

Abstract
The sweetpotato whitefly, Bemisia tabaci (Gennadius) is fed on by a wide variety of generalist predators, but there is little information on these predator-prey interactions. A laboratory investigation was conducted to quantify the foraging behavior of the adults of five common whitefly predators presented with a surfeit of whitefly eggs, nymphs, and adults. The beetles, Hippodamia convergens Guérin-Méneville and Collops vittatus (Say) fed mostly on whitefly eggs, but readily and rapidly preyed on all of the whitefly lifestages. The true bugs, Geocoris punctipes (Say) and Orius tristicolor (Say) preyed almost exclusively on adult whiteflies, while Lygus hesperus Knight preyed almost exclusively on nymphs. The true bugs had much longer prey handling times than the beetles and spent much more of their time feeding (35–42%) than the beetles (6–7%). These results indicate that generalist predators vary significantly in their interaction with this host, and that foraging behavior should be considered during development of a predator-based biological control program for B. tabaci.
| Abbreviation: | |
|---|---|
| ELISA | enzyme-linked immunosorbent assay |
Keywords: Predator-prey interactions, prey choice, sequence analysis, biological control, insect behavior
Introduction
The sweetpotato whitefly, Bemisia tabaci (Gennadius), strain B (= silverleaf whitefly, B. argentifolii Bellows and Perring) and other whitefly species are worldwide pests of many ornamental, greenhouse, and agricultural crops (Perring 1996; Naranjo and Ellsworth 2001). An enormous effort has been dedicated to studying the natural enemies of these pests (Gerling and Mayer 1996; Gerling et al. 2001). To date, most of the research on whitefly natural enemies has focused on evaluating the behavior and efficacy of parasitoids (Gerling 1986, 1992; Gerling et al. 2001). In contrast, while it is known that many predator species feed on whiteflies (Gerling 1990; Cock 1994; Breene et al. 1994; Nordlund and Legaspi 1996; Gerling et al. 2001), there is little published information on their prey preferences or specific foraging patterns. For example, with the exception of a few lacewing species (Or and Gerling 1985; Gerling 1986, 1990; Butler and Henneberry 1988; Kapadia and Puri 1989; Breene et al. 1992, 1994; Legaspi et al. 1994), very little is known about any predator's prey preference, consumption rate, or handling time for the various whitefly lifestages (Nordlund and Legaspi 1996; Hagler 2002).
Progress toward determining these basic foraging characteristics has been slow, due in part to the difficulty of observing whitefly predation in the field. Whiteflies and many of their predators are small, elusive, and cryptic. Additionally, adult whiteflies and their predators are highly mobile. As a consequence, direct field observations of whitefly predation are tedious, time consuming, and can be disruptive to the normal foraging process (personal observation). Furthermore, continuous observation of an individual predator over an extended period of time is not practical in the field for those species that disperse readily and forage on the underside of the leaf. Because of these technical challenges, indirect techniques are used to examine the influence of whitefly predators. For example, we developed a whitefly-specific enzyme-linked immunosorbent assay (ELISA) for identifying whitefly remains in the guts of field-collected predators. However, this assay had three limitations: (1) the whitefly “egg-specific” monoclonal antibody used in the assay did not differentiate between an egg and a gravid female meal, (2) it did not detect predation on whitefly nymphs and adult males, and (3) the assay was not quantifiable (Hagler et al. 1993). As a consequence, the gut content immunoassay could not detect how many whiteflies were consumed or which lifestage were consumed (i.e., an egg, adult female, or a mixture of both).
This study was conducted to learn more about the interactions of whitefly predators with their host. We directly observed the behaviors of 130 individuals, representing five predator species, in feeding arenas containing an abundance of whitefly eggs, nymphs, and adults on a cotton leaf disk. The whitefly predators examined were: Hippodamia convergens Guérin-Méneville (Coleoptera: Coccinellidae), Collops vittatus (Say) (Coleoptera: Melyridae), Geocoris punctipes (Say) (Heteroptera: Lygaeidae), Orius tristicolor (Say) (Heteroptera: Anthocoridae), and Lygus hesperus Knight (Heteroptera: Miridae). These predators were selected for this study because assays of predator gut content by ELISA have shown that they readily prey on whitefly (Hagler and Naranjo 1994a,b). Moreover, L. hesperus was included because it has been shown that this omnivorous pest is also a prolific whitefly predator (Hagler and Naranjo 1994a). Our specific objectives were to: (1) determine if these predators prefer a particular whitefly life stage, (2) determine the amount of and rate of whitefly consumption, and (3) compare predator foraging patterns between species.
Materials and Methods
Predators
Adult predators of the five test species were collected with 36.0-cm diameter sweep nets from cotton, Gossypium hirsutum L. fields located at The University of Arizona's Agricultural Research Station, Maricopa, Arizona. The entire contents of the sweep net samples (e.g., arthropods and plant debris) were placed in a large communal rearing cage containing a potted cotton plant and held at 27°C in our laboratory. The predators were allowed to feed freely on any available prey collected in the sweep nets. The day before behavioral observations, a single adult predator was removed from the communal rearing container, placed individually into a clean Petri dish (9.0-cm diameter), and provided with only water.
Feeding arena
Single ‘Delta Pine 5415’ cotton plants were grown in 15.2-cm diameter pots in a greenhouse. Plants that were four to five weeks old were infested with adult whiteflies each week. When the plants were approximately eight to nine weeks old, a single leaf was removed from a plant and cut to fit exactly into the bottom of a 3.5-cm diameter plastic Petri dish (the feeding arena). We selected leaves that contained numerous whitefly eggs and nymphs. The number of whitefly eggs and nymphs were counted on each leaf disk and placed abaxial side up into the bottom of the feeding arena. Adult whiteflies were then introduced into the arena and the Petri dish lid was placed over the top of the arena. A typical feeding arena contained a 3.5-cm diameter cotton leaf disk infested with an average (± SD) of 837.2 ± 67.8, 704.4 ± 60.0 and 43.1 ± 1.5 eggs, nymphs, and adults (males and females at a 1:1 sex ratio), respectively.
Behavioral observations
A single, randomly selected predator (each predator species had an approximate sex ratio of 1:1), that had been held overnight without food was placed into the feeding arena and continuously monitored for 1 h under a dissecting microscope. Preliminary observations of the predator's behavior revealed several distinct behaviors that were common for each predator species. Subsequently, a behavioral ethogram (Lehner 1979) was developed and its components were programmed into The Observer®, a software program designed specifically for animal behavior research (Noldus Information Technology, Version 3.0, www.noldus.com). Descriptions of the predator behaviors that were observed and tabulated during this study are given in Table 1. For most of the 1 hour observations a new feeding arena was used for each observation. In a few instances, limitations in the number of available prey and plant material meant that a predator was removed from the arena after a 1 hour observation and replaced with another randomly selected predator of the same species. No more than two predators were observed consecutively in the same arena. All observations were made between 08:00 and 12:00 at 27°C. The feeding arenas were replaced daily with fresh plant and prey material.
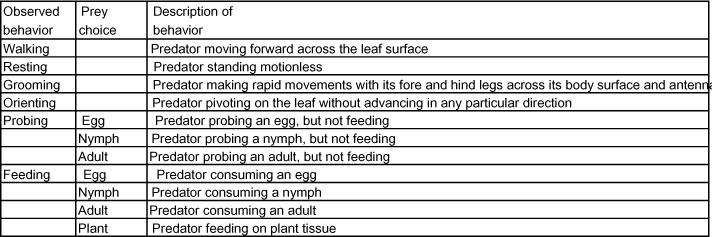
Table 1.
Descriptions of the behavioral events recorded for the predators exposed to a cotton leaf disk containing the various lifestages of Bemisia tabaci.
Data analysis
The prey choice and the amount of time that each predator species spent feeding (i.e., handling time) on the various whitefly lifestages did not conform to the assumption of normality, so a Kruskal-Wallis one-way ANOVA on the ranked data was used to determine significant differences between prey choice and prey handling time for each predator species. For the prey choice analysis, a Student Newman-Keuls multiple comparison test was used to separate statistical differences in prey choice (P = 0.05, SigmaStat Ver. 2.03, SPSS Inc., www.spss.com). For the prey handling time analysis, Dunn's multiple comparison test was used to separate statistical differences in handling time (P = 0.05).
Behavior transition matrices were determined for each predator species by transferring the observational data obtained from the Observer® from columns of sequences into matrices of preceding and succeeding behavioral events as described by Lehner (1979) and Isaacs et al. (1999). Thereafter, each first-order transition (Slater 1973) from one behavior to the other was analyzed by a χ2 test to identify those transitional behaviors that were significantly greater than expected by chance. This was done only for those transitions with a frequency greater than 1.0% of the total number of transitions in order to reduce the likelihood of making type II statistical errors (Isaacs et al. 1999). The critical P-value assigned to the χ2 test was 0.001.
Results
Hippodamia convergens
Adult H. convergens were most commonly observed eating whitefly eggs. On average, 18.3 ± 4.1 eggs, 6.2 ± 1.5 nymphs and 9.6 ± 1.5 adults were consumed per hour of observation (Figure 1A). The time spent feeding on individual whitefly eggs was significantly less than the time spent feeding on nymphs and adults. The average feeding durations were 5.0 ± 0.4, 9.2 ± 0.9, and 11.0 ± 0.4 seconds for eggs, nymphs, and adults, respectively (Figure 1B). Hippodamia convergens was rarely observed eating the cotton leaf.
Figure 1.
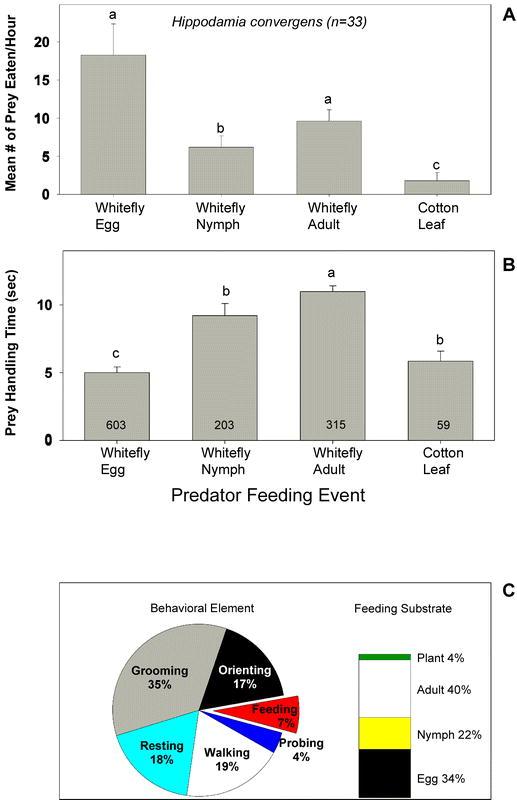
(A) Mean (± SD) frequency of Hippodamia convergens feeding on the various whitefly lifestages or on the cotton leaf. The letters above the error bars indicate significant differences as determined by the Student Newman-Keuls multiple comparison test (P = 0.05, n = 33 individuals observed for 1 hour each). (B) The mean (± SD) amount of time H. convergens spent feeding on the various whitefly lifestages or on the cotton leaf. The numbers inside the vertical bars are the total number of observations recorded during 33 hours of observation. The letters above the error bars indicate significant differences as determined by Dunn's multiple comparison test (P = 0.05). (C) Time budgets for H. convergens in the feeding arena. Results are expressed as the percentage of total time spent in each behavioral element and on each feeding substrate over 33 hours of observation (n = 33 individuals).
Observations revealed that the proportion of time that H. convergens spent on each behavior was grooming (35%) > walking (19%), resting (18%), orienting (17%) > feeding (7%) > probing (4%) (Figure 1C). Even though H. convergens consumed twice as many eggs as adults, the majority of its feeding time (40%) was spent feeding on adults (Figure 1C).
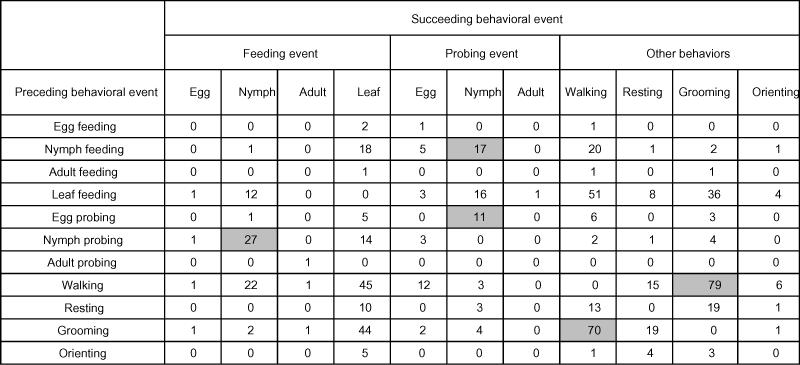
Hippodamia convergens was very active, yielding a total of 3,464 behavioral events during 33 hours of observation (105.0 events per hour). The general sequence of behavioral events exhibited by H. convergens as revealed by the analysis of the transition matrix showed that there is a high likelihood that H. convergens will proceed from:
egg feeding to egg feeding or walking
adult feeding to orienting or walking
nymph feeding to nymph feeding or orienting
leaf feeding to walking
grooming to walking, resting or orienting
orienting to grooming, adult feeding or nymph feeding
adult probing to walking
egg probing to egg feeding
resting to grooming
walking to grooming, adult feeding, adult probing or leaf feeding.
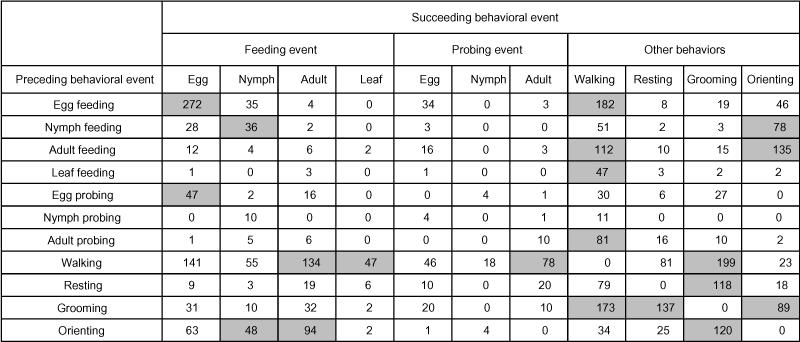
The most common behavioral sequence recorded was egg feeding followed by an additional egg feeding 7.8% (e.g., 272 of the 3464 behavioral events) of the time (Table 2). If an individual ate more than one egg in succession, an average of 3.1 ± 1.2 (range 2 to 9) eggs were consecutively consumed. Interestingly, another of the significant transitions was nymph feeding followed immediately by another nymph feeding 1.0% of the time. If an individual ate more than one nymph in succession, an average of 2.7 ± 0.9 (range 2 to 5) nymphs were consecutively consumed.
Table 2.
Frequencies of transitions between all observed behavioral events for adult Hippodamia convergens on cotton leaves that also contained whitefly eggs, nymphs, and adults. Transitions that were significantly more common than expected by chance are shown in grey boxes (chi-square, adjusted P<0.001).
Collops vittatus
Adult C. vittatus were most commonly observed feeding on whitefly eggs. On average, 13.0 ± 5.6 eggs, 3.6 ± 1.2 nymphs, and 7.6 ± 1.1 adults were consumed per hour of observation (Figure 2A). The feeding duration for individual eggs was significantly less than for whitefly nymphs and adults (Figure 2B). The average feeding durations were 4.6 ± 0.5, 6.6 ± 1.0, and 14.3 ± 0.6 seconds for eggs, nymphs, and adults, respectively (Figure 2B). C. vittatus was rarely observed eating the cotton leaf (Figure 2B).
Figure 2.
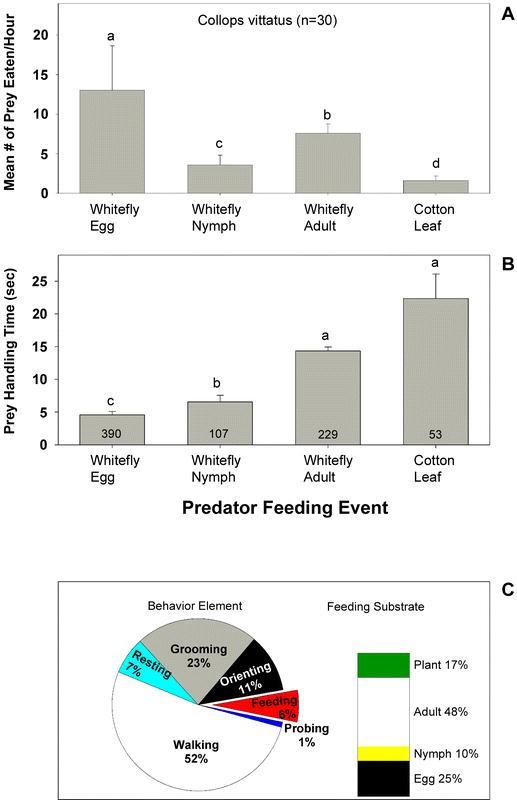
(A) Mean (± SD) frequency of Collops vittatus feeding on the various whitefly lifestages or on the cotton leaf. The letters above the error bars indicate significant differences as determined by the Student Newman-Keuls multiple comparison test (P = 0.05, n = 30 individuals observed for 1 hour each). (B) The mean (± SD) amount of time C. vittatus spent feeding on the various whitefly lifestages or on the cotton leaf. The numbers inside the vertical bars are the total number of observations recorded during 30 hours of observation. The letters above the error bars indicate significant differences as determined by Dunn's multiple comparison test (P = 0.05). (C) Time budgets for C. vittatus in the feeding arena. Results are expressed as the percentage of total time spent in each behavioral element and on each feeding substrate over 30 hours of observation (n = 30 individuals).
Observations revealed that the proportion of time that C. vittatus spent on each behavior was walking (52%) > grooming (23%) > orienting (11%) > resting (7%), feeding (6%) > probing (1%) (Figure 2C). Although C. vittatus ate significantly more eggs than adults, almost one half of its feeding time was spent consuming adult whiteflies (Figure 2C).
C. vittatus was very active yielding a total of 2,571 behavioral events during 30 hours of observation (85.7 events per hour). The behavioral sequence analysis revealed that there is a high likelihood that C. vittatus will proceed from:
adult feeding to walking and orienting
egg feeding to egg feeding, walking or orienting
nymph feeding to nymph feeding
leaf feeding to walking
grooming to walking
orienting to egg feeding
resting to walking and grooming
walking to grooming, adult feeding, resting, egg feeding or leaf feeding (Table 3).
The most common behavioral sequences recorded were walking followed by grooming 13.0% of the time and grooming followed by walking 12.6% of the time. As with H. convergens, egg feeding followed immediately by another egg feeding (8.9% of the time) and nymph feeding followed immediately by another nymph feeding (1.1% of the time) were significant transitional behaviors. On average, if an individual ate more than one particular prey type in succession, 3.5 ± 1.7 (range 2 to 15) eggs and 2.4 ± 0.5 (range 2 to 3) nymphs were consecutively eaten.
Table 3.
Frequencies of transitions between all observed behavioral events for adult Collops vittatus on cotton leaves that also contained whitefly eggs, nymphs, and adults. Transitions that were significantly more common than expected by chance are shown in grey boxes (chi-square, adjusted P<0.001).
Geocoris punctipes
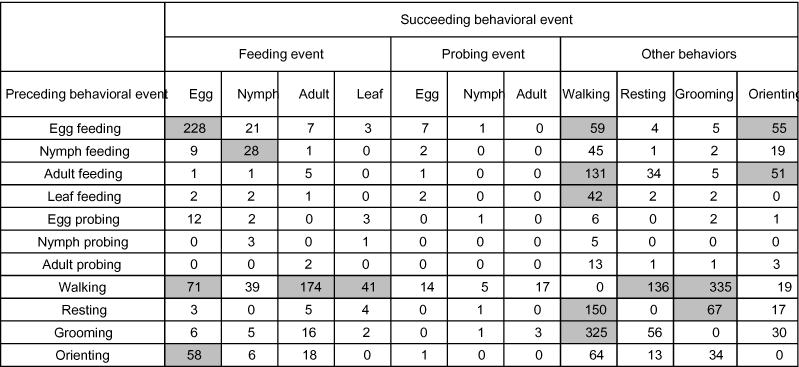
Adult G. punctipes were commonly observed feeding (9.8 ± 2.7 times per hour) on the cotton leaf for extended periods of time (98.0 ± 17.4 seconds per feeding event) (Figure 3A–B). The frequency of predation on adult whiteflies was significantly greater than on eggs and nymphs (Figure 3B). Very few whitefly eggs and nymphs were eaten during the 18 hours of observation. On average, 0.1 ± 0.1 eggs, 1.1 ± 0.3 nymphs, and 3.7 ± 1.0 adults, were consumed per hour of observation. The average feeding duration was 8.5 ± 0.5, 54.0 ±11.8, and 111.5 ± 9.8 seconds for eggs, nymphs, and adults, respectively (Figure 3B).
Figure 3.

(A) Mean (±S D) frequency of Geocoris punctipes feeding on the various whitefly lifestages or on the cotton leaf. The letters above the error bars indicate significant differences as determined by the Student Newman-Keuls multiple comparison test (P = 0.05) (n = 18 individuals observed for 1 hour each). (B) The mean (± SD) amount of time G. punctipes spent feeding on the various whitefly lifestages or on the cotton leaf. The numbers inside the vertical bars are the total number of observations recorded during 18 hours of observation. The letters above the error bars indicate significant differences as determined by Dunn's multiple comparison test (P = 0.05). (C) Time budgets for G. punctipes in the feeding arena. Results are expressed as the percentage of total time spent in each behavioral element and on each feeding substrate over 18 hours of observation (n = 18 individuals).
Observations revealed that the proportion of time that G. punctipes spent on each behavior was feeding (40%) > resting (20%), walking (19%), grooming (18%) > orienting (2%) probing (1%) (Figure 3C). Overall, G. punctipes spent two-thirds of its feeding time eating the cotton leaf.
G. punctipes was an active predator yielding a total of 1,554 behavioral events during 18 hours of observation (86.3 events per hour). The general sequence of behavioral events exhibited by G. punctipes as revealed by the analysis of the transition matrix showed that there is a high likelihood that G. punctipes will proceed from:
leaf feeding to orienting
grooming to resting
orienting to leaf feeding
adult probing to adult feeding
resting to walking or grooming
walking to resting.
The most common sequence recorded was walking followed by resting 13.4% of the time, and resting followed by walking 12.4% of the time (Table 4).
Table 4.
Frequencies of transitions between all observed behavioral events for adult Geocoris punctipes on cotton leaves that also contained whitefly eggs, nymphs, and adults. Transitions that were significantly more common than expected by chance are shown in grey boxes (chi-square, adjusted P<0.001).
Orius tristicolor
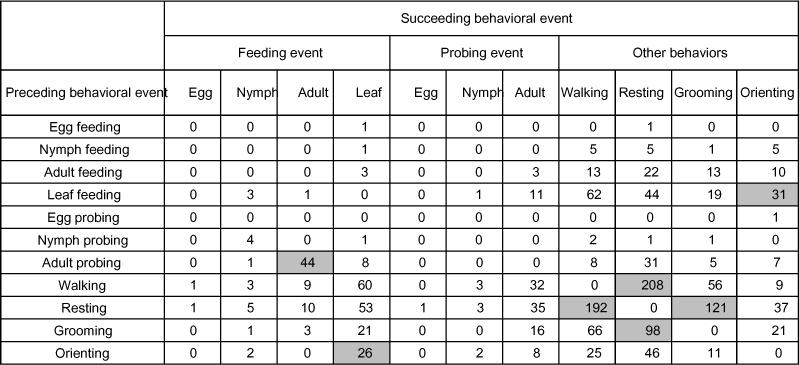
Adult O. tristicolor were most frequently observed eating the cotton leaf (3.7 ± 2.0 times per hour) (Figure 4A). On average, 0.1 ± 0.1 eggs, 0.6 ± 0.4 nymphs, and 2.1 ± 0.5 adults were consumed per hour of observation. The feeding duration on adult whiteflies was significantly greater than for the egg and nymphal stages. The average feeding durations were 45.0 ± 12.0, 44.4 ± 9.7, and 455.1 ± 65.3 seconds for eggs, nymphs, and adults, respectively (Figure 4B).
Figure 4.
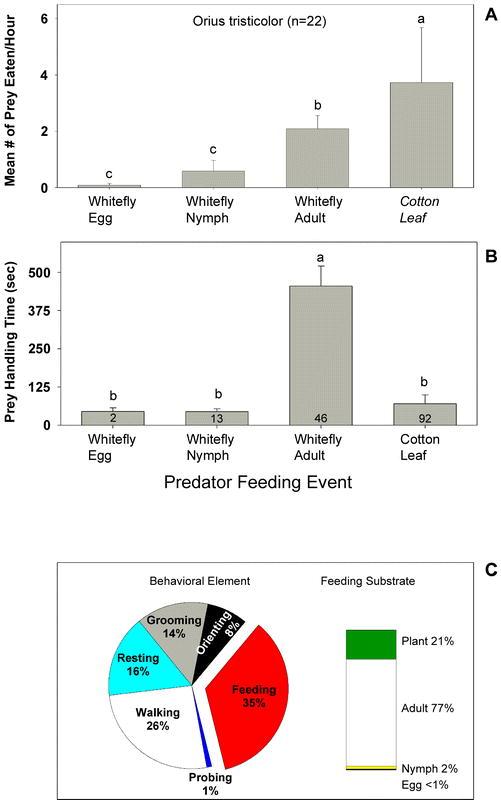
(A) Mean (± SD) frequency of Orius tristicolor feeding on the various whitefly lifestages or on the cotton leaf. The letters above the error bars indicate significant differences as determined by the Student Newman-Keuls multiple comparison test (P = 0.05, n = 22 individuals observed for 1 hour each). (B) The mean (± SD) amount of time O. tristicolor spent feeding on the various whitefly lifestages or on the cotton leaf. The numbers inside the vertical bars are the total number of observations recorded during 22 hours of observation. The letters above the error bars indicate significant differences as determined by Dunn's multiple comparison test (P = 0.05). (C) Time budgets for O. tristicolor in the feeding arena. Results are expressed as the percentage of total time spent in each behavioral element and on each feeding substrate over 22 hours of observation (n = 22 individuals).
Observations revealed that the proportion of time that adult O. tristicolor spent on each behavior was feeding (35%) > walking (26%) > resting (16%), grooming (14%) > orienting (8%) > probing (1%) (Figure 4C). Overall, O. tristicolor spent three-fourths of their feeding time eating adult whiteflies. Less than 3.0% of their feeding time was spent eating eggs and nymphs (Figure 4C).
A total of 1,489 behavioral events were recorded during 22 hours of observation (67.7 per hour). The general sequence of behavioral events exhibited by O. tristicolor as revealed by the analysis of the transition matrix showed that there is a high likelihood that O. tristicolor will proceed from:
grooming to walking
leaf feeding to orienting
orienting to leaf feeding
adult probing to walking and adult feeding
walking to grooming, resting and adult probing.
The most common sequence recorded was walking followed by grooming 16.8% of the time and grooming followed by walking 16.5% of the time (Table 5).
Table 5.
Frequencies of transitions between all observed behavioral events for adult Orius tristicolor on cotton leaves that also contained whitefly eggs, nymphs, and adults. Transitions that were significantly more common than expected by chance are shown in grey boxes (chi-square, adjusted P<0.001).
Lygus hesperus
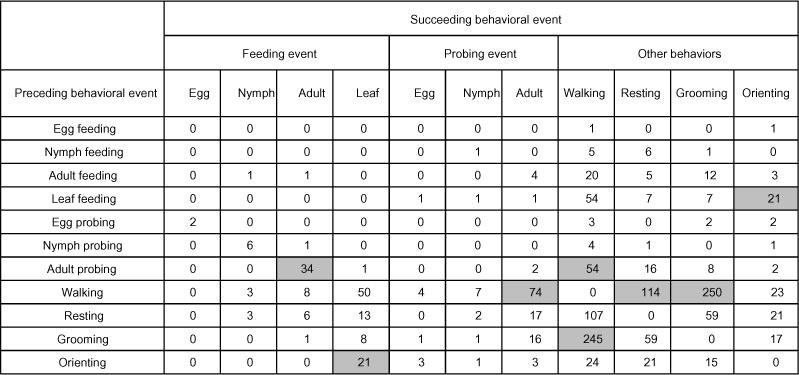
Adult L. hesperus were most commonly observed feeding (5.33 ± 0.71 times per hour) for extended periods of time (235.3 ± 123.2 seconds per feeding event) on the cotton leaf (Fig. 5A–B). Adult L. hesperus were next most commonly observed preying on whitefly nymphs. Very few whitefly eggs (4) or adults (3) were eaten during the 27 hours of observation. On average, 0.2 ± 0.1 eggs, 2.4 ± 1.0 nymphs, and 0.1 ± 0.1 adults were consumed per hour of observation (Figure 5A). The average prey feeding durations were 28.0 ± 17.2, 93.0 ± 11.4, and 221.7 ± 26.7 seconds for eggs, nymphs, and adults, respectively (Figure 5B).
Figure 5.

(A) Mean (± SD) frequency of Lygus hesperus feeding on the various whitefly lifestages or on the cotton leaf. The letters above the error bars indicate significant differences as determined by the Student Newman-Keuls multiple comparison test (P = 0.05, n = 27 individuals observed for 1 hour each). (B) The mean (± SD) amount of time L. hesperus spent feeding on the various whitefly lifestages or on the cotton leaf. The numbers inside the vertical bars are the total number of observations recorded during 27 hours of observation. The letters above the error bars indicate significant differences as determined by Dunn's multiple comparison test (P = 0.05). (C) Time budgets for L. hesperus in the feeding arena. Results are expressed as the percentage of total time spent in each behavioral element and on each feeding substrate over 27 hours of observation (n = 27 individuals).
Observations revealed that the proportion of time that L. hesperus spent on each behavior was feeding (42%) > grooming (24%) > walking (19%) > resting (11%) > orienting (2%), probing (2%) (Figure 5C). The majority (83%) of their feeding time was spent eating the plant (Figure 5C).
L. hesperus was not as active as the other predators examined in this study yielding a total of 670 behavioral events during 27 hours of observation (28.8 events per hour). The general sequence of behavioral events recorded for L. hesperus, as revealed by the analysis of the transition matrix, showed that there is a high likelihood that L. hesperus will proceed from:
nymph feeding to nymph probing and nymph probing to nymph feeding
egg probing to nymph feeding
grooming to walking and walking to grooming.
The most common sequence recorded was walking followed by grooming 11.8% of the time and grooming followed by walking 10.4% of the time (Table 6).
Table 6.
Frequencies of transitions between all observed behavioral events for adult Lygus hesperus on cotton leaves that also contained whitefly eggs, nymphs, and adults. Transitions that were significantly more common than expected by chance are shown in grey boxes (chi-square, adjusted P<0.001).
Discussion
Generally, insect predators either stalk or ambush their prey (Murdoch et al. 1985; O'Brien et al. 1989; Sabelis 1992; Hagler 2000). Certain assumptions about prey selection have been made for some whitefly predators based solely on their predominant hunting strategy (Breene 1992; Breene et al. 1992; Nordlund and Legaspi 1996). For example, ambush predators that place themselves in a strategic location and wait until prey move into their field of attack are considered to be best adapted for capturing mobile prey such as adult whiteflies (Sabelis 1992; Breene 1992). Conversely, stalking predators that constantly roam their environment searching for prey are considered to be best adapted for capturing immobile prey such as whitefly eggs and nymphs (Breene et al. 1992, 1994; Nordlund and Legaspi 1996). The predators examined in this study are generally regarded as stalking predators. As a consequence, an overwhelming majority of studies examining whitefly predation have been restricted to studies with the sessile whitefly lifestages conducted in no-choice arenas containing either eggs or nymphs or in choice arenas containing both eggs and nymphs (Elbadry 1968; Gerling 1986; Butler and Henneberry 1988; Kapadia and Puri 1991; Hoelmer et al. 1993, 1994; Legaspi et al. 1994; Nordlund and Legaspi 1996; Guershon and Gerling 1999). To our knowledge, only a few no-choice experiments have been conducted to identify the sources of adult whitefly predation. Of these, various species of true bugs, flies, and spiders have been identified as predators of adults (Sussman 1988; Cohen and Byrne 1992; Breene et al. 1992, 1994). Furthermore, to date only one predator evaluation has been conducted in which a predator was simultaneously presented with a choice of whitefly eggs, nymphs and adults. In that study, Hagler (2002) showed that the predatory fly, Drapetis nr. divergens fed exclusively on adult whiteflies. Clearly, as Nordlund and Legaspi (1996) pointed out, more studies are needed to evaluate predation on all whitefly stages.
The results from our study revealed that the beetles, H. convergens and C. vittatus, readily and rapidly preyed on all whitefly lifestages with a preference for eggs > adults > nymphs. Both species spent about 7.0% of their overall time budget feeding, but H. convergens spent much more time immobile/resting (18.4% of their time) and grooming (35.0%) behaviors whereas C. vittatus spent 52% of their time budget walking in the arena. An interesting foraging characteristic exhibited by the beetles was revealed in the analyses of their transitional behaviors. H. convergens and C. vittatus were frequently observed feeding consecutively on many individuals of the same whitefly lifestage. For example, an initial egg feeding was frequently followed by additional egg feedings or initial nymph feeding was frequently followed by additional nymph feedings. Whether this pattern of foraging behavior was due to associative learning is an area for future research.
Prey selection by the true bugs, G. punctipes, O. tristicolor, and L. hesperus yielded some surprising results. For example, G. punctipes and O. tristicolor preyed on adults over three times more frequently than on eggs and nymphs combined. In contrast, L. hesperus, a notorious pest in many cropping systems, was observed feeding on nymphs over nine times more frequently than on eggs and adults combined. Our observations of G. punctipes and those of Cohen and Byrne (1992) indicate that adult whiteflies rarely escaped when attacked by true bugs. Further observations revealed that the true bugs did not feed on nearly as many whiteflies as the beetles and spent a considerably longer time (e.g., 5 to 25 times longer) eating their prey. The increased handling time exhibited by the true bugs when compared to the beetles is due primarily to differences in their feeding mechanisms. The chewing mouthparts of beetles enable them simultaneously to kill by crushing or macerating and to rapidly devour their prey (see Video 1 available at http://insectscience.org/4.1/). Conversely, the true bugs have piercing and sucking mouthparts. The stylets are used to pierce the prey, to disrupt the cell walls and cellular contents, and to deliver saliva containing potent digestive enzymes (see Video 2 available at http://insectscience.org/4.1/). Together, the mechanical damage and the enzymes break down the tissue into a slurry of small particles, which are ingested along with the saliva (Cohen 1998). Overall, the time budgets exhibited for each of the true bugs were similar with all three species spending approximately one-third of their time in the immobile resting and grooming behaviors, two common characteristics of an ambush predator. Furthermore, approximately 40% of their time was spent feeding on either plant or insect tissue. However, there was a sharp contrast between the amounts of time that G. punctipes and L. hesperus spent feeding on plant tissue compared to O. tristicolor. Analysis of the feeding times revealed that L. hesperus, G. punctipes, and O. tristicolor spent 83, 66, and 21% of their feeding time on plant tissue, respectively. The majority of feeding by O. tristicolor's (77%) was spent eating adult whiteflies.
The prey choices observed in the feeding arena provide some insight to the results obtained from previous gut content examinations on these predators using a whitefly-specific ELISA (Hagler and Naranjo 1994a,b). In those studies, ∼35% of all field collected G. punctipes and O. tristicolor contained whitefly egg protein in their guts (Hagler and Naranjo 1994a). However, as discussed above, a positive ELISA reaction can be obtained for a predator that eats a whitefly egg or a gravid adult female (Hagler et al. 1993). Originally we believed that these predators were feeding primarily on whitefly eggs. The results from this study suggest that the majority of positive responses yielded by field-collected G. punctipes and O. tristicolor might be more likely due to predation on adult females. Prey selection by L. hesperus, H. convergens, and C. vittatus make it impossible to differentiate a positive ELISA response for the presence of whitefly between an egg and an adult female. For instance, while L. hesperus preyed almost exclusively on whitefly nymphs, their rare occurrence of predation on eggs and adults was about the same. Conversely, H. convergens and C. vittatus readily preyed on all whitefly lifestages. The voracious appetite exhibited by the beetles for every whitefly lifestage suggests that a positive whitefly ELISA response is probably due to the beetles feeding on a combination of eggs and adult females.
We would be remiss if we did not discuss some of the potential drawbacks associated with studies of predation conducted in confined feeding arenas. First, the whitefly population in each arena was very high. This could lead to an overestimation of predation because it minimizes the time needed for the predator to search for its prey. However, the whitefly densities in our arenas were not atypical of the densities often found late in the season in the cotton growing regions of the Southwestern United States (Ellsworth et al. 1993; Natwick et al. 1995). Second, the predators and whiteflies were fully enclosed in a Petri dish. The enclosed arena can reduce the ability of an adult whitefly to escape and the ability of the predator to move to a different plant or patch. This too would likely lead to an overestimation of predation rates. Moreover, no alternate prey species were made available to the predators in the enclosed arenas. The presence of alternate prey would likely reduce the number of whiteflies consumed by these generalist predators, especially if the alternate prey species were preferred or prey selection were random. Finally, in some instances, two predators were observed consecutively in the same feeding arena. While we did not observe any noticeable differences in the behaviors between the two individuals, it is possible that the behavior of the first predator could have indirectly affected the behavior of the second due to an induced response mediated by the plant, herbivore, and/or predator (Dicke et al. 1990, 2003; Dicke 1994, 2000; Dicke and van Loon 2000; Cortesero et al. 2000). The investigation of chemically induced responses between the trophic levels was beyond the scope of this investigation.
The major objective of this study was to identify prey selection by these five whitefly predators. Additionally, we identified the propensity of these predators to feed on cotton foliage. The frequency and duration of cotton feeding was highly variable between the beetles and the true bugs. The beetles were sometimes observed briefly nibbling or “skimming” the surface of the cotton leaf, probably to obtain water, sugar and other micronutrients. On the other hand, the true bugs were often observed inserting their piercing and sucking mouthparts deep into leaf veins and feeding for extended periods of time. It appears from this study and others that plant feeding is an important component to the nutritional ecology of many heteropterans (Naranjo and Gibson 1996; Wiedenmann et al. 1996; Coll 1996, 1998; Coll and Izraylevich 1997; Coll and Guershon 2002). For example, while it has been determined that Geocoris spp. can be reared in the laboratory exclusively on a meat diet (Cohen 1985), their fitness can be increased with the addition of plant nutrients to their diet (Dunbar and Bacon 1972; Tamaki and Weeks 1972; Naranjo and Gibson 1996). For instance, Tamaki and Weeks (1972) showed that Geocoris bullatus and G. pallens had increased longevity and fecundity and a faster rate of development when a diet of aphids was supplemented with plant material. Others have shown that facultative phytophagy by true bugs provides the insects with water, but also may provide some nutrients that supplement prey diet and help the predator survive periods when prey are scarce (Ridgway and Jones 1968; Stoner 1972; Salas-Aguilar and Ehler 1977; Kiman and Yeargan 1985; Wiedenmann and O'Neil 1991, 1992; Naranjo and Gibson 1996; Armer et al. 1998; Coll and Guershon 2002). Eubanks and Denno (2000) reported that the availability of high quality bean pods in the diet of Geocoris spp. resulted in a shift from carnivory to phytophagy.
Armer et al. (1998) state that a continuum exists among heteropteran feeding characteristics ranging from facultative phytophagous predators to facultative predaceous herbivores (Wiedenmann et al. 1996). While facultative herbivory is common among many predaceous true bugs (Polis et al. 1989; Rosenheim et al. 1995; Naranjo and Gibson 1996; Polis and Winemiller 1996), it is surprising that a greater research effort has not been made towards determining what advantages in fitness are derived by “predators” that feed on plants and “herbivores” that feed on insects. Insight into whether these true bugs are primarily predators or primarily herbivores might be determined by studies of their digestive enzyme complex. Specifically, researchers have determined that the presence of certain digestive enzymes indicates an insect's ability to efficiently eat plant tissue, animal tissue, or both (Baptist 1941; Adams and McAllen 1956; Strong and Kruitwagen 1968; Tingey and Pillemer 1977; Cohen 1995, 1998). For example, Agustí and Cohen (2000) conducted a study of the digestive enzyme complex of two omnivorous Lygus spp. They concluded that L. hesperus was better suited to carnivory than L. lineolaris because it has greater trypsin-like activity, and L. lineolaris was better suited to herbivory because it had greater α-amylase and pectinase activity.
In conclusion, our study provides insight into the host range and feeding characteristics of five common whitefly predators. The beetles preyed on all of the whitefly lifestages with a preference for eggs and adults, whereas G. punctipes and O. tristicolor preferred adults, and L. hesperus preferred nymphs. Based solely on the results from this laboratory study and from previous predator gut content evaluations (Hagler and Naranjo 1994a,b), we conclude that adult H. convergens and C. vittatus will be more effective whitefly biological control agents than the adult true bugs, G. punctipes and O. tristicolor in the cotton system of the Southwestern United States. Specifically, H. convergens and C. vittatus fed on many more whiteflies in less time than G. punctipes and O. tristicolor and their prey choice was directed more against the egg stage than the other species tested. Furthermore, we have shown that the proportion of beetles feeding on whiteflies in the field is over twice as high as that of the true bugs. While L. hesperus is a predator of whitefly (Hagler and Naranjo 1994a) and many other pest species (Knight 1941; Lindquist and Sorensen 1970; Cleveland 1987; Bryan et al. 1976; Wheeler 1976; Hagler and Naranjo 1994a,b), it is not a viable biological control agent because of its propensity to feed on plant meristematic tissue.
This study and other laboratory studies indicate that predatory beetles possess many characteristics that make them superior biological candidates, but we caution that such laboratory studies should not be considered a complete representation of what happens under field conditions. Predator behavior studies such as this will ultimately need to be combined with studies of nutrition (Naranjo and Stimac 1985; Coll 1996; Coll and Izraylevich 1997), physiology (Cohen and Byrne 1992; Agusti and Cohen 2000), and predator population dynamics (Hagler and Naranjo 1994a,b) to determine the predatory potential of each of these insects in a whitefly biological control program.
Acknowledgments
We thank Sunny Carrington, Melani Charney, Rochelle Christensen, Shana England, and Dan Langhorst for technical assistance. We are most grateful to Jackie Blackmer, Hollis Flint, Dan Gerling, Debbie Hagler, Kim Hoelmer, and three anonymous reviewers for reviewing earlier versions of this manuscript.
Disclaimer
This article reports the results of research only. Mention of a proprietary product does not constitute an endorsement or a recommendation for its use by the USDA.
References
- Adams JB, McAllan JW. Pectinase in the saliva of Myzus persicae (Homoptera: Aphididae) Canadian Journal of Zoology. 1956;34:541–542. [Google Scholar]
- Agusti N, Cohen AC. Lygus hesperus and L. lineolaris (Hemiptera: Miridae), phytophages, zoophages, or omnivores:Evidence of feeding adaptations suggested by the salivary and midgut digestive enzymes. Journal of Entomological Science. 2000;35:176–186. [Google Scholar]
- Armer CA, Wiedenmann RN, Bush DR. Plant feeding site selection on soybean by the facultatively phytophagous predator Orius insidiosus. Entomologia Experimentalis et Applicata. 1998;86:109–118. [Google Scholar]
- Baptist BA. The morphology and physiology of the salivary glands of Hemiptera-Heteroptera. Quarterly Journal of Microscopic Science. 1941;83:91–139. [Google Scholar]
- Breene RG. A new approach for matching biological control agents to pests. The IPM Practitioner. 1992;14:1–9. [Google Scholar]
- Breene RG, Dean DA, Quarles W. Predators of sweetpotato whitefly. The IPM Practitioner. 1994;16:1–9. [Google Scholar]
- Breene RG, Meagher RL, Nordlund DA, Wang YT. Biological control of Bemisia tabaci (Homoptera: Aleyrodidae) in a greenhouse using Chrysoperla rufilabris (Neuroptera: Chrysopidae) Biological Control. 1992;2:9–14. [Google Scholar]
- Bryan DE, Jackson CG, Carranza RL, Neemann EG. Lygus hesperus: Production and development in the laboratory. Journal of Economic Entomology. 1976;69:127–129. [Google Scholar]
- Butler GD, Henneberry TJ. Laboratory studies of Chrysoperla carnea predation on Bemisia tabaci. The Southwestern Entomologist. 1988;13:165–170. [Google Scholar]
- Cleveland TC. Predation by tarnished plant bugs (Heteroptera: Miridae) of Heliothis (Lepidoptera: Noctuidae) eggs and larvae. Environmental Entomology. 1987;16:37–40. [Google Scholar]
- Cock MJW. Integrated management of whitefly pest problems in the Middle and Near East with special emphasis on biological control. Arab Journal of Plant Protection. 1994;12:127–136. [Google Scholar]
- Cohen AC. Simple method for rearing the insect predator Geocoris punctipes (Heteroptera: Lygaeidae) on a meat diet. Journal of Economic Entomology. 1985;78:1173–1175. [Google Scholar]
- Cohen AC. Extra-oral digestion in predatory Arthropoda. Annual Review of Entomology. 1995;40:85–103. [Google Scholar]
- Cohen AC. Solid-to-liquid feeding: The inside(s) story of extra-oral digestion in predaceous Arthropoda. American Entomologist. 1998;44:103–117. [Google Scholar]
- Cohen AC, Byrne DN. Geocoris punctipes as a predator of Bemisia tabaci: A laboratory evaluation. Entomologia Experimentalis et Applicata. 1992;64:195–202. [Google Scholar]
- Coll M. Feeding and ovipositing on plants by an omnivorous insect predator. Oecologia. 1996;105:214–220. doi: 10.1007/BF00328549. [DOI] [PubMed] [Google Scholar]
- Coll M. 1997 Feeding and living on plants in predatory Heteroptera. In: Coll M, Ruberson JR, editors. Predatory Heteroptera: Their Ecology and Use in Biological Control, Lanham, MD: Thomas Say Publications in Entomology. [Google Scholar]
- Coll M, Guershon M. Omnivory in terrestrial arthopods: Mixing plant and prey diets. Annual Review of Entomology. 2002;47:267–297. doi: 10.1146/annurev.ento.47.091201.145209. [DOI] [PubMed] [Google Scholar]
- Coll M, Izraylevich S. When predators also feed on plants: Effects of competition and plant quality on omnivore-prey population dynamics. Annals of the Entomological Society of America. 1997;90:155–161. [Google Scholar]
- Cortesero AM, Stapel JO, Lewis WJ. Understanding and manipulating plant attributes to enhance biological control. Biological Control. 2000;17:35–49. [Google Scholar]
- Dicke M. Local and systematic production of volatile herbivore-induced terpenoids: Their role in plant-carnivore mutualism. Journal of Plant Physiology. 1994;143:465–472. [Google Scholar]
- Coll M. 1998 Living and feeding on plants in predatory Heteroptera. In: Coll M, Ruberson JR, editors. Predatory Heteroptera: Their ecology and use in biological control. 89–129.Lanham, MD: Thomas Say Publications in Entomology. [Google Scholar]
- Dicke M. Chemical ecology of host-plant selection by herbivorous arthropods: A multitrophic perspective. Biochemical Systematics and Ecology. 2000;28:601–617. doi: 10.1016/s0305-1978(99)00106-4. [DOI] [PubMed] [Google Scholar]
- Dicke M, Sabelis MW, Takabayashi J, Bruin J, Posthumus MA. Plant strategies of manipulating predator-prey interactions through allelochemicals: Prospects for application in pest control. Journal of Chemical Ecology. 1990;16:3091–3118. doi: 10.1007/BF00979614. [DOI] [PubMed] [Google Scholar]
- Dicke M, van Poecke MP, de Boer JG. Inducible indirect defence of plants: From mechanisms to ecological functions. Basic and Applied Ecology. 2003;4:27–42. [Google Scholar]
- Dicke M, van Loon JJA. Multiple effects of herbivore-induced plant volatiles in an evolutionary context. Entomologia Experimentalis et Applicata. 2000;97:237–249. [Google Scholar]
- Dunbar DM, Bacon OG. Feeding, development, and reproduction of Geocoris punctipes (Heteroptera: Lygaeidae) on eight diets. Annals of the Entomological Society of America. 1972;65:892–895. [Google Scholar]
- Elbadry EA. Biological studies on Amblyseius aleyrodis a predator of the cotton whitefly (Acarina, Phytoseiidae) Entomophaga. 1968;13:323–329. [Google Scholar]
- Ellsworth PC, Meade DL, Husman SH, Ramsey CS, Silvertooth JC, and Malcuit JE. 1993 Sweetpotato whitefly preference and performance on medium maturity cotton varieties in Arizona. In: Silvertooth J, editor. Cotton: A college of agriculture report, Series P-94. pp. 275–279. [Google Scholar]
- Eubanks MD, Denno RF. Host plants mediate omnivore-herbivore interactions and influence prey suppression. Ecology. 2000;81:936–947. [Google Scholar]
- Gerling D. Natural enemies of Bemisia tabaci, biological characteristics and potential as biological control agents: A review. Agriculture, Ecosystems and Environment. 1986;17:99–110. [Google Scholar]
- Gerling D. 1990 Natural enemies of whiteflies: Predators and parasitoids. In: Gerling D, editor. Whiteflies: Their Bionomics, Pest Status and Management. 147–185.Andover, Hants, UK: Intercept Ltd. [Google Scholar]
- Gerling D. Approaches to the biological control of whiteflies. The Florida Entomologist. 1992;75:446–456. [Google Scholar]
- Gerling D, Alomar O, Arno J. Biological control of Bemisia tabaci using predators and parasitoids. Crop Protection. 2001;20:779–799. [Google Scholar]
- Gerling D, Mayer RT. Bemisia: 1995 Taxonomy, Biology, Damage, Control and Management Interecpt 1996 [Google Scholar]
- Guershon M, Gerling D. Predatory behavior of Delphastus pusillus in relation to the phenotypic plasticity of Bemisia tabaci nymphs. Entomologia Experimentalis et Applicata. 1999;92:239–248. [Google Scholar]
- Hagler JR. 2000 Biological Control of Insects. In: Rechcigl JR, Rechcigl NA, editors. Insect Pest Management: Techniques for Environmental Protection. 207–241.New York, NY: Lewis Publishers. [Google Scholar]
- Hagler JR. Foraging behavior, host stage selection and gut content analysis of field collected Drapetis nr. divergens: A predatory fly of Bemisia argentifolli. Southwestern Entomologist. 2002;27:241–249. [Google Scholar]
- Hagler JR, Brower AG, Tu Z, Byrne DN, Bradley-Dunlop D, Enriquez FJ. Development of a monoclonal antibody to detect predation of the sweetpotato whitefly, Bemisia tabaci. Entomologia Experimentalis et Applicata. 1993;68:231–236. [Google Scholar]
- Hagler JR, Naranjo SE. Determining the frequency of heteropteran predation on sweetpotato whitefly and pink bollworm using multiple ELISAs. Entomologia Experimentalis et Applicata. 1994a;72:59–66. [Google Scholar]
- Hagler JR, Naranjo SE. Qualitative survey of two coleopteran predators of Bemisia tabaci (Homoptera: Aleyrodidae) and Pectinophora gossypiella (Lepidoptera: Gelechiidae) using a multiple prey gut content ELISA. Environmental Entomology. 1994b;23:193–197. [Google Scholar]
- Hoelmer KA, Osborne LS, Yokomi RK. Reproduction and feeding behavior of Delphastus pusillus (Coleoptera: Coccinellidae), a predator of Bemisia tabaci (Homoptera: Aleyrodidae) Journal of Economic Entomology. 1993;86:322–329. [Google Scholar]
- Hoelmer KA, Osborne LS, Yokomi RK. Interactions of the whitefly predator Delphastus pusillus (Coleoptera: Coccinellidae) with parasitized sweetpotato whitefly (Homoptera: Aleyrodidae) Environmental Entomology. 1994;23:136–139. [Google Scholar]
- Isaacs R, Cahill M, Byrne DN. Host plant evaluation behaviour of Bemisia tabaci and its modification by external or internal uptake of imidacloprid. Physiological Entomology. 1999;24:1–8. [Google Scholar]
- Kapadia MN, Puri SN. Seasonal incidence of natural enemies of Bemisia tabaci (Gennadius) on cotton. Indian Journal of Ecology. 1989;16:164–168. [Google Scholar]
- Kapadia MN, Puri SN. Biology and comparative predation efficacy of three heteropteran species recorded as predators of Bemisia tabaci in Maharashtra. Entomophaga. 1991;36:555–559. [Google Scholar]
- Kiman ZB, Yeargan KV. Development and reproduction of the predator Orius insidiosus (Hemiptera: Anthocoridae) reared on diets of selected plant material and arthropod prey. Annals of the Entomological Society of America. 1985;78:464–467. [Google Scholar]
- Knight HH. 1941 The plant bugs or Miridae of Illinois. Bulletin of the Illinois State Natural History Survey. No. 22. [Google Scholar]
- Legaspi JC, Carruthers RI, Nordlund DA. Life history of Chrysoperla rufilabris (Neuroptera: Chrysopidae) provided sweetpotato whitefly Bemisia tabaci (Homoptera: Aleyrodidae) and other food. Biological Control. 1994;4:178–184. [Google Scholar]
- Lehner PN. 1979 Handbook of Ethological Methods. STPM, New York: Garland. [Google Scholar]
- Lindquist RK, Sorensen EL. Interrelationships among aphids, tarnished plant bugs, and alfalfas. Journal of Economic Entomology. 1970;63:192–195. [Google Scholar]
- Murdoch WW, Chesson J, Chesson PL. Biological control in theory and practice. American Naturalist. 1985;125:344–366. [Google Scholar]
- Naranjo SE, Ellsworth PC. Special issue: Challenges and opportunities for pest management of Bemisia tabaci in the new century. Crop Protection. 2001;20:707–869. [Google Scholar]
- Naranjo SE, Gibson RL. 1996 Phytophagy in Predaceous Heteroptera: Effects on life history and population dynamics. In: Alomar O, Wiedenmann RN, editors. Zoophytophagous Heteroptera: Implications for Life History and Integrated Pest Management. 57–93.Lanham, MD: Thomas Say Publications in Entomology. [Google Scholar]
- Naranjo SE, Stimac JL. Development, survival, and reproduction of Geocoris punctipes (Hemiptera: Lygaeidae): Effects of plant feeding on soybean and associated weeds. Environmental Entomology. 1985;14:523–530. [Google Scholar]
- Natwick ET, Chu CC, Perkins HH, Henneberry TJ, Cohen AC. Pima and Upland cotton susceptibility to Bemisia argentifolii under desert conditions. Southwestern Entomologist. 1995;20:429–438. [Google Scholar]
- Nordlund DA, Legaspi JC. 1996 Whitefly predators and their potential for use in biological control. In: Gerling D, Mayer RT, editors. Bemisia 1995: Taxonomy, Biology, Damage, Control and Management. 499–513.Andover, Hants, UK: Intercept Ltd. [Google Scholar]
- O'Brien WJ, Evans BI, Browman HI. Flexible search tactics and efficient foraging in saltatory animals. Oecologia. 1989;80:100–110. doi: 10.1007/BF00789938. [DOI] [PubMed] [Google Scholar]
- Or R, Gerling D. The green lacewing, Chrysoperla carnea as a predator of Bemisia tabaci. Phytoparasitica. 1985;13:75. [Google Scholar]
- Perring TM. 1996 Biological differences of two species of Bemisia that contribute to adaptive advantage. Bemisia 1995: Taxonomy, Biology, Damage, Control and Management. 3–16.Andover, Hants, UK: Intercept Ltd. [Google Scholar]
- Polis GA, Myers CA, Holt RD. The ecology and evolution of intraguild predation-potential competitors that eat each other. Annual Review of Ecology and Systematics. 1989;20:297–330. [Google Scholar]
- Polis GA, Winemiller KO. 1996 Food Webs: Integration of Patterns and Dynamics. New York, NY: Chapman and Hall. [Google Scholar]
- Ridgway RL, Jones SL. Plant feeding by Geocoris pallens and Nabis americoferus. Annals of the Entomological Society of America. 1968;61:232–233. [Google Scholar]
- Rosenheim JA, Kaya HK, Ehler LE, Marois JJ, Jaffee BA. Intraguild predation among biological-control agents theory and evidence. Biological Control. 1995;5:303–335. [Google Scholar]
- Sabelis MW. 1992 Predatory arthropods. In: Crawley MJ, editor. Natural Enemies, the Population Biology of Predators, Parasites and Diseases. 225–264.Cambridge, MA: Blackwell Scientific Publications. [Google Scholar]
- Salas-Aguilar J, Ehler LE. Feeding habits of Orius tristicolor. Annals of the Entomological Society of America. 1977;70:60–62. [Google Scholar]
- Slater PJB. 1973 Describing sequences of behavior. In: Batesont PPG, Klopfer PH, editors. Perspectives in Ethology. 131–153.Plenum, New York: Plenum Press. [Google Scholar]
- Stoner A. Plant feeding by Nabis, a predaceous genus. Environmental Entomology. 1972;1:557–558. [Google Scholar]
- Strong FE, Kruitwagen EC. Polygalactronase in the salivary apparatus of Lygus hesperus (Hemiptera) Journal of Insect Physiology. 1968;14:1113–1119. [Google Scholar]
- Sussman L. 1988 The cotton insects of Israel and aspects of the biology of Deraeocoris pallens Reuter (Heteroptera, Miridae). MS thesis. Tel Aviv Univ. 154. pp. [Google Scholar]
- Tamaki G, Weeks RE. Biology and ecology of two predators, Geocoris pallens Stål and G. bullatus (Say) USDA Technical Bulletin. 1972;1446:1–46. [Google Scholar]
- Tingey WM, Pillemer EA. Lygus bugs: Crop resistance and physiological nature of feeding injury. Bulletin of the Entomological Society of America. 1977;23:277–287. [Google Scholar]
- Wheeler AG Jr. 1976 Lygus bugs as facultative predators. In: Scott DR, O'Keeffe LE, editors. In: Lygus bug: Host plant interactions. Proceedings of a workshop at the XV International Congress of Entomology. 19–26.August. 28–35, Moscow, ID: University Press of Idaho. [Google Scholar]
- Wiedenmann RN, Legaspi JC, and O'Neil RJ. 1996 Impact of prey density and facultative plant feeding on the life history of the predator Podisus maculiventris. In: Alomar O, Wiedenmann RN, editors. Zoophytophagous Heteroptera: Implications for Life History and Integrated Pest Management. 94–118.Lanham, MD: Thomas Say Publications in Entomology. [Google Scholar]
- Wiedenmann RN, O'Niel RJ. Searching behavior and time budgets of the predator Podisus maculiventris. Entomologia Experimentalis et Applicata. 1991;60:83–93. [Google Scholar]
- Wiedenmann RN, O'Niel RJ. Searching strategy of the predator Podisus maculiventris (Say) (Heteroptera: Pentatomidae) Environmental Entomology. 1992;21:1–9. [Google Scholar]
- Wiedenmann RN, Wilson LT. 1996 Potential for using zoophytophagous Heteroptera in IPM strategies. In: Alomar O, Wiedenmann RN, editors. Zoophytophagous Heteroptera: Implications for Life History and Integrated Pest Management. 190–202.Lanham, MD: Thomas Say Publications in Entomology. [Google Scholar]