

Abstract
Background
In the Metazoan nucleus, core histones assemble the genomic DNA to form nucleosome arrays, which are further compacted into dense chromatin structures by the linker histone H1. The extraordinary density of chromatin creates an obstacle for accessing the genetic information. Regulation of chromatin dynamics is therefore critical to cellular homeostasis, and histone chaperones serve as prominent players in these processes. In the current study, we examined the role of specific histone chaperones in negotiating the inherently repressive chromatin structure during transcriptional activation.
Results
Using a model promoter, we demonstrate that the human nucleosome assembly protein family members hNap1 and SET/Taf1β stimulate transcription in vitro during pre-initiation complex formation, prior to elongation. This stimulatory effect is dependent upon the presence of activators, p300, and Acetyl-CoA. We show that transcription from our chromatin template is strongly repressed by H1, and that both histone chaperones enhance RNA synthesis by overcoming H1-induced repression. Importantly, both hNap1 and SET/Taf1β directly bind H1, and function to enhance transcription by evicting the linker histone from chromatin reconstituted with H1. In vivo studies demonstrate that SET/Taf1β, but not hNap1, strongly stimulates activated transcription from the chromosomally-integrated model promoter, consistent with the observation that SET/Taf1β is nuclear, whereas hNap1 is primarily cytoplasmic. Together, these observations indicate that SET/Taf1β may serve as a critical regulator of H1 dynamics and gene activation in vivo.
Conclusions
These studies uncover a novel function for SET that mechanistically couples transcriptional derepression with H1 dynamics. Furthermore, they underscore the significance of chaperone-dependent H1 displacement as an essential early step in the transition of a promoter from a dense chromatin state into one that is permissive to transcription factor binding and robust activation.
Keywords: Linker histone, H1, Histone chaperone, SET/Taf1β, Chromatin, Transcription activation
Background
In the Metazoan nuclei, the extraordinary compaction of the genetic material is achieved through organization of the chromosomal DNA into extensively folded chromatin structures. Nucleosomes, the most basic unit of chromatin, are formed by wrapping 147 bp of DNA around two copies each of the core histones H2A, H2B, H3, and H4 [1]. The binding of the linker histone H1 further compacts the DNA into more condensed chromatin arrays [2]. The extensive compaction imparted by nucleosomes and the linker histones, in concert with other chromatin-associated proteins, creates a dense physical barrier that restricts access to the genetic information. As such, compacted chromatin structures are inherently incompatible with multiple nuclear processes, including gene expression. To enable access to promoters and regulatory elements of genes targeted for transcriptional activation, cells must possess mechanisms that actively locate these regions to locally relax the repressive chromatin fiber and expose specific DNA sequences.
The linker histone H1 plays a pivotal role in the formation and stabilization of extensively folded chromatin structures. In mammals there are at least 11 variants of H1. While structurally similar, with each subtype sharing a tripartite domain organization, the variants differ in their patterns of expression, chromosomal distribution and chromatin binding dynamics [3, 4]. The highly conserved central globular domain of the linker histones binds the nucleosome core particle at the entry/exit point of the DNA, further stabilizing the nucleosome and facilitating folding and compaction nucleosomal arrays into higher-ordered chromatin fibers [2, 4, 5]. Consistent with the repressive nature of H1, the association of the linker histones with chromatin inversely correlates with transcriptionally active gene regions. Recent genome-wide profiling studies revealed that the promoters of most transcriptionally active, or transcriptionally poised, genes are depleted of both nucleosomes and the linker histones [6–9]. However, unlike nucleosome-depleted regions that are generally localized to core promoters and enhancers, H1 displacement is considerably more extensive, extending significantly upstream and downstream of the transcription start site (TSS) [7–9]. These data further support the view that H1 binding is incompatible with gene expression, and that a critical early step in transcriptional activation is linker histone removal, followed by localized chromatin decompaction to accommodate the binding of the transcription machinery. Moreover, these observations indicate that specific cellular proteins and/or pathways must exist to facilitate H1 displacement from target gene regions primed for activation.
While many nuclear proteins modulate chromatin dynamics, the ATP-independent histone chaperones have established roles in numerous chromatin-related cellular processes, including gene expression [10, 11]. Members of the of nucleosome assembly protein (NAP) superfamily are amongst the best-characterized histone chaperones. The NAP proteins expressed throughout eukaryotes are structurally conserved and obligate dimers (or multimers), and their role in the regulation of chromatin dynamics is well documented [12]. Interestingly, despite the observation that their tertiary structure is conserved, differences exist in individual family members that likely support similar, yet distinct, chromatin-related functional properties [10, 11]. Members of the NAP superfamily bind the core histones with high affinity, and have been shown to promote nucleosome assembly [12], H2A/H2B dimer eviction and exchange [13–16], and acetylation-dependent nucleosome disassembly [17, 18]. Together, these activities support a prominent role for NAP-family proteins in regulated gene expression. In humans, there are at least eight NAP-superfamily members. These proteins differ with respect to their patterns of expression and subcellular distribution [19–21]. A subset of the NAP-family, NAP1L1-5, shares a greater degree of sequence homology, and thus the ability to heterodimerize given shared tissue expression and subcellular localization [19]. Of interest, all NAP1L1-5 proteins have been found in neurons, and a subset of these, hNAP1L2, L3, L5, exhibit restricted neuronal expression and dynamic changes in subcellular localization during differentiation [19]. The NAP-family members include hNAP1 (hNAP1L1) and hNAP2 (hNAP1L4) are ubiquitously expressed, as is the more distantly related NAP-family member, SET/Taf1 [also called SET, template activating factor-1 (Taf-1), INHAT, and IPP2A].
SET is predicted to share an overall structural homology with the NAP-superfamily member in mammals, despite the fact that it shares only 27 % amino acid sequence identity with hNAP1L1 [19]. The SET gene expresses two isoforms, SET/Taf1α and SET/Taf1β. Like the NAP proteins, SET is an obligate dimer, but does not heterodimerize with other NAP-family members, a finding consistent with observed pronounced differences in the dimerization helix [19, 22]. SET was originally discovered as a potent inhibitor of protein phosphatase 2A (PP2A) [23, 24]. Later, recurrent translocations involving SET and CAN (also called Nup214), a nucleoporin, were found to be associated with several distinct malignancies, including myeloid leukemias, colorectal, oral, and ovarian cancers [25–27]. The SET-CAN fusion protein dysregulates several potentially oncogenic nuclear processes, including aberrant retention of proteins targeted for export, stimulation of the wnt signaling pathway, and transcriptional activation of HOXA target genes [28–30]. Furthermore, misregulation of SET disrupts the catalytic activity of PP2A, which is linked to aberrant patterns of gene expression via effects on histone post-translational modifications [31], further linking SET to transcriptional regulation. Moreover, SET has recently been shown to play a role in chromosome segregation, suggesting that the SET/PP2A pathway is critical in the cell division cycle [32, 33]. Together, these diverse functions attributed to SET may be etiologically linked to malignant transformation associated with the deregulation of this important protein.
Although pleiotropic, SET has been shown to function in the regulated expression of numerous cellular genes. Its precise role in transcription, however, remains controversial. For example, several studies demonstrate that SET functions as a transcriptional activator via mechanisms that oppose the repressive effect of chromatin [31, 34–37], while other studies demonstrate SET functions as a repressor, primarily through inhibition of CBP/p300 acetylation activity [38–42]. Notably, the modulation of chromatin structure is a reoccurring theme in many studies that link SET and transcriptional regulation. Although the NAP-family proteins are well-characterized core histone chaperones, recent findings reveal that both NAP1 (hNAP1) and SET also interact with linker histone H1, and have been linked to chromatin decondensation through modulation of linker histone H1 dynamics [43–45]. However, the mechanisms by which these two human histone chaperones function in linker histone dynamics, and the potential outcome of the histone chaperone-H1 interaction on regulated gene expression, remain elusive.
In this report, we investigated the function of hNAP1 (hNAP1L1) and SET in a cell-free transcription system using a natural model promoter assembled into chromatin. We find that both chaperones enhance activated transcription, dependent upon the presence of activators, p300 and acetyl-CoA (AC-CoA). The stimulatory activity of hNAP1 and SET is significantly enhanced on chromatin templates assembled with H1. Importantly, both chaperones evict the linker histone from chromatin templates, providing a mechanistic foundation for their shared anti-repressive effects. Transfection of the histone chaperones into cells containing the integrated model promoter linked to a reporter plasmid reveals a potent transcriptional stimulatory function for SET, but not hNAP1. These data are consistent with the observation that SET is localized primarily in the nucleus. Further, transcriptional enhancement by SET is only observed with the integrated promoter, and under conditions of activated transcription. Together, these data uncover a novel transcription stimulatory function for SET that occurs early in transcriptional initiation: opposing H1 repression mediated through the targeted eviction of the linker histone from chromatin. Displacement of H1 is required for chromatin unfolding and the subsequent binding of the transcription machinery, creating a chromatin environment compatible with transcriptional activation.
Results
NAP1 proteins enhance activated transcription on chromatin templates during PIC assembly
To investigate the role of NAP-family histone chaperones in transcription, we utilized a chromatin-based in vitro transcription system that we previously demonstrated to be highly responsive to activators, coactivators, and acetyl-CoA (Ac-CoA) [18, 46–48]. This model system is composed of a 588 bp (or 900 bp, see below) fragment carrying the natural HTLV-1 promoter linked to a G-less cassette immediately downstream of the TSS (Fig. 1a). The promoter fragment carries an upstream biotin linkage to enable immobilization on a streptavidin-coated magnetic bead. The HTLV-1 transcriptional control region carries three reiterated 21 bp enhancer elements, called viral cyclic AMP response elements (vCREs), located upstream of the TSS (Fig. 1a). The vCREs serve as binding sites for the cellular transcription factor CREB and the potent HTLV-1-encoded transcription factor Tax. DNA-bound Tax and ser-133 phosphorylated CREB (pCREB) together strongly recruit the coactivators CBP/p300 [48, 49] (Fig. 1a, b). CBP and p300 are ubiquitously expressed, highly homologous histone acetyltransferases (HATs) involved in the regulation of many classes of genes throughout metazoans [50]. Tax and pCREB recruitment of the coactivators is essential for potent, Ac-CoA-dependent transcription on chromatin templates in vitro [18, 47, 48].
Fig. 1.
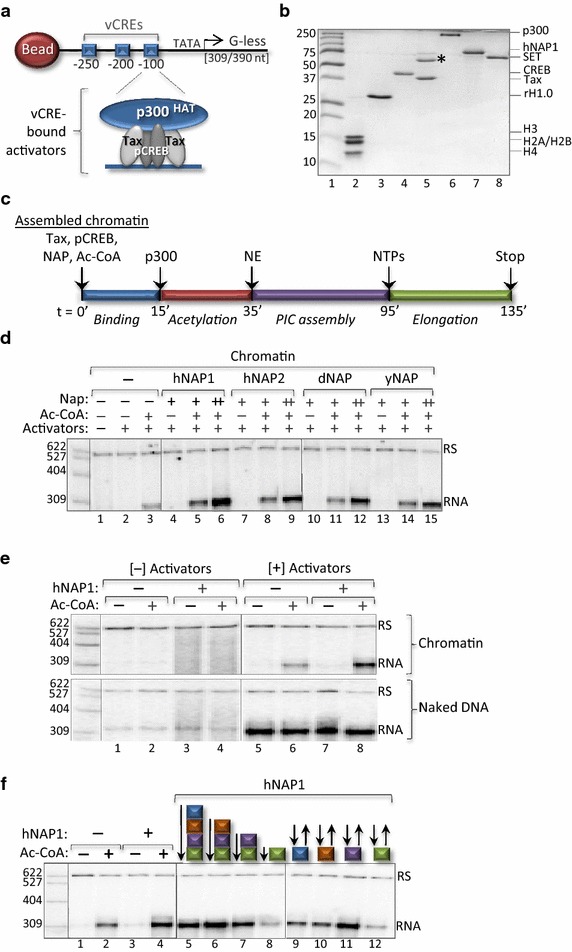
NAP proteins enhance activated transcription from an immobilized chromatin template. a Schematic depicting the model transcription template: the HTLV-1 promoter/G-less cassette. b Purified recombinant proteins. The indicated proteins used in the experiments described herein were separated by SDS-PAGE and visualized by Coomassie staining. Molecular weight size markers (kDa) (lane 1), X. laevis histone octamer (10 pmol) (lane 2), linker histone H1.0 (20 pmol) (lane 3), pCREB (Ser-133, 20 pmol) (lane 4), Tax (20 pmol) (lane 5); asterisk denotes GroEL that co-purifies with Tax, p300 (10 pmol) (lane 6), GST-hNap1 (20 pmol) (lane 7), and GST-SET/Taf1β (16 pmol) (lane 8). c Schematic representation of the in vitro transcription assay showing the individual, temporally-distinct steps. d NAP-family proteins enhance activated transcription from the chromatin-assembled promoter template. The repressive effect of chromatin was relieved by the addition of purified, recombinant activators (Tax, pCREB, and p300) and Ac-CoA. The indicated purified, recombinant histone chaperone (0.17 [+] or 0.67 μM [++]) was added in the presence of activators (Tax, pCREB and p300), and in the absence or presence of Ac-CoA (lanes 4–15). The amount of hNAP1 that gave the greatest enhancement (0.67 μM) was used in subsequent experiments. The transcript (RNA), recovery standard (RS), and size markers are shown. Solid vertical lines designate where the image (from the same gel) was spliced to remove irrelevant lanes, or portions of the image arranged to simplify presentation of the data. The experiment was performed at least three times, and a representative experiment is shown. For this, and subsequent in vitro transcription reactions, size markers derived from radiolabeled HpaII-digested pBR322 are shown on the left (bp). Prior to RNA isolation, a radiolabeled 622 bp recovery standard (RS) was added to each reaction to monitor transcript recovery. e hNAP1 stimulation of transcription requires activators, Ac-CoA, and chromatin. In vitro transcription assays were performed on chromatin-assembled (upper panel) or naked DNA (lower panel) templates, in the absence or presence of activators, as indicated. f hNAP1 enhances transcription during the PIC assembly step. In vitro transcription reactions, performed in the absence or presence of Ac-CoA and/or hNAP1 (added at the start of the reaction), is shown for reference (lanes 1–4). The effect of hNAP1 addition at the start of each individual step, denoted by color (see panel b) is shown (lanes 5–8). The effect of hNAP1, added at the start of the indicated step and removed from the reaction at the end of the indicated step (see “Results” for details), is shown (lanes 9–12). All reactions were performed in the presence of activators (Tax, pCREB, p300)
Using our model cell-free transcription system we investigated whether NAP-family histone chaperones from different species functioned to influence activated transcription. Unless indicated otherwise, all transcription reactions were performed as follows: the immobilized chromatin-assembled model promoter templates [50 ng (0.004 µM)] were incubated in the presence of purified, recombinant activators [pCREB (0.03 µM), Tax (0.06 µM), p300 (0.02 µM)], the indicated histone chaperone (0.67 µM), and Ac-CoA (100 µM) (Fig. 1b). We tested purified, recombinant NAP from yeast, Drosophila, and the two ubiquitously expressed human NAP-family proteins, hNAP1 and hNAP2. A schematic depicting the temporal steps in the in vitro transcription reaction is shown in Fig. 1c. We first assembled chromatin on our immobilized model promoter by the method of salt dilution, and measured the effect of the NAP proteins on transcription (Fig. 1d). As expected, the chromatin-assembled promoter template was fully repressed in the absence of activators and Ac-CoA (lanes 1, 2). The addition of activators and Ac-CoA, however, produced an RNA transcript of the appropriate length (lane 3). Importantly, the addition of two amounts of each of the NAP proteins further increased the level of transcription in a dose- and Ac-CoA-dependent manner (lanes 4–15). We observed the most dramatic transcriptional enhancement in the presence of human NAP1. For this reason, and because the other components in our transcription system are of human origin, we selected hNAP1 for use in the subsequent studies presented herein. We next examined the enhancement effect of hNAP1 under a variety of transcription conditions (Fig. 1e). Consistent with a role for NAP-family proteins in modulating chromatin dynamics, we found that the transcription stimulatory effect of hNAP1 was only observed on chromatin templates, as hNAP1 addition had no effect on transcription reactions performed on naked DNA. Furthermore, transcriptional enhancement by hNAP1 required the presence of Tax, pCREB, p300, and Ac-CoA (Fig. 1e, and data not shown).
In the preceding experiments, NAP1 was added at the start of each reaction (t = 0′), and was present through termination (stop). To assess whether hNAP1 functioned optimally at a specific, temporally-distinct step in the transcription assay (see Fig. 1c), we tested the effect of hNAP1 when added at the start of each phase of the reaction. Figure 1f shows that hNAP1 enhanced transcription, except when added at the start of the elongation phase (lanes 5–8). This observation suggests that hNAP1 functions during assembly of the pre-initiation complex (PIC). To investigate this possibility further, we took advantage of our immobilized template system to specifically examine the effect of hNAP1 during each individual step in the reaction. hNAP1 was added at the start of each step, however, the supernatant containing hNAP1 was removed following magnetic isolation the transcription templates at the end of each step, and replaced with one prepared in parallel in the absence of hNAP1. Using this approach, the equilibrium binding conditions of the other reaction components remained essentially constant. Figure 1f shows that hNAP1 significantly enhanced transcription when present during PIC assembly (lane 11). It is unclear why NAP exhibited a repressive effect when added during the elongation phase of the reaction (lane 12). Collectively, these data demonstrate that hNAP1 is not a prototypical transcriptional activator, but rather functions on chromatin templates during the PIC assembly step to further stimulate activated transcription.
Human NAP1 modulates the repressive function of histone H1 to enhance transcription
We next focused on the mechanism of hNAP1 transcriptional enhancement during PIC formation. Assembly of the PIC occurs following the addition of nuclear extract, as it supplies RNAP II, mediator, and the numerous additional factors essential for transcription initiation. Previous studies have reported that nuclear extracts also contain the linker histone H1, and that H1 is repressive to in vitro transcription [51–53]. Based on these observations, we hypothesized that hNAP1 enhances activated transcription via counteracting the repression induced by H1. To directly test this hypothesis, we added purified, recombinant H10 (rH10) at the beginning of the PIC assembly step. Figure 2a shows that exogenous H10 repressed activated transcription (lanes 5–7). However, addition of increasing amounts of hNAP1 reversed rH10 repression in a dose-dependent manner (lanes 8–10). Addition of rH10 to reactions performed on naked DNA templates produced only a modest effect on transcription, indicating that H1-repression requires a chromatin context (Fig. 2b). Together, these data confirm that H1 is repressive to transcription in our in vitro system, and provide evidence that hNAP1 functions to enhance transcription by directly counteracting H1. We next examined the transcriptional stimulatory effect of hNAP1 on our 900 bp promoter fragment assembled into chromatin in the presence of equimolar amounts of rH10 and histone octamer (H10-chromatin). To verify the incorporation of the linker histone into our chromatin templates, we measured salt-dependent chromatin condensation [54]. Relative to chromatin arrays assembled with our promoter fragment in the absence of the linker histone, the rH10-chromatin arrays precipitated at a lower Mg2+ concentration, indicating that H1 incorporation facilitated inter-array self-association and chromatin condensation (Fig. 2c). We next compared the transcriptional competences of rH10-chromatin vs. chromatin to ascertain the functional consequences of H1 incorporation. Figure 2d shows that rH10-chromatin was refractory to transcriptional activation (compare lanes 1, 2 and 5, 6). Importantly, the addition of hNAP1 reversed the potent repression induced by H1 incorporation into chromatin (compare lanes 6 and 8).
Fig. 2.
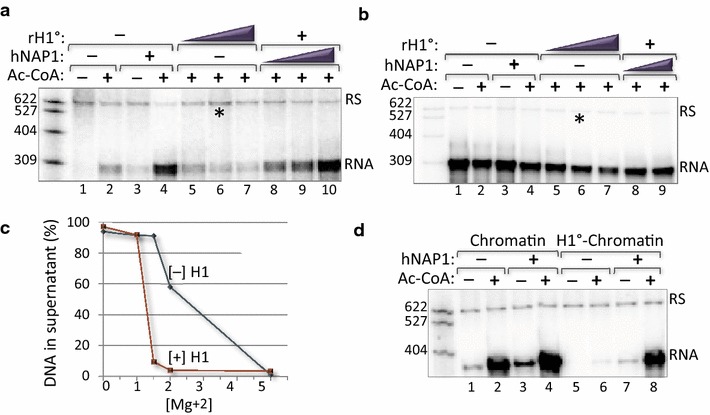
hNAP1 opposes the repressive function of linker histone H1 to enhance activated transcription in vitro. a hNAP1 reverses H1-induced transcriptional repression. In vitro transcription reactions were performed on the 588 bp HTLV-1/G-less cassette fragment as described (Fig. 1b), in the presence of activators and in the absence or presence of Ac-CoA, hNAP1, or rH10. Reactions were supplemented with increasing amounts of rH10 (0.03, 0.06, and 0.12 µM) at the start of the PIC assembly step (lanes 5–7). Reactions containing a constant amount of rH10 (0.06 µM) were supplemented with increasing amounts of hNAP1 (0.17, 0.34 and 0.67 µM) (lanes 8–10). Asterisk denotes the rH10 concentration used in reactions supplemented with varying amounts of histone chaperone. b Transcription from unassembled (naked) promoter templates is only modestly affected by H1 and hNAP1. The experiment was performed as described in panel a, except only two amounts of hNAP1 (0.34 and 0.67 µM) were added to reactions containing rH10 (lanes 8, 9). c rH10-induced chromatin compaction. Chromatin was assembled on the 900 bp HTLV-1/G-less cassette fragment in the absence or presence of equimolar amounts of rH1.0 (H1:octamer), as indicated. The chromatin templates were incubated with increasing amounts of MgCl2, precipitated, and the percentage of DNA from chromatin prepared in the absence ([−H1], blue) or presence ([+H1], red) remaining in the supernatant following centrifugation was plotted. The graph shown is representative of three experiments. d hNAP1 relieves potent transcriptional repression induced by H10-chromatin. Chromatin was assembled on the 900 bp HTLV-1/G-less cassette fragment in the absence or presence of equimolar rH1.0 (H1:octamer) (see “Methods”). hNAP1 (0.67 µM) was added to in vitro transcription reactions performed on chromatin (lanes 1–4) or H1-chromatin (lanes 5–8) templates, in the presence of activators, and in the absence or presence of Ac-CoA. Note that the 900 bp fragment produces an RNA transcript 390 nt in length
The NAP-family protein, SET/Taf1β, similarly modulates the repressive function of histone H1 to enhance transcription
The data presented above provide strong evidence that hNAP1 enhances transcription through opposing H1 repression. The physiological significance of these observations is uncertain, however, as many NAP-family members have been reported to reside primarily, although not exclusively, in the cytoplasm [21, 55, 56]. SET/Taf1β (SET) shares structural similarity to NAP1, however, it has been shown to reside primarily in the nucleus [45, 57]. Further, SET has been implicated in chromatin decondensation, H1 dynamics, transcriptional regulation, and PIC formation [34, 35, 38, 39, 45, 57–60]. Based on these observations, we turned our focus to SET as a potential candidate histone chaperone that may also function to enhance transcription by opposing H1-repression. We first analyzed the sub-cellular localization of hNAP1 and SET/Taf1β by immunofluorescence and cell fractionation. Figure 3a, b reveal that, in the cell lines examined, hNAP1 is predominately cytoplasmic, whereas SET is predominately nuclear. However, it should be noted that the NAP-family proteins carry both nuclear import and export signals. As such, NAP may perform chromatin-related nuclear activities either transiently, under certain cellular conditions (e.g., during development or a specific phase of the cell cycle), and/or function at relatively low concentrations. We next examined SET function during the individual steps in the transcription reaction. Similar to hNAP1 (see Fig. 1e), SET enhanced activated transcription during the PIC assembly step of initiation (Fig. 3c). Increasing concentrations of SET produced a dose-dependent stimulation of transcription from the chromatin-assembled templates, but had no effect on transcription from naked DNA (Fig. 3d). Importantly, SET also reversed the repression induced by the addition of rH10 to the transcription reactions, and countered the potent repression induced by rH10-chromatin (Fig. 3e, f). Taken together, these observations indicate that both hNAP1 and SET likely function in a mechanistically similar way to enhance transcription by counteracting H1 repression.
Fig. 3.
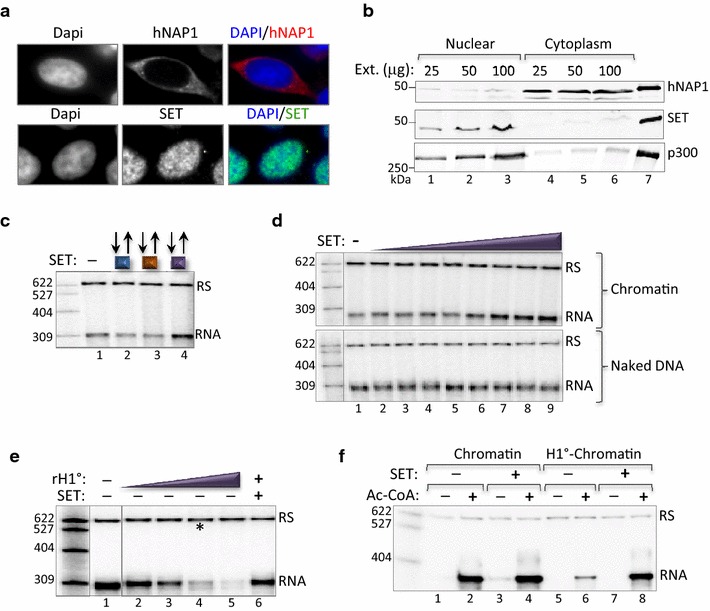
SET/Taf1β (SET), opposes the repressive function of H1 to enhance activated transcription in vitro. a Immunofluorescence labeling of hNAP1 (red, upper panel) and SET (green, lower panel) reveals distinct subcellular localization of the histone chaperones in HEK-293T cells. DAPI labeling of DNA is shown in blue. For single channel staining, black and white gave better resolution. For dual channel staining, color images were used to distinguish between the two antibodies. b Western blot analysis of hNAP1 and SET in nuclear and cytoplasmic extracts prepared from CEM T-cells. Immunostaining of nuclear p300 was performed to monitor efficiency of fractionation. Recombinant, purified hNAP1, SET, and p300 were analyzed in parallel as a positive control (lane 7). Recombinant hNAP1 migrates slightly slower due to the His6 purification tag. The position of the molecular weight size markers (kDa) is shown. c Like hNAP1, SET enhances activates transcription during the PIC assembly step. The in vitro transcription reaction was performed exactly as described for hNAP1 in Fig. 1e (lanes 9–12), in the presence of activators and SET (0.67 µM). d Dose-dependent enhancement of transcription by SET requires chromatin. In vitro transcription assay performed with increasing amounts of SET (0.03, 0.06, 0.12, 0.2, 0.27, 0.33 and 0.67 µM) added during PIC assembly produced a dose-dependent enhancement of transcription from chromatin templates (upper panel), but not from naked DNA (lower panel). The experiment was performed in the presence of activators and Ac-CoA. e SET reverses rH10-induced transcriptional repression. The in vitro transcription experiment was performed as described in Fig. 2a, with increasing amounts of rH10 (0.015, 0.03, 0.06, and 0.12 μM) (lanes 2–5), or a constant amount of rH10 (0.06 μM*) and SET (0.34 μM) (lane 6), in the presence of activators and Ac-CoA. f SET reverses transcriptional repression induced by H10-chromatin. In vitro transcription reactions were performed on the 900 bp HTLV-1/G-less cassette fragment, exactly as described for Fig. 2d, using SET (0.67 µM) in place of hNAP1. Reactions were performed in the presence of activators, and the absence or presence of SET and/or Ac-CoA, as indicated
SET/Taf1β and hNAP1 directly bind H1 and evict the linker histone from chromatin
We next investigated the mechanism of histone chaperone-mediated anti-repression. Using the immobilized template assay, and binding conditions designed to precisely recapitulate the transcription reactions shown in Figs. 2a and 3e, we found that H1 supplied by the nuclear extract, or rH10 added exogenously to the reaction, directly bound the chromatin-assembled promoter template (Fig. 4a, b, lane 1). Interestingly, based on the input amounts, the nuclear extract contributes significantly more total H1 to the binding reaction than exogenously added rH10 (Panels a, b, lanes 3). However, the rH10 appears to bind more efficiently to the templates, suggesting that a significant percentage of the endogenous H1 may be modified or in complex with other components in the reaction, and thus is unavailable for binding. Of note, the addition of hNAP1 and SET to the binding reactions reduced H1 binding to the template (Fig. 4a, b, lane 2). These data confirm that H1 is present in the nuclear extracts and that it binds to the transcription templates. Further, H1 binding likely imparts a moderately repressive effect on transcription that is relieved, via displacement, by the histone chaperones. We next measured the effect of hNAP1 and SET on H1 incorporated into chromatin. Following H1-chromatin assembly, the immobilized templates were incubated in the absence or presence of activators, p300, hNAP1 and SET. Figure 4c shows that both chaperones significantly reduced the amount of H1 incorporated into chromatin, and that H1 displacement was independent of activators and histone acetylation by Ac-CoA (lanes 3–6). Analysis of the supernatant fraction confirmed the presence of evicted H1, but not the core histones (data not shown). Of note, the chaperone-mediated release of rH10 from the templates correlated with increased transcription factor/coactivator binding, an observation consistent with chromatin relaxation following H1 release. To further decipher the mechanism of chaperone-dependent H1 displacement, we performed GST pull-down assays and confirmed that both chaperones directly bind rH10 (Fig. 4d) [43–45]. Interestingly, these observations support a pathway for chaperone-mediated H1 eviction; however, the mechanism of chaperone recruitment to H1-containing promoters remains elusive. Previous reports, together with our unpublished studies, have shown that the NAP-family proteins directly bind to CBP/p300 [61, 62], suggesting a potential mechanism of coactivator-mediated recruitment to promoters targeted for activation. However, as shown in Fig. 4d, we did not observe enhanced association of either hNAP1 or SET in binding reactions containing p300. In general, we found that chaperone binding to the immobilized template varies between experiments, but is generally low or undetectable. From these observations, we hypothesized that a chaperone–coactivator complex exists in solution, and that the chaperone is released upon coactivator recruitment to the promoter. To test this hypothesis, we examined full-length p300 binding to GST-SET, in the absence or presence of the enhancer-bound Tax/pCREB complex. GST pull-down assays confirmed complex formation between p300 and SET in solution (Fig. 4e, lanes 2, 3). However, when the p300/SET interaction was analyzed in the presence of the Tax/pCREB/vCRE DNA complex, which binds p300 with high affinity, the p300/SET interaction was abolished (lanes 4, 5). In contrast, when the p300/SET complex was similarly challenged with Tax/vCRE DNA, which does not bind p300, the p300/SET interaction was unaffected (lanes 5, 6). These data support a model in which the histone chaperone is initially recruited to the promoter via direct interaction with CBP/p300, followed by chaperone release from the coactivator to facilitate H1 eviction. Together, these findings provide a mechanistic foundation for the anti-repression activity of the histone chaperones.
Fig. 4.
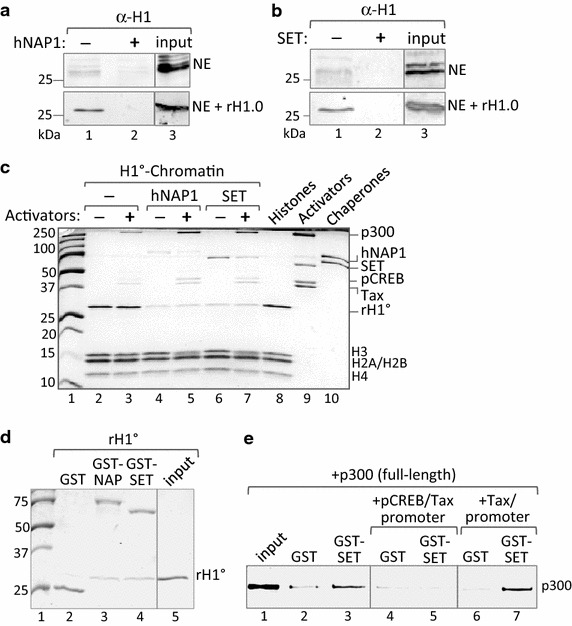
hNAP1 and SET evict H1 from chromatin. a, b hNAP1 and SET remove endogenous H1 and rH10 from chromatin templates. Immobilized chromatin templates (0.33 ng/µl) were incubated with CEM NE (upper panel) (~0.55 µg/µl), or NE supplemented with rH10 (lower panel) (0.067 µM; ~fourfold excess rH10 relative to octamer) in the absence or presence of hNAP1 (panel a) or SET (panel b) (0.67 µM each). The binding assays were designed to replicate the conditions of the in vitro transcription reactions shown in Fig. 2 (hNAP1) and 3 (SET) (i.e., transcription in the presence of NE, or NE supplemented with exogenous rH10). For each panel, input NE (15 %) and rH10 (100 %) are shown (lane 3). Note that input H1 appears similar due to saturating amounts of protein. H1 binding was assessed by Western blot using an antibody against rH10. c hNAP1 and SET remove H1 incorporated into chromatin. The 900 bp immobilized promoter fragment was assembled into H10-chromatin, followed by incubation with activators, Ac-CoA, and histone chaperones, as indicated. For reference, input amounts (100 %) of histones (octamer, rH10), activators (Tax, CREB, p300), and chaperones were analyzed in parallel (lanes 7–9). Note that in the reactions containing activators, p300, Ac-CoA, and histone chaperones, p300 acetylation caused diffuse migration of the H3 band (lanes 4, 6). Under the conditions of the experiment, only histone H1 was evicted from the promoter template. Reactions were analyzed by SDS-PAGE followed by Coomassie staining. Molecular weight size markers (kDa) are shown (lane 1). d hNAP1 and SET directly bind rH10 as demonstrated by GST pull-down assay. Input rH10 (50 %) (lane 5) and molecular weight size markers (kDa) (lane 1) are shown. Binding reactions were analyzed by SDS-PAGE followed by Coomassie staining. e SET binding to full-length p300 is abolished in the presence of enhancer-bound Tax/pCREB complex. GST pull-down assays were performed to measure the binding of p300 to SET in the absence or presence of HTLV-1 promoter-bound activators (Tax/pCREB). Full-length p300 bound to GST-SET alone (lanes 2, 3), but was disrupted in the presence of the Tax/pCREB-bound promoter DNA (which serves as a high affinity binding site for p300) (lanes 4, 5). The p300/SET interaction was unaffected under the same conditions, but in the absence of pCREB (lanes 6, 7). Input p300 (100 %) is shown (lane 1). Binding was analyzed by Western blot using an antibody against p300
SET/Taf1β enhances activator-dependent transcription in vivo
We next asked whether the NAP family histone chaperones functioned to enhance activated transcription in vivo. To recapitulate our in vitro model system, we utilized a cell line that carries the chromosomally-integrated HTLV-1 promoter linked to luciferase [63]. Of note, we previously demonstrated that Tax-induced transcriptional activation in this cell line correlated with eviction of H1 from the HTLV-1 promoter [64]. To test whether expression of the chaperones enhanced activated transcription, we co-transfected an expression plasmid for Tax, in the absence or presence of increasing amounts of the expression plasmids for SET or hNAP1. As expected, Tax dramatically stimulated expression from the integrated HTLV-1 promoter (150-fold) (Fig. 5a). Importantly, co-transfection of SET, but not hNAP1, significantly enhanced Tax transactivation in a dose-dependent manner. Expression of the chaperones was confirmed by immunoblot analysis (Fig. 5b). Consistent with our in vitro data, the stimulatory effect of SET required conditions of activated transcription, as SET had no effect in the absence of Tax (Fig. 5c). Moreover, Tax-activated transcription was refractory to stimulation by SET when assayed on a transiently transfected HTLV-1 promoter reporter plasmid (Fig. 5d), consistent with a requirement for properly assembled H1-chromatin [65]. To investigate the potential stimulatory effect of other NAP-superfamily histone chaperones on transcriptional activation, we also tested Taf1α, an isoform of SET/Taf1β, and hNAP2 (hNAP1L4). Like SET, transfection of Taf1α also enhanced Tax transactivation (Fig. 5e). Similarly, like hNAP1, the related hNAP2 had no effect on transcription from the integrated model promoter (Fig. 5f). These findings consistent with the respective subcellular localization of the histone chaperones, as well as the in vitro binding and functional data, and together support a role for SET in transcriptional activation via modulation of H1 dynamics.
Fig. 5.
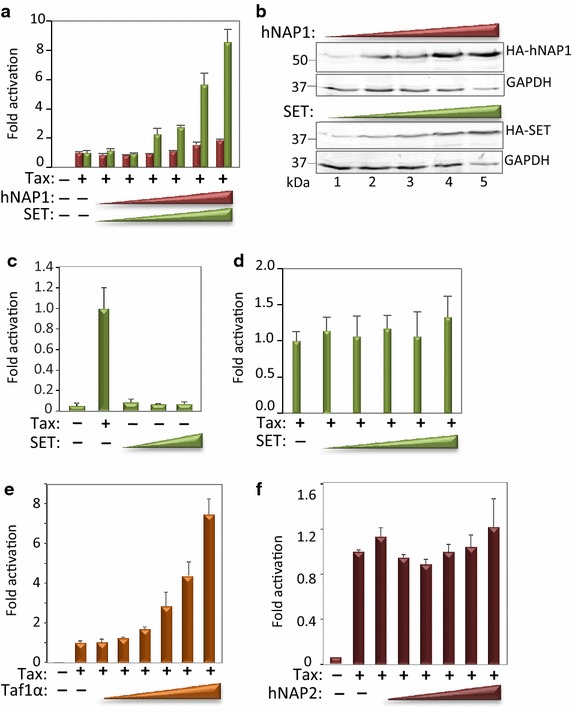
SET enhances activated transcription in vivo. a SET, but not hNAP1, enhanced activated transcription from the chromosomally-integrated model promoter. CHOK-Luc cells, carrying multiple integrated copies of the HTLV-1 promoter linked to luciferase [63] were transiently transfected with an expression plasmid for Tax alone (10 ng) or co-transfected with increasing amounts (25, 50, 100, 200, 400, 500 ng) of HA-hNAP1 or HA-SET expression plasmids. The signal obtained in the presence of Tax alone was normalized to 1, and fold activation by the histone chaperones, relative to Tax alone, is reported. The experiment was performed in triplicate, and is representative of three biological replicates. b Expression of HA-hNAP1 (upper panel) and HA-SET (lower panel) following transfection. CHOK-Luc cells were transfected as described in panel a, and lysates analyzed by immunoblot using an anti-HA antibody. Immunoblot analysis of GAPDH was performed in parallel as a loading control. c SET enhancement of transcription requires co-transfection of Tax. CHOK-Luc cells were transiently transfected with an expression plasmid for Tax (10 ng) or the expression plasmid for SET (200, 400, 500 ng), as indicated. d The transiently transfected HTLV-1 promoter/luciferase reporter plasmid is refractory to enhancement by co-transfected SET. Jurkat T-cells were transiently co-transfected with the HTLV-1/Luc reporter plasmid (100 ng) [87], and expression plasmids for Tax and SET, as described in panel a. e SET/Taf1α, an isoform of SET/Taf1β, enhanced activated transcription from the integrated model promoter. The experiment was performed as described in panel a, with co-transfected Taf1α in place of SET. f Co-transfection of hNAP2 had a negligible effect on activated transcription from the integrated model promoter. The experiment was performed as described in panel a
Discussion
The studies presented herein provide the first report to directly link the histone chaperones NAP1 (hNAP1L1) and SET/Taf1β with histone H1 displacement and transcriptional activation. Using a powerful, chromatin-based in vitro system, we demonstrate that both human NAP1 and SET enhance transcription from an immobilized native promoter template. Transcriptional enhancement is dependent upon a chromatin environment, and the presence of activators (Tax and pCREB), p300, and Ac-CoA. The underlying requirement for transcriptionally permissive conditions indicates that the histone chaperones exert their stimulatory effect in a manner distinct from conventional DNA-binding transcription factors. To explore the mechanism of transcriptional enhancement, we examined histone chaperone function during each temporally-distinct step of the initiation reaction. We initially found that both hNAP1 and SET exert their stimulatory activity during PIC assembly, immediately following nuclear extract addition. Based on the unconventional activation properties of the histone chaperones, we considered the possibility that they functioned to enhance transcription via displacement of a repressor, thus facilitating PIC formation. Previous studies found that nuclear extracts contain the repressive linker histone H1, and that H1 is potently repressive to transcription [51–53]. Since both chaperones bind to the linker histones, we reasoned that hNAP1 and SET might function as anti-repressors; enhancing transcription by opposing H1-induced repression. Using the immobilized template assay, we find that endogenous and exogenous H1 bind to the chromatin-assembled model promoter template, and that exogenous H1 potently represses transcription. The level of transcription stimulation by the chaperones directly correlates with the amount of H1 present in the assay. While both chaperones stimulated transcription on chromatin templates without exogenously added H1 (likely through displacement of H1 in the nuclear extract), their overall stimulatory effect is appreciably enhanced in the presence of exogenously added H1. Significantly, restoration of high-level activated transcription by the histone chaperones directly correlates with reduced H1 binding to the template, increased activator and coactivator binding, and histone H3 acetylation; all conditions essential for strong transcription. Our findings are consistent with a previous study showing that both NAP1 and SET enhance activated transcription on chromatin templates in vitro, at a step prior to elongation [35]. However, the mechanism of transcriptional enhancement—specifically the involvement of H1—was not identified.
We find that in vitro, NAP1 and SET function in a mechanistically indistinguishable manner to derepress transcription from our model promoter. In vivo, however, SET, but not hNAP1, significantly enhances transcription from the stably integrated HTLV-1 promoter/reporter construct. This observation is consistent with the predominant cytoplasm localization of NAP1 and nuclear localization of SET. Because H1 is an abundant nuclear protein, it is improbable that hNAP1 would have a general role in modulating H1 dynamics in the cell lines examined. It should be noted, however, that NAP1 carries both nuclear import and export signals [12, 66]. Furthermore, NAP1 has been shown to bind both core and linker histones, and has been implicated in numerous transcription-related processes [10, 11]. Based on these observations, the nuclear distribution of hNAP1, and thus its role in H1 dynamics, may be influenced by post-translational modification (phosphorylation or glutamylation), host cell and/or stage of differentiation, or the presence of a compatible dimerization partner. We also found that the anti-repressive activity of SET in vivo is dependent upon Tax expression, a result that corroborates our in vitro observations. Furthermore, SET enhanced transcription from the chromosomally integrated model promoter/reporter construct, but had no effect when the same reporter construct was transiently transfected into the cells. This finding underscores the requirement for a native chromatin environment for SET transcriptional enhancement function, and further implicates H1 as a major target of SET. Moreover, we previously detected significant eviction of H1 from the integrated HTLV-1 promoter region in this same cell line [64]. H1 loss was dependent upon Tax expression, and correlated with activator and coactivator binding, RNA polymerase II recruitment, and strong transcriptional activation. Our previous study demonstrating activator-dependent H1 eviction, together with the studies presented herein, are consistent with a model of chaperone-mediated anti-repression via eviction of H1. Displacement of the linker histone is likely an early event required for relaxation of the chromatin fiber, followed by nucleosome mobilization and PIC formation. Although the role of the linker histones in gene regulation has been controversial [4, 67], recent genome-wide profiling studies demonstrate significantly reduced H1 association at active promoter regions [7–9]. These observations are consistent with a model of activated transcription in which H1 displacement is an early event in transcriptional initiation. The strong correlation between our in vivo and in vitro data support a role for SET as an H1 chaperone responsible for transcriptional derepression mediated through the targeted displacement of the repressive linker histone.
These studies begin to address the mechanism by which SET is recruited to the promoters of genes targeted for H1 displacement. In a previously published study examining heat shock genes in Drosophila polytene chromosomes, SET was shown to dramatically redistribute to transcriptionally active gene regions following heat shock [31], however, the mechanism/s that facilitated targeted relocalization of SET to the heat shock loci remained elusive. As described herein, H1 eviction directly correlates with the binding of the activators and p300 to the model promoter. These observations infer a causal relationship between activator/coactivator binding and H1 dissociation. Further, they implicate a role for the transcription factors and/or p300 in the delivery of SET to the viral enhancer elements for H1 dissociation. In support of this idea, previous studies indicate that SET may directly interact with CBP/p300 and specific transcription factors [68]. These studies offer precedence for a mechanism of direct recruitment of SET to enhancer regions prior to transcriptional activation. While we did not detect stable association of SET with promoter-bound p300 in vitro, we have confirmed an interaction between the two proteins in solution. Moreover, we find that the SET/p300 complex is destabilized upon p300 binding to the HTLV-1 enhancer. These data provide preliminary support for a model of targeted recruitment of SET via coactivator binding and subsequent release at the promoter to facilitate local eviction of H1. Of note, we observe the binding of activators and p300 to H1-chromatin, albeit at a reduced level relative to chromatin depleted of H1 (see Fig. 4c). These observations provide support for a model in which activators and coactivators associate with the promoters of genes targeted for activation prior to chromatin decondensation, and thus provide a mechanism for the precise delivery of SET, followed by H1 displacement. As such, H1 displacement may be an early event in initiation, but one that occurs subsequent to the binding of specific transcription factors and/or coactivators that serve to elicit the cascade of events that culminate in transcriptional activation.
Conclusions
We biochemically characterized a novel anti-repressor function of members of the NAP superfamily; the opposition of H1 transcriptional repression mediated by the physical displacement of H1 from chromatin. Eviction of the linker histone by the chaperones SET and hNAP1 promotes chromatin decompaction and the creation of a relaxed chromatin environment permissive to PIC formation and transcriptional initiation. Although NAP1 and SET function indistinguishably in vitro to displace H1 from chromatin and enhance activated transcription, the abundant nuclear localization of SET, together with strong enhancement of activated transcription in vivo, collectively support a biologically-relevant role for SET as a novel transcriptional anti-repressor that functions via modulation of H1 dynamics. Of note, previous in vivo studies correlated SET overexpression with chromosomal decondensation and enhanced H1 exchange [45, 58, 69, 70], whereas silencing of SET correlated with abnormally condensed chromosomes [45]. Our novel biochemical studies integrate the numerous previous studies on SET, and identify a direct link between SET and H1 mobilization. Together, our findings support a mechanism for achieving chromatin decompaction in vivo, and point to a physiologically-relevant role for SET in targeting specific genes for transcriptional activation.
Methods
Plasmid constructs, protein expression, purification, nuclear extract
The mammalian reporter plasmid, HTLV-Luc [71], and expression plasmids, pSG-Tax [72], pcDNA-HA-hNAP1 and pCMV-myc-hNAP2 [73] have been described previously. The HA-SET/Taf1 (pcDNA-HA-Taf1α/β) mammalian expression plasmid was subcloned from the bacterial expression plasmid [74]. Each of the following recombinant proteins used in this study were expressed and purified as described in their accompanying references, or using established protocols: Tax-His6 [75], ser-133 phosphorylated CREB (pCREB) [76, 77], GST-hNAP1 and His6-hNAP1 [73], His6-dNAP [78], His6-GST-SET/Taf1β, and linker histone rH1.0 [79, 80]. Purified, recombinant X. laevis core histones were assembled into octamer as previously described [46, 81].
Cell culture, nuclear extracts, luciferase assays
CHOK1-Luc hamster ovary cells (gift from Dr. Kuan-Teh Jeang) [63], HEK-293T, and HeLa cells were cultured in DMEM (Sigma) supplemented with 10 % fetal bovine serum (FBS), 2 mM l-glutamine, and penicillin–streptomycin. CHOK1-Luc cells were supplemented with 500 μg/ml G418 to maintain the integrated HTLV-1/Luc reported construct. CEM T-cells and Jurkat T-cells were maintained in IMDM (Gibco) supplemented with 10 % FBS, 2 mM l-glutamine, and penicillin–streptomycin. HeLa S3 cells, grown in suspension for nuclear extract preparation, were cultured in IMDM supplemented with 2 % FBS, 2 mM l-glutamine, and penicillin–streptomycin. Nuclear extracts for transcription assays were prepared from HeLa S3 or CEM cells as previously described [82]. Both extracts gave the same results in the transcription assays. Luciferase assays using CHOK1-Luc cells [63] or Jurkat T-cells were performed as previously described [64]. Briefly, 1 × 105 cells were seeded into a 24-well plate and transfected with the indicated plasmids and a constant amount of DNA (1 μg) using Lipofectamine reagent (Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s protocol. After 48 h the cells were harvested and lysed, and luciferase activity was measured using a dual-luciferase reporter assay system (Promega). Luciferase activity was detected using a Turner Designs luminometer (TD-20/20). To control for transfection efficiency, cells were co-transfected with Renilla, and firefly luciferase activity was normalized to Renilla activity (pRL-TK, Promega, Madison, WI, USA). Because the luciferase signal is very low in the absence of transfected Tax, we set the signal obtained in the presence of Tax to 1, and normalized all other samples relative to the signal obtained with Tax alone. The transient transfection assays were performed in triplicate with at least three biological replicates.
Chromatin assembly
Immobilized templates (588 or 900 bp) carrying the HTLV-1 promoter linked to a G-less cassette (HTLV-1/G-less) were generated by PCR using an upstream biotinylated primer. The biotinylated promoter fragments were immobilized on streptavidin-coated magnetic Dynabeads (Invitrogen) according to the manufacturer’s instruction. The immobilized DNA templates were assembled into chromatin (0.8:1 histone octamer/DNA ratio [wt/wt]) by salt deposition as described previously [48]. Briefly, bead-bound DNA fragment and histone octamer were mixed at 4 °C at 1300 rpm in 10 mM Tris, pH 8.0 mM EDTA (TE) containing 1 M NaCl and 0.1 % Chaps. The NaCl concentration was diluted stepwise to 0.8, 0.6, 0.4, 0.2, 0.1 M, with a 30 min. agitation per step. All transcription reactions, except those with H10-chromatin, were performed with the 588 bp promoter fragment, which gives a transcript 310 nt in length. For H10-chromatin assembly, equimolar amounts of rH10 and histone octamer were added at the beginning of chromatin assembly using the 900 bp fragment. The longer fragment gives a transcript 390 nt in length. The longer fragment was initially used to promote higher order H1-chromatin structure formation, however we obtained essentially identical transcription results with either the 588 or 900 bp fragments assembled into H1-chromatin. The assembled templates were stored at 4 °C in storage buffer (1 mM EDTA, 10 mM NaCl, 0.1 % NP40, 0.5 % Chaps, 20 % glycerol, 2 mM DTT) at a DNA concentration of 100 ng/μl, and used within 1 week.
In vitro transcription assay
The immobilized chromatin-assembled promoter templates (50 ng; 0.004 μM) were used in transcription assays as previously described [17, 18]. Briefly, the bead-bound promoter fragments were incubated with pCREB, Tax, and Ac-CoA (as indicated in each reaction) for 15′ at 30 °C with shaking (1300 rpm) followed by the addition of p300 for 20′ (see Fig. 1c). Nuclear extract (~15 μg) was added to each reaction and incubated for an additional 60′ in a final volume of 30 μl in 0.5× TM buffer (25 mM Tris, pH 7.9, 50 mM KCl, 6.25 mM MgCl2, 10 % glycerol, 0.5 mM EDTA, 0.5 mM DTT). RNA synthesis was initiated by addition of 224 μM ATP, 224 μM CTP, and 0.75 μM [α-32P] UTP (3000 Ci/mmol), and the elongation reaction incubated for an additional 40 min. Reactions were stopped, RNA was isolated following digestion with Proteinase K, and transcripts were separated by denaturing PAGE and analyzed using ImageQuant® software. Molecular weight markers (radiolabeled HpaII-digested pBR322) were used to estimate the sizes of the RNA products. A labeled 622-bp DNA fragment was added to each reaction mixture as a recovery standard. Unless otherwise indicated, constant amounts of the following purified, recombinant proteins were used: pCREB (0.03 μM), Tax (0.06 μM), p300 (0.02 μM), hNAP1 (0.67 μM), SET/Taf1β (0.67 μM) and Ac-CoA (100 μM). Experiments in which the chaperone was added and removed at a specific step were performed as follows: after the indicated incubation, the supernatant was removed by magnetic isolation of the immobilized transcription templates, and replaced with supernatant generated in parallel, but lacking the histone chaperone. Using this approach, the overall steady-state concentrations of each reaction component remained constant. All transcription experiments were performed at least three times, with each replicate giving essentially identical results.
Immobilized template assay
Binding reactions were prepared as described for the in vitro transcription assays, except the reactions were increased 20-fold (1 μg of DNA) to enable visualization of template-bound proteins by Coomassie-stained gels. Unless otherwise indicated, reactions contained purified, recombinant pCREB (0.03 μM), Tax (0.06 μM), p300 (0.02 μM), hNAP1 or SET/Taf1β (0.67 μM), and Ac-CoA (100 μM) in 0.5× TM buffer. At the completion of the binding reaction, the bead-bound templates were isolated by magnetic separation, and washed twice with 0.5× TM buffer. Template-bound proteins were separated by 15 % SDS-PAGE and analyzed by Western blot (Fig. 4a, b) or Coomassie blue staining (Fig. 4c).
Analysis of H1-chromatin
Magnesium-dependent chromatin condensation assays were performed as previously described [83, 84]. Chromatin and H1-chromatin was assembled as described above, except using the method of salt dialysis, as described [85]. Following dialysis, arrays were incubated in the presence of the indicated concentration of MgCl2 and subjected to centrifugation for 10 min at 16,100g. DNA from both the pellet and supernatant fractions was purified by phenol/chloroform extraction and ethanol precipitation, and analyzed by 1 % agarose gel electrophoresis followed by ethidium bromide staining. DNA from each fraction was quantified using ImageQuant® software and plotted as percentage of total DNA in each reaction.
GST pull-down assay
GST pull-down assays were performed as described [86]. Briefly, purified GST alone, or GST-fused histone chaperone (25 pmol) were bound to 20 μl glutathione agarose beads in binding buffer (50 mM Tris–HCl, pH 7.9, 150 mM KCl, 12.5 mM MgCl2, 1 mM EDTA, 20 % glycerol, 0.5 % NP40, 0.1 % Chaps, and 100 ng/μl BSA) for 1 h at 4 °C, followed by incubation with an equal concentration of rH1.0 for an additional 16 h at 4 °C. The beads were washed extensively, and GST-bound proteins analyzed by 15 % SDS-PAGE and Coomassie blue staining (Fig. 4d) or western blot (Fig. 4e). For the GST pull-down reactions using full-length p300, the binding reactions contained GST alone or GST-SET (20 pmol) and full-length p300 (5 pmol). The binding reactions were performed in the absence or presence of activator/DNA complex composed of the 588 bp HTLV-1 promoter DNA fragment (2.2 pmol; 6.6 pmol binding sites) and either Tax alone (which does not bind DNA in the absence of CREB), or Tax/pCREB (30 pmol each, 15 pmol dimers). Binding reactions were performed as described above in the presence of 0.5× TM buffer supplemented with 0.1 % NP40 and 500 ng/μl BSA.
Antibodies, western blot and immunofluorescence labeling
Western blot and immunofluorescence labeling were performed according to the manufacturer’s instructions. Anti-hNAP1 (1:2000, Abcam #21630), anti-SET/Taf1β (1:2000, Abcam #1183), anti-p300 (1:300, Santa Cruz Biotechnology #584), anti-H1 (1:300, Santa Cruz Biotechnology #8030), anti-HA (Santa Cruz Biotechnology #SC-805), and anti-GAPDH (Santa Cruz Biotechnology #SC-137179) were used for western blot analysis. Immunofluorescence labeling in HEK-293T cells was performed using anti-hNAP1 (1:150, Abnova #H00004673) and anti-SET/Taf1β (1:200, Abcam #1183) primary antibodies and Alexa Fluor® 488 goat anti-rabbit IgG (H+L) (1:200, Invitrogen #A11008) and Alexa Fluor® 594 goat anti-mouse IgG (H+L) (1:200, Invitrogen #A11005) secondary antibodies. An Olympus IX81 spinning disk confocal microscope with 60×/1.42 NA objective and a Photometrics Cascade II CCD camera with SlideBook (Intelligent Imaging Innovations) software were used for fluorescence images. Metamorph software was used to process images.
Image processing
Solid vertical lines on some gels indicate where the image (from the same gel) was spliced to either remove irrelevant lanes or enhance presentation of the data. Minimal and uniform brightness/contrast was applied to some gel images.
Authors’ contributions
QZ, MKI and JKN conceived the study. QZ, HAG and MKI performed the experiments. JKN, HAG and QZ wrote the manuscript. MKI provided critical comments on revising the manuscript. All authors read and approved the final manuscript.
Acknowledgements
We thank L. Frappier for the SET/Taf1β expression plasmid, Dr. Hataichanok (Mam) Sherman and the Protein Expression and Purification Facility (CSU) for providing many of the purified proteins, members of Karolin Luger’s laboratory for optimization of rH10 purification, and members of the Nyborg laboratory and P01 group for numerous helpful discussions. This research was supported by P01GM88409 from the National Institutes of Health (to JKN).
Compliance with ethical guidelines
Competing interests The authors declare that they have no conflict of interest.
Abbreviations
- NAP
nucleosome assembly protein
- SET
also called SET/Taf1β, template activating factor-1 (Taf-1), INHAT, and IPP2A
- TSS
transcription start site
- CREB
cAMP response element binding protein
- CBP
CREB binding protein
- pCREB
phosphorylated CREB
- Ac-CoA
acetyl coenzyme A
- HTLV-1
human T cell leukemia virus, type 1
- PIC
preinitiation complex assembly
- vCRE
viral cAMP response element
- GST
glutathione-S-transferase
Contributor Information
Qian Zhang, Email: qian.zhang@colostate.edu.
Holli A. Giebler, Email: holli.giebler@colostate.edu
Marisa K. Isaacson, Email: misaacson2@pace.edu
Jennifer K. Nyborg, Phone: (970) 491-0420, Email: Jennifer.nyborg@colostate.edu
References
- 1.Luger K, Mader AW, Richmond RK, Sargent DF, Richmond TJ. Crystal structure of the nucleosome core particle at 2.8 a resolution. Nature. 1997;389(6648):251–260. doi: 10.1038/38444. [DOI] [PubMed] [Google Scholar]
- 2.Woodcock CL, Ghosh RP. Chromatin higher-order structure and dynamics. Cold Spring Harb Perspect Biol. 2010;2(5):a000596. doi: 10.1101/cshperspect.a000596. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Happel N, Doenecke D. Histone h1 and its isoforms: contribution to chromatin structure and function. Gene. 2009;431(1–2):1–12. doi: 10.1016/j.gene.2008.11.003. [DOI] [PubMed] [Google Scholar]
- 4.Bustin M, Catez F, Lim JH. The dynamics of histone H1 function in chromatin. Mol Cell. 2005;17(5):617–620. doi: 10.1016/j.molcel.2005.02.019. [DOI] [PubMed] [Google Scholar]
- 5.Zhou BR, Feng H, Kato H, Dai L, Yang Y, Zhou Y, et al. Structural insights into the histone H1-nucleosome complex. Proc Natl Acad Sci USA. 2013;110(48):19390–19395. doi: 10.1073/pnas.1314905110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Heintzman ND, Stuart RK, Hon G, Fu Y, Ching CW, Hawkins RD, et al. Distinct and predictive chromatin signatures of transcriptional promoters and enhancers in the human genome. Nat Genet. 2007;39(3):311–318. doi: 10.1038/ng1966. [DOI] [PubMed] [Google Scholar]
- 7.Izzo A, Kamieniarz-Gdula K, Ramirez F, Noureen N, Kind J, Manke T, et al. The genomic landscape of the somatic linker histone subtypes h1.1 to h1.5 in human cells. Cell Rep. 2013;3(6):2142–2154. doi: 10.1016/j.celrep.2013.05.003. [DOI] [PubMed] [Google Scholar]
- 8.Cao K, Lailler N, Zhang Y, Kumar A, Uppal K, Liu Z, et al. High-resolution mapping of H1 linker histone variants in embryonic stem cells. PLoS Genet. 2013;9(4):e1003417. doi: 10.1371/journal.pgen.1003417. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Millan-Arino L, Islam AB, Izquierdo-Bouldstridge A, Mayor R, Terme JM, Luque N, et al. Mapping of six somatic linker histone H1 variants in human breast cancer cells uncovers specific features of H1.2. Nucleic Acids Res. 2014;42(7):4474–4493. doi: 10.1093/nar/gku079. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Hansen JC, Nyborg JK, Luger K, Stargell LA. Histone chaperones, histone acetylation, and the fluidity of the chromogenome. J Cell Physiol. 2010;224(2):289–299. doi: 10.1002/jcp.22150. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Das C, Tyler JK, Churchill ME. The histone shuffle: histone chaperones in an energetic dance. Trends Biochem Sci. 2010;35(9):476–489. doi: 10.1016/j.tibs.2010.04.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Park YJ, Luger K. Structure and function of nucleosome assembly proteins. Biochem Cell Biol. 2006;84(4):549–558. doi: 10.1139/o06-088. [DOI] [PubMed] [Google Scholar]
- 13.Lorch Y, Maier-Davis B, Kornberg RD. Chromatin remodeling by nucleosome disassembly in vitro. Proc Natl Acad Sci USA. 2006;103(9):3090–3093. doi: 10.1073/pnas.0511050103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Park YJ, Chodaparambil JV, Bao Y, McBryant SJ, Luger K. Nucleosome assembly protein 1 exchanges histone H2A-H2B dimers and assists nucleosome sliding. J Biol Chem. 2005;280(3):1817–1825. doi: 10.1074/jbc.M411347200. [DOI] [PubMed] [Google Scholar]
- 15.Andrews AJ, Chen X, Zevin A, Stargell LA, Luger K. The histone chaperone NAP1 promotes nucleosome assembly by eliminating nonnucleosomal histone DNA interactions. Mol Cell. 2010;37(6):834–842. doi: 10.1016/j.molcel.2010.01.037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Ito T, Ikehara T, Nakagawa T, Kraus WL, Muramatsu M. P300-mediated acetylation facilitates the transfer of histone H2A-H2B dimers from nucleosomes to a histone chaperone. Genes Dev. 2000;14(15):1899–1907. [PMC free article] [PubMed] [Google Scholar]
- 17.Sharma N, Nyborg JK. The coactivators CBP/p300 and the histone chaperone nap1 promote transcription-independent nucleosome eviction at the HTLV-1 promoter. Proc Natl Acad Sci USA. 2008;105(23):7959–7963. doi: 10.1073/pnas.0800534105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Luebben WR, Sharma N, Nyborg JK. Nucleosome eviction and activated transcription require p300 acetylation of histone H3 lysine 14. Proc Natl Acad Sci USA. 2010;107(45):19254–19259. doi: 10.1073/pnas.1009650107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Attia M, Rachez C, Avner P, Rogner UC. Nucleosome assembly proteins and their interacting proteins in neuronal differentiation. Arch Biochem Biophys. 2013;534(1–2):20–26. doi: 10.1016/j.abb.2012.09.011. [DOI] [PubMed] [Google Scholar]
- 20.Attia M, Rachez C, De Pauw A, Avner P, Rogner UC. Nap1l2 promotes histone acetylation activity during neuronal differentiation. Mol Cell Biol. 2007;27(17):6093–6102. doi: 10.1128/MCB.00789-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Okuwaki M, Kato K, Nagata K. Functional characterization of human nucleosome assembly protein 1-like proteins as histone chaperones. Genes Cells. 2010;15(1):13–27. doi: 10.1111/j.1365-2443.2009.01361.x. [DOI] [PubMed] [Google Scholar]
- 22.Muto S, Senda M, Akai Y, Sato L, Suzuki T, Nagai R, et al. Relationship between the structure of SET/TAF-Iβ/INHAT and its histone chaperone activity. Proc Natl Acad Sci USA. 2007;104:4285–4290. doi: 10.1073/pnas.0603762104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Li M, Guo H, Damuni Z. Purification and characterization of two potent heat-stable protein inhibitors of protein phosphatase 2a from bovine kidney. Biochemistry. 1995;34(6):1988–1996. doi: 10.1021/bi00006a020. [DOI] [PubMed] [Google Scholar]
- 24.Li M, Makkinje A, Damuni Z. The myeloid leukemia-associated protein set is a potent inhibitor of protein phosphatase 2a. J Biol Chem. 1996;271(19):11059–11062. doi: 10.1074/jbc.271.19.11059. [DOI] [PubMed] [Google Scholar]
- 25.von Lindern M, van Baal S, Wiegant J, Raap A, Hagemeijer A, Grosveld G. Can, a putative oncogene associated with myeloid leukemogenesis, may be activated by fusion of its 3′ half to different genes: characterization of the set gene. Mol Cell Biol. 1992;12(8):3346–3355. doi: 10.1128/mcb.12.8.3346. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Adachi Y, Pavlakis GN, Copeland TD. Identification and characterization of set, a nuclear phosphoprotein encoded by the translocation break point in acute undifferentiated leukemia. J Biol Chem. 1994;269(3):2258–2262. [PubMed] [Google Scholar]
- 27.Rosati R, La Starza R, Barba G, Gorello P, Pierini V, Matteucci C, et al. Cryptic chromosome 9q34 deletion generates TAF-1alpha/can and TAF-1beta/can fusion transcripts in acute myeloid leukemia. Haematologica. 2007;92(2):232–235. doi: 10.3324/haematol.10538. [DOI] [PubMed] [Google Scholar]
- 28.Saito S, Miyaji-Yamaguchi M, Nagata K. Aberrant intracellular localization of set-can fusion protein, associated with a leukemia, disorganizes nuclear export. Int J Cancer. 2004;111(4):501–507. doi: 10.1002/ijc.20296. [DOI] [PubMed] [Google Scholar]
- 29.Van Vlierberghe P, van Grotel M, Tchinda J, Lee C, Beverloo HB, van der Spek PJ, et al. The recurrent set-nup214 fusion as a new hoxa activation mechanism in pediatric T-cell acute lymphoblastic leukemia. Blood. 2008;111(9):4668–4680. doi: 10.1182/blood-2007-09-111872. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Dong L, Zhu J, Wen X, Jiang T, Chen Y. Involvement of set in the wnt signaling pathway and the development of human colorectal cancer. Oncology letters. 2014;7(4):1203–1208. doi: 10.3892/ol.2014.1866. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Nowak SJ, Pai CY, Corces VG. Protein phosphatase 2a activity affects histone H3 phosphorylation and transcription in drosophila melanogaster. Mol Cell Biol. 2003;23(17):6129–6138. doi: 10.1128/MCB.23.17.6129-6138.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Chambon JP, Touati SA, Berneau S, Cladiere D, Hebras C, Groeme R, et al. The pp2a inhibitor i2 pp2a is essential for sister chromatid segregation in oocyte meiosis II. Curr Biol. 2013;23(6):485–490. doi: 10.1016/j.cub.2013.02.004. [DOI] [PubMed] [Google Scholar]
- 33.Qi ST, Wang ZB, Ouyang YC, Zhang QH, Hu MW, Huang X, et al. Overexpression of setbeta, a protein localizing to centromeres, causes precocious separation of chromatids during the first meiosis of mouse oocytes. J Cell Sci. 2013;126(Pt 7):1595–1603. doi: 10.1242/jcs.116541. [DOI] [PubMed] [Google Scholar]
- 34.Okuwaki M, Nagata K. Template activating factor-i remodels the chromatin structure and stimulates transcription from the chromatin template. J Biol Chem. 1998;273(51):34511–34518. doi: 10.1074/jbc.273.51.34511. [DOI] [PubMed] [Google Scholar]
- 35.Gamble MJ, Erdjument-Bromage H, Tempst P, Freedman LP, Fisher RP. The histone chaperone taf-i/set/inhat is required for transcription in vitro of chromatin templates. Mol Cell Biol. 2005;25(2):797–807. doi: 10.1128/MCB.25.2.797-807.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Gamble MJ, Fisher RP. Set and parp1 remove dek from chromatin to permit access by the transcription machinery. Nat Struct Mol Biol. 2007;14(6):548–555. doi: 10.1038/nsmb1248. [DOI] [PubMed] [Google Scholar]
- 37.Mansouri S, Wang S, Frappier L. A role for the nucleosome assembly proteins taf-ibeta and nap1 in the activation of bzlf1 expression and Epstein–Barr virus reactivation. PLoS One. 2013;8(5):e63802. doi: 10.1371/journal.pone.0063802. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Seo SB, McNamara P, Heo S, Turner A, Lane WS, Chakravarti D. Regulation of histone acetylation and transcription by inhat, a human cellular complex containing the set oncoprotein. Cell. 2001;104(1):119–130. doi: 10.1016/S0092-8674(01)00196-9. [DOI] [PubMed] [Google Scholar]
- 39.Loven MA, Muster N, Yates JR, Nardulli AM. A novel estrogen receptor alpha-associated protein, template-activating factor ibeta, inhibits acetylation and transactivation. Mol Endocrinol. 2003;17(1):67–78. doi: 10.1210/me.2002-0280. [DOI] [PubMed] [Google Scholar]
- 40.Schneider R, Bannister AJ, Weise C, Kouzarides T. Direct binding of inhat to H3 tails disrupted by modifications. J Biol Chem. 2004;279(23):23859–23862. doi: 10.1074/jbc.C400151200. [DOI] [PubMed] [Google Scholar]
- 41.Macfarlan T, Parker JB, Nagata K, Chakravarti D. Thanatos-associated protein 7 associates with template activating factor-ibeta and inhibits histone acetylation to repress transcription. Mol Endocrinol. 2006;20(2):335–347. doi: 10.1210/me.2005-0248. [DOI] [PubMed] [Google Scholar]
- 42.Ichijo T, Chrousos GP, Kino T. Activated glucocorticoid receptor interacts with the inhat component SET/TAF-1beta and releases it from a glucocorticoid-responsive gene promoter, relieving repression: implications for the pathogenesis of glucocorticoid resistance in acute undifferentiated leukemia with set-can translocation. Mol Cell Endocrinol. 2008;283(1–2):19–31. doi: 10.1016/j.mce.2007.10.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Kepert JF, Mazurkiewicz J, Heuvelman GL, Toth KF, Rippe K. Nap1 modulates binding of linker histone H1 to chromatin and induces an extended chromatin fiber conformation. J Biol Chem. 2005;280(40):34063–34072. doi: 10.1074/jbc.M507322200. [DOI] [PubMed] [Google Scholar]
- 44.Shintomi K, Iwabuchi M, Saeki H, Ura K, Kishimoto T, Ohsumi K. Nucleosome assembly protein-1 is a linker histone chaperone in xenopus eggs. Proc Natl Acad Sci USA. 2005;102(23):8210–8215. doi: 10.1073/pnas.0500822102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Kato K, Okuwaki M, Nagata K. Role of template activating factor-I as a chaperone in linker histone dynamics. J Cell Sci. 2011;124(Pt 19):3254–3265. doi: 10.1242/jcs.083139. [DOI] [PubMed] [Google Scholar]
- 46.Georges SA, Kraus WL, Luger K, Nyborg JK, Laybourn PJ. p300-mediated tax transactivation from recombinant chromatin: histone tail deletion mimics coactivator function. Mol Cell Biol. 2002;22(1):127–137. doi: 10.1128/MCB.22.1.127-137.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Georges SA, Giebler HA, Cole PA, Luger K, Laybourn PJ, Nyborg JK. Tax recruitment of CBP/p300, via the KIX domain, reveals a potent requirement for acetyltransferase activity that is chromatin dependent and histone tail independent. Mol Cell Biol. 2003;23(10):3392–3404. doi: 10.1128/MCB.23.10.3392-3404.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Geiger TR, Sharma N, Kim YM, Nyborg JK. The human T-cell leukemia virus type 1 Tax protein confers CBP/p300 recruitment and transcriptional activation properties to phosphorylated CREB. Mol Cell Biol. 2008;28(4):1383–1392. doi: 10.1128/MCB.01657-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Kim YM, Ramirez JA, Mick JE, Giebler HA, Yan JP, Nyborg JK. Molecular characterization of the tax-containing HTLV-1 enhancer complex reveals a prominent role for CREB phosphorylation in Tax transactivation. J Biol Chem. 2007;282(26):18750–18757. doi: 10.1074/jbc.M700391200. [DOI] [PubMed] [Google Scholar]
- 50.Van Orden K, Nyborg JK. Insight into the tumor suppressor function of CBP through the viral oncoprotein Tax. Gene Expr. 2000;9(1–2):29–36. doi: 10.3727/000000001783992678. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Laybourn PJ, Kadonaga JT. Role of nucleosomal cores and histone H1 in regulation of transcription by RNA polymerase II. Science. 1991;254(5029):238–245. doi: 10.1126/science.1718039. [DOI] [PubMed] [Google Scholar]
- 52.Croston GE, Laybourn PJ, Paranjape SM, Kadonaga JT. Mechanism of transcriptional antirepression by gal4-vp16. Genes Dev. 1992;6(12A):2270–2281. doi: 10.1101/gad.6.12a.2270. [DOI] [PubMed] [Google Scholar]
- 53.Croston GE, Kerrigan LA, Lira LM, Marshak DR, Kadonaga JT. Sequence-specific antirepression of histone H1-mediated inhibition of basal RNA polymerase II transcription. Science. 1991;251(4994):643–649. doi: 10.1126/science.1899487. [DOI] [PubMed] [Google Scholar]
- 54.Schwarz PM, Hansen JC. Formation and stability of higher order chromatin structures. Contributions of the histone octamer. J Biol Chem. 1994;269(23):16284–16289. [PubMed] [Google Scholar]
- 55.Marheineke K, Krude T. Nucleosome assembly activity and intracellular localization of human caf-1 changes during the cell division cycle. J Biol Chem. 1998;273(24):15279–15286. doi: 10.1074/jbc.273.24.15279. [DOI] [PubMed] [Google Scholar]
- 56.Dong A, Liu Z, Zhu Y, Yu F, Li Z, Cao K, et al. Interacting proteins and differences in nuclear transport reveal specific functions for the NAP1 family proteins in plants. Plant Physiol. 2005;138(3):1446–1456. doi: 10.1104/pp.105.060509. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Nagata K, Saito S, Okuwaki M, Kawase H, Furuya A, Kusano A, et al. Cellular localization and expression of template-activating factor I in different cell types. Exp Cell Res. 1998;240(2):274–281. doi: 10.1006/excr.1997.3930. [DOI] [PubMed] [Google Scholar]
- 58.Leung JW, Leitch A, Wood JL, Shaw-Smith C, Metcalfe K, Bicknell LS, et al. Set nuclear oncogene associates with microcephalin/mcph1 and regulates chromosome condensation. J Biol Chem. 2011;286(24):21393–21400. doi: 10.1074/jbc.M110.208793. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Kadota S, Nagata K. Silencing of ifn-stimulated gene transcription is regulated by histone h1 and its chaperone TAF-I. Nucleic Acids Res. 2014 doi: 10.1093/nar/gku485. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Kato K, Miyaji-Yamaguchi M, Okuwaki M, Nagata K. Histone acetylation-independent transcription stimulation by a histone chaperone. Nucleic Acids Res. 2007;35(3):705–715. doi: 10.1093/nar/gkl1077. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Karetsou Z, Martic G, Sflomos G, Papamarcaki T. The histone chaperone set/taf-ibeta interacts functionally with the CREB-binding protein. Biochem Biophys Res Commun. 2005;335(2):322–327. doi: 10.1016/j.bbrc.2005.06.210. [DOI] [PubMed] [Google Scholar]
- 62.Shikama N, Chan HM, Krstic-Demonacos M, Smith L, Lee CW, Cairns W, et al. Functional interaction between nucleosome assembly proteins and p300/CREB-binding protein family coactivators. Mol Cell Biol. 2000;20(23):8933–8943. doi: 10.1128/MCB.20.23.8933-8943.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Okada M, Jeang KT. Differential requirements for activation of integrated and transiently transfected human T-cell leukemia virus type 1 long terminal repeat. J Virol. 2002;76(24):12564–12573. doi: 10.1128/JVI.76.24.12564-12573.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Lemasson I, Polakowski NJ, Laybourn PJ, Nyborg JK. Tax-dependent displacement of nucleosomes during transcriptional activation of human T-cell leukemia virus type 1. J Biol Chem. 2006;281(19):13075–13082. doi: 10.1074/jbc.M512193200. [DOI] [PubMed] [Google Scholar]
- 65.Hebbar PB, Archer TK. Altered histone h1 stoichiometry and an absence of nucleosome positioning on transfected DNA. J Biol Chem. 2008;283(8):4595–4601. doi: 10.1074/jbc.M709121200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Keck KM, Pemberton LF. Histone chaperones link histone nuclear import and chromatin assembly. Biochim Biophys Acta. 2013;1819(3–4):277–289. [PubMed] [Google Scholar]
- 67.Sandaltzopoulos R, Blank T, Becker PB. Transcriptional repression by nucleosomes but not H1 in reconstituted preblastoderm drosophila chromatin. EMBO J. 1994;13(2):373–379. doi: 10.1002/j.1460-2075.1994.tb06271.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Telese F, Bruni P, Donizetti A, Gianni D, D’Ambrosio C, Scaloni A, et al. Transcription regulation by the adaptor protein fe65 and the nucleosome assembly factor set. EMBO Rep. 2005;6(1):77–82. doi: 10.1038/sj.embor.7400309. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Matsumoto K, Nagata K, Miyaji-Yamaguchi M, Kikuchi A, Tsujimoto M. Sperm chromatin decondensation by template activating factor i through direct interaction with basic proteins. Mol Cell Biol. 1999;19(10):6940–6952. doi: 10.1128/mcb.19.10.6940. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Karetsou Z, Martic G, Tavoulari S, Christoforidis S, Wilm M, Gruss C, et al. Prothymosin alpha associates with the oncoprotein set and is involved in chromatin decondensation. FEBS Lett. 2004;577(3):496–500. doi: 10.1016/j.febslet.2004.09.091. [DOI] [PubMed] [Google Scholar]
- 71.Kibler KV, Jeang KT. CREB/ATF-dependent repression of cyclin a by human T-cell leukemia virus type 1 Tax protein. J Virol. 2001;75(5):2161–2173. doi: 10.1128/JVI.75.5.2161-2173.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Rousset R, Desbois C, Bantignies F, Jalinot P. Effects on NF-kappa B/p105 processing of the interaction between the HTLV-1 transactivator tax and the proteasome. Nature. 1996;381(6580):328–331. doi: 10.1038/381328a0. [DOI] [PubMed] [Google Scholar]
- 73.Rehtanz M, Schmidt HM, Warthorst U, Steger G. Direct interaction between nucleosome assembly protein 1 and the papillomavirus e2 proteins involved in activation of transcription. Mol Cell Biol. 2004;24(5):2153–2168. doi: 10.1128/MCB.24.5.2153-2168.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Wang S, Frappier L. Nucleosome assembly proteins bind to Epstein–Barr virus nuclear antigen 1 and affect its functions in DNA replication and transcriptional activation. J Virol. 2009;83(22):11704–11714. doi: 10.1128/JVI.00931-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Zhao LJ, Giam CZ. Interaction of the human t-cell lymphotrophic virus type 1 (HTLV-1) transcriptional activator Tax with cellular factors that bind specifically to the 21-base-pair repeats in the HTLV-1 enhancer. Proc Natl Acad Sci USA. 1991;88(24):11445–11449. doi: 10.1073/pnas.88.24.11445. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Lopez DI, Mick JE, Nyborg JK. Purification of creb to apparent homogeneity: removal of truncation products and contaminating nucleic acid. Protein Expr Purif. 2007;55(2):406–418. doi: 10.1016/j.pep.2007.06.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Franklin AA, Kubik MF, Uittenbogaard MN, Brauweiler A, Utaisincharoen P, Matthews MA, et al. Transactivation by the human t-cell leukemia virus tax protein is mediated through enhanced binding of activating transcription factor-2 (ATF-2) and cAMP element-binding protein (CREB) J Biol Chem. 1993;268(28):21225–21231. [PubMed] [Google Scholar]
- 78.Fyodorov DV, Kadonaga JT. Chromatin assembly in vitro with purified recombinant ACF and NAP-1. Methods Enzymol. 2003;371:499–515. doi: 10.1016/S0076-6879(03)71037-4. [DOI] [PubMed] [Google Scholar]
- 79.Lu X, Hansen JC. Identification of specific functional subdomains within the linker histone H10 C-terminal domain. J Biol Chem. 2004;279(10):8701–8707. doi: 10.1074/jbc.M311348200. [DOI] [PubMed] [Google Scholar]
- 80.Hieb AR, D’Arcy S, Kramer MA, White AE, Luger K. Fluorescence strategies for high-throughput quantification of protein interactions. Nucleic Acids Res. 2012;40(5):e33. doi: 10.1093/nar/gkr1045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Luger K, Rechsteiner TJ, Flaus AJ, Waye MM, Richmond TJ. Characterization of nucleosome core particles containing histone proteins made in bacteria. J Mol Biol. 1997;272(3):301–311. doi: 10.1006/jmbi.1997.1235. [DOI] [PubMed] [Google Scholar]
- 82.Dynan WS. DNAse I footprinting as an assay for mammalian gene regulatory proteins. Genet Eng. 1987;9:75–87. [Google Scholar]
- 83.Lal A, Navarro F, Maher CA, Maliszewski LE, Yan N, O’Day E, et al. Mir-24 inhibits cell proliferation by targeting e2f2, myc, and other cell-cycle genes via binding to “seedless” 3′utr microrna recognition elements. Mol Cell. 2009;35(5):610–625. doi: 10.1016/j.molcel.2009.08.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Carruthers LM, Bednar J, Woodcock CL, Hansen JC. Linker histones stabilize the intrinsic salt-dependent folding of nucleosomal arrays: mechanistic ramifications for higher-order chromatin folding. Biochemistry. 1998;37(42):14776–14787. doi: 10.1021/bi981684e. [DOI] [PubMed] [Google Scholar]
- 85.Luger K, Rechsteiner TJ, Richmond TJ. Preparation of nucleosome core particle from recombinant histones. Methods Enzymol. 1999;304:3–19. doi: 10.1016/S0076-6879(99)04003-3. [DOI] [PubMed] [Google Scholar]
- 86.Scoggin KE, Ulloa A, Nyborg JK. The oncoprotein Tax binds the SRC-1-interacting domain of CBP/p300 to mediate transcriptional activation. Mol Cell Biol. 2001;21(16):5520–5530. doi: 10.1128/MCB.21.16.5520-5530.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Giebler HA, Loring JE, van Orden K, Colgin MA, Garrus JE, Escudero KW, et al. Anchoring of CREB binding protein to the human T-cell leukemia virus type 1 promoter: a molecular mechanism of tax transactivation. Mol Cell Biol. 1997;17(9):5156–5164. doi: 10.1128/mcb.17.9.5156. [DOI] [PMC free article] [PubMed] [Google Scholar]