

Abstract
The history of ketamine and phencyclidine from their development as potential clinical anaesthetics through drugs of abuse and animal models of schizophrenia to potential rapidly acting antidepressants is reviewed. The discovery in 1983 of the NMDA receptor antagonist property of ketamine and phencyclidine was a key step to understanding their pharmacology, including their psychotomimetic effects in man. This review describes the historical context and the course of that discovery and its expansion into other hallucinatory drugs. The relevance of these findings to modern hypotheses of schizophrenia and the implications for drug discovery are reviewed. The findings of the rapidly acting antidepressant effects of ketamine in man are discussed in relation to other glutamatergic mechanisms.
Tables of Links
| TARGETS | |
|---|---|
| GPCRsa | Ligand-gated ion channelsb |
| κ receptor | AMPA receptors |
| μ receptor | GluN2A |
| ACh receptors (muscarinic) | GluN2B |
| Cannabinoid receptors | GluN2C |
| D2 receptor | GluN2D |
| Metabotrophic glutamate receptors | Kainate receptors |
| Enzymesd | NMDA receptors |
| Cholinesterases | Nicotinic ACh receptors |
| GAD-67 | |
| GSK-3 | Ion channelc |
| mTOR | HCN1 |
| PKB (Akt) |
| LIGANDS | ||
|---|---|---|
| 5-HT | Dopamine | Morphine |
| ACh | Ethylketocyclazocine | Naloxone |
| AMPA | HA-966 | NMDA |
| Amphetamine | Ifenprodil | Noradrenaline |
| Bicuculline | Kainate | Pentazocine |
| Chlorpromazine | Ketamine (CI-581) | Phencyclidine |
| Cyclazocine | Levorphanol | Pregnenolone |
| D-AP5 | LSD | Quisqualate |
| Dextromethorphan | Memantine | U50488H |
| Dizocilpine (MK-801) |
These Tables list key protein targets and ligands in this article which are hyperlinked to corresponding entries in http://www.guidetopharmacology.org, the common portal for data from the IUPHAR/BPS Guide to PHARMACOLOGY (Pawson et al., 2014) and are permanently archived in the Concise Guide to PHARMACOLOGY 2013/14
Alexander et al., 2013a,b,c,d,,,).
Preamble
In April 1981, Nabil Anis and David Lodge showed for the first time that ketamine was a selective antagonist of the NMDA subtype of glutamate receptor. Unlike the discovery of bicuculline as a GABA antagonist, reported by Graham Johnston in an earlier publication in this series of historical reviews (Johnston, 2013), the editors of Nature were not impressed, the finding ‘not being of sufficient general interest’. Fortunately, the British Journal of Pharmacology took a different view and the resulting paper has now over 1000 citations (Anis et al., 1983). Both before and since this discovery, ketamine and its congener, phencyclidine, have captured the interest of clinicians, basic scientists and sections of the general public. This review will attempt to describe some of this history and bring the reader up to date with the latest twists of this fascinating story in the context of these drugs as NMDA receptor antagonists.
Discovery of phencyclidine and ketamine: early observations
About 27 years earlier, phencyclidine had been synthesized by chemists at Parke Davis Company but not published for some 10 years (Maddox et al., 1965). Initial pharmacology, however, was described by Chen et al. (1959) who noted lack of sensation, hyperlocomotion, ataxia and catalepsy in rats and pigeons. They ascribed these to monoaminergic mechanisms and showed partial reversal by the neuroleptic chlorpromazine (Chen et al., 1959). At about the same time, Ed Domino commenced a fuller study of the neuropharmacology of phencyclidine (Domino, 1964). In rats, he described a ‘drunken state’ with increased locomotor activity leading to ataxia and catatonia at higher doses. Dogs also showed a similar state called ‘canine delirium, although in monkeys a more satisfactory anaesthetic state was achieved but with some catatonia’ (Domino, 1964). Such catatonia had been observed by one of the present authors in the mid-1960s, as a veterinarian using phencyclidine to ‘immobilise’ monkeys for chest radiography for tuberculosis (D. Lodge, unpublished).
Earlier, Greifenstein and co-workers had examined the anaesthetic utility of phencyclidine in man (Greifenstein et al., 1958; Meyer et al., 1959); it proved to be safe with good analgesia but with alarming signs of delirium or psychosis in the recovery stage including a state of centrally mediated sensory deprivation, investigated later in detail by Domino and colleagues (see Domino and Luby, 2012). Although phencyclidine continued to be used as a veterinary anaesthetic particularly for primates, the severe emergence phenomena prevented its further use in man. Parke Davis chemists made many analogues of phencyclidine using rats, dogs and monkeys to find compounds with shorter duration and less potential for delirium; this culminated in ketamine (CI-581; McCarthy et al., 1965), which became a successful human and veterinary anaesthetic. Its properties differed from the standard barbiturate and gaseous anaesthetics of the time, ketamine being a good analgesic, poor muscle relaxant and weak sympathomimetic with a good margin of safety (White and Holtzman, 1982; White et al., 1982; personal observations). Although emergence phenomena were less severe than with phencyclidine (Domino et al., 1965), they have nevertheless limited its use mostly to paediatric and geriatric anaesthesia where safety is the prime consideration (but see Rappaport et al., 2015 for current discussion). It also maintains a particular place on the battlefield where analgesia and safety are paramount. Interestingly in 1982, ketamine became difficult to obtain for our own electrophysiological studies, as all supplies were prioritized to the Falkland Islands!
Psychotomimetic effects: early observations
The schizophrenia-like effects of phencyclidine, and later of ketamine, were known from the outset and indeed phencyclidine was called ‘a new schizophrenomimetic drug’ in the title of Luby’s first study in 1959 (Luby et al., 1959). These effects of phencyclidine and ketamine were a focus of research from the early 1960s. From comparisons made with other psychogens, such as lysergic acid diethylamide (LSD) and mescaline, and from procedures such as sensory isolation and sleep deprivation, it was concluded that phencyclidine-induced psychosis was most akin to schizophrenia (Luby et al., 1959; Domino et al., 1965); a conclusion that has been reiterated with further supportive evidence for both phencyclidine and ketamine over the subsequent half century (e.g. Javitt and Zukin, 1991; Krystal et al., 1994; Olney and Farber, 1995; Malhotra et al., 1996; Newcomer et al., 1999; Jentsch and Roth, 1999; Geyer et al., 2001; Lahti et al., 2001; Tsai and Coyle, 2002; Morris et al., 2005; Ehrlichman et al., 2009; Saunders et al., 2012; Moghaddam and Krystal, 2012; Gil-da-Costa et al., 2013); this is particularly so following repeated abuse (Javitt and Zukin, 1991; Jentsch and Roth, 1999). Negative, positive, cognitive and electrophysiological signs of schizophrenia are all reported to be replicated to some extent by phencyclidine and ketamine (Javitt and Zukin, 1991; Krystal et al., 1994). Indeed, so much so that in the late 1970s and early 1980s, pharmaceutical companies and academics, hoping for antipsychotic agents as therapy for schizophrenia, invested heavily in searching for antagonists of phencyclidine (Kamenka et al., 1983).
Underlying these overt behavioural effects of phencyclidine and ketamine were changes in the CNS both in the EEG and in glucose metabolism.
The EEG showed bizarre and complex changes in rhythmical activity, reduced evoked potentials and inhibition of some pathways in a dose-dependent manner (Domino, 1964). The EEG was dominated by increased delta and reduced alpha waves in the thalamo-neocortical pathways, accompanied by theta bursts in the hippocampal formation, suggesting a dissociation of sensory and limbic systems (Miyasaka and Domino, 1968). For example, in one of the earliest studies, Rodin et al. (1959), cited by Domino and Luby (2012), reported ‘rather profound slowing of the EEG with pronounced theta activity’. With ketamine, ‘abolishing of the alpha waves and induction of theta activity were the most consistent and typical EEG effects’ (Domino et al., 1965). These complex disruptions of the EEG signal by ketamine and phencyclidine and their relationship to desynchronization seen in schizophrenia has remained a subject of research for the ensuing 50 years (see Table 2012).
Table 1.
Some shared GABAergic features between schizophrenia and the effect of arylcyclohexylamines
| Feature | Schizophrenia | Phencyclidine/ketamine |
|---|---|---|
| Reduction in markers of cortical GABAergic interneurones (e.g. PV, GAD-67) | Akbarian et al., 1995; Beasley and Reynolds, 1997; Woo et al., 1998; Guidotti et al., 2000; Hashimoto et al., 2003; Reynolds et al., 2004 | Cochran et al., 2003; Keilhoff et al., 2004; Reynolds et al., 2004; Cunningham et al., 2006; Behrens et al., 2007; Morrow et al., 2007; Broberg et al., 2008; Wang et al., 2008 |
| Decreased chandelier synapses and up-regulation of GABA receptors | Woo et al., 1998; Volk et al., 2002 | Pierri et al., 1999; Abe et al., 2000; Wang et al., 2008; du Bois et al., 2009; Beninger et al., 2010 |
| Disinhibition increasing glutamate release with risk of excitotoxicity via AMPA receptors | Harrison and Weinberger, 2005; Coyle, 2012; Kraguljac et al., 2013; Poels et al., 2014 | Olney and Farber, 1995; Moghaddam et al., 1997; Lorrain et al., 2003 |
| EEG changes, including desynchronization and alterations in gamma and lower frequencies | Friston, 1998; Cho et al., 2006; Roopun et al., 2008; Minzenberg et al., 2010; Uhlhaas and Singer, 2010 | Cunningham et al., 2006; Ehrlichman et al., 2009; McNally et al., 2011; Troyano-Rodriguez et al., 2014; Moran et al., 2015 |
| Deficit in mismatch negativity and pre-pulse inhibition | Javitt et al., 1996; Umbricht and Krljes, 2005; Braff et al., 2007 | Javitt et al., 1996; Heekeren et al., 2008; Amann et al., 2010; Gil-da-Costa et al., 2013 |
Desynchronization was also implied from brain metabolism studies using the 2-deoxyglucose method which showed increases in limbic areas, particularly hippocampus, cingulate gyrus and entorhinal cortex, but decreases in more afferent-related areas, including inferior colliculus and sensory motor cortex (Nelson et al., 1980; Crosby et al., 1982; Hammer et al., 1982).
Abuse of phencyclidine and ketamine: early observations
Despite apparently unpleasant sensory changes described above, both phencyclidine and ketamine soon became substances of abuse. Commonly known as ‘Angel Dust’ or ‘PeaCePill’, phencyclidine appeared on the streets of West Coast USA by 1965, reaching epidemic proportions, such that in Washington, DC, there were more psychiatric admissions due to its overdose than for alcohol abuse and schizophrenia combined (Luisada and Reddick, 1975; Petersen and Stillman, 1978), symptoms often being mistaken for schizophrenia. Such psychotomimetic episodes may persist for several weeks following its last use (Luby et al., 1959; Domino et al., 1965), and even after long abstinence, some abusers progressed to schizophrenia (Luisada, 1978). Fortunately, phencyclidine is no longer so widely abused (Moeller et al., 2008). Ketamine, however, has become a common recreational drug, particularly prevalent at rave parties and music venues throughout the world, where it appears as Special K, Kit Kat, Cat Valium, etc. (Nutt et al., 2007; Morris and Wallach, 2014). Presumably in human endophenotypes that encompass the ‘Special K’ abusers, the ‘out-of-body’, ‘dissociative’ experiences and hallucinations, possibly related to facilitation of dopaminergic systems (Wise, 1996), outweigh the drug-induced disabilities and risks of near-death experiences in the ‘K-hole’.
Putative mechanisms of action: early considerations
Because dopamine is implicated in the aetiology of schizophrenia and in drug abuse, the interaction of phencyclidine with dopaminergic and other neurotransmitter systems was widely investigated (see Johnson, 1983). Phencyclidine inhibits dopamine uptake and enhances dopamine release (Garey and Heath, 1976; Smith et al., 1977; Vickroy and Johnson, 1980; 1982,), and the behavioural actions of phencyclidine could be reversed by functional antagonists of dopaminergic transmission (Freeman and Bunney, 1984). Similar effects of phencyclidine on noradrenaline and 5-hydroxytryptamine (5-HT) transport were also reported (Taube et al., 1975; Garey and Heath, 1976; Smith et al., 1977). The cholinergic system also provided potential targets for phencyclidine, which has modest affinity for nicotinic and muscarinic receptors as well as for cholinesterases (Kloog et al., 1977; Vincent et al., 1978; Albuquerque et al., 1980). Other proposed mechanisms for the action of phencyclidine included block of potassium channels (Albuquerque et al., 1981; Blaustein and Ickowicz, 1983) and μ-opioid receptors (Vincent et al., 1978). These somewhat confusing multifarious putative mechanisms received two new challenges between 1979 and 1983.
Firstly, the description of specific sub-micromolar binding of tritiated phencyclidine to brain membranes (Vincent et al., 1979; Zukin and Zukin, 1979) gave a new impetus to the field but also a major challenge; binding of phencyclidine was displaced by other arylcyclohexylamines with a rank order similar to their behavioural potencies. However, although more than 40 compounds were examined, including monoamines, acetylcholine, amino acid neurotransmitters, analogues and antagonists including neuroleptic and other centrally active drugs, there was no clear link to the above-mentioned putative mechanisms. The one chemically unrelated compound that displaced phencyclidine binding was the benzomorphan, SKF10 047 or N-allyl-normetazocine (Zukin and Zukin, 1979), a protypical sigma opiate (Martin et al., 1976). In the following few years, cross displacement in binding studies in rat and human tissue between arylcyclohexylamines, sigma opiates (e.g. SKF10 047 and cyclazocine), dioxolanes (e.g. dexoxadrol) and morphinans (e.g. dextrorphan) suggested a common site of action of these psychotomimetics (Quirion et al., 1981; Zukin and Zukin, 1981; Hampton et al., 1982; Zukin, 1982; Sircar and Zukin, 1983; Murray and Leid, 1984; Zukin et al., 1984).
Secondly, drug discrimination tests for phencyclidine showed that other arylcyclohexylamines, and many of the above benzomorphans, generalized to the subjective phencyclidine cue (Holtzman, 1980). In the following few years, cross generalization within the arylcyclohexylamines, sigma opiates, dioxolanes and morphinans in rats, pigeons and monkeys was well documented (Holtzman, 1980; 1982,; Brady and Balster, 1981; Herling et al., 1981; Shannon, 1981; 1982a,b; 1983,,,; Brady et al., 1982a,b,; White and Holtzman, 1982). There was a good correlation between displacement of phencyclidine binding and generalization to the phencyclidine cue (Javitt and Zukin, 1991). Importantly, other classes of psychoactive drugs not displacing phencyclidine binding did not generalize to the phencyclidine cue.
In common with arylcyclohexylamines, the drugs that displaced phencyclidine binding and that generalized to the phencyclidine cue had known psychotomimetic properties in man. These included the sigma opiates (e.g. cyclazocine and SKF10.047) (Keats and Telford, 1964; Haertzen, 1970; Martin et al., 1976), dioxolanes (e.g. etoxadrol) (Wilson et al., 1970; Frederickson et al., 1976) and morphinans (e.g. dextrorphan and dextromethorphan) (Isbell and Fraser, 1953; Jasinski, 1979).
The above, and much more, was detailed at an excellent meeting in Montpellier in 1983 organized by Ed Domino, a pioneering leader in all matters related to phencyclidine (Kamenka et al., 1983).
So by the early 1980s, it seemed likely that this phencyclidine binding site mediated the psychotomimetic effects in man of arylcyclohexylamines and the above structurally diverse drugs. The challenge was to find the central synaptic correlate.
Ketamine and spinal reflexes
In 1981, Lodge and Anis were studying the effects on synaptic transmission of short acting i.v. anaesthetics (e.g. methohexitone, alphaxolone/alphadolone and ketamine) on spinal reflexes (Lodge and Anis, 1982; 1984,). To their surprise, ketamine (2.5–10 mg·kg−1, i.v.) preferentially reduced polysynaptic, rather than monosynaptic, reflexes but was without effects on synaptic inhibition and dorsal root potentials. By contrast and as expected, barbiturate and steroid anaesthetics reduced both mono- and polysynaptic reflexes by enhancing GABAergic inhibition (Lodge and Anis, 1984). Not known to these authors at the outset, this sensitivity of spinal polysynaptic reflexes to ketamine had been reported some 10 years earlier (Tang and Schroeder, 1973; Chen and Chow, 1975). Subsequently, when ketamine was administered directly into the region of single neurones in the spinal cord, polysynaptic excitations were selectively reduced (Anis et al., 1983). This was a key experiment because it demonstrated that the action of ketamine was close to the soma and dendrites of the postsynaptic cell and most likely at the excitatory synapse itself. It had been recently demonstrated that ketamine did not affect the uptake or release of amino acids (Minchin, 1981), leaving the possibility that it acted on the postsynaptic receptors.
At this time it had just been established that, in the mammalian CNS, glutamate acted on three subtypes of ionotropic receptor, namely NMDA, quisqualate and kainate receptors (Davies and Watkins, 1979; McLennan and Lodge, 1979; Watkins and Evans, 1981). Because quisqualate also acts at metabotropic glutamate receptors and because AMPA is a selective ionotropic receptor agonist (Krogsgaard-Larsen et al., 1980), the name quisqualate was replaced by AMPA (Watkins and Evans, 1981). Importantly, in relation to the effects of ketamine, studies with competitive antagonists had already demonstrated that postsynaptic NMDA receptors mediated spinal polysynaptic excitation (Biscoe et al., 1977; Lodge et al., 1978; Davies and Watkins, 1979).
The NMDA receptor, ketamine and phencyclidine
To determine whether the action of ketamine was on postsynaptic receptors, a series of experiments were commenced using the microelectrophoresis technique to eject ketamine and some of the above excitatory amino acid agonists into the vicinity of single spinal neurones. Local application of ketamine reduced or abolished responses to N-methyl-DL-aspartate (NMDA) and left responses to quisqualate and kainate largely unchanged (Figure 1; Anis et al., 1983). This exciting discovery was quickly followed by the same experiment with intravenous ketamine (2.5–10 mg·kg−1). In parallel with the reduction of polysynaptic reflexes, responses induced by exogenous activation of the NMDA receptor were reduced within 5 min of injection and recovered slowly over the next 30–120 min. Interestingly, nicotinic responses of Renshaw cells were also reduced but to a lesser extent (Anis et al., 1983). Ketamine is a racemic mixture, the ((S)+) enantiomer being approximately two to four times more potent than the ((R)-) enantiomer as an anaesthetic (Ryder et al., 1978; Franks and Lieb, 1994), as an analgesic (Klepstad et al., 1990), as an inhibitor of phencyclidine binding (Zukin, 1982; Murray and Leid, 1984) and in drug discrimination studies (Brady and Balster, 1982). The (+) rather than the (−) enantiomer is psychotomimetic in man (Vollenweider et al., 1997). As an NMDA receptor antagonist, (+)-ketamine was about 3 times more potent than the (−) enantiomer but only 1.5 times more potent as a nicotinic antagonist (Lodge and Anis, 1982; Lodge et al., 1982).
Figure 1.
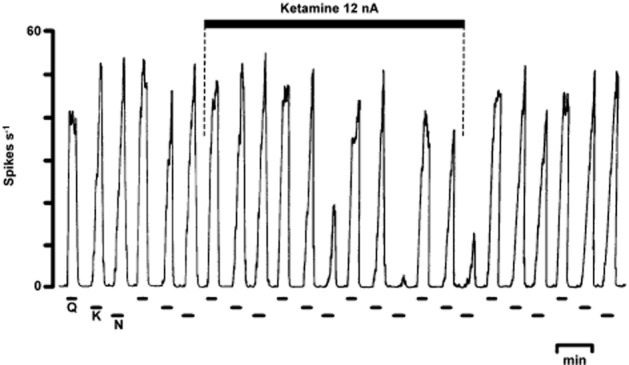
Original record from Anis et al. (1983) showing one of the earliest experiments demonstrating the selectivity of ketamine for NMDA. The recording shows the firing rate of a spinal neurone from a pentobarbitone-anaesthetized cat in response to the electrophoretic ejection of quisqualate, kainate and N-methyl-DL-aspartate. The co-ejection of ketamine almost abolishes the latter with only minor effects on responses to the non-NMDA receptor agonists and recovery occurs within 5 min of stopping the ketamine ejection. Other details are in Anis et al. (1983).
Was the NMDA receptor antagonism of ketamine also present in ‘illegal’ phencyclidine? An old veterinary supply of injectable phencyclidine and some phencyclidine powder was obtained from the Metropolitan Police, and checked for purity! Compared with ketamine, phencyclidine proved to be similarly selective as an NMDA receptor antagonist but approximately 10 times more potent and longer lasting following either local or systemic administration (Lodge and Anis, 1982; Anis et al., 1983). Phencyclidine, 0.2–0.5 mg·kg−1 i.v., reduced responses to NMDA for several hours. Interestingly, thienylcyclohexylpiperidine was even more potent than phencyclidine (Lodge et al., 1988c). The enantiomers of both 4- and 5-methyl substituted phencyclidine showed notable stereoselectivity as NMDA receptor antagonists; the (+) enantiomers being 5–10 times more potent and more selective than the (−) enantiomers. By contrast, as antagonists of acetylcholine on Renshaw cells, there was only minor stereoselectivity (Berry et al., 1983; Lacey and Henderson, 1986).
