

Abstract
In response to rapid changes in their environment, bacteria control a number of processes, including motility, cell division, biofilm formation, and virulence. Research presented in January 2015 at the biennial Bacterial Locomotion and Signal Transduction (BLAST) meeting in Tucson, AZ, illustrates the