oue N, Matsubara A, et al. Reg IV is an independent prognostic factor for relapse in patients with clinically localized prostate cancer. Cancer Sci. 2008;99:1570–7. doi: 10.1111/j.1349-7006.2008.00846.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sasahira T, Oue N, Kirita T, et al. Reg IV expression is associated with cell growth and prognosis of adenoid cystic carcinoma in the salivary gland. Histopathology. 2008;53:667–75. doi: 10.1111/j.1365-2559.2008.03188.x. [DOI] [PubMed] [Google Scholar]
- Tamura H, Ohtsuka M, Washiro M, et al. Reg IV expression and clinicopathologic features of gallbladder carcinoma. Hum Pathol. 2009;40:1686–92. doi: 10.1016/j.humpath.2009.06.001. [DOI] [PubMed] [Google Scholar]
- Heiskala K, Arola J, Heiskala M, Andersson LC. Expression of Reg IV and Hath1 in neuroendocrine neoplasms. Histol Histopathol. 2010;25:63–72. doi: 10.14670/HH-25.63. [DOI] [PubMed] [Google Scholar]
- Zhang J, Liu WL, Tang DC, et al. Identification and characterization of a novel member of olfactomedin-related protein family, hGC-1, expressed during myeloid lineage development. Gene. 2002;283:83–93. doi: 10.1016/s0378-1119(01)00763-6. [DOI] [PubMed] [Google Scholar]
- van der Flier LG, Haegebarth A, Stange DE, van de Wetering M, Clevers H. OLFM4 is a robust marker for stem cells in human intestine and marks a subset of colorectal cancer cells. Gastroenterology. 2009;137:15–7. doi: 10.1053/j.gastro.2009.05.035. [DOI] [PubMed] [Google Scholar]
- Oue N, Sentani K, Noguchi T, et al. Serum olfactomedin 4 (GW112, hGC-1) in combination with Reg IV is a highly sensitive biomarker for gastric cancer patients. Int J Cancer. 2009;125:2383–92. doi: 10.1002/ijc.24624. [DOI] [PubMed] [Google Scholar]
- Qiu W, Wang X, Leibowitz B, et al. Chemoprevention by nonsteroidal anti-inflammatory drugs eliminates oncogenic intestinal stem cells via SMAC-dependent apoptosis. Proc Natl Acad Sci U S A. 2010;107:20027–32. doi: 10.1073/pnas.1010430107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reynolds A, Wharton N, Parris A, et al. Canonical Wnt signals combined with suppressed TGFbeta/BMP pathways promote renewal of the native human colonic epithelium. Gut. 2014;63:610–21. doi: 10.1136/gutjnl-2012-304067. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu W, Yan M, Liu Y, et al. Olfactomedin 4 down-regulates innate immunity against Helicobacter pylori infection. Proc Natl Acad Sci U S A. 2010;107:11056–61. doi: 10.1073/pnas.1001269107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schuijers J, van der Flier LG, van Es J, Clevers H. Robust cre-mediated recombination in small intestinal stem cells utilizing the olfm4 locus. Stem Cell Reports. 2014;3:234–41. doi: 10.1016/j.stemcr.2014.05.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu W, Liu Y, Zhu J, Wright E, Ding I, Rodgers GP. Reduced hGC-1 protein expression is associated with malignant progression of colon carcinoma. Clin Cancer Res. 2008;14:1041–9. doi: 10.1158/1078-0432.CCR-07-4125. [DOI] [PubMed] [Google Scholar]
- Seko N, Oue N, Noguchi T, et al. Olfactomedin 4 (GW112, hGC-1) is an independent prognostic marker for survival in patients with colorectal cancer. Exp Ther Med. 2010;1:73–8. doi: 10.3892/etm_00000013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Duan C, Liu X, Liang S, et al. Oestrogen receptor-mediated expression of Olfactomedin 4 regulates the progression of endometrial adenocarcinoma. J Cell Mol Med. 2014;18:863–74. doi: 10.1111/jcmm.12232. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sentani K, Oue N, Naito Y, et al. Upregulation of HOXA10 in gastric cancer with the intestinal mucin phenotype: reduction during tumor progression and favorable prognosis. Carcinogenesis. 2012;33:1081–8. doi: 10.1093/carcin/bgs121. [DOI] [PubMed] [Google Scholar]
- McGinnis W, Krumlauf R. Homeobox genes and axial patterning. Cell. 1992;68:283–302. doi: 10.1016/0092-8674(92)90471-n. [DOI] [PubMed] [Google Scholar]