The NMDA receptor and other psychotomimetics
Next, other structurally different psychotomimetics mentioned above (see Figure 2) were examined. The sigma opiates, α-cyclazocine and SKF10 047, proved to be selective antagonists of NMDA, cyclazocine being approximately twice as potent as SKF10 047; the (−) enantiomer of cyclazocine and the (+) enantiomer of SKF10 047 were somewhat more potent than their respective enantiomers (Berry et al., 1984b; Lodge and Anis, 1984; Lodge and Berry, 1984; Lodge et al., 1984). By contrast, two protypical κ-receptor opiates, ethylketocyclazocine and U50 488H, and the μ-receptor ligands, morphine and naloxone, were without significant or selective effects on responses to NMDA. Furthermore, naloxone did not prevent the effects of cyclazocine or ketamine as NMDA receptor antagonists (Anis et al., 1983; Lodge et al., 1988b). Interestingly at concentrations that reduced responses to NMDA, the (+), but not the (−) enantiomer, of SKF10 047 enhanced responses to acetylcholine on Renshaw cells (Berry et al., 1984a). Etoxadrol and dexoxadrol, but not levoxadrol, potently blocked responses to NMDA (Berry et al., 1984b) as did dextrorphan and dextromethorphan but not the former’s enantiomer, levorphanol (Church et al., 1985).
Figure 2.
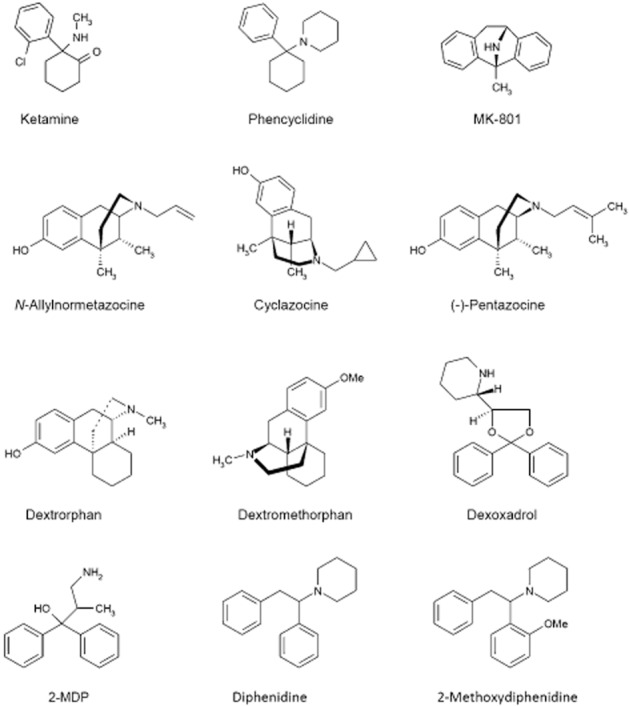
Chemical structure of some channel-blocking NMDA antagonists.
Three new structurally different classes of phencyclidine-like compounds were published during the course of the above studies. Benz(f)isoquinolines (Mendelsohn et al., 1984), 2-methyl-3,3-diphenyl-3-propanolamine (2-MDP; Tang et al., 1984) and dizocilpine (MK-801) shared binding and behavioural properties with phencyclidine (Wong et al., 1986; Sircar et al., 1987; Koek et al., 1988). All three showed selective NMDA receptor antagonism: the NMDA receptor antagonist profile within the benz(f)isoquinolines correlated with their phencyclidine-like properties (Berry and Lodge, 1984) as did the threefold greater potency of (−)-, versus (+)-, 2-MDP (Blake et al., 1986). MK-801 was by far the most potent compound as an NMDA receptor antagonist (Davies et al., 1988), as a displacer of phencyclidine binding (Wong et al., 1986) and in phencyclidine discrimination studies (Willetts and Balster, 1988). Subsequently, β-cyclazocine was shown to approach MK-801 in potency and stereoselectivity as an NMDA receptor antagonist (Church et al., 1991), but with less selectivity in drug discrimination (Slifer and Balster, 1988) and binding studies (Todd et al., 1990).
Relation to biochemical and behavioural effects of psychotomimetics
The bulk of these early observations was made using the technique of microelectrophoresis to administer both glutamate receptor agonists and potential antagonists into the region of single spinal neurones. This technique, although providing excellent information concerning selectivity between agonists, does not allow accurate assessment of drug concentrations and hence should only be used as a guide to potency. Nevertheless, when comparing activity of compounds with similar physicochemical characteristics and particularly between enantiomers, potency can be reasonably estimated. Figure 3 plots the relative potency of pairs of enantiomers as NMDA receptor antagonists versus their potency in binding assays (Figure 3A) and drug discrimination assays (Figure 3B). It can be seen that there is close agreement between stereoselectivity in these various assays.
Figure 3.
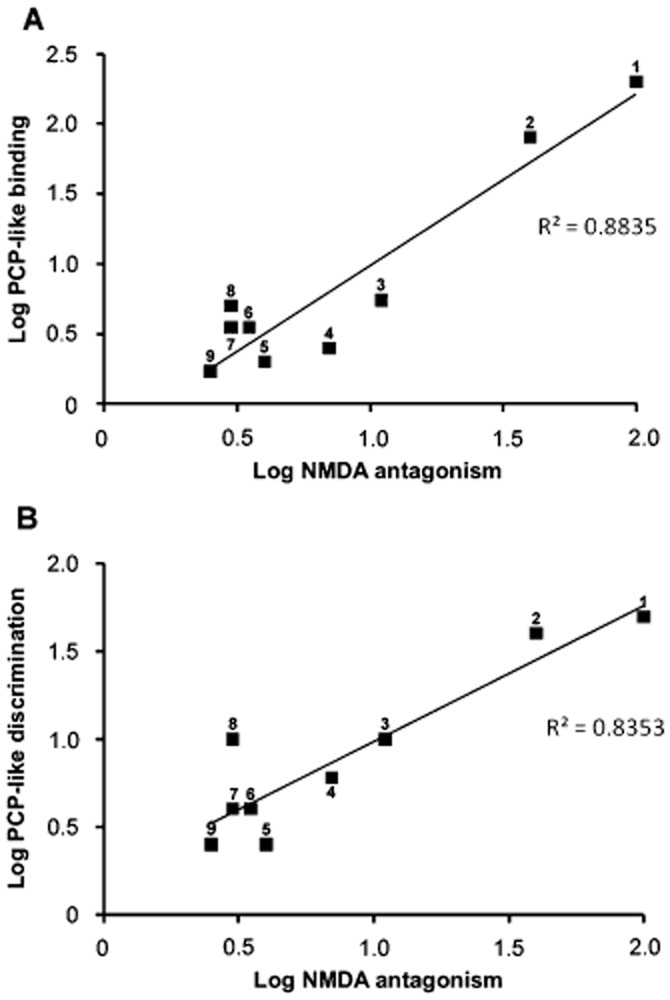
Stereoselective potency between pairs of isomers as NMDA receptor antagonists versus their stereoselectivity in phencyclidine-like binding assays (A) and in drug discrimination assays (B). Data are compiled from references cited in the text, showing potency comparisons between isomers (e.g. Berry et al., 1984a,b,; Church et al., 1985; 1991,). Each numbered point represents the stereoselectivity of the following pairs of isomers: 1 = (−) versus (+) s-cyclazocine: 2 = dexoxadrol versus levoxadrol: 3 = (+) versus (−) 3-methylphencyclidine: 4 = dextrorphan versus levorphanol: 5 = (+) versus (−) SKF10,047: 6 = (−) versus (+) α-cyclazocine: 7 = (+) versus (−) ketamine: 8 = (−) versus (+) 2-MDP: 9 = (−) versus (+) pentazocine β.
Recognizing the limitations of potency values from the above in vivo experiments, in vitro studies were used to provide alternative potency values and to study the nature of the interaction between psychotomimetics and the NMDA receptor. Using grease-seal preparations of frog and rat spinal cords and of rat cerebro-cortical slices, many of the above compounds were tested against depolarizations induced by agonists of NMDA, AMPA and kainate receptors. Effects in vitro were very comparable with those in vivo; ketamine, phencyclidine and other related psychotomimetics selectively antagonized depolarization by NMDA but not by quisqualate/AMPA and kainate (Martin and Lodge, 1985; 1988,; Davies et al., 1988; Aram et al., 1989). Because agonist concentration–response curves were not shifted in parallel, potencies were assessed as IC50 values versus 40 μM NMDA. Such in vitro potencies correlated closely with those from phencyclidine binding assays (Figure 4A).
Figure 4.
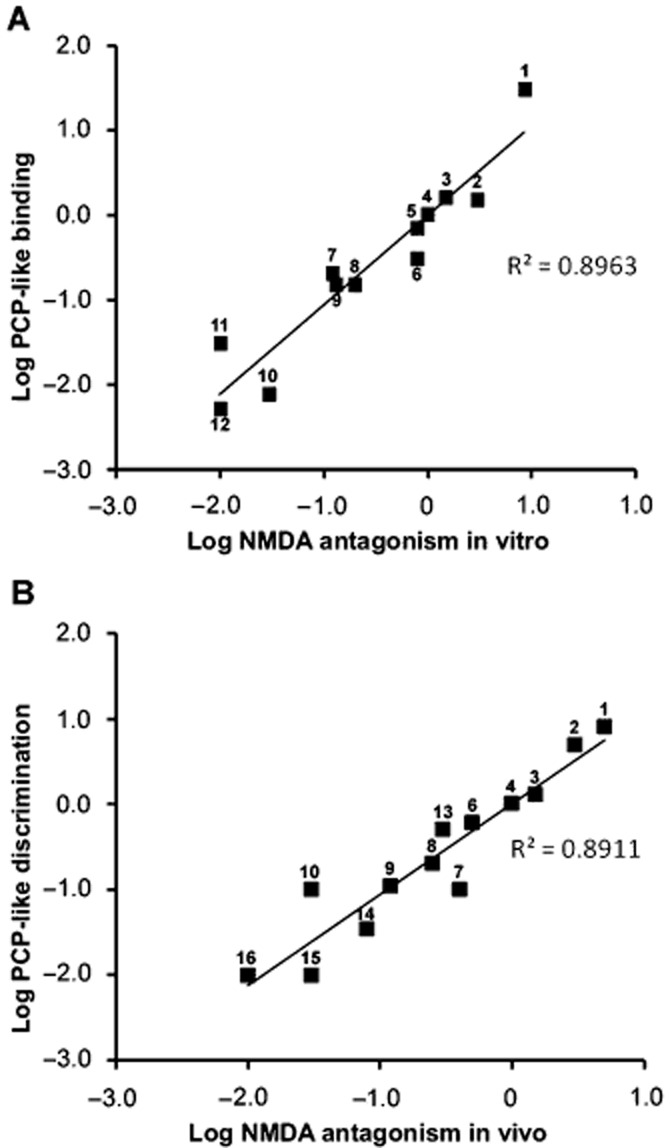
Comparison of potency of structurally diverse compounds, expressed relative to phencyclidine, as NMDA receptor antagonists in vitro versus relative potency in binding assays (A) and as NMDA receptor antagonists in vivo versus relative potency in drug discrimination assays (B). Data are compiled from references cited in the text; potencies were compared in the same animals and often on the same neurones (e.g. Berry et al., 1984b; Church and Lodge, 1990). Each numbered point represents a single compound: 1 = MK-801: 2 = (−)-β-cyclazocine: 3 = thienylcyclohexylpiperidine: 4 = phencyclidine: 5 = LY154045: 6 = α-cyclazocine: 7 = dextrorphan: 8 = SKF10,047: 9 = ketamine: 10 = (+)-s-cyclazocine: 11 = pentazocine: 12 = LY154005: 13 = dexoxadrol: 14 = dextromethorphan: 15 = levorphanol: 16 = levoxadrol: Note that the compounds plotted in (A) are not identical to those in (B).
The potencies of compounds following in vivo administration can more appropriately be compared with potencies in behavioural assays. Figure 4B shows that there is a good correlation between NMDA receptor antagonism and phencyclidine-like discrimination assays. Such a relationship between compounds of diverse structures speaks to the central role of NMDA receptor antagonism in the common behavioural features of these psychotomimetics.
Correlating potency of compounds as NMDA receptor antagonists with dysphoric and psychotomimetic potency in man is more tenuous. Doses producing such effects, for example, 0.05 mg·kg−1 MK-801 (Erowid Experience Vaults, 2013), 0.1 mg·kg−1 phencyclidine (Domino and Luby, 2012) and 2 mg·kg−1 ketamine (White and Holtzman, 1982; White et al., 1982), overlap with NMDA receptor blocking doses in the rat (Lodge et al., 1988a). Similarly, the doses in man of 0.05 mg·kg−1 cyclazocine (Martin et al., 1965), 0.3 mg·kg−1 dexoxadrol (Lasagna and Pearson, 1965), 1 mg·kg−1 dextrorphan (Isbell and Fraser, 1953) and 2 mg·kg−1 pentazocine (Jasinski et al., 1970) are not dissimilar from those in the rat that antagonize NMDA receptors (Lodge et al., 1988a).
In conclusion, these various potency comparisons strongly suggested that the phencyclidine binding site is linked to the NMDA receptor and that NMDA receptor antagonism underlies the behavioural effects of these structurally diverse compounds in laboratory animals and presumably in man. As the effects in man are reminiscent of schizophrenia (see above), it seemed likely that NMDA receptor dysfunction could be linked to the disease process (Lodge and Berry, 1984).
Mechanism of action
Ion channels coupled to NMDA receptors (Figure 5) are activated by synaptically released L-glutamate, are permeable to calcium (Ascher and Nowak, 1986; MacDermott et al., 1986), require co-activation with glycine or D-serine (Johnson and Ascher, 1987) and are inhibited by magnesium ions (Evans et al., 1977) in a voltage-dependent manner (Mayer et al., 1984; Nowak et al., 1984). Competitive interaction at the glutamate recognition site of the NMDA receptor depends on the amino and two carboxylic moieties of glutamate (Curtis and Watkins, 1960). No such charge distribution can be assigned to the above psychotomimetic structures (Figure 2). The non-parallel shift in agonist concentration–response curves by higher concentrations of these NMDA receptor antagonists (Lodge and Johnston, 1985; Snell and Johnson, 1985) and the non-additive effects of combinations of these drugs with competitive antagonists (Harrison and Simmonds, 1985; Lodge and Johnston, 1985; Martin and Lodge, 1985) confirmed that these compounds act at a site distinct from the agonist recognition site (Figure 5A). This was quickly followed by electrophysiological demonstrations that the action of ketamine was use- and voltage-dependent (MacDonald et al., 1987) and further confirmed with other compounds (Wong et al., 1986; Davies et al., 1988; Huettner and Bean, 1988). Similarly, binding to the phencyclidine site was shown to be agonist dependent (Fagg, 1987; Foster and Wong, 1987; Loo et al., 1987; Kloog et al., 1988). The above findings are all consistent with open channel block of the NMDA receptor by the arylcyclohexylamines and related compounds (Figure 5B).
Figure 5.
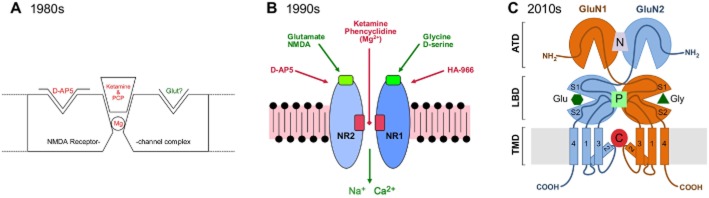
Historical development of crude models of NMDA receptor-channel complexes with putative sites of action of key compounds. (A) and (B) were used by Lodge at conferences during the 1980s and 1990s, respectively, reflecting his knowledge of structure function early in each decade. (C) reflects current ideas showing the relationship of the amino terminal (ATD), ligand binding (LBD) and transmembrane (TMD) domains with putative binding sites for negative allosteric (N; e.g. ifenprodil), positive allosteric (P; e.g. pregnenolone) and channel-blocking (C; e.g. ketamine) compounds, kindly provided by David Jane and www.hellobio.com.
Demonstration of its channel-blocking site curtailed search for displacers of phencyclidine binding as antipsychotics but brought the NMDA receptor to the forefront of schizophrenia research. Inasmuch as phencyclidine and ketamine model the disease symptoms, NMDA receptor dysfunction is the cornerstone of the glutamate hypothesis of schizophrenia.
This use and voltage dependency of the open channel block, defined by on and off kinetics, varies considerably between individual compounds within the various compound classes. For example, ketamine and memantine are less use and more voltage dependent than MK-801, with phencyclidine and dextrorphan being intermediate (Davies et al., 1988; MacDonald et al., 1991; Parsons et al., 1993; 1995,; Gilling et al., 2009). It should be noted that most pharmacodynamic measures are conducted in vitro at temperatures below the normal physiological ranges, and therefore likely to slow the on and off kinetics and hence exaggerate apparent use dependency (Davies et al., 1988).
These pharmacodynamic properties all contribute towards the different profiles of these uncompetitive NMDA receptor antagonists. They do not, however, appear to explain why ketamine is a dissociative anaesthetic and analgesic whereas memantine has found a niche for the neurodegeneration of Alzheimer’s disease, as both compounds have similar potencies and pharmacodynamics (Gilling et al., 2009; Emnett et al., 2013). Interestingly, the pharmacokinetics of memantine are closer to those of MK-801 than of ketamine, and it seems likely that this property, along with the diversity of their effects on other voltage- and receptor-coupled channels rather than their pharmacodynamics per se, determines their different therapeutic profiles (Gilling et al., 2009).
Subunit selectivity
Further likely influences on the different behavioural profiles of non-competitive NMDA receptor antagonists are potency differences between subtypes of NMDA receptors; excitatory NMDA receptors are tetrameric combinations usually comprising of two GluN1 and two GluN2 subunits, with four possible genes (A–D) coding for the latter (Monaghan et al., 2012). The binding site for uncompetitive NMDA receptor antagonists, and for magnesium, is deep within the channel near the asparagine residues (the N-site) of the pore-lining M2 loops of the GluN1 and GluN2 subunits (Burnashev et al., 1992; Kashiwagi et al., 2002; Kotermanski and Johnson, 2009; see Figure 5C). In earlier studies, MK-801 was suggested to be more potent on NMDA receptors containing the GluN2A and GluN2B subunits (Yamakura et al., 1993; Bresink et al., 1996). More recently, potencies of phencyclidine-like compounds were shown to vary between the four GluN2 subunits. For example, (+)-ketamine, memantine and phencyclidine were respectively about nine, five and four times less potent on GluN1/GluN2A than on GluN1/GluN2B-D, whereas dextrorphan showed a small preference for GluN2C subunits and (+)-MK-801 was almost equipotent across all four subtypes (Dravid et al., 2007). Interestingly (+)-ketamine was less potent than the racemate on GluN1/GluN2A and GluN1/GluN2B receptors, but about 1.5 times more potent on GluN1/GluN2C and GluN1/GluN2D heteromers. If these data on receptors expressed in oocytes reflect the in vivo situation, they suggest the possibility that the behavioural effects of ketamine are more likely to be mediated by GluN2C- or GluN2D-containing receptors rather than GluN2A or GluN2B subunits. In agreement with this is the observation that phencyclidine-induced locomotor activity is significantly less in GluN2D knockout mice than in wild-type mice (Hagino et al., 2010; Yamamoto et al., 2013). Against this are the important roles for GluN2A and GluN2B subunits in synaptic activity in higher centres (Bartlett et al., 2007; Anastasio et al., 2009; McNally et al., 2011; Kocsis, 2012; Hanson et al., 2013). Furthermore, GluN2B antagonists have psychotomimetic properties in man (Preskorn et al., 2008) and generalize to the phencyclidine cue in rats and monkeys (Chaperon et al., 2003; De Vry and Jentzsch, 2003; Nicholson et al., 2007).
The early therapeutic promise
Developed as an anaesthetic, it was the analgesic property of ketamine that attracted most early clinical interest. Although its use is limited by emergent psychotomimetic episodes, ketamine is still widely used in acute, neuropathic and palliative care cases (Chizh, 2007; Teasell et al., 2010; Niesters et al., 2013; Persson, 2013). Because it blocks the NMDA receptor-mediated plasticity that results in ‘wind-up’ of spinal neurones in response to repetitive nociceptive stimuli (Davies and Lodge, 1987), ketamine is useful in reducing the allodynia and hyperalgesia of neuropathic pain.
Coincident with the discovery of ketamine as an NMDA receptor antagonist, the anticonvulsant and neuroprotective properties of competitive NMDA receptor antagonists (Croucher et al., 1982; Simon et al., 1984) indicated other potential therapeutic considerations for non-competitive NMDA receptor antagonists. Indeed, the rank order of potency as neuroprotectants in vitro, MK-801 > phencyclidine > ketamine > (±)SKF10 047 = cyclazocine > pentazocine (Olney et al., 1986), closely parallels their potency as NMDA receptor antagonists. Evidence for such a correlation was supported by the in vivo demonstration of the effectiveness of ketamine and MK-801 in global ischaemia in both gerbils, rats and cats (Church et al., 1988; Gill et al., 1988; Marcoux et al., 1988; Park et al., 1988a,b,). Such neuroprotective potential was however confounded by other data from Olney’s group, which showed that, when administered alone at doses somewhat above those required for neuroprotection, MK-801, phencyclidine and ketamine induced neuronal vacuoles and cell death in the rat cerebral cortex (Olney et al., 1989). The extension of such neurotoxic effects of dissociative anaesthetics to large increases in apoptosis in fetal and neonatal rat brains following repeated dosing (Ikonomidou et al., 1999; Wang et al., 2001) has further questioned the use of ketamine as an analgesic/anaesthetic agent, particularly in paediatric practice (Rappaport et al., 2015).
These considerations, together with sympathomimetic and emergent psychotomimetic effects, have largely prevented these drugs from becoming established as neuroprotective agents (reviewed by Hudetz and Pagel, 2010), although there is still some enthusiasm for their candidature (e.g. Gakuba et al., 2011). Interestingly, the reduced pH of ischaemic and hypoxic tissues would enhance the potency of these channel blockers (Dravid et al., 2007).
Similarly, despite early enthusiasm for uncompetitive NMDA receptor antagonists as potential anticonvulsants (Aram et al., 1989; Chapman and Meldrum, 1989; Tricklebank et al., 1989), their use in epilepsy is largely limited to refractory status epilepticus where ketamine is relatively effective and safe (Gaspard et al., 2013; Zeiler et al., 2014).
Ketamine, phencyclidine, schizophrenia and NMDA receptor antagonism
As stated above, behavioural effects of phencyclidine and ketamine in man replicate to a large degree the positive, negative and cognitive symptoms of schizophrenia and exacerbate symptoms in schizophrenic patients (Javitt and Zukin, 1991; Krystal et al., 1994; Malhotra et al., 1996; 1997,; Lahti et al., 2001). So, at face value, the actions of ketamine and phencyclidine could provide insights into the aetiology and treatment of schizophrenia. Indeed, arguments have been made that these drugs can be used to provide animal models of certain aspects of the disease (e.g. Jentsch and Roth, 1999; Cunningham et al., 2006; Buccafusco and Terry, 2009). To think, however, that a single drug can recreate in animals a disease with such complex aetiologies as that of human schizophrenia is clearly naive, but as a way of testing potential therapies against some of the symptoms of schizophrenia, the concept is more plausible (but see Gilmour et al., 2012). Nevertheless, the possibility of modelling particular symptom domains is likely to be a more feasible goal using genetic and environmental perturbations concerned with the aetiology of schizophrenia (Pratt et al., 2012; Lustig et al., 2013; Foussias et al., 2015; Young and Geyer, 2015).
One oft stated concern in relation to implying a purely glutamatergic hypothesis is that many of the above non-competitive NMDA receptor antagonists act on molecular substrates other than NMDA receptors (see above) and that these make important contributions to their schizophrenia-like symptoms (Lodge and Johnson, 1990). However, in addition to the work cited above, many other studies have tried and failed to correlate activity on non-NMDA receptor substrates with behavioural effects. Some useful comparisons follow: Lacey and Henderson in 1986 showed that phencyclidine was 10 times more potent as an NMDA receptor antagonist (IC50 0.4 μM) compared with effects on noradrenaline, enkephalin and action potentials and had no effect on membrane currents at up to 100 μM; furthermore, only NMDA receptor antagonism showed stereoselectivity with methyl-phencyclidine (Lacey and Henderson, 1986). In a recent review, the potency of ketamine at sigma, κ, muscarinic, cannabinoid and GABA receptors was shown to be two orders of magnitude less than at NMDA receptors; only as a stimulant of D2 dopamine receptors was submicromolar equipotency reported (Frohlich and Van Horn, 2014). Tellingly, in contrast to their psychotomimetic effects, ketamine had similar and phencyclidine greater potency relative to MK-801 as activators of D2 receptors (Seeman et al., 2009)! Potassium channels and monoaminergic systems are a target for phencyclidine and ketamine but require dissociative concentrations in the tens of micromolar (and see Fletcher et al., 1989), except for HCN1 channels (Chen et al., 2009), but the latter appear not to be sensitive to 80 μM MK-801 (Tokay et al., 2009). Ketamine blocks ganglionic, α3β4 and α7 nicotinic receptors with an IC50 of about 1.4, 3.1 and 20 μM respectively (Friederich et al., 2000; Yamakura et al., 2000; Moaddel et al., 2013). Memantine and MK-801 were somewhat less potent as α3β4 channel blockers (Buisson and Bertrand, 1998); rank orders that do not correlate with the above behavioural effects. It is however important to note that nicotine and/or nicotinic agonists may nevertheless help in the treatment of schizophrenia (e.g. Buccafusco and Terry, 2009).
Although it is undoubtedly true that some of the non-NMDA receptor actions contribute to the behavioural profile of each dissociative compound, there are compelling arguments supporting NMDA receptor antagonism as the likely major contributor to the psychotomimesis:
There is a good correlation between potency of drugs, including between enantiomers, as NMDA receptor antagonists and their psychotomimetic effects in laboratory animals and to a limited extent in man (see above and Figures 4).
Competitive NMDA receptor antagonists share some behavioural effects with phencyclidine-like compounds, including drug discrimination cues (Koek et al., 1987; see Willetts et al., 1990 for critical review), but more importantly have psychotomimetic effects in man (Grotta et al., 1995; Herrling, 1997; Davis et al., 2000; Muir, 2006).
NMDA receptor antibodies have been implicated in the aetiology of some schizophrenias, although causality is unproven (Deakin et al., 2014; Kayser and Dalmau, 2014; for reviews see Coutinho et al., 2014; Pearlman and Najjar, 2014).
Genetic risk factors for schizophrenia include several genes that impinge directly or indirectly on NMDA receptor function (Harrison and Weinberger, 2005; Gilmour et al., 2012; Labrie et al., 2012a; Weickert et al., 2013; Harrison, 2015).
Genetic manipulations in mice affecting NMDA receptor activity induce (Belforte et al., 2010; Labrie et al., 2012b; Wei et al., 2014; Born et al., 2015; Takagi et al., 2015) or reduce (Hagino et al., 2010; Yamamoto et al., 2013) aspects of a schizophrenia-like phenotype.
Some clinical trials with drugs designed to up-regulate NMDA receptor function have had positive outcomes (reviewed in Coyle, 2012). Other manipulations of glutamatergic transmission show promise in animal models and are being assessed therapeutically (reviewed in Javitt, 2012 and Dunlop and Brandon, 2015).
Agonists of group II metabotropic glutamate receptors reduce hyperlocomotion induced by NMDA receptor antagonists in rats (Moghaddam and Adams, 1998; Cartmell et al., 1999), ketamine-induced cognitive deficits in man (Krystal et al., 2005), and positive and negative symptoms of schizophrenia in man (Patil et al., 2007). Subsequent trials have however failed to confirm efficacy in schizophrenics; explanations of this discrepancy will be of value for future therapies.
Implications for schizophrenia
So what can studies with NMDA receptor antagonism by ketamine and phencyclidine teach us about schizophrenia and potential therapeutic strategies? Other than the overt behavioural effects, are there other features of the disease spectrum that are replicated by the non-competitive NMDA receptor antagonists mentioned above? Two such features to be considered are changes in dopaminergic transmission and the neuropathology of schizophrenia.
Thus, one key question is the compatibility of the glutamate and dopamine hypotheses. The main tenets of the dopamine hypothesis of schizophrenia are that dopamine, particularly D2, receptor antagonists are antipsychotic and that amphetamine by increasing dopamine release can mimic aspects of the disease. Disruptions in dopaminergic activity especially in prefrontal cortical and striatal areas are a hallmark of schizophrenia and aspects of these complex changes are replicated by acute and chronic ketamine or phencyclidine dosing (Javitt and Zukin, 1991; Jentsch and Roth, 1999; Javitt, 2007; Coyle, 2012; Moghaddam and Krystal, 2012). In particular, systemic phencyclidine and SKF10 047 indirectly excite midbrain dopaminergic neurones (Freeman and Bunney, 1984), acute ketamine treatment increases dopamine release in prefrontal cortex rather than striatum (Verma and Moghaddam, 1996) whereas chronic treatment with phencyclidine increases striatal and reduces prefrontal dopamine (Jentsch et al., 1997; 1998,), more consistent with the hypofrontality of schizophrenia. The complex neurotransmitter feedback loops between prefrontal cortex, hippocampus, ventral tegmentum and striatum suggest that, for example, reduced glutamatergic activity in the hippocampus can result in the above dopaminergic pattern of schizophrenia (Goto and Grace, 2007; Howes et al., 2015) and the reduced glutamate activity in the frontal areas of this disease (Marsman et al., 2013).
The second key question, the ability of dissociative anaesthetics to mimic aspects of the disease pathology, is covered in the next section.
Neuropathology underlying cognitive dysfunction
The potential of NMDA receptor antagonists to disrupt normal synaptic development of neuronal circuitry follows from the seminal observation by Collingridge et al. (1983). Using the newly discovered competitive NMDA receptor antagonist, D-2-amino-5-phosphonovalerate (D-APV; D-AP5; Davies et al., 1981), the requirement for NMDA receptor activation in plasticity was established in 1983 with the classical demonstration of its role in long-term potentiation (LTP; Collingridge et al., 1983). Coincidentally in the same year, without knowledge of their NMDA receptor antagonist properties, phencyclidine and ketamine were shown to reduce LTP (Stringer and Guyenet, 1983). NMDA receptors not only mediate LTP but also long-term depression (reviewed in Collingridge et al., 2013). Early observations, that local NMDA receptor antagonism disrupted the normal pattern of synaptic connections in the visual cortex in vivo (Cline et al., 1987; Kleinschmidt et al., 1987), indicated the importance of these receptors in neurodevelopment. This trophic role of NMDA receptors, likely mediated by the entry of calcium through the NMDA receptor channel (Collingridge and Singer, 1990; Cline and Tsien, 1991) during development, is potentially a key aetiological factor predisposing to schizophrenia. For example, during critical phases of development, exposure to ketamine, phencyclidine or other disruptors of NMDA receptor function is likely to result in dysregulation of synapse formation and brain circuitry (see below), characteristics of schizophrenic brain (Harrison, 1999; Snyder and Gao, 2013).
In contrast to established neuroprotective properties of NMDA receptor antagonists (see above), severe acute neuropathology occurs with high doses of dissociative anaesthetics. Thus, Olney and colleagues showed that acute high doses of ketamine, phencyclidine and MK-801 led to neuronal cytotoxicity including that of cortical pyramidal cells and inhibitory interneurones in adult rats, drawing parallels at that time with the neuropathology of schizophrenia (Olney et al., 1989; Olney and Farber, 1995). Similar lesions were also reported following prolonged administration of competitive NMDA receptor antagonists (Ellison, 1994; 1995,).
Chronic administration of lower doses during the perinatal period and into adulthood results in more subtle effects on neurogenesis, cell survival, migration, proliferation and synaptogenesis (Ikonomidou et al., 1999; Wang et al., 2001; Keilhoff et al., 2004; Maeda et al., 2007; Namba et al., 2011; Toriumi et al., 2012; Sabbagh et al., 2013; Musaelyan et al., 2014; Uchida et al., 2014). Such changes are more representative of schizophrenic brains, which have reduced cortical neuropil, disturbed neuronal migration and circuitry but lack severe degenerative pathology (Bogerts, 1997; Jones, 1997; Weinberger, 1997; Harrison, 1999; Catts et al., 2013). Such chronic exposure to dissociative anaesthetics also leads to behavioural correlates of the disease (Jentsch and Roth, 1999; Wang et al., 2001; Egerton et al., 2008; Rodefer et al., 2008; Goetghebeur and Dias, 2009; Amitai and Markou, 2010; Neill et al., 2010).
Importantly, in addition to these similarities to schizophrenia, chronic administration of ketamine and phencyclidine also disrupted metabolic connectivities (Cochran et al., 2003; Dawson et al., 2013) and desynchronized electrical activity between prefrontal cortex and thalamic (Troyano-Rodriguez et al., 2014) and hippocampal (Moran et al., 2015) regions.
There is however compelling evidence in schizophrenia for specific disruption of GABAergic function, which led to the modern GABAergic hypothesis of schizophrenia; down-regulation of fast-spiking parvalbumin (PV)-containing and GAD67-positive GABAergic neuronal networks is proposed as a major contributor to schizophrenia, potentially explaining aspects of glutamatergic and dopaminergic dysregulation (Olney and Farber, 1997; Lewis and Gonzalez-Burgos, 2006; Nakazawa et al., 2012; Lewis, 2014). Synaptic excitation of PV-containing GABAergic interneurones, chandelier and basket cells, depends largely on NMDA receptor activation (Jones and Bühl, 1993; Grunze et al., 1996; Goldberg et al., 2003). This may underlie the intriguing parallels between schizophrenia and dissociative anaesthetics (see Table 2012).
The similarities in Table 2012 suggest that exposure to dissociative anaesthetics during early development and adolescence could contribute to the PV cell damage and lead to schizophrenic symptoms later in life. This, however, is likely to explain a very small minority of cases. Rather than dissociative anaesthetics, other influences, such as oxidative stress, viral infections along with genetic predisposition, are likely causes of GABAergic dysfunction in the majority of schizophrenics (reviewed in Harrison et al., 2012; Nakazawa et al., 2012; Powell et al., 2012; Lewis, 2014).
Thus, to conclude this section, ketamine and phencyclidine offer two ways of mimicking aspects of schizophrenia pathology. Firstly, chronic doses during development and in adulthood result in dysregulation of the synaptic connectivity that mirrors schizophrenic pathology and behaviour. Secondly, in more mature brains, acute and sub-acute doses of ketamine or phencyclidine, by blocking the excitatory drive of PV-GABAergic interneurones and thus disinhibiting hippocampal and cortical networks, results in psychotomimetic activity. Such behaviours will be more marked if development of GABAergic pathways has been compromised during development.
Modern-day recreational use of ketamine and related drugs
Because of its lower potency and shorter duration of action, ‘Special K’ has less severe psychiatric issues than ‘Angel Dust’, but can lead to death in combination with other illicit drugs and/or alcohol and fatal accidents due to disturbed perceptions. The difference between the two drugs has long been recognized by UK and US drug authorities, which classified phencyclidine and ketamine as Class A/Schedule II and Class C/Schedule III drugs respectively. The use of ketamine in the UK alone is likely to be in excess of 100 000 each year (Morgan and Curran, 2012). Ketamine is in the top 6 drugs of abuse for causing social and physical harm (Nutt et al., 2007), being implicated in tens of deaths each year; the major risk of physical harm is a result of ketamine’s dissociative and analgesic properties (see Morgan and Curran, 2012; Office of National Statistics, 2013). In 2014, ketamine was upgraded to Class B in the UK. In addition to psychiatric effects, continued abuse of ketamine can lead to systemic toxicities, including chronic and painful urinary cystitis (Shahani et al., 2007; Chu et al., 2008; Morgan and Curran, 2012). In attempts to avoid such pathology and also legislative strictures, numerous analogues of ketamine and phencyclidine have been made available on the street (Roth et al., 2013; Morris and Wallach, 2014). One of the latest is diphenidine, a diarylethylamine, an old compound (Christiaen, 1924) that disappeared from the scientific literature until patented for neurotoxic shock (Gray and Cheng, 1989). It subsequently reappeared, along with some of its derivatives, as a recreational drug in 2013, coincident with the rescheduling of ketamine! The non-medical uses of this and other NMDA receptor antagonists have been thoroughly reviewed recently (Morris and Wallach, 2014). As with ketamine, diphenidine and its 2-methoxy derivative inhibit NMDA receptor-mediated synaptic events in the CNS (Figure 6). The slow onset of diphenidine (Figure 6) approaches the kinetics of MK-801 in slice preparations (Wong et al., 1986; Davies et al., 1988) and may reflect a greater use dependency or possibly trapping of the antagonist in the NMDA receptor channel compared with ketamine.
Figure 6.
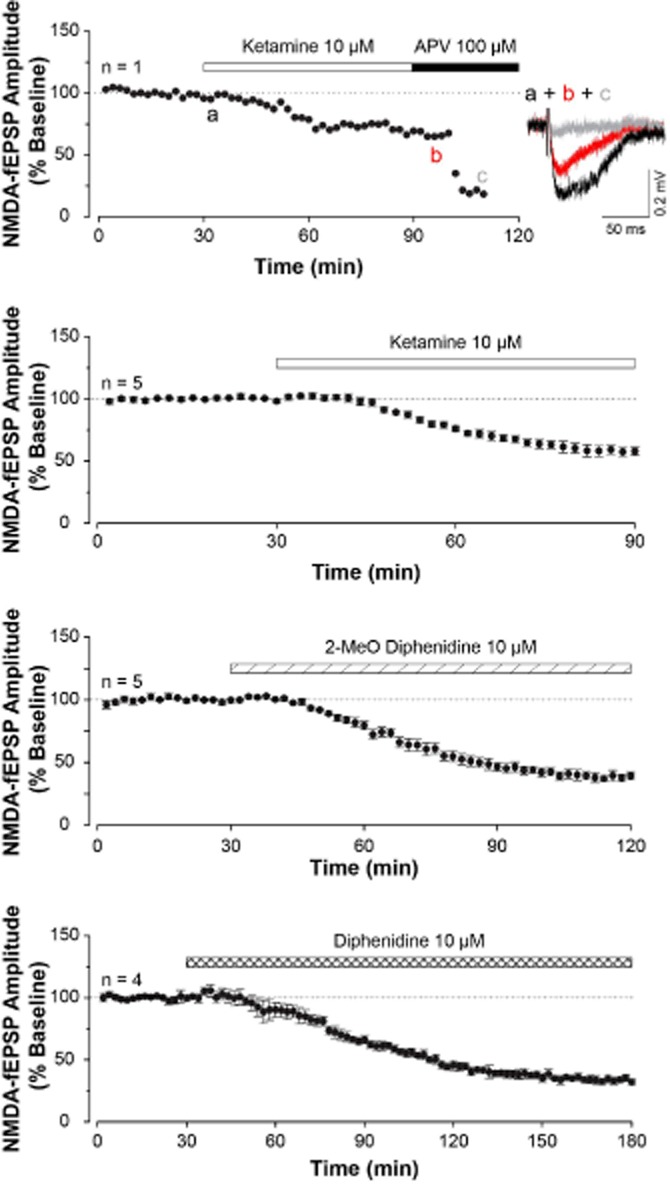
Newer diarylethylamine street drugs, like ketamine, reduce NMDA receptor-mediated synaptic excitation. The graphs plot the amplitude of field EPSPs in the CA1 region of a hippocampal slice following stimulation of the Schaffer collateral input. Slices had been treated with AMPA and GABA receptor antagonists to isolate the NMDA receptor component of the synaptic event. The top graph is a single experiment showing the effect of ketamine 10 μM and D-AP5 100 μM, demonstrating that these were NMDA receptor-mediated responses. The inset shows the raw traces. The subsequent three graphs show pooled data from four or five experiments illustrating NMDA receptor antagonism by ketamine, 2-methoxy-diphenidine and diphenidine. Note the slower time course to reach near plateau reduction of the EPSP. These data suggest that diphenidine is somewhat more potent than 2-methoxydiphenidine, which in turn is more potent than ketamine.
It appears that clandestine synthetic medicinal chemists will always be able to outwit legislation based on known structures because the possibilities of finding novel compounds to modulate NMDA receptor activity are near to infinite! But the consequences of their subsequent misuse can be fatal (Elliott et al., 2015).
Future therapeutic potential of ketamine and related drugs
In addition to the previously considered therapeutic uses of non-competitive NMDA receptor antagonists, the recent exciting discovery of antidepressant-like effects of ketamine in man (Berman et al., 2000; Zarate et al., 2006) has re-awakened interest in these drugs. The background to this discovery is worthy of consideration.
Largely, because depression is treated by drugs that enhance monoaminergic neurotransmission albeit with delayed onset, depression is considered mainly a monoaminergic disorder (Schildkraut and Kety, 1967; Millan, 2004). However, there is a long history suggesting a potential glutamatergic role in the aetiology and treatment of depression (see Alt et al., 2006), including the observation that some antidepressants are NMDA receptor antagonists (Evans and Watkins, 1981; Reynolds and Miller, 1988; Sernagor et al., 1989; Sills and Loo, 1989; Watanabe et al., 1993). Two major observations from Skolnick’s laboratory, however, clearly demonstrated the link between antidepressant activity and glutamate. Firstly, 25 years ago, they made the crucial observation that competitive channel-blocking and glycine-site NMDA receptor antagonists were all effective in animal models of depression (Trullas and Skolnick, 1990). Secondly, positive allosteric potentiators of AMPA receptors were similarly effective in such animal models (Li et al., 2001; 2003,). A third later observation that antagonists of glutamatergic autoreceptors have antidepressant potential (Chaki et al., 2004; Yoshimizu et al., 2006) confirmed the importance of glutamatergic systems to mood disturbances.
The innovative translation to man using ketamine (Berman et al., 2000) and the subsequent double-blind confirmation in treatment-resistant patients (Zarate et al., 2006) gave a massive impetus to the search for glutamatergic, rather than monoaminergic, antidepressants. An interesting feature of these new antidepressant therapies is the rapid onset of the change in mood both in animal models and in man. This feature is particularly valuable for suicidal patients. Unlike the weeks of therapy required for 5-HT (serotonin) and noradrenaline uptake inhibitors, mood is elevated rapidly after single injections of ketamine which provide relief for days and even weeks (see Aan Het Rot et al., 2012; Caddy et al., 2014; Duman, 2014). Although the antidepressant activity of ketamine is not doubted, mild psychotomimetic effects may alert the patients to the administration of drug rather than placebo. The low doses, typically 0.5 mg·kg−1, providing plasma levels of approximately 0.5 μM and presumably similar brain levels, are likely to show preference for GluN2B-, C- and D-, rather than GluN2A- containing NMDA receptors, but with only partial antagonism (Dravid et al., 2007). In addition to NMDA receptor-channel blockers including magnesium (see Serefko et al., 2013; Zarate et al., 2013), glycine-site (Danysz and Parsons, 1998), GluN2B allosteric and gaseous (Preskorn et al., 2008; Lima-Ojeda et al., 2013; Miller et al., 2014; Nagele et al., 2014) NMDA receptor antagonists have shown rapid antidepressant potential in man and/or animal models. Similarly AMPA receptor potentiators (Alt et al., 2006) and group II metabotropic glutamate receptor antagonists (Chaki et al., 2004; Dwyer et al., 2012) have rapid antidepressant effects.
Although these diverse drugs have different modes of action, a functional increase in glutamatergic neurotransmission is hypothesized as the linker. In the case of ketamine and other NMDA receptor antagonists, this is most likely via inhibition of GABAergic pathways (see above). The resulting surge in excitatory transmission is presumably the trigger for changes to long-term plasticity that underlies the depressed mood. Antagonism of AMPA receptors prevents the rapid antidepressant action of ketamine (see Koike and Chaki, 2014). Brief periods of increased excitatory transmission are known to de-potentiate and/or de-depress established plasticity (Collingridge et al., 2010). Brain-derived neurotrophic factor (BDNF), tropomyosin-related kinase B, mammalian target of rapamycin (mTOR), PKB, glycogen synthase kinase-3 (GSK-3) and synapse remodelling are well documented mediators of such processes (Duman et al., 2012; Duman, 2014) and provide further therapeutic targets. However, the ubiquitous nature of these latter targets raises concerns of deleterious on-target secondary actions, including non-neuronal tissues.
Whether ketamine itself, with its potential psychotomimetic and other unwanted side effects, could become an accepted therapy is still under discussion. It is interesting to speculate why other similarly acting NMDA receptor antagonists such as memantine, phencyclidine and MK-801 do not show such robust antidepressant effects. Presumably some combination of NMDA subunit selectivity, pharmacodynamics and pharmacokinetics gives ketamine its advantage. The challenge is now to find and replicate that advantageous combination and to avoid ketamine’s unwanted side effects.
Conclusion
The history of ketamine and phencyclidine as therapeutics has been a roller coaster of optimism and pessimism. Ketamine’s accepted positions as an adjunct in anaesthesia and in pain management and as a dangerous street drug illustrate this dichotomy. The value of ketamine in providing insights into the role of NMDA receptors in brain function, into the understanding of schizophrenia and potentially into new antidepressant drugs has led to the present swell of interest in this enigmatic molecule. Like the past, the future will no doubt contain surprises.
Acknowledgments
Professors KM Johnson, E. Domino, PM Beart and GL Collingridge commented on earlier versions of the manuscript. D. L. wishes to thank all his previous collaborators for providing the data that have been quoted in this review and for funding from the MRC and Wellcome Trust at that time. We also thank Simon Brandt for provision of diphenidine analogues and Andy Doherty and David Jane for help with figures.
Glossary
- 2-MDP
2-methyl-3,3-diphenyl-3-propanolamine
- D-AP5/D-APV
D-2-amino-5-phosphonovalerate
- GAD67
glutamic acid dehydrogenase
- LTP
long-term potentiation
- MK-801
dizocilpine
- PV
parvalbumin
- SKF10,047
N-allyl-normetazocine
Author contributions
D. L. and M. S. M. wrote the first draft and collated the bibliography. D. L. and M. S. M. revised the manuscript. M. S. M. contributed the data in Figure 4.
Conflict of interest
D. L. and M. S. M. have no conflicts of interest to disclose.
Disclosure statement
D. L. and M. S. M. have received contributions to their salaries in previous years from Eli Lilly & Co. M. S. M. is currently at UCL supported by the Wellcome Trust.
References
- Aan Het Rot M, Zarate CA, Charney DS, Mathew SJ. Ketamine for depression: where do we go from here? Biol Psychiatry. 2012;72:537–547. doi: 10.1016/j.biopsych.2012.05.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Abe S, Suzuki T, Ito T, Baba A, Hori T, Kurita H, et al. Differential expression of GABA(A) receptor subunit mRNAs and ligand binding sites in rat brain following phencyclidine administration. Synapse. 2000;38:51–60. doi: 10.1002/1098-2396(200010)38:1<51::AID-SYN6>3.0.CO;2-A. [DOI] [PubMed] [Google Scholar]
- Akbarian S, Huntsman MM, Kim JJ, Tafazzoli A, Potkin SG, Bunney WE, et al. GABAA receptor subunit gene expression in human prefrontal cortex: comparison of schizophrenics and controls. Cereb Cortex. 1995;5:550–560. doi: 10.1093/cercor/5.6.550. [DOI] [PubMed] [Google Scholar]
- Albuquerque EX, Tsai MC, Aronstam RS, Witkop B, Eldefrawi AT, Eldefrawi ME. Phencyclidine interactions with the ionic channel of the acetylcholine receptor and electrogenic membrane. Proc Natl Acad Sci U S A. 1980;77:1224–1228. doi: 10.1073/pnas.77.2.1224. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Albuquerque EX, Aguayo LG, Warnick JE, Weinstein H, Glick SD, Maayani S, et al. The behavioral effects of phencyclidines may be due to their blockade of potassium channels. Proc Natl Acad Sci U S A. 1981;78:7792–7796. doi: 10.1073/pnas.78.12.7792. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alexander SPH, Benson HE, Faccenda E, Pawson AJ, Sharman JL, Spedding M, et al. The Concise Guide to PHARMACOLOGY 2013/14: G protein-coupled receptors. Br J Pharmacol. 2013a;170:1459–1581. doi: 10.1111/bph.12445. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alexander SPH, Benson HE, Faccenda E, Pawson AJ, Sharman JL, Spedding M, et al. The Concise Guide to PHARMACOLOGY 2013/14: ligand-gated ion channels. Br J Pharmacol. 2013b;170:1582–1606. doi: 10.1111/bph.12446. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alexander SPH, Benson HE, Faccenda E, Pawson AJ, Sharman JL, Catterall WA, et al. The concise guide to pharmacology 2013/14: ion channels. Br J Pharmacol. 2013c;170:1607–1651. doi: 10.1111/bph.12447. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alexander SPH, Benson HE, Faccenda E, Pawson AJ, Sharman JL, Spedding M, et al. The Concise Guide to PHARMACOLOGY 2013/14: enzymes. Br J Pharmacol. 2013d;170:1797–1867. doi: 10.1111/bph.12451. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alt A, Nisenbaum ES, Bleakman D, Witkin JM. A role for AMPA receptors in mood disorders. Biochem Pharmacol. 2006;71:1273–1288. doi: 10.1016/j.bcp.2005.12.022. [DOI] [PubMed] [Google Scholar]
- Amann LC, Gandal MJ, Halene TB, Ehrlichman RS, White SL, McCarren HS, et al. Mouse behavioral endophenotypes for schizophrenia. Brain Res Bull. 2010;83:147–161. doi: 10.1016/j.brainresbull.2010.04.008. [DOI] [PubMed] [Google Scholar]
- Amitai N, Markou A. Disruption of performance in the five-choice serial reaction time task induced by administration of N-methyl-D-aspartate receptor antagonists: relevance to cognitive dysfunction in schizophrenia. Biol Psychiatry. 2010;68:5–16. doi: 10.1016/j.biopsych.2010.03.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anastasio NC, Xia Y, O’Connor ZR, Johnson KM. Differential role of N-methyl-D-aspartate receptor subunits 2A and 2B in mediating phencyclidine-induced perinatal neuronal apoptosis and behavioral deficits. Neuroscience. 2009;163:1181–1191. doi: 10.1016/j.neuroscience.2009.07.058. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anis NA, Berry SC, Burton NR, Lodge D. The dissociative anaesthetics, ketamine and phencyclidine, selectively reduce excitation of central mammalian neurones by N-methyl-aspartate. Br J Pharmacol. 1983;79:565–575. doi: 10.1111/j.1476-5381.1983.tb11031.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aram JA, Martin D, Tomczyk M, Zeman S, Millar J, Pohler G, et al. Neocortical epileptogenesis in vitro: studies with N-methyl-D-aspartate, phencyclidine, sigma and dextromethorphan receptor ligands. J Pharmacol Exp Ther. 1989;248:320–328. [PubMed] [Google Scholar]
- Ascher P, Nowak L. A patch-clamp study of excitatory amino acid activated channels. Adv Exp Med Biol. 1986;203:507–511. doi: 10.1007/978-1-4684-7971-3_39. [DOI] [PubMed] [Google Scholar]
- Bartlett TE, Bannister NJ, Collett VJ, Dargan SL, Massey PV, Bortolotto ZA, et al. Differential roles of NR2A and NR2B-containing NMDA receptors in LTP and LTD in the CA1 region of two-week old rat hippocampus. Neuropharmacology. 2007;52:60–70. doi: 10.1016/j.neuropharm.2006.07.013. [DOI] [PubMed] [Google Scholar]
- Beasley CL, Reynolds GP. Parvalbumin-immunoreactive neurons are reduced in the prefrontal cortex of schizophrenics. Schizophr Res. 1997;24:349–355. doi: 10.1016/s0920-9964(96)00122-3. [DOI] [PubMed] [Google Scholar]
- Behrens MM, Ali SS, Dao DN, Lucero J, Shekhtman G, Quick KL, et al. Ketamine-induced loss of phenotype of fast-spiking interneurons is mediated by NADPH-oxidase. Science. 2007;318:1645–1647. doi: 10.1126/science.1148045. [DOI] [PubMed] [Google Scholar]
- Belforte JE, Zsiros V, Sklar ER, Jiang Z, Yu G, Li Y, et al. Postnatal NMDA receptor ablation in corticolimbic interneurons confers schizophrenia-like phenotypes. Nat Neurosci. 2010;13:76–83. doi: 10.1038/nn.2447. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beninger RJ, Beuk J, Banasikowski TJ, Adel M, van Boivin GA, Reynolds JN. Subchronic phencyclidine in rats: alterations in locomotor activity, maze performance, and GABA(A) receptor binding. Behav Pharmacol. 2010;21:1–10. doi: 10.1097/FBP.0b013e3283347091. [DOI] [PubMed] [Google Scholar]
- Berman RM, Cappiello A, Anand A, Oren DA, Heninger GR, Charney DS, et al. Antidepressant effects of ketamine in depressed patients. Biol Psychiatry. 2000;47:351–354. doi: 10.1016/s0006-3223(99)00230-9. [DOI] [PubMed] [Google Scholar]
- Berry SC, Lodge D. Benz(f)isoquinolines as excitatory amino acid antagonists: an indication of their mechanism of action? Biochem Pharmacol. 1984;33:3829–3832. doi: 10.1016/0006-2952(84)90047-9. [DOI] [PubMed] [Google Scholar]
- Berry SC, Burton NR, Anis NA, Lodge D. Stereoselective effects of two phencyclidine derivatives on N-methylaspartate excitation of spinal neurones in the cat and rat. Eur J Pharmacol. 1983;96:261–267. doi: 10.1016/0014-2999(83)90315-1. [DOI] [PubMed] [Google Scholar]
- Berry SC, Anis NA, Lodge D. The effect of the dioxolanes on amino acid induced excitation in the mammalian spinal cord. Brain Res. 1984a;307:85–90. doi: 10.1016/0006-8993(84)90463-3. [DOI] [PubMed] [Google Scholar]
- Berry SC, Dawkins SL, Lodge D. Comparison of sigma- and kappa-opiate receptor ligands as excitatory amino acid antagonists. Br J Pharmacol. 1984b;83:179–185. doi: 10.1111/j.1476-5381.1984.tb10133.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Biscoe TJ, Evans RH, Francis AA, Martin MR, Watkins JC, Davies J, et al. D-alpha-aminoadipate as a selective antagonist of amino acid-induced and synaptic excitation of mammalian spinal neurones. Nature. 1977;270:743–745. doi: 10.1038/270743a0. [DOI] [PubMed] [Google Scholar]
- Blake JC, Davies SN, Church J, Martin D, Lodge D. 2-Methyl-3,3-diphenyl-3-propanolamine (2-MDP) selectively antagonises N-methyl-aspartate (NMA) Pharmacol Biochem Behav. 1986;24:23–25. doi: 10.1016/0091-3057(86)90038-9. [DOI] [PubMed] [Google Scholar]
- Blaustein MP, Ickowicz RK. Phencyclidine in nanomolar concentrations binds to synaptosomes and blocks certain potassium channels. Proc Natl Acad Sci U S A. 1983;80:3855–3859. doi: 10.1073/pnas.80.12.3855. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bogerts B. The temporolimbic system theory of positive schizophrenic symptoms. Schizophr Bull. 1997;23:423–435. doi: 10.1093/schbul/23.3.423. [DOI] [PubMed] [Google Scholar]
- du Bois TM, Deng C, Han M, Newell KA, Huang X-F. Excitatory and inhibitory neurotransmission is chronically altered following perinatal NMDA receptor blockade. Eur Neuropsychopharmacol. 2009;19:256–265. doi: 10.1016/j.euroneuro.2008.12.002. [DOI] [PubMed] [Google Scholar]
- Born G, Grayton HM, Langhorst H, Dudanova I, Rohlmann A, Woodward BW, et al. Genetic targeting of NRXN2 in mice unveils role in excitatory cortical synapse function and social behaviors. Front Synaptic Neurosci. 2015;7:3. doi: 10.3389/fnsyn.2015.00003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brady KT, Balster RL. Discriminative stimulus properties of phencyclidine and five analogues in the squirrel monkey. Pharmacol Biochem Behav. 1981;14:213–218. doi: 10.1016/0091-3057(81)90245-8. [DOI] [PubMed] [Google Scholar]
- Brady KT, Balster RL. Discriminative stimulus properties of ketamine stereoisomers in phencyclidine-trained rats. Pharmacol Biochem Behav. 1982;17:291–295. doi: 10.1016/0091-3057(82)90083-1. [DOI] [PubMed] [Google Scholar]
- Brady KT, Balster RL, May EL. Stereoisomers of N-allylnormetazocine: phencyclidine-like behavioral effects in squirrel monkeys and rats. Science. 1982a;215:178–180. doi: 10.1126/science.6274022. [DOI] [PubMed] [Google Scholar]
- Brady KT, Woolverton WL, Balster RL. Discriminative stimulus and reinforcing properties of etoxadrol and dexoxadrol in monkeys. J Pharmacol Exp Ther. 1982b;220:56–62. [PubMed] [Google Scholar]
- Braff DL, Light GA, Swerdlow NR. Prepulse inhibition and P50 suppression are both deficient but not correlated in schizophrenia patients. Biol Psychiatry. 2007;61:1204–1207. doi: 10.1016/j.biopsych.2006.08.015. [DOI] [PubMed] [Google Scholar]
- Bresink I, Benke TA, Collett VJ, Seal AJ, Parsons CG, Henley JM, et al. Effects of memantine on recombinant rat NMDA receptors expressed in HEK 293 cells. Br J Pharmacol. 1996;119:195–204. doi: 10.1111/j.1476-5381.1996.tb15971.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Broberg BV, Dias R, Glenthøj BY, Olsen CK. Evaluation of a neurodevelopmental model of schizophrenia – early postnatal PCP treatment in attentional set-shifting. Behav Brain Res. 2008;190:160–163. doi: 10.1016/j.bbr.2008.02.020. [DOI] [PubMed] [Google Scholar]
- Buccafusco JJ, Terry A., V A reversible model of the cognitive impairment associated with schizophrenia in monkeys: potential therapeutic effects of two nicotinic acetylcholine receptor agonists. Biochem Pharmacol. 2009;78:852–862. doi: 10.1016/j.bcp.2009.06.102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Buisson B, Bertrand D. Open-channel blockers at the human alpha4beta2 neuronal nicotinic acetylcholine receptor. Mol Pharmacol. 1998;53:555–563. doi: 10.1124/mol.53.3.555. [DOI] [PubMed] [Google Scholar]
- Burnashev N, Schoepfer R, Monyer H, Ruppersberg JP, Günther W, Seeburg PH, et al. Control by asparagine residues of calcium permeability and magnesium blockade in the NMDA receptor. Science. 1992;257:1415–1419. doi: 10.1126/science.1382314. [DOI] [PubMed] [Google Scholar]
- Caddy C, Giaroli G, White TP, Shergill SS, Tracy DK. Ketamine as the prototype glutamatergic antidepressant: pharmacodynamic actions, and a systematic review and meta-analysis of efficacy. Ther Adv Psychopharmacol. 2014;4:75–99. doi: 10.1177/2045125313507739. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cartmell J, Monn JA, Schoepp DD. The metabotropic glutamate 2/3 receptor agonists LY354740 and LY379268 selectively attenuate phencyclidine versus d-amphetamine motor behaviors in rats. J Pharmacol Exp Ther. 1999;291:161–170. [PubMed] [Google Scholar]
- Catts VS, Fung SJ, Long LE, Joshi D, Vercammen A, Allen KM, et al. Rethinking schizophrenia in the context of normal neurodevelopment. Front Cell Neurosci. 2013;7:60. doi: 10.3389/fncel.2013.00060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chaki S, Yoshikawa R, Hirota S, Shimazaki T, Maeda M, Kawashima N, et al. MGS0039: a potent and selective group II metabotropic glutamate receptor antagonist with antidepressant-like activity. Neuropharmacology. 2004;46:457–467. doi: 10.1016/j.neuropharm.2003.10.009. [DOI] [PubMed] [Google Scholar]
- Chaperon F, Müller W, Auberson YP, Tricklebank MD, Neijt HC. Substitution for PCP, disruption of prepulse inhibition and hyperactivity induced by N-methyl-D-aspartate receptor antagonists: preferential involvement of the NR2B rather than NR2A subunit. Behav Pharmacol. 2003;14:477–487. doi: 10.1097/01.fbp.0000091471.79060.ed. [DOI] [PubMed] [Google Scholar]
- Chapman AG, Meldrum BS. Non-competitive N-methyl-D-aspartate antagonists protect against sound-induced seizures in DBA/2 mice. Eur J Pharmacol. 1989;166:201–211. doi: 10.1016/0014-2999(89)90060-5. [DOI] [PubMed] [Google Scholar]
- Chen C, Chow S. Effects of ketamine on synaptic transmission in cat spinal cord. Neuropharmacology. 1975;14:147–149. doi: 10.1016/0028-3908(75)90037-4. [DOI] [PubMed] [Google Scholar]
- Chen G, Ensor CR, Russell D, Bohner B. The pharmacology of 1-(1-phenylcyclohexyl) piperidine HCl. J Pharmacol Exp Ther. 1959;127:241–250. [PubMed] [Google Scholar]
- Chen X, Shu S, Bayliss DA. HCN1 channel subunits are a molecular substrate for hypnotic actions of ketamine. J Neurosci. 2009;29:600–609. doi: 10.1523/JNEUROSCI.3481-08.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chizh BA. Low dose ketamine: a therapeutic and research tool to explore N-methyl-D-aspartate (NMDA) receptor-mediated plasticity in pain pathways. J Psychopharmacol. 2007;21:259–271. doi: 10.1177/0269881105062484. [DOI] [PubMed] [Google Scholar]
- Cho RY, Konecky RO, Carter CS. Impairments in frontal cortical gamma synchrony and cognitive control in schizophrenia. Proc Natl Acad Sci U S A. 2006;103:19878–19883. doi: 10.1073/pnas.0609440103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Christiaen E. Reaction of organo-magnesium compounds on α-amino nitriles. Bull Des Soc Chim Belg. 1924;33:483. [Google Scholar]
- Chu PS-K, Ma W-K, Wong SC-W, Chu RW-H, Cheng C-H, Wong S, et al. The destruction of the lower urinary tract by ketamine abuse: a new syndrome? BJU Int. 2008;102:1616–1622. doi: 10.1111/j.1464-410X.2008.07920.x. [DOI] [PubMed] [Google Scholar]
- Church J, Lodge D. Cyclazocine and pentazocine as N-methylaspartate antagonists on cat and rat spinal neurons in vivo. J Pharmacol Exp Ther. 1990;253:636–645. [PubMed] [Google Scholar]
- Church J, Lodge D, Berry SC. Differential effects of dextrorphan and levorphanol on the excitation of rat spinal neurons by amino acids. Eur J Pharmacol. 1985;111:185–190. doi: 10.1016/0014-2999(85)90755-1. [DOI] [PubMed] [Google Scholar]
- Church J, Zeman S, Lodge D. The neuroprotective action of ketamine and MK-801 after transient cerebral ischemia in rats. Anesthesiology. 1988;69:702–709. doi: 10.1097/00000542-198811000-00011. [DOI] [PubMed] [Google Scholar]
- Church J, Millar JD, Jones MG, Lodge D. NMDA receptor antagonist effects of the stereoisomers of beta-cyclazocine in rats, in vivo and in vitro. Eur J Pharmacol. 1991;192:337–342. doi: 10.1016/0014-2999(91)90222-c. [DOI] [PubMed] [Google Scholar]
- Cline HT, Tsien RW. Glutamate-induced increases in intracellular Ca2+ in cultured frog tectal cells mediated by direct activation of NMDA receptor channels. Neuron. 1991;6:259–267. doi: 10.1016/0896-6273(91)90361-3. [DOI] [PubMed] [Google Scholar]
- Cline HT, Debski EA, Constantine-Paton M. N-methyl-D-aspartate receptor antagonist desegregates eye-specific stripes. Proc Natl Acad Sci U S A. 1987;84:4342–4345. doi: 10.1073/pnas.84.12.4342. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cochran SM, Kennedy M, McKerchar CE, Steward LJ, Pratt JA, Morris BJ. Induction of metabolic hypofunction and neurochemical deficits after chronic intermittent exposure to phencyclidine: differential modulation by antipsychotic drugs. Neuropsychopharmacology. 2003;28:265–275. doi: 10.1038/sj.npp.1300031. [DOI] [PubMed] [Google Scholar]
- Collingridge GL, Singer W. Excitatory amino acid receptors and synaptic plasticity. Trends Pharmacol Sci. 1990;11:290–296. doi: 10.1016/0165-6147(90)90011-v. [DOI] [PubMed] [Google Scholar]
- Collingridge GL, Kehl SJ, McLennan H. Excitatory amino acids in synaptic transmission in the Schaffer collateral-commissural pathway of the rat hippocampus. J Physiol. 1983;334:33–46. doi: 10.1113/jphysiol.1983.sp014478. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Collingridge GL, Peineau S, Howland JG, Wang YT. Long-term depression in the CNS. Nat Rev Neurosci. 2010;11:459–473. doi: 10.1038/nrn2867. [DOI] [PubMed] [Google Scholar]
- Collingridge GL, Volianskis A, Bannister N, France G, Hanna L, Mercier M, et al. The NMDA receptor as a target for cognitive enhancement. Neuropharmacology. 2013;64:13–26. doi: 10.1016/j.neuropharm.2012.06.051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Coutinho E, Harrison P, Vincent A. Do neuronal autoantibodies cause psychosis? A neuroimmunological perspective. Biol Psychiatry. 2014;75:269–275. doi: 10.1016/j.biopsych.2013.07.040. [DOI] [PubMed] [Google Scholar]
- Coyle JT. NMDA receptor and schizophrenia: a brief history. Schizophr Bull. 2012;38:920–926. doi: 10.1093/schbul/sbs076. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crosby G, Crane AM, Sokoloff L. Local changes in cerebral glucose utilization during ketamine anesthesia. Anesthesiology. 1982;56:437–443. doi: 10.1097/00000542-198206000-00005. [DOI] [PubMed] [Google Scholar]
- Croucher MJ, Collins JF, Meldrum BS. Anticonvulsant action of excitatory amino acid antagonists. Science. 1982;216:899–901. doi: 10.1126/science.7079744. [DOI] [PubMed] [Google Scholar]
- Cunningham MO, Hunt J, Middleton S, LeBeau FE, Gillies MJ, Davies CH, et al. Region-specific reduction in entorhinal gamma oscillations and parvalbumin-immunoreactive neurons in animal models of psychiatric illness. J Neurosci. 2006;26:2767–2776. doi: 10.1523/JNEUROSCI.5054-05.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Curtis DR, Watkins JC. The excitation and depression of spinal neurones by structurally related amino acids. J Neurochem. 1960;6:117–141. doi: 10.1111/j.1471-4159.1960.tb13458.x. [DOI] [PubMed] [Google Scholar]
- Danysz W, Parsons CG. Glycine and N-methyl-D-aspartate receptors: physiological significance and possible therapeutic applications. Pharmacol Rev. 1998;50:597–664. [PubMed] [Google Scholar]
- Davies J, Watkins JC. Selective antagonism of amino acid-induced and synaptic excitation in the cat spinal cord. J Physiol. 1979;297:621–635. doi: 10.1113/jphysiol.1979.sp013060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Davies J, Francis AA, Jones AW, Watkins JC. 2-Amino-5-phosphonovalerate (2APV), a potent and selective antagonist of amino acid-induced and synaptic excitation. Neurosci Lett. 1981;21:77–81. doi: 10.1016/0304-3940(81)90061-6. [DOI] [PubMed] [Google Scholar]
- Davies SN, Lodge D. Evidence for involvement of N-methylaspartate receptors in ‘wind-up’ of class 2 neurones in the dorsal horn of the rat. Brain Res. 1987;424:402–406. doi: 10.1016/0006-8993(87)91487-9. [DOI] [PubMed] [Google Scholar]
- Davies SN, Martin D, Millar JD, Aram JA, Church J, Lodge D. Differences in results from in vivo and in vitro studies on the use-dependency of N-methylaspartate antagonism by MK-801 and other phencyclidine receptor ligands. Eur J Pharmacol. 1988;145:141–151. doi: 10.1016/0014-2999(88)90225-7. [DOI] [PubMed] [Google Scholar]
- Davis SM, Lees KR, Albers GW, Diener HC, Markabi S, Karlsson G, et al. Selfotel in acute ischemic stroke: possible neurotoxic effects of an NMDA antagonist. Stroke. 2000;31:347–354. doi: 10.1161/01.str.31.2.347. [DOI] [PubMed] [Google Scholar]
- Dawson N, Morris BJ, Pratt JA. Subanaesthetic ketamine treatment alters prefrontal cortex connectivity with thalamus and ascending subcortical systems. Schizophr Bull. 2013;39:366–377. doi: 10.1093/schbul/sbr144. [DOI] [PMC free article] [PubMed] [Google Scholar]
- De Vry J, Jentzsch KR. Role of the NMDA receptor NR2B subunit in the discriminative stimulus effects of ketamine. Behav Pharmacol. 2003;14:229–235. doi: 10.1097/00008877-200305000-00007. [DOI] [PubMed] [Google Scholar]
- Deakin J, Lennox BR, Zandi MS. Antibodies to the N-methyl-D-aspartate receptor and other synaptic proteins in psychosis. Biol Psychiatry. 2014;75:284–291. doi: 10.1016/j.biopsych.2013.07.018. [DOI] [PubMed] [Google Scholar]
- Domino EF. Neurobiology of phencyclidine (Sernyl), a drug with an unusual spectrum of pharmacological activity. Int Rev Neurobiol. 1964;6:303–347. doi: 10.1016/s0074-7742(08)60772-2. [DOI] [PubMed] [Google Scholar]
- Domino EF, Luby ED. Phencyclidine/schizophrenia: one view toward the past, the other to the future. Schizophr Bull. 2012;38:914–919. doi: 10.1093/schbul/sbs011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Domino EF, Chodoff P, Corssen G. Pharmacologic effects of CI-581, a new dissociative anesthetic, in man. Clin Pharmacol Ther. 1965;6:279–291. doi: 10.1002/cpt196563279. [DOI] [PubMed] [Google Scholar]
- Dravid SM, Erreger K, Yuan H, Nicholson K, Le P, Lyuboslavsky P, et al. Subunit-specific mechanisms and proton sensitivity of NMDA receptor channel block. J Physiol. 2007;581:107–128. doi: 10.1113/jphysiol.2006.124958. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Duman RS. Pathophysiology of depression and innovative treatments: remodeling glutamatergic synaptic connections. Dialogues Clin Neurosci. 2014;16:11–27. doi: 10.31887/DCNS.2014.16.1/rduman. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Duman RS, Li N, Liu R-J, Duric V, Aghajanian G. Signaling pathways underlying the rapid antidepressant actions of ketamine. Neuropharmacology. 2012;62:35–41. doi: 10.1016/j.neuropharm.2011.08.044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dunlop J, Brandon NJ. Schizophrenia drug discovery and development in an evolving era: are new drug targets fulfilling expectations? J Psychopharmacol. 2015;29:230–238. doi: 10.1177/0269881114565806. [DOI] [PubMed] [Google Scholar]
- Dwyer JM, Lepack AE, Duman RS. mTOR activation is required for the antidepressant effects of mGluR2/3 blockade. Int J Neuropsychopharmacol. 2012;15:429–434. doi: 10.1017/S1461145711001702. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Egerton A, Reid L, McGregor S, Cochran SM, Morris BJ, Pratt JA. Subchronic and chronic PCP treatment produces temporally distinct deficits in attentional set shifting and prepulse inhibition in rats. Psychopharmacology (Berl) 2008;198:37–49. doi: 10.1007/s00213-008-1071-5. [DOI] [PubMed] [Google Scholar]
- Ehrlichman RS, Gandal MJ, Maxwell CR, Lazarewicz MT, Finkel LH, Contreras D, et al. N-methyl-d-aspartic acid receptor antagonist-induced frequency oscillations in mice recreate pattern of electrophysiological deficits in schizophrenia. Neuroscience. 2009;158:705–712. doi: 10.1016/j.neuroscience.2008.10.031. [DOI] [PubMed] [Google Scholar]
- Elliott SP, Brandt SD, Wallach J, Morris H, Kavanagh PV. First reported fatalities associated with the ‘research chemical’ 2-methoxydiphenidine. J Anal Toxicol. 2015;39:287–293. doi: 10.1093/jat/bkv006. [DOI] [PubMed] [Google Scholar]
- Ellison G. Competitive and non-competitive NMDA antagonists induce similar limbic degeneration. Neuroreport. 1994;5:2688–2692. doi: 10.1097/00001756-199412000-00070. [DOI] [PubMed] [Google Scholar]
- Ellison G. The N-methyl-D-aspartate antagonists phencyclidine, ketamine and dizocilpine as both behavioral and anatomical models of the dementias. Brain Res Brain Res Rev. 1995;20:250–267. doi: 10.1016/0165-0173(94)00014-g. [DOI] [PubMed] [Google Scholar]
- Emnett CM, Eisenman LN, Taylor AM, Izumi Y, Zorumski CF, Mennerick S. Indistinguishable synaptic pharmacodynamics of the N-methyl-D-aspartate receptor channel blockers memantine and ketamine. Mol Pharmacol. 2013;84:935–947. doi: 10.1124/mol.113.089334. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Erowid Experience Vaults. 2013. Dizocilpine Reports. [Online] Available at: https://www.erowid.org/experiences/subs/exp_Dizocilpine.shtml (accessed 2/25 2015)
- Evans RH, Watkins JC. Pharmacological antagonists of excitant amino acid action. Life Sci. 1981;28:1303–1308. doi: 10.1016/0024-3205(81)90401-x. [DOI] [PubMed] [Google Scholar]
- Evans RH, Francis AA, Watkins JC. Selective antagonism by Mg2+ of amino acid-induced depolarization of spinal neurones. Experientia. 1977;33:489–491. doi: 10.1007/BF01922227. [DOI] [PubMed] [Google Scholar]
- Fagg GE. Phencyclidine and related drugs bind to the activated N-methyl-D-aspartate receptor-channel complex in rat brain membranes. Neurosci Lett. 1987;76:221–227. doi: 10.1016/0304-3940(87)90719-1. [DOI] [PubMed] [Google Scholar]
- Fletcher EJ, Drew C, Lodge D, O’Shaughnessy CT. Efflux of rubidium in rat cortical synaptosomes is blocked by sigma and dextromethorphan binding site ligands. Neuropharmacology. 1989;28:661–666. doi: 10.1016/0028-3908(89)90148-2. [DOI] [PubMed] [Google Scholar]
- Foster AC, Wong EH. The novel anticonvulsant MK-801 binds to the activated state of the N-methyl-D-aspartate receptor in rat brain. Br J Pharmacol. 1987;91:403–409. doi: 10.1111/j.1476-5381.1987.tb10295.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Foussias G, Siddiqui I, Fervaha G, Agid O, Remington G. Dissecting negative symptoms in schizophrenia: opportunities for translation into new treatments. J Psychopharmacol. 2015;29:116–126. doi: 10.1177/0269881114562092. [DOI] [PubMed] [Google Scholar]
- Franks NP, Lieb WR. Molecular and cellular mechanisms of general anaesthesia. Nature. 1994;367:607–614. doi: 10.1038/367607a0. [DOI] [PubMed] [Google Scholar]
- Frederickson EL, Longnecker DE, Allen GW. Clinical investigation of a new intravenous anesthetic – etoxadrol hydrochloride (CL-1848; U-37862A) Anesth Analg. 1976;55:335–339. doi: 10.1213/00000539-197605000-00010. [DOI] [PubMed] [Google Scholar]
- Freeman AS, Bunney BS. The effects of phencyclidine and N-allylnormetazocine on midbrain dopamine neuronal activity. Eur J Pharmacol. 1984;104:287–293. doi: 10.1016/0014-2999(84)90404-7. [DOI] [PubMed] [Google Scholar]
- Friederich P, Dybek A, Urban BW. Stereospecific interaction of ketamine with nicotinic acetylcholine receptors in human sympathetic ganglion-like SH-SY5Y cells. Anesthesiology. 2000;93:818–824. doi: 10.1097/00000542-200009000-00032. [DOI] [PubMed] [Google Scholar]
- Friston KJ. The disconnection hypothesis. Schizophr Res. 1998;30:115–125. doi: 10.1016/s0920-9964(97)00140-0. [DOI] [PubMed] [Google Scholar]
- Frohlich J, Van Horn JD. Reviewing the ketamine model for schizophrenia. J Psychopharmacol. 2014;28:287–302. doi: 10.1177/0269881113512909. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gakuba C, Gauberti M, Mazighi M, Defer G, Hanouz J-L, Vivien D. Preclinical evidence toward the use of ketamine for recombinant tissue-type plasminogen activator-mediated thrombolysis under anesthesia or sedation. Stroke. 2011;42:2947–2949. doi: 10.1161/STROKEAHA.111.620468. [DOI] [PubMed] [Google Scholar]
- Garey RE, Heath RG. The effects of phencyclidine on the uptake of 3H-catecholamines by rat striatal and hypothalamic synaptosomes. Life Sci. 1976;18:1105–1110. doi: 10.1016/0024-3205(76)90145-4. [DOI] [PubMed] [Google Scholar]
- Gaspard N, Foreman B, Judd LM, Brenton JN, Nathan BR, McCoy BM, et al. Intravenous ketamine for the treatment of refractory status epilepticus: a retrospective multicenter study. Epilepsia. 2013;54:1498–1503. doi: 10.1111/epi.12247. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Geyer MA, Krebs-Thomson K, Braff DL, Swerdlow NR. Pharmacological studies of prepulse inhibition models of sensorimotor gating deficits in schizophrenia: a decade in review. Psychopharmacology (Berl) 2001;156:117–154. doi: 10.1007/s002130100811. [DOI] [PubMed] [Google Scholar]
- Gil-da-Costa R, Stoner GR, Fung R, Albright TD. Nonhuman primate model of schizophrenia using a noninvasive EEG method. Proc Natl Acad Sci U S A. 2013;110:15425–15430. doi: 10.1073/pnas.1312264110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gill R, Foster AC, Woodruff GN. MK-801 is neuroprotective in gerbils when administered during the post-ischaemic period. Neuroscience. 1988;25:847–855. doi: 10.1016/0306-4522(88)90040-1. [DOI] [PubMed] [Google Scholar]
- Gilling KE, Jatzke C, Hechenberger M, Parsons CG. Potency, voltage-dependency, agonist concentration-dependency, blocking kinetics and partial untrapping of the uncompetitive N-methyl-D-aspartate (NMDA) channel blocker memantine at human NMDA (GluN1/GluN2A) receptors. Neuropharmacology. 2009;56:866–875. doi: 10.1016/j.neuropharm.2009.01.012. [DOI] [PubMed] [Google Scholar]
- Gilmour G, Dix S, Fellini L, Gastambide F, Plath N, Steckler T, et al. NMDA receptors, cognition and schizophrenia – testing the validity of the NMDA receptor hypofunction hypothesis. Neuropharmacology. 2012;62:1401–1412. doi: 10.1016/j.neuropharm.2011.03.015. [DOI] [PubMed] [Google Scholar]
- Goetghebeur P, Dias R. Comparison of haloperidol, risperidone, sertindole, and modafinil to reverse an attentional set-shifting impairment following subchronic PCP administration in the rat – a back translational study. Psychopharmacology (Berl) 2009;202:287–293. doi: 10.1007/s00213-008-1132-9. [DOI] [PubMed] [Google Scholar]
- Goldberg JH, Yuste R, Tamas G. Ca2+ imaging of mouse neocortical interneurone dendrites: contribution of Ca2+-permeable AMPA and NMDA receptors to subthreshold Ca2+dynamics. J Physiol. 2003;551:67–78. doi: 10.1113/jphysiol.2003.042598. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goto Y, Grace AA. The dopamine system and the pathophysiology of schizophrenia: a basic science perspective. Int Rev Neurobiol. 2007;78:41–68. doi: 10.1016/S0074-7742(06)78002-3. [DOI] [PubMed] [Google Scholar]
- Gray NM, Cheng BK. 1989. 1,2-Diarylethylamines for treatment of neurotoxic injury.
- Greifenstein FE, Devault M, Yoshitake J, Gajewski JE. A study of a 1-arylcyclohexylamine for anesthesia. Anesth Analg. 1958;37:283–294. [PubMed] [Google Scholar]
- Grotta J, Clark W, Coull B, Pettigrew LC, Mackay B, Goldstein LB, et al. Safety and tolerability of the glutamate antagonist CGS 19755 (Selfotel) in patients with acute ischemic stroke. Results of a phase IIa randomized trial. Stroke. 1995;26:602–605. doi: 10.1161/01.str.26.4.602. [DOI] [PubMed] [Google Scholar]
- Grunze HC, Rainnie DG, Hasselmo ME, Barkai E, Hearn EF, McCarley RW, et al. NMDA-dependent modulation of CA1 local circuit inhibition. J Neurosci. 1996;16:2034–2043. doi: 10.1523/JNEUROSCI.16-06-02034.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guidotti A, Auta J, Davis JM, Di-Giorgi-Gerevini V, Dwivedi Y, Grayson DR, et al. Decrease in reelin and glutamic acid decarboxylase67 (GAD67) expression in schizophrenia and bipolar disorder: a postmortem brain study. Arch Gen Psychiatry. 2000;57:1061–1069. doi: 10.1001/archpsyc.57.11.1061. [DOI] [PubMed] [Google Scholar]
- Haertzen CA. Subjective effects of narcotic antagonists cyclazocine and nalorphine on the Addiction Research Center Inventory (ARCI) Psychopharmacologia. 1970;18:366–377. doi: 10.1007/BF00402763. [DOI] [PubMed] [Google Scholar]
- Hagino Y, Kasai S, Han W, Yamamoto H, Nabeshima T, Mishina M, et al. Essential role of NMDA receptor channel ε4 subunit (GluN2D) in the effects of phencyclidine, but not methamphetamine. PLoS ONE. 2010;5:e13722. doi: 10.1371/journal.pone.0013722. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hammer RP, Herkenham M, Pert CB, Quirion R. Correlation of regional brain metabolism with receptor localization during ketamine anesthesia: combined autoradiographic 2-[3H]deoxy-D-glucose receptor binding technique. Proc Natl Acad Sci U S A. 1982;79:3067–3070. doi: 10.1073/pnas.79.9.3067. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hampton RY, Medzihradsky F, Woods JH, Dahlstrom PJ. Stereospecific binding of 3H-phencyclidine in brain membranes. Life Sci. 1982;30:2147–2154. doi: 10.1016/0024-3205(82)90288-0. [DOI] [PubMed] [Google Scholar]
- Hanson JE, Weber M, Meilandt WJ, Wu T, Luu T, Deng L, et al. GluN2B antagonism affects interneurons and leads to immediate and persistent changes in synaptic plasticity, oscillations, and behavior. Neuropsychopharmacology. 2013;38:1221–1233. doi: 10.1038/npp.2013.19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harrison NL, Simmonds MA. Quantitative studies on some antagonists of N-methyl D-aspartate in slices of rat cerebral cortex. Br J Pharmacol. 1985;84:381–391. doi: 10.1111/j.1476-5381.1985.tb12922.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harrison PJ. The neuropathology of schizophrenia. A critical review of the data and their interpretation. Brain. 1999;122:593–624. doi: 10.1093/brain/122.4.593. [DOI] [PubMed] [Google Scholar]
- Harrison PJ. Recent genetic findings in schizophrenia and their therapeutic relevance. J Psychopharmacol. 2015;29:85–96. doi: 10.1177/0269881114553647. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harrison PJ, Weinberger DR. Schizophrenia genes, gene expression, and neuropathology: on the matter of their convergence. Mol Psychiatry. 2005;10:40–68. doi: 10.1038/sj.mp.4001558. [DOI] [PubMed] [Google Scholar]
- Harrison PJ, Pritchett D, Stumpenhorst K, Betts JF, Nissen W, Schweimer J, et al. Genetic mouse models relevant to schizophrenia: taking stock and looking forward. Neuropharmacology. 2012;62:1164–1167. doi: 10.1016/j.neuropharm.2011.08.009. [DOI] [PubMed] [Google Scholar]
- Hashimoto T, Volk DW, Eggan SM, Mirnics K, Pierri JN, Sun Z, et al. Gene expression deficits in a subclass of GABA neurons in the prefrontal cortex of subjects with schizophrenia. J Neurosci. 2003;23:6315–6326. doi: 10.1523/JNEUROSCI.23-15-06315.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heekeren K, Daumann J, Neukirch A, Stock C, Kawohl W, Norra C, et al. Mismatch negativity generation in the human 5HT2A agonist and NMDA antagonist model of psychosis. Psychopharmacology (Berl) 2008;199:77–88. doi: 10.1007/s00213-008-1129-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Herling S, Coale EH, Hein DW, Winger G, Woods JH. Similarity of the discriminative stimulus effects of ketamine, cyclazocine, and dextrorphan in the pigeon. Psychopharmacology (Berl) 1981;73:286–291. doi: 10.1007/BF00422419. [DOI] [PubMed] [Google Scholar]
- Herrling P. Excitatory Amino Acids: Clinical Results with Antagonists. London: Academic Press; 1997. [Google Scholar]
- Holtzman SG. Phencyclidine-like discriminative effects of opioids in the rat. J Pharmacol Exp Ther. 1980;214:614–619. [PubMed] [Google Scholar]
- Holtzman SG. Phencyclidine-like discriminative stimulus properties of opioids in the squirrel monkey. Psychopharmacology (Berl) 1982;77:295–300. doi: 10.1007/BF00432758. [DOI] [PubMed] [Google Scholar]
- Howes O, McCutcheon R, Stone J. Glutamate and dopamine in schizophrenia: an update for the 21st century. J Psychopharmacol. 2015;29:97–115. doi: 10.1177/0269881114563634. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hudetz JA, Pagel PS. Neuroprotection by ketamine: a review of the experimental and clinical evidence. J Cardiothorac Vasc Anesth. 2010;24:131–142. doi: 10.1053/j.jvca.2009.05.008. [DOI] [PubMed] [Google Scholar]
- Huettner JE, Bean BP. Block of N-methyl-D-aspartate-activated current by the anticonvulsant MK-801: selective binding to open channels. Proc Natl Acad Sci U S A. 1988;85:1307–1311. doi: 10.1073/pnas.85.4.1307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ikonomidou C, Bosch F, Miksa M, Bittigau P, Vöckler J, Dikranian K, et al. Blockade of NMDA receptors and apoptotic neurodegeneration in the developing brain. Science. 1999;283:70–74. doi: 10.1126/science.283.5398.70. [DOI] [PubMed] [Google Scholar]
- Isbell H, Fraser HF. Actions and addiction liabilities of dromoran derivatives in man. J Pharmacol Exp Ther. 1953;107:524–530. [PubMed] [Google Scholar]
- Jasinski DR. Human pharmacology of narcotic antagonists. Br J Clin Pharmacol. 1979;7(Suppl. 3):287S–290S. doi: 10.1111/j.1365-2125.1979.tb04702.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jasinski DR, Martin WR, Hoeldtke RD. Effects of short- and long-term administration of pentazocine in man. Clin Pharmacol Ther. 1970;11:385–403. doi: 10.1002/cpt1970113385. [DOI] [PubMed] [Google Scholar]
- Javitt DC. Glutamate and schizophrenia: phencyclidine, N-methyl-D-aspartate receptors, and dopamine-glutamate interactions. Int Rev Neurobiol. 2007;78:69–108. doi: 10.1016/S0074-7742(06)78003-5. [DOI] [PubMed] [Google Scholar]
- Javitt DC. Twenty-five years of glutamate in schizophrenia: are we there yet? Schizophr Bull. 2012;38:911–913. doi: 10.1093/schbul/sbs100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Javitt DC, Zukin SR. Recent advances in the phencyclidine model of schizophrenia. Am J Psychiatry. 1991;148:1301–1308. doi: 10.1176/ajp.148.10.1301. [DOI] [PubMed] [Google Scholar]
- Javitt DC, Steinschneider M, Schroeder CE, Arezzo JC. Role of cortical N-methyl-D-aspartate receptors in auditory sensory memory and mismatch negativity generation: implications for schizophrenia. Proc Natl Acad Sci U S A. 1996;93:11962–11967. doi: 10.1073/pnas.93.21.11962. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jentsch JD, Roth RH. The neuropsychopharmacology of phencyclidine: from NMDA receptor hypofunction to the dopamine hypothesis of schizophrenia. Neuropsychopharmacology. 1999;20:201–225. doi: 10.1016/S0893-133X(98)00060-8. [DOI] [PubMed] [Google Scholar]
- Jentsch JD, Redmond DE, Elsworth JD, Taylor JR, Youngren KD, Roth RH. Enduring cognitive deficits and cortical dopamine dysfunction in monkeys after long-term administration of phencyclidine. Science. 1997;277:953–955. doi: 10.1126/science.277.5328.953. [DOI] [PubMed] [Google Scholar]
- Jentsch JD, Elsworth JD, Taylor JR, Redmond DE, Roth RH. Dysregulation of mesoprefrontal dopamine neurons induced by acute and repeated phencyclidine administration in the nonhuman primate: implications for schizophrenia. Adv Pharmacol. 1998;42:810–814. doi: 10.1016/s1054-3589(08)60870-4. [DOI] [PubMed] [Google Scholar]
- Johnson JW, Ascher P. Glycine potentiates the NMDA response in cultured mouse brain neurons. Nature. 1987;325:529–531. doi: 10.1038/325529a0. [DOI] [PubMed] [Google Scholar]
- Johnson KM. Phencyclidine: behavioral and biochemical evidence supporting a role for dopamine. Fed Proc. 1983;42:2579–2583. [PubMed] [Google Scholar]
- Johnston GAR. Advantages of an antagonist: bicuculline and other GABA antagonists. Br J Pharmacol. 2013;169:328–336. doi: 10.1111/bph.12127. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jones EG. Cortical development and thalamic pathology in schizophrenia. Schizophr Bull. 1997;23:483–501. doi: 10.1093/schbul/23.3.483. [DOI] [PubMed] [Google Scholar]
- Jones RS, Bühl EH. Basket-like interneurones in layer II of the entorhinal cortex exhibit a powerful NMDA-mediated synaptic excitation. Neurosci Lett. 1993;149:35–39. doi: 10.1016/0304-3940(93)90341-h. [DOI] [PubMed] [Google Scholar]
- Kamenka JM, Domino EF, Geneste P. Phencyclidine and Related Arylcyclohexylamines: Present and Future Applications. Ann Arbor, MI: NPP Books; 1983. [Google Scholar]
- Kashiwagi K, Masuko T, Nguyen CD, Kuno T, Tanaka I, Igarashi K, et al. Channel blockers acting at N-methyl-D-aspartate receptors: differential effects of mutations in the vestibule and ion channel pore. Mol Pharmacol. 2002;61:533–545. doi: 10.1124/mol.61.3.533. [DOI] [PubMed] [Google Scholar]
- Kayser MS, Dalmau J. Anti-NMDA receptor encephalitis, autoimmunity, and psychosis. Schizophr Res. 2014 doi: 10.1016/j.schres.2014.10.007. doi: 10.1016/j.schres.2014.10.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Keats AS, Telford J. Studies of analgesic drugs. VIII. A narcotic antagonist analgesic without psychotomimetic effects. J Pharmacol Exp Ther. 1964;143:157–164. [PubMed] [Google Scholar]
- Keilhoff G, Bernstein H-G, Becker A, Grecksch G, Wolf G. Increased neurogenesis in a rat ketamine model of schizophrenia. Biol Psychiatry. 2004;56:317–322. doi: 10.1016/j.biopsych.2004.06.010. [DOI] [PubMed] [Google Scholar]
- Kleinschmidt A, Bear MF, Singer W. Blockade of ‘NMDA’ receptors disrupts experience-dependent plasticity of kitten striate cortex. Science. 1987;238:355–358. doi: 10.1126/science.2443978. [DOI] [PubMed] [Google Scholar]
- Klepstad P, Maurset A, Moberg ER, Oye I. Evidence of a role for NMDA receptors in pain perception. Eur J Pharmacol. 1990;187:513–518. doi: 10.1016/0014-2999(90)90379-k. [DOI] [PubMed] [Google Scholar]
- Kloog Y, Rehavi M, Maayani S, Sokolovsky M. Anticholinesterase and antiacetylcholine activity of 1-phenylcyclohexylamine derivatives. Eur J Pharmacol. 1977;45:221–227. doi: 10.1016/0014-2999(77)90002-4. [DOI] [PubMed] [Google Scholar]
- Kloog Y, Haring R, Sokolovsky M. Kinetic characterization of the phencyclidine-N-methyl-D-aspartate receptor interaction: evidence for a steric blockade of the channel. Biochemistry. 1988;27:843–848. doi: 10.1021/bi00403a001. [DOI] [PubMed] [Google Scholar]
- Kocsis B. Differential role of NR2A and NR2B subunits in N-methyl-D-aspartate receptor antagonist-induced aberrant cortical gamma oscillations. Biol Psychiatry. 2012;71:987–995. doi: 10.1016/j.biopsych.2011.10.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koek W, Woods JH, Ornstein P. A simple and rapid method for assessing similarities among directly observable behavioral effects of drugs: PCP-like effects of 2-amino-5-phosphonovalerate in rats. Psychopharmacology (Berl) 1987;91:297–304. doi: 10.1007/BF00518181. [DOI] [PubMed] [Google Scholar]
- Koek W, Woods JH, Winger GD. MK-801, a proposed noncompetitive antagonist of excitatory amino acid neurotransmission, produces phencyclidine-like behavioral effects in pigeons, rats and rhesus monkeys. J Pharmacol Exp Ther. 1988;245:969–974. [PubMed] [Google Scholar]
- Koike H, Chaki S. Requirement of AMPA receptor stimulation for the sustained antidepressant activity of ketamine and LY341495 during the forced swim test in rats. Behav Brain Res. 2014;271:111–115. doi: 10.1016/j.bbr.2014.05.065. [DOI] [PubMed] [Google Scholar]
- Kotermanski SE, Johnson JW. Mg2+ imparts NMDA receptor subtype selectivity to the Alzheimer’s drug memantine. J Neurosci. 2009;29:2774–2779. doi: 10.1523/JNEUROSCI.3703-08.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kraguljac N, V, White DM, Reid MA, Lahti AC. Increased hippocampal glutamate and volumetric deficits in unmedicated patients with schizophrenia. JAMA Psychiatry. 2013;70:1294–1302. doi: 10.1001/jamapsychiatry.2013.2437. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Krogsgaard-Larsen P, Honoré T, Hansen JJ, Curtis DR, Lodge D. New class of glutamate agonist structurally related to ibotenic acid. Nature. 1980;284:64–66. doi: 10.1038/284064a0. [DOI] [PubMed] [Google Scholar]
- Krystal JH, Karper LP, Seibyl JP, Freeman GK, Delaney R, Bremner JD, et al. Subanesthetic effects of the noncompetitive NMDA antagonist, ketamine, in humans. Arch Gen Psychiatry. 1994;51:199–214. doi: 10.1001/archpsyc.1994.03950030035004. [DOI] [PubMed] [Google Scholar]
- Krystal JH, Abi-Saab W, Perry E, D’Souza DC, Liu N, Gueorguieva R, et al. Preliminary evidence of attenuation of the disruptive effects of the NMDA glutamate receptor antagonist, ketamine, on working memory by pretreatment with the group II metabotropic glutamate receptor agonist, LY354740, in healthy human subjects. Psychopharmacology (Berl) 2005;179:303–309. doi: 10.1007/s00213-004-1982-8. [DOI] [PubMed] [Google Scholar]
- Labrie V, Pai S, Petronis A. Epigenetics of major psychosis: progress, problems and perspectives. Trends Genet. 2012a;28:427–435. doi: 10.1016/j.tig.2012.04.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Labrie V, Wong AHC, Roder JC. Contributions of the D-serine pathway to schizophrenia. Neuropharmacology. 2012b;62:1484–1503. doi: 10.1016/j.neuropharm.2011.01.030. [DOI] [PubMed] [Google Scholar]
- Lacey MG, Henderson G. Actions of phencyclidine on rat locus coeruleus neurones in vitro. Neuroscience. 1986;17:485–494. doi: 10.1016/0306-4522(86)90261-7. [DOI] [PubMed] [Google Scholar]
- Lahti AC, Weiler MA, Tamara M, Parwani A, Tamminga CA. Effects of ketamine in normal and schizophrenic volunteers. Neuropsychopharmacology. 2001;25:455–467. doi: 10.1016/S0893-133X(01)00243-3. [DOI] [PubMed] [Google Scholar]
- Lasagna L, Pearson JW. Analgesic and psychotomimetic properties of dexoxadrol. Proc Soc Exp Biol Med. 1965;118:352–354. doi: 10.3181/00379727-118-29840. [DOI] [PubMed] [Google Scholar]
- Lewis DA. Inhibitory neurons in human cortical circuits: substrate for cognitive dysfunction in schizophrenia. Curr Opin Neurobiol. 2014;26:22–26. doi: 10.1016/j.conb.2013.11.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lewis DA, Gonzalez-Burgos G. Pathophysiologically based treatment interventions in schizophrenia. Nat Med. 2006;12:1016–1022. doi: 10.1038/nm1478. [DOI] [PubMed] [Google Scholar]
- Li X, Tizzano JP, Griffey K, Clay M, Lindstrom T, Skolnick P. Antidepressant-like actions of an AMPA receptor potentiator (LY392098) Neuropharmacology. 2001;40:1028–1033. doi: 10.1016/s0028-3908(00)00194-5. [DOI] [PubMed] [Google Scholar]
- Li X, Witkin JM, Need AB, Skolnick P. Enhancement of antidepressant potency by a potentiator of AMPA receptors. Cell Mol Neurobiol. 2003;23:419–430. doi: 10.1023/A:1023648923447. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lima-Ojeda JM, Vogt MA, Pfeiffer N, Dormann C, Köhr G, Sprengel R, et al. Pharmacological blockade of GluN2B-containing NMDA receptors induces antidepressant-like effects lacking psychotomimetic action and neurotoxicity in the perinatal and adult rodent brain. Prog Neuropsychopharmacol Biol Psychiatry. 2013;45:28–33. doi: 10.1016/j.pnpbp.2013.04.017. [DOI] [PubMed] [Google Scholar]
- Lodge D, Anis NA. Effects of phencyclidine on excitatory amino acid activation of spinal interneurones in the cat. Eur J Pharmacol. 1982;77:203–204. doi: 10.1016/0014-2999(82)90022-x. [DOI] [PubMed] [Google Scholar]
- Lodge D, Anis NA. Effects of ketamine and three other anaesthetics on spinal reflexes and inhibitions in the cat. Br J Anaesth. 1984;56:1143–1151. doi: 10.1093/bja/56.10.1143. [DOI] [PubMed] [Google Scholar]
- Lodge D, Berry SC. Psychotomimetic effects of sigma opiates may be mediated by block of central excitatory synapses utilizing receptors for aspartate-like amino acids. Neurol Neurobiol. 1984;12:503–518. [Google Scholar]
- Lodge D, Johnson KM. Noncompetitive excitatory amino acid receptor antagonists. Trends Pharmacol Sci. 1990;11:81–86. doi: 10.1016/0165-6147(90)90323-z. [DOI] [PubMed] [Google Scholar]
- Lodge D, Johnston GA. Effect of ketamine on amino acid-evoked release of acetylcholine from rat cerebral cortex in vitro. Neurosci Lett. 1985;56:371–375. doi: 10.1016/0304-3940(85)90271-x. [DOI] [PubMed] [Google Scholar]
- Lodge D, Headley PM, Curtis DR. Selective antagonism by D-alpha-aminoadipate of amino acid and synaptic excitation of cat spinal neurons. Brain Res. 1978;152:603–608. doi: 10.1016/0006-8993(78)91117-4. [DOI] [PubMed] [Google Scholar]
- Lodge D, Anis NA, Burton NR. Effects of optical isomers of ketamine on excitation of cat and rat spinal neurones by amino acids and acetylcholine. Neurosci Lett. 1982;29:281–286. doi: 10.1016/0304-3940(82)90330-5. [DOI] [PubMed] [Google Scholar]
- Lodge D, Berry SC, Church J, Martin D, McGhee A, Lai HM, et al. Isomers of cyclazocine as excitatory amino acid antagonists. Neuropeptides. 1984;5:245–248. doi: 10.1016/0143-4179(84)90073-8. [DOI] [PubMed] [Google Scholar]
- Lodge D, Aram JA, Church J, Davies SN, Fletcher E, Martin D. Electrophysiological studies of the interaction between phencyclidine/sigma receptors agonists and excitatory amino acid neurotransmission on central mammalian neurones. In: Domino EF, Kamenka JM, editors. Sigma and Phencyclidine-like Compounds as Molecular Probes in Biology. Ann Arbor, MI: NPP Books; 1988a. pp. 239–250. [Google Scholar]
- Lodge D, Aram JA, Curch J, Davies SN, Martin D, Millar J. Sigma opiates and excitatory amino acids. In: Lodge D, et al., editors. Excitatory Amino Acids in Health and Disease. London: John Wiley & Sons; 1988b. pp. 237–259. [Google Scholar]
- Lodge D, Davies SN, Jones MG, Millar J, Manallack DT, Ornstein PL, et al. A comparison between the in vivo and in vitro activity of five potent and competitive NMDA antagonists. Br J Pharmacol. 1988c;95:957–965. doi: 10.1111/j.1476-5381.1988.tb11726.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Loo PS, Braunwalder AF, Lehmann J, Williams M, Sills MA. Interaction of L-glutamate and magnesium with phencyclidine recognition sites in rat brain: evidence for multiple affinity states of the phencyclidine/N-methyl-D-aspartate receptor complex. Mol Pharmacol. 1987;32:820–830. [PubMed] [Google Scholar]
- Lorrain DS, Baccei CS, Bristow LJ, Anderson JJ, Varney MA. Effects of ketamine and N-methyl-D-aspartate on glutamate and dopamine release in the rat prefrontal cortex: modulation by a group II selective metabotropic glutamate receptor agonist LY379268. Neuroscience. 2003;117:697–706. doi: 10.1016/s0306-4522(02)00652-8. [DOI] [PubMed] [Google Scholar]
- Luby ED, Cohen BD, Rosenbaum G, Gottlieb JS, Kelley R. Study of a new schizophrenomimetic drug – Sernyl. AMA Arch Neurol Psychiatry. 1959;81:363–369. doi: 10.1001/archneurpsyc.1959.02340150095011. [DOI] [PubMed] [Google Scholar]
- Luisada P, Reddick C. 1975. An epidemic of drug-induced schizophrenia. 128th Annu. Meet. Am. Psychiatr. Assoc. Anaheim, California. May, 5–9.
- Luisada PV. The phencyclidine psychosis: phenomenology and treatment. NIDA Res Monogr. 1978;21:241–253. [PubMed] [Google Scholar]
- Lustig C, Kozak R, Sarter M, Young JW, Robbins TW. CNTRICS final animal model task selection: control of attention. Neurosci Biobehav Rev. 2013;37:2099–2110. doi: 10.1016/j.neubiorev.2012.05.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- MacDermott AB, Mayer ML, Westbrook GL, Smith SJ, Barker JL. NMDA-receptor activation increases cytoplasmic calcium concentration in cultured spinal cord neurones. Nature. 1986;321:519–522. doi: 10.1038/321519a0. [DOI] [PubMed] [Google Scholar]
- MacDonald JF, Miljkovic Z, Pennefather P. Use-dependent block of excitatory amino acid currents in cultured neurons by ketamine. J Neurophysiol. 1987;58:251–266. doi: 10.1152/jn.1987.58.2.251. [DOI] [PubMed] [Google Scholar]
- MacDonald JF, Bartlett MC, Mody I, Pahapill P, Reynolds JN, Salter MW, et al. Actions of ketamine, phencyclidine and MK-801 on NMDA receptor currents in cultured mouse hippocampal neurones. J Physiol. 1991;432:483–508. doi: 10.1113/jphysiol.1991.sp018396. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maddox VH, Godefroi EF, Parcell RF. The Synthesis of Phencyclidine and Other 1-Arylcyclohexylamines. J Med Chem. 1965;8:230–235. doi: 10.1021/jm00326a019. [DOI] [PubMed] [Google Scholar]
- Maeda K, Sugino H, Hirose T, Kitagawa H, Nagai T, Mizoguchi H, et al. Clozapine prevents a decrease in neurogenesis in mice repeatedly treated with phencyclidine. J Pharmacol Sci. 2007;103:299–308. doi: 10.1254/jphs.fp0061424. [DOI] [PubMed] [Google Scholar]
- Malhotra AK, Pinals DA, Weingartner H, Sirocco K, Missar CD, Pickar D, et al. NMDA receptor function and human cognition: the effects of ketamine in healthy volunteers. Neuropsychopharmacology. 1996;14:301–307. doi: 10.1016/0893-133X(95)00137-3. [DOI] [PubMed] [Google Scholar]
- Malhotra AK, Pinals DA, Adler CM, Elman I, Clifton A, Pickar D, et al. Ketamine-induced exacerbation of psychotic symptoms and cognitive impairment in neuroleptic-free schizophrenics. Neuropsychopharmacology. 1997;17:141–150. doi: 10.1016/S0893-133X(97)00036-5. [DOI] [PubMed] [Google Scholar]
- Marcoux FW, Goodrich JE, Dominick MA. Ketamine prevents ischemic neuronal injury. Brain Res. 1988;452:329–335. doi: 10.1016/0006-8993(88)90037-6. [DOI] [PubMed] [Google Scholar]
- Marsman A, van den Heuvel MP, Klomp DWJ, Kahn RS, Luijten PR, Hulshoff Pol HE. Glutamate in schizophrenia: a focused review and meta-analysis of 1H-MRS studies. Schizophr Bull. 2013;39:120–129. doi: 10.1093/schbul/sbr069. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martin D, Lodge D. Ketamine acts as a non-competitive N-methyl-D-aspartate antagonist on frog spinal cord in vitro. Neuropharmacology. 1985;24:999–1003. doi: 10.1016/0028-3908(85)90128-5. [DOI] [PubMed] [Google Scholar]
- Martin D, Lodge D. Phencyclidine receptors and N-methyl-D-aspartate antagonism: electrophysiologic data correlates with known behaviours. Pharmacol Biochem Behav. 1988;31:279–286. doi: 10.1016/0091-3057(88)90346-2. [DOI] [PubMed] [Google Scholar]
- Martin WR, Fraser HF, Gorodetzky CW, Rosenberg DE. Studies of the dependence-producing potential of the narcotic antagonist 2-cyclopropylmethyl-2′-hydroxy-5,9-dimethyl-6,7-benzomorphan (cyclazocine, WIN-20,740, ARC II-c-3) J Pharmacol Exp Ther. 1965;150:426–436. [PubMed] [Google Scholar]
- Martin WR, Eades CG, Thompson JA, Huppler RE, Gilbert PE. The effects of morphine- and nalorphine- like drugs in the nondependent and morphine-dependent chronic spinal dog. J Pharmacol Exp Ther. 1976;197:517–532. [PubMed] [Google Scholar]
- Mayer ML, Westbrook GL, Guthrie PB. Voltage-dependent block by Mg2+ of NMDA responses in spinal cord neurones. Nature. 1984;309:261–262. doi: 10.1038/309261a0. [DOI] [PubMed] [Google Scholar]
- McCarthy DA, Chen G, Kaump DH, Ensor C. General Anesthetic and Other Pharmacological Properties of 2-(O-Chlorophenyl)-2-Methylamino Cyclohexanone HCl (CI-581) J New Drugs. 1965;5:21–33. doi: 10.1002/j.1552-4604.1965.tb00219.x. [DOI] [PubMed] [Google Scholar]
- McLennan H, Lodge D. The antagonism of amino acid-induced excitation of spinal neurones in the cat. Brain Res. 1979;169:83–90. doi: 10.1016/0006-8993(79)90375-5. [DOI] [PubMed] [Google Scholar]
- McNally JM, McCarley RW, McKenna JT, Yanagawa Y, Brown RE. Complex receptor mediation of acute ketamine application on in vitro gamma oscillations in mouse prefrontal cortex: modeling gamma band oscillation abnormalities in schizophrenia. Neuroscience. 2011;199:51–63. doi: 10.1016/j.neuroscience.2011.10.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mendelsohn LG, Kerchner GA, Kalra V, Zimmerman DM, Leander JD. Phencyclidine receptors in rat brain cortex. Biochem Pharmacol. 1984;33:3529–3535. doi: 10.1016/0006-2952(84)90133-3. [DOI] [PubMed] [Google Scholar]
- Meyer JS, Greifenstein F, Devault M. A new drug causing symptoms of sensory deprivation. J Nerv Ment Dis. 1959;129:54–61. [Google Scholar]
- Millan MJ. The role of monoamines in the actions of established and ‘novel’ antidepressant agents: a critical review. Eur J Pharmacol. 2004;500:371–384. doi: 10.1016/j.ejphar.2004.07.038. [DOI] [PubMed] [Google Scholar]
- Miller OH, Yang L, Wang C-C, Hargroder EA, Zhang Y, Delpire E, et al. GluN2B-containing NMDA receptors regulate depression-like behavior and are critical for the rapid antidepressant actions of ketamine. Elife. 2014;3 doi: 10.7554/eLife.03581. doi: 10.7554/eLife.03581. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Minchin MCW. The effect of anaesthetics on the uptake and release of γ-aminobutyrate and D-aspartate in rat brain slices. Br J Pharmacol. 1981;73:681–689. doi: 10.1111/j.1476-5381.1981.tb16803.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Minzenberg MJ, Firl AJ, Yoon JH, Gomes GC, Reinking C, Carter CS. Gamma oscillatory power is impaired during cognitive control independent of medication status in first-episode schizophrenia. Neuropsychopharmacology. 2010;35:2590–2599. doi: 10.1038/npp.2010.150. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miyasaka M, Domino EF. Neural mechanisms of ketamine-induced anesthesia. Int J Neuropharmacol. 1968;7:557–573. doi: 10.1016/0028-3908(68)90067-1. [DOI] [PubMed] [Google Scholar]
- Moaddel R, Abdrakhmanova G, Kozak J, Jozwiak K, Toll L, Jimenez L, et al. Sub-anesthetic concentrations of (R,S)-ketamine metabolites inhibit acetylcholine-evoked currents in α7 nicotinic acetylcholine receptors. Eur J Pharmacol. 2013;698:228–234. doi: 10.1016/j.ejphar.2012.11.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moeller KE, Lee KC, Kissack JC. Urine drug screening: practical guide for clinicians. Mayo Clin Proc. 2008;83:66–76. doi: 10.4065/83.1.66. [DOI] [PubMed] [Google Scholar]
- Moghaddam B, Adams BW. Reversal of phencyclidine effects by a group II metabotropic glutamate receptor agonist in rats. Science. 1998;281:1349–1352. doi: 10.1126/science.281.5381.1349. [DOI] [PubMed] [Google Scholar]
- Moghaddam B, Krystal JH. Capturing the angel in ‘angel dust’: twenty years of translational neuroscience studies of NMDA receptor antagonists in animals and humans. Schizophr Bull. 2012;38:942–949. doi: 10.1093/schbul/sbs075. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moghaddam B, Adams B, Verma A, Daly D. Activation of glutamatergic neurotransmission by ketamine: a novel step in the pathway from NMDA receptor blockade to dopaminergic and cognitive disruptions associated with the prefrontal cortex. J Neurosci. 1997;17:2921–2927. doi: 10.1523/JNEUROSCI.17-08-02921.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Monaghan DT, Irvine MW, Costa BM, Fang G, Jane DE. Pharmacological modulation of NMDA receptor activity and the advent of negative and positive allosteric modulators. Neurochem Int. 2012;61:581–592. doi: 10.1016/j.neuint.2012.01.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moran RJ, Jones MW, Blockeel AJ, Adams RA, Stephan KE, Friston KJ. Losing control under ketamine: suppressed cortico-hippocampal drive following acute ketamine in rats. Neuropsychopharmacology. 2015;40:268–277. doi: 10.1038/npp.2014.184. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morgan CJA, Curran HV. Ketamine use: a review. Addiction. 2012;107:27–38. doi: 10.1111/j.1360-0443.2011.03576.x. [DOI] [PubMed] [Google Scholar]
- Morris BJ, Cochran SM, Pratt JA. PCP: from pharmacology to modelling schizophrenia. Curr Opin Pharmacol. 2005;5:101–106. doi: 10.1016/j.coph.2004.08.008. [DOI] [PubMed] [Google Scholar]
- Morris H, Wallach J. From PCP to MXE: a comprehensive review of the non-medical use of dissociative drugs. Drug Test Anal. 2014;6:614–632. doi: 10.1002/dta.1620. [DOI] [PubMed] [Google Scholar]
- Morrow BA, Elsworth JD, Roth RH. Repeated phencyclidine in monkeys results in loss of parvalbumin-containing axo-axonic projections in the prefrontal cortex. Psychopharmacology (Berl) 2007;192:283–290. doi: 10.1007/s00213-007-0708-0. [DOI] [PubMed] [Google Scholar]
- Muir KW. Glutamate-based therapeutic approaches: clinical trials with NMDA antagonists. Curr Opin Pharmacol. 2006;6:53–60. doi: 10.1016/j.coph.2005.12.002. [DOI] [PubMed] [Google Scholar]
- Murray TF, Leid ME. Interaction of dextrorotatory opioids with phencyclidine recognition sites in rat brain membranes. Life Sci. 1984;34:1899–1911. doi: 10.1016/0024-3205(84)90121-8. [DOI] [PubMed] [Google Scholar]
- Musaelyan K, Egeland M, Fernandes C, Pariante CM, Zunszain PA, Thuret S. Modulation of adult hippocampal neurogenesis by early-life environmental challenges triggering immune activation. Neural Plast. 2014;2014:194396. doi: 10.1155/2014/194396. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nagele P, Duma A, Kopec M, Gebara MA, Parsoei A, Walker M, et al. Nitrous oxide for treatment-resistant major depression: a proof-of-concept trial. Biol Psychiatry. 2014 doi: 10.1016/j.biopsych.2014.11.016. doi: 10.1016/j.biopsych.2014.11.016. [DOI] [PubMed] [Google Scholar]
- Nakazawa K, Zsiros V, Jiang Z, Nakao K, Kolata S, Zhang S, et al. GABAergic interneuron origin of schizophrenia pathophysiology. Neuropharmacology. 2012;62:1574–1583. doi: 10.1016/j.neuropharm.2011.01.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Namba T, Ming G-L, Song H, Waga C, Enomoto A, Kaibuchi K, et al. NMDA receptor regulates migration of newly generated neurons in the adult hippocampus via Disrupted-In-Schizophrenia 1 (DISC1) J Neurochem. 2011;118:34–44. doi: 10.1111/j.1471-4159.2011.07282.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Neill JC, Barnes S, Cook S, Grayson B, Idris NF, McLean SL, et al. Animal models of cognitive dysfunction and negative symptoms of schizophrenia: focus on NMDA receptor antagonism. Pharmacol Ther. 2010;128:419–432. doi: 10.1016/j.pharmthera.2010.07.004. [DOI] [PubMed] [Google Scholar]
- Nelson SR, Howard RB, Cross RS, Samson F. Ketamine-induced changes in regional glucose utilization in the rat brain. Anesthesiology. 1980;52:330–334. doi: 10.1097/00000542-198004000-00009. [DOI] [PubMed] [Google Scholar]
- Newcomer JW, Farber NB, Jevtovic-Todorovic V, Selke G, Melson AK, Hershey T, et al. Ketamine-induced NMDA receptor hypofunction as a model of memory impairment and psychosis. Neuropsychopharmacology. 1999;20:106–118. doi: 10.1016/S0893-133X(98)00067-0. [DOI] [PubMed] [Google Scholar]
- Nicholson KL, Mansbach RS, Menniti FS, Balster RL. The phencyclidine-like discriminative stimulus effects and reinforcing properties of the NR2B-selective N-methyl-D-aspartate antagonist CP-101 606 in rats and rhesus monkeys. Behav Pharmacol. 2007;18:731–743. doi: 10.1097/FBP.0b013e3282f14ed6. [DOI] [PubMed] [Google Scholar]
- Niesters M, Martini C, Dahan A. Ketamine for chronic pain: risks and benefits. Br J Clin Pharmacol. 2013;77:357–367. doi: 10.1111/bcp.12094. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nowak L, Bregestovski P, Ascher P. Magnesium gates glutamate-activated channels in mouse central neurones. Nature. 1984;307:462–465. doi: 10.1038/307462a0. [DOI] [PubMed] [Google Scholar]
- Nutt D, King LA, Saulsbury W, Blakemore C. Development of a rational scale to assess the harm of drugs of potential misuse. Lancet. 2007;369:1047–1053. doi: 10.1016/S0140-6736(07)60464-4. [DOI] [PubMed] [Google Scholar]
- Office for National Statistics. 2013. Drug related deaths involving ketamine by age group. [online] Available at: http://www.ons.gov.uk/ons/search/index.html?newquery=drug-related+death (accessed 2/25 2015)
- Olney JW, Farber NB. NMDA antagonists as neurotherapeutic drugs, psychotogens, neurotoxins, and research tools for studying schizophrenia. Neuropsychopharmacology. 1995;13:335–345. doi: 10.1016/0893-133X(95)00079-S. [DOI] [PubMed] [Google Scholar]
- Olney JW, Farber NB. Discussion of Bogerts’ temporolimbic system theory of paranoid schizophrenia. Schizophr Bull. 1997;23:533–536. doi: 10.1093/schbul/23.3.533. [DOI] [PubMed] [Google Scholar]
- Olney JW, Price MT, Fuller TA, Labruyere J, Samson L, Carpenter M, et al. The anti-excitotoxic effects of certain anesthetics, analgesics and sedative-hypnotics. Neurosci Lett. 1986;68:29–34. doi: 10.1016/0304-3940(86)90224-7. [DOI] [PubMed] [Google Scholar]
- Olney JW, Labruyere J, Price MT. Pathological changes induced in cerebrocortical neurons by phencyclidine and related drugs. Science. 1989;244:1360–1362. doi: 10.1126/science.2660263. [DOI] [PubMed] [Google Scholar]
- Park CK, Nehls DG, Graham DI, Teasdale GM, McCulloch J. Focal cerebral ischaemia in the cat: treatment with the glutamate antagonist MK-801 after induction of ischaemia. J Cereb Blood Flow Metab. 1988a;8:757–762. doi: 10.1038/jcbfm.1988.124. [DOI] [PubMed] [Google Scholar]
- Park CK, Nehls DG, Graham DI, Teasdale GM, McCulloch J. The glutamate antagonist MK-801 reduces focal ischemic brain damage in the rat. Ann Neurol. 1988b;24:543–551. doi: 10.1002/ana.410240411. [DOI] [PubMed] [Google Scholar]
- Parsons CG, Gruner R, Rozental J, Millar J, Lodge D. Patch clamp studies on the kinetics and selectivity of N-methyl-D-aspartate receptor antagonism by memantine (1-amino-3,5-dimethyladamantan) Neuropharmacology. 1993;32:1337–1350. doi: 10.1016/0028-3908(93)90029-3. [DOI] [PubMed] [Google Scholar]
- Parsons CG, Quack G, Bresink I, Baran L, Przegalinski E, Kostowski W, et al. Comparison of the potency, kinetics and voltage-dependency of a series of uncompetitive NMDA receptor antagonists in vitro with anticonvulsive and motor impairment activity in vivo. Neuropharmacology. 1995;34:1239–1258. doi: 10.1016/0028-3908(95)00092-k. [DOI] [PubMed] [Google Scholar]
- Patil ST, Zhang L, Martenyi F, Lowe SL, Jackson KA, Andreev B, V, et al. Activation of mGlu2/3 receptors as a new approach to treat schizophrenia: a randomized Phase 2 clinical trial. Nat Med. 2007;13:1102–1107. doi: 10.1038/nm1632. [DOI] [PubMed] [Google Scholar]
- Pawson AJ, Sharman JL, Benson HE, Faccenda E, Alexander SP, Buneman OP, et al. NC-IUPHAR. The IUPHAR/BPS Guide to PHARMACOLOGY: an expert-driven knowledgebase of drug targets and their ligands. Nucl Acids Res. 2014;42(Database Issue):D1098–1106. doi: 10.1093/nar/gkt1143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pearlman DM, Najjar S. Meta-analysis of the association between N-methyl-d-aspartate receptor antibodies and schizophrenia, schizoaffective disorder, bipolar disorder, and major depressive disorder. Schizophr Res. 2014;157:249–258. doi: 10.1016/j.schres.2014.05.001. [DOI] [PubMed] [Google Scholar]
- Persson J. Ketamine in pain management. CNS Neurosci Ther. 2013;19:396–402. doi: 10.1111/cns.12111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Petersen RC, Stillman RC. Phencyclidine abuse: an appraisal. Natl Inst Drug Abus Res Monogr. 1978;21:1–17. [PubMed] [Google Scholar]
- Pierri JN, Chaudry AS, Woo TU, Lewis DA. Alterations in chandelier neuron axon terminals in the prefrontal cortex of schizophrenic subjects. Am J Psychiatry. 1999;156:1709–1719. doi: 10.1176/ajp.156.11.1709. [DOI] [PubMed] [Google Scholar]
- Poels EMP, Kegeles LS, Kantrowitz JT, Javitt DC, Lieberman JA, Abi-Dargham A, et al. Glutamatergic abnormalities in schizophrenia: a review of proton MRS findings. Schizophr Res. 2014;152:325–332. doi: 10.1016/j.schres.2013.12.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Powell SB, Sejnowski TJ, Behrens MM. Behavioral and neurochemical consequences of cortical oxidative stress on parvalbumin-interneuron maturation in rodent models of schizophrenia. Neuropharmacology. 2012;62:1322–1331. doi: 10.1016/j.neuropharm.2011.01.049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pratt J, Winchester C, Dawson N, Morris B. Advancing schizophrenia drug discovery: optimizing rodent models to bridge the translational gap. Nat Rev Drug Discov. 2012;11:560–579. doi: 10.1038/nrd3649. [DOI] [PubMed] [Google Scholar]
- Preskorn SH, Baker B, Kolluri S, Menniti FS, Krams M, Landen JW. An innovative design to establish proof of concept of the antidepressant effects of the NR2B subunit selective N-methyl-D-aspartate antagonist, CP-101,606, in patients with treatment-refractory major depressive disorder. J Clin Psychopharmacol. 2008;28:631–637. doi: 10.1097/JCP.0b013e31818a6cea. [DOI] [PubMed] [Google Scholar]
- Quirion R, Hammer RP, Herkenham M, Pert CB. Phencyclidine (angel dust)/sigma ‘opiate’ receptor: visualization by tritium-sensitive film. Proc Natl Acad Sci U S A. 1981;78:5881–5885. doi: 10.1073/pnas.78.9.5881. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rappaport BA, Suresh S, Hertz S, Evers AS, Orser BA. Anesthetic neurotoxicity – clinical implications of animal models. N Engl J Med. 2015;372:796–797. doi: 10.1056/NEJMp1414786. [DOI] [PubMed] [Google Scholar]
- Reynolds GP, Abdul-Monim Z, Neill JC, Zhang Z-J. Calcium binding protein markers of GABA deficits in schizophrenia – postmortem studies and animal models. Neurotox Res. 2004;6:57–61. doi: 10.1007/BF03033297. [DOI] [PubMed] [Google Scholar]
- Reynolds IJ, Miller RJ. Tricyclic antidepressants block N-methyl-D-aspartate receptors: similarities to the action of zinc. Br J Pharmacol. 1988;95:95–102. doi: 10.1111/j.1476-5381.1988.tb16552.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rodefer JS, Nguyen TN, Karlsson J-J, Arnt J. Reversal of subchronic PCP-induced deficits in attentional set shifting in rats by sertindole and a 5-HT6 receptor antagonist: comparison among antipsychotics. Neuropsychopharmacology. 2008;33:2657–2666. doi: 10.1038/sj.npp.1301654. [DOI] [PubMed] [Google Scholar]
- Rodin EA, Luby ED, Meyer JS. Electroencephalographic findings associated with sernyl infusion. Electroencephalogr Clin Neurophysiol. 1959;11:796–798. doi: 10.1016/0013-4694(59)90122-1. [DOI] [PubMed] [Google Scholar]
- Roopun AK, Cunningham MO, Racca C, Alter K, Traub RD, Whittington MA. Region-specific changes in gamma and beta2 rhythms in NMDA receptor dysfunction models of schizophrenia. Schizophr Bull. 2008;34:962–973. doi: 10.1093/schbul/sbn059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roth BL, Gibbons S, Arunotayanun W, Huang X-P, Setola V, Treble R, et al. The ketamine analogue methoxetamine and 3- and 4-methoxy analogues of phencyclidine are high affinity and selective ligands for the glutamate NMDA receptor. PLoS ONE. 2013;8:e59334. doi: 10.1371/journal.pone.0059334. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ryder S, Way WL, Trevor AJ. Comparative pharmacology of the optical isomers of ketamine in mice. Eur J Pharmacol. 1978;49:15–23. doi: 10.1016/0014-2999(78)90217-0. [DOI] [PubMed] [Google Scholar]
- Sabbagh JJ, Murtishaw AS, Bolton MM, Heaney CF, Langhardt M, Kinney JW. Chronic ketamine produces altered distribution of parvalbumin-positive cells in the hippocampus of adult rats. Neurosci Lett. 2013;550:69–74. doi: 10.1016/j.neulet.2013.06.040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saunders JA, Gandal MJ, Siegel SJ. NMDA antagonists recreate signal-to-noise ratio and timing perturbations present in schizophrenia. Neurobiol Dis. 2012;46:93–100. doi: 10.1016/j.nbd.2011.12.049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schildkraut JJ, Kety SS. Biogenic amines and emotion. Science. 1967;156:21–37. doi: 10.1126/science.156.3771.21. [DOI] [PubMed] [Google Scholar]
- Seeman P, Guan H-C, Hirbec H. Dopamine D2 high receptors stimulated by phencyclidines, lysergic acid diethylamide, salvinorin A, and modafinil. Synapse. 2009;63:698–704. doi: 10.1002/syn.20647. [DOI] [PubMed] [Google Scholar]
- Serefko A, Szopa A, Wlaź P, Nowak G, Radziwoń-Zaleska M, Skalski M, et al. Magnesium in depression. Pharmacol Reports. 2013;65:547–554. doi: 10.1016/s1734-1140(13)71032-6. [DOI] [PubMed] [Google Scholar]
- Sernagor E, Kuhn D, Vyklicky L, Mayer ML. Open channel block of NMDA receptor responses evoked by tricyclic antidepressants. Neuron. 1989;2:1221–1227. doi: 10.1016/0896-6273(89)90306-1. [DOI] [PubMed] [Google Scholar]
- Shahani R, Streutker C, Dickson B, Stewart RJ. Ketamine-associated ulcerative cystitis: a new clinical entity. Urology. 2007;69:810–812. doi: 10.1016/j.urology.2007.01.038. [DOI] [PubMed] [Google Scholar]
- Shannon HE. Evaluation of phencyclidine analogs on the basis of their discriminative stimulus properties in the rat. J Pharmacol Exp Ther. 1981;216:543–551. [PubMed] [Google Scholar]
- Shannon HE. Pharmacological analysis of the phencyclidine-like discriminative stimulus properties of narcotic derivatives in rats. J Pharmacol Exp Ther. 1982a;222:146–151. [PubMed] [Google Scholar]
- Shannon HE. Phencyclidine-like discriminative stimuli of (+)- and (−)-N-allylnormetazocine in rats. Eur J Pharmacol. 1982b;84:225–228. doi: 10.1016/0014-2999(82)90207-2. [DOI] [PubMed] [Google Scholar]
- Shannon HE. Pharmacological evaluation of N-allynormetazocine (SKF 10,047) on the basis of its discriminative stimulus properties in the rat. J Pharmacol Exp Ther. 1983;225:144–152. [PubMed] [Google Scholar]
- Sills MA, Loo PS. Tricyclic antidepressants and dextromethorphan bind with higher affinity to the phencyclidine receptor in the absence of magnesium and L-glutamate. Mol Pharmacol. 1989;36:160–165. [PubMed] [Google Scholar]
- Simon RP, Swan JH, Griffiths T, Meldrum BS. Blockade of N-methyl-D-aspartate receptors may protect against ischemic damage in the brain. Science. 1984;226:850–852. doi: 10.1126/science.6093256. [DOI] [PubMed] [Google Scholar]
- Sircar R, Zukin SR. Characterization of specific sigma opiate/phencyclidine (PCP)-binding sites in the human brain. Life Sci. 1983;33(Suppl. 1):259–262. doi: 10.1016/0024-3205(83)90492-7. [DOI] [PubMed] [Google Scholar]
- Sircar R, Rappaport M, Nichtenhauser R, Zukin SR. The novel anticonvulsant MK-801: a potent and specific ligand of the brain phencyclidine/sigma-receptor. Brain Res. 1987;435:235–240. doi: 10.1016/0006-8993(87)91606-4. [DOI] [PubMed] [Google Scholar]
- Slifer BL, Balster RL. Phencyclidine-like discriminative stimulus effects of the stereoisomers of alpha- and beta-cyclazocine in rats. J Pharmacol Exp Ther. 1988;244:606–612. [PubMed] [Google Scholar]
- Smith RC, Meltzer HY, Arora RC, Davis JM. Effects of phencyclidine on [3H]catecholamine and [3H]serotonin uptake in synaptosomal preparations from rat brain. Biochem Pharmacol. 1977;26:1435–1439. doi: 10.1016/0006-2952(77)90370-7. [DOI] [PubMed] [Google Scholar]
- Snell LD, Johnson KM. Antagonism of N-methyl-D-aspartate-induced transmitter release in the rat striatum by phencyclidine-like drugs and its relationship to turning behavior. J Pharmacol Exp Ther. 1985;235:50–57. [PubMed] [Google Scholar]
- Snyder MA, Gao W-J. NMDA hypofunction as a convergence point for progression and symptoms of schizophrenia. Front Cell Neurosci. 2013;7:31. doi: 10.3389/fncel.2013.00031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stringer JL, Guyenet PG. Elimination of long-term potentiation in the hippocampus by phencyclidine and ketamine. Brain Res. 1983;258:159–164. doi: 10.1016/0006-8993(83)91244-1. [DOI] [PubMed] [Google Scholar]
- Takagi S, Balu DT, Coyle JT. Subchronic pharmacological and chronic genetic NMDA receptor hypofunction differentially regulate the Akt signaling pathway and Arc expression in juvenile and adult mice. Schizophr Res. 2015;162:216–221. doi: 10.1016/j.schres.2014.12.034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tang AH, Schroeder LA. Spinal-cord depressant effects of ketamine and etoxadrol in the cat and the rat. Anesthesiology. 1973;39:37–43. doi: 10.1097/00000542-197307000-00007. [DOI] [PubMed] [Google Scholar]
- Tang AH, Cangelosi AA, Code RA, Franklin SR. Phencyclidine-like behavioral effects of 2-methyl-3,3-diphenyl-3-propanolamine (2-MDP) Pharmacol Biochem Behav. 1984;20:209–213. doi: 10.1016/0091-3057(84)90244-2. [DOI] [PubMed] [Google Scholar]
- Taube HD, Montel H, Hau G, Starke K. Phencyclidine and ketamine: comparison with the effect of cocaine on the noradrenergic neurones of the rat brain cortex. Naunyn Schmiedebergs Arch Pharmacol. 1975;291:47–54. doi: 10.1007/BF00510820. [DOI] [PubMed] [Google Scholar]
- Teasell RW, Mehta S, Aubut J-AL, Foulon B, Wolfe DL, Hsieh JTC, et al. A systematic review of pharmacologic treatments of pain after spinal cord injury. Arch Phys Med Rehabil. 2010;91:816–831. doi: 10.1016/j.apmr.2010.01.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Todd SL, Balster RL, Martin BR. Affinity of the enantiomers of alpha- and beta-cyclazocine for binding to the phencyclidine and mu opioid receptors. Life Sci. 1990;46:895–901. doi: 10.1016/0024-3205(90)90120-g. [DOI] [PubMed] [Google Scholar]
- Tokay T, Rohde M, Krabbe S, Rehberg M, Bender RA, Köhling R, et al. HCN1 channels constrain DHPG-induced LTD at hippocampal Schaffer collateral-CA1 synapses. Learn Mem. 2009;16:769–776. doi: 10.1101/lm.1556009. [DOI] [PubMed] [Google Scholar]
- Toriumi K, Mouri A, Narusawa S, Aoyama Y, Ikawa N, Lu L, et al. Prenatal NMDA receptor antagonism impaired proliferation of neuronal progenitor, leading to fewer glutamatergic neurons in the prefrontal cortex. Neuropsychopharmacology. 2012;37:1387–1396. doi: 10.1038/npp.2011.324. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tricklebank MD, Singh L, Oles RJ, Preston C, Iversen SD. The behavioural effects of MK-801: a comparison with antagonists acting non-competitively and competitively at the NMDA receptor. Eur J Pharmacol. 1989;167:127–135. doi: 10.1016/0014-2999(89)90754-1. [DOI] [PubMed] [Google Scholar]
- Troyano-Rodriguez E, Lladó-Pelfort L, Santana N, Teruel-Martí V, Celada P, Artigas F. Phencyclidine inhibits the activity of thalamic reticular gamma-aminobutyric acidergic neurons in rat brain. Biol Psychiatry. 2014;76:937–945. doi: 10.1016/j.biopsych.2014.05.019. [DOI] [PubMed] [Google Scholar]
- Trullas R, Skolnick P. Functional antagonists at the NMDA receptor complex exhibit antidepressant actions. Eur J Pharmacol. 1990;185:1–10. doi: 10.1016/0014-2999(90)90204-j. [DOI] [PubMed] [Google Scholar]
- Tsai G, Coyle JT. Glutamatergic mechanisms in schizophrenia. Annu Rev Pharmacol Toxicol. 2002;42:165–179. doi: 10.1146/annurev.pharmtox.42.082701.160735. [DOI] [PubMed] [Google Scholar]
- Uchida T, Furukawa T, Iwata S, Yanagawa Y, Fukuda A. Selective loss of parvalbumin-positive GABAergic interneurons in the cerebral cortex of maternally stressed Gad1-heterozygous mouse offspring. Transl Psychiatry. 2014;4:e371. doi: 10.1038/tp.2014.13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Uhlhaas PJ, Singer W. Abnormal neural oscillations and synchrony in schizophrenia. Nat Rev Neurosci. 2010;11:100–113. doi: 10.1038/nrn2774. [DOI] [PubMed] [Google Scholar]
- Umbricht D, Krljes S. Mismatch negativity in schizophrenia: a meta-analysis. Schizophr Res. 2005;76:1–23. doi: 10.1016/j.schres.2004.12.002. [DOI] [PubMed] [Google Scholar]
- Verma A, Moghaddam B. NMDA receptor antagonists impair prefrontal cortex function as assessed via spatial delayed alternation performance in rats: modulation by dopamine. J Neurosci. 1996;16:373–379. doi: 10.1523/JNEUROSCI.16-01-00373.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vickroy TW, Johnson KM. In vivo administration of phencyclidine inhibits 3H-dopamine accumulation by rat brain striatal slices. Subst Alcohol Actions Misuse. 1980;1:351–354. [PubMed] [Google Scholar]
- Vickroy TW, Johnson KM. Similar dopamine-releasing effects of phencyclidine and nonamphetamine stimulants in striatal slices. J Pharmacol Exp Ther. 1982;223:669–674. [PubMed] [Google Scholar]
- Vincent JP, Cavey D, Kamenka JM, Geneste P, Lazdunski M. Interaction of phencyclidines with the muscarinic and opiate receptors in the central nervous system. Brain Res. 1978;152:176–182. doi: 10.1016/0006-8993(78)90145-2. [DOI] [PubMed] [Google Scholar]
- Vincent JP, Kartalovski B, Geneste P, Kamenka JM, Lazdunski M. Interaction of phencyclidine (‘angel dust’) with a specific receptor in rat brain membranes. Proc Natl Acad Sci U S A. 1979;76:4678–4682. doi: 10.1073/pnas.76.9.4678. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Volk DW, Pierri JN, Fritschy J-M, Auh S, Sampson AR, Lewis DA. Reciprocal alterations in pre- and postsynaptic inhibitory markers at chandelier cell inputs to pyramidal neurons in schizophrenia. Cereb Cortex. 2002;12:1063–1070. doi: 10.1093/cercor/12.10.1063. [DOI] [PubMed] [Google Scholar]
- Vollenweider FX, Leenders KL, Oye I, Hell D, Angst J. Differential psychopathology and patterns of cerebral glucose utilisation produced by (S)- and (R)-ketamine in healthy volunteers using positron emission tomography (PET) Eur Neuropsychopharmacol. 1997;7:25–38. doi: 10.1016/s0924-977x(96)00042-9. [DOI] [PubMed] [Google Scholar]
- Wang C, McInnis J, Ross-Sanchez M, Shinnick-Gallagher P, Wiley JL, Johnson KM. Long-term behavioral and neurodegenerative effects of perinatal phencyclidine administration: implications for schizophrenia. Neuroscience. 2001;107:535–550. doi: 10.1016/s0306-4522(01)00384-0. [DOI] [PubMed] [Google Scholar]
- Wang CZ, Yang SF, Xia Y, Johnson KM. Postnatal phencyclidine administration selectively reduces adult cortical parvalbumin-containing interneurons. Neuropsychopharmacology. 2008;33:2442–2455. doi: 10.1038/sj.npp.1301647. [DOI] [PubMed] [Google Scholar]
- Watanabe Y, Saito H, Abe K. Tricyclic antidepressants block NMDA receptor-mediated synaptic responses and induction of long-term potentiation in rat hippocampal slices. Neuropharmacology. 1993;32:479–486. doi: 10.1016/0028-3908(93)90173-z. [DOI] [PubMed] [Google Scholar]
- Watkins JC, Evans RH. Excitatory amino acid transmitters. Annu Rev Pharmacol Toxicol. 1981;21:165–204. doi: 10.1146/annurev.pa.21.040181.001121. [DOI] [PubMed] [Google Scholar]
- Wei J, Graziane NM, Wang H, Zhong P, Wang Q, Liu W, et al. Regulation of N-methyl-D-aspartate receptors by disrupted-in-schizophrenia-1. Biol Psychiatry. 2014;75:414–424. doi: 10.1016/j.biopsych.2013.06.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weickert CS, Fung SJ, Catts VS, Schofield PR, Allen KM, Moore LT, et al. Molecular evidence of N-methyl-D-aspartate receptor hypofunction in schizophrenia. Mol Psychiatry. 2013;18:1185–1192. doi: 10.1038/mp.2012.137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weinberger DR. On localizing schizophrenic neuropathology. Schizophr Bull. 1997;23:537–540. doi: 10.1093/schbul/23.3.537. [DOI] [PubMed] [Google Scholar]
- White JM, Holtzman SG. Properties of pentazocine as a discriminative stimulus in the squirrel monkey. J Pharmacol Exp Ther. 1982;223:396–401. [PubMed] [Google Scholar]
- White PF, Way WL, Trevor AJ. Ketamine – Its pharmacology and therapeutic uses. Anesthesiology. 1982;56:119–136. doi: 10.1097/00000542-198202000-00007. [DOI] [PubMed] [Google Scholar]
- Willetts J, Balster RL. Phencyclidine-like discriminative stimulus properties of MK-801 in rats. Eur J Pharmacol. 1988;146:167–169. doi: 10.1016/0014-2999(88)90498-0. [DOI] [PubMed] [Google Scholar]
- Willetts J, Balster RL, Leander JD. The behavioral pharmacology of NMDA receptor antagonists. Trends Pharmacol Sci. 1990;11:423–428. doi: 10.1016/0165-6147(90)90150-7. [DOI] [PubMed] [Google Scholar]
- Wilson RD, Traber DL, Barratt E, Creson DL, Schmitt RC, Allen CR. Evaluation of CL-1848C: a new dissociative anesthetic in normal human volunteers. Anesth Analg. 1970;49:236–241. [PubMed] [Google Scholar]
- Wise RA. Addictive drugs and brain stimulation reward. Annu Rev Neurosci. 1996;19:319–340. doi: 10.1146/annurev.ne.19.030196.001535. [DOI] [PubMed] [Google Scholar]
- Wong EH, Kemp JA, Priestley T, Knight AR, Woodruff GN, Iversen LL. The anticonvulsant MK-801 is a potent N-methyl-D-aspartate antagonist. Proc Natl Acad Sci U S A. 1986;83:7104–7108. doi: 10.1073/pnas.83.18.7104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Woo TU, Whitehead RE, Melchitzky DS, Lewis DA. A subclass of prefrontal gamma-aminobutyric acid axon terminals are selectively altered in schizophrenia. Proc Natl Acad Sci U S A. 1998;95:5341–5346. doi: 10.1073/pnas.95.9.5341. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yamakura T, Mori H, Masaki H, Shimoji K, Mishina M. Different sensitivities of NMDA receptor channel subtypes to non-competitive antagonists. Neuroreport. 1993;4:687–690. doi: 10.1097/00001756-199306000-00021. [DOI] [PubMed] [Google Scholar]
- Yamakura T, Chavez-Noriega LE, Harris RA. Subunit-dependent inhibition of human neuronal nicotinic acetylcholine receptors and other ligand-gated ion channels by dissociative anesthetics ketamine and dizocilpine. Anesthesiology. 2000;92:1144–1153. doi: 10.1097/00000542-200004000-00033. [DOI] [PubMed] [Google Scholar]
- Yamamoto H, Kamegaya E, Sawada W, Hasegawa R, Yamamoto T, Hagino Y, et al. Involvement of the N-methyl-D-aspartate receptor GluN2D subunit in phencyclidine-induced motor impairment, gene expression, and increased Fos immunoreactivity. Mol Brain. 2013;6:56. doi: 10.1186/1756-6606-6-56. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yoshimizu T, Shimazaki T, Ito A, Chaki S. An mGluR2/3 antagonist, MGS0039, exerts antidepressant and anxiolytic effects in behavioral models in rats. Psychopharmacology (Berl) 2006;186:587–593. doi: 10.1007/s00213-006-0390-7. [DOI] [PubMed] [Google Scholar]
- Young J, Geyer M. Developing treatments for cognitive deficits in schizophrenia: the challenge of translation. J Psychopharmacol. 2015;29:178–196. doi: 10.1177/0269881114555252. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zarate C, Duman RS, Liu G, Sartori S, Quiroz J, Murck H. New paradigms for treatment-resistant depression. Ann N Y Acad Sci. 2013;1292:21–31. doi: 10.1111/nyas.12223. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zarate CA, Singh JB, Carlson PJ, Brutsche NE, Ameli R, Luckenbaugh DA, et al. A randomized trial of an N-methyl-D-aspartate antagonist in treatment-resistant major depression. Arch Gen Psychiatry. 2006;63:856–864. doi: 10.1001/archpsyc.63.8.856. [DOI] [PubMed] [Google Scholar]
- Zeiler FA, Teitelbaum J, Gillman LM, West M. NMDA antagonists for refractory seizures. Neurocrit Care. 2014;20:502–513. doi: 10.1007/s12028-013-9939-6. [DOI] [PubMed] [Google Scholar]
- Zukin RS, Zukin SR. Demonstration of [3H]cyclazocine binding to multiple opiate receptor sites. Mol Pharmacol. 1981;20:246–254. [PubMed] [Google Scholar]
- Zukin SR. Differing stereospecificities distinguish opiate receptor subtypes. Life Sci. 1982;31:1307–1310. doi: 10.1016/0024-3205(82)90368-x. [DOI] [PubMed] [Google Scholar]
- Zukin SR, Zukin RS. Specific [3H]phencyclidine binding in rat central nervous system. Proc Natl Acad Sci U S A. 1979;76:5372–5376. doi: 10.1073/pnas.76.10.5372. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zukin SR, Brady KT, Slifer BL, Balster RL. Behavioral and biochemical stereoselectivity of sigma opiate/PCP receptors. Brain Res. 1984;294:174–177. doi: 10.1016/0006-8993(84)91326-x. [DOI] [PubMed] [Google Scholar]