- Li B, Cao X, Weng C, et al. HoxA10 induces proliferation in human prostate carcinoma PC-3 cell line. Cell Biochem Biophys. 2014;70:1363–8. doi: 10.1007/s12013-014-0065-7. [DOI] [PubMed] [Google Scholar]
- Calvo R, West J, Franklin W, et al. Altered HOX and WNT7A expression in human lung cancer. Proc Natl Acad Sci U S A. 2000;97:12776–81. doi: 10.1073/pnas.97.23.12776. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jiang Y, Chu Y, Tang W, Wan Y, Zhang L, Cheng W. Transcription factor WT1 and promoter CpG hypomethylation coactivate HOXA10 expression in ovarian cancer. Curr Pharm Des. 2014;20:1647–54. doi: 10.2174/13816128113199990545. [DOI] [PubMed] [Google Scholar]
- Bromleigh VC, Freedman LP. p21 is a transcriptional target of HOXA10 in differentiating myelomonocytic cells. Genes Dev. 2000;14:2581–6. doi: 10.1101/gad.817100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yoshida H, Broaddus R, Cheng W, Xie S, Naora H. Deregulation of the HOXA10 homeobox gene in endometrial carcinoma: role in epithelial–mesenchymal transition. Cancer Res. 2006;66:889–97. doi: 10.1158/0008-5472.CAN-05-2828. [DOI] [PubMed] [Google Scholar]
- Anami K, Oue N, Noguchi T, et al. Search for transmembrane protein in gastric cancer by the Escherichia coli ampicillin secretion trap: expression of DSC2 in gastric cancer with intestinal phenotype. J Pathol. 2010;221:275–84. doi: 10.1002/path.2717. [DOI] [PubMed] [Google Scholar]
- Stokes DL. Desmosomes from a structural perspective. Curr Opin Cell Biol. 2007;19:565–71. doi: 10.1016/j.ceb.2007.09.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yin T, Green KJ. Regulation of desmosome assembly and adhesion. Semin Cell Dev Biol. 2004;15:665–77. doi: 10.1016/j.semcdb.2004.09.005. [DOI] [PubMed] [Google Scholar]
- Anami K, Oue N, Noguchi T, et al. TSPAN8, identified by Escherichia coli ampicillin secretion trap, is associated with cell growth and invasion in gastric cancer. Gastric Cancer. doi: 10.1007/s10120-015-0478-z. [Epub ahead of print] [DOI] [PubMed] [Google Scholar]
- Zoller M. Tetraspanins: push and pull in suppressing and promoting metastasis. Nat Rev Cancer. 2009;9:40–55. doi: 10.1038/nrc2543. [DOI] [PubMed] [Google Scholar]
- Nazarenko I, Rana S, Baumann A, et al. Cell surface tetraspanin Tspan8 contributes to molecular pathways of exosome-induced endothelial cell activation. Cancer Res. 2010;70:1668–78. doi: 10.1158/0008-5472.CAN-09-2470. [DOI] [PubMed] [Google Scholar]
- Oo HZ, Sentani K, Sakamoto N, et al. Identification of novel transmembrane proteins in scirrhous-type gastric cancer by the Escherichia coli ampicillin secretion trap (CAST) method: TM9SF3 participates in tumor invasion and serves as a prognostic factor. Pathobiology. 2014;81:138–48. doi: 10.1159/000357821. [DOI] [PubMed] [Google Scholar]
- Pruvot B, Laurens V, Salvadori F, Solary E, Pichon L, Chluba J. Comparative analysis of nonaspanin protein sequences and expression studies in zebrafish. Immunogenetics. 2010;62:681–99. doi: 10.1007/s00251-010-0472-x. [DOI] [PubMed] [Google Scholar]
- Cornillon S, Pech E, Benghezal M, et al. Phg1p is a nine-transmembrane protein superfamily member involved in dictyostelium adhesion and phagocytosis. J Biol Chem. 2000;275:34287–92. doi: 10.1074/jbc.M006725200. [DOI] [PubMed] [Google Scholar]
- Sanada Y, Oue N, Mitani Y, Yoshida K, Nakayama H, Yasui W. Down-regulation of the claudin-18 gene, identified through serial analysis of gene expression data analysis, in gastric cancer with an intestinal phenotype. J Pathol. 2006;208:633–42. doi: 10.1002/path.1922. [DOI] [PubMed] [Google Scholar]
- Tsukita S, Furuse M, Itoh M. Multifunctional strands in tight junctions. Nat Rev Mol Cell Biol. 2001;2:285–93. doi: 10.1038/35067088. [DOI] [PubMed] [Google Scholar]
- Niimi T, Nagashima K, Ward JM, et al. claudin-18, a novel downstream target gene for the T/EBP/NKX2.1 homeodomain transcription factor, encodes lung- and stomach-specific isoforms through alternative splicing. Mol Cell Biol. 2001;21:7380–90. doi: 10.1128/MCB.21.21.7380-7390.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hayashi D, Tamura A, Tanaka H, et al. Deficiency of claudin-18 causes paracellular H+ leakage, up-regulation of interleukin-1beta, and atrophic gastritis in mice. Gastroenterology. 2012;142:292–304. doi: 10.1053/j.gastro.2011.10.040. [DOI] [PubMed] [Google Scholar]
- Cancer Genome Atlas Research Network. Comprehensive molecular characterization of gastric adenocarcinoma. Nature. 2014;513:202–9. doi: 10.1038/nature13480. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Woll S, Schlitter AM, Dhaene K, et al. Claudin 18.2 is a target for IMAB362 antibody in pancreatic neoplasms. Int J Cancer. 2014;134:731–9. doi: 10.1002/ijc.28400. [DOI] [PubMed] [Google Scholar]
- Silberg DG, Swain GP, Suh ER, Traber PG. Cdx1 and cdx2 expression during intestinal development. Gastroenterology. 2000;119:961–71. doi: 10.1053/gast.2000.18142. [DOI] [PubMed] [Google Scholar]
- Silberg DG, Sullivan J, Kang E, et al. Cdx2 ectopic expression induces gastric intestinal metaplasia in transgenic mice. Gastroenterology. 2002;122:689–96. doi: 10.1053/gast.2002.31902. [DOI] [PubMed] [Google Scholar]
- Park ET, Gum JR, Kakar S, Kwon SW, Deng G, Kim YS. Aberrant expression of SOX2 upregulates MUC5AC gastric foveolar mucin in mucinous cancers of the colorectum and related lesions. Int J Cancer. 2008;122:1253–60. doi: 10.1002/ijc.23225. [DOI] [PubMed] [Google Scholar]
- Tani Y, Akiyama Y, Fukamachi H, Yanagihara K, Yuasa Y. Transcription factor SOX2 up-regulates stomach-specific pepsinogen A gene expression. J Cancer Res Clin Oncol. 2007;133:263–9. doi: 10.1007/s00432-006-0165-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Benahmed F, Gross I, Gaunt SJ, et al. Multiple regulatory regions control the complex expression pattern of the mouse Cdx2 homeobox gene. Gastroenterology. 2008;135:1238–47. doi: 10.1053/j.gastro.2008.06.045. 47.e1–3. [DOI] [PubMed] [Google Scholar]
- Hinoi T, Lucas PC, Kuick R, Hanash S, Cho KR, Fearon ER. CDX2 regulates liver intestine-cadherin expression in normal and malignant colon epithelium and intestinal metaplasia. Gastroenterology. 2002;123:1565–77. doi: 10.1053/gast.2002.36598. [DOI] [PubMed] [Google Scholar]
- Naito Y, Oue N, Hinoi T, et al. Reg IV is a direct target of intestinal transcriptional factor CDX2 in gastric cancer. PLoS ONE. 2012;7:e47545. doi: 10.1371/journal.pone.0047545. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Takakura Y, Hinoi T, Oue N, et al. CDX2 regulates multidrug resistance 1 gene expression in malignant intestinal epithelium. Cancer Res. 2010;70:6767–78. doi: 10.1158/0008-5472.CAN-09-4701. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kakiuchi M, Nishizawa T, Ueda H, et al. Recurrent gain-of-function mutations of RHOA in diffuse-type gastric carcinoma. Nat Genet. 2014;46:583–7. doi: 10.1038/ng.2984. [DOI] [PubMed] [Google Scholar]
- Forbes SA, Beare D, Gunasekaran P, et al. COSMIC: exploring the world’s knowledge of somatic mutations in human cancer. Nucleic Acids Res. 2015;43:D805–11. doi: 10.1093/nar/gku1075. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang K, Yuen ST, Xu J, et al. Whole-genome sequencing and comprehensive molecular profiling identify new driver mutations in gastric cancer. Nat Genet. 2014;46:573–82. doi: 10.1038/ng.2983. [DOI] [PubMed] [Google Scholar]
- Uozaki H, Barua RR, Minhua S, et al. Transcriptional factor typing with SOX2, HNF4aP1, and CDX2 closely relates to tumor invasion and Epstein–Barr virus status in gastric cancer. Int J Clin Exp Pathol. 2011;4:230–40. [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Fig. S1. Schematic outline of the Serial Analysis of Gene Expression (SAGE) method.
Fig. S2. Schematic outline of the Escherichia coli ampicillin secretion trap (CAST) method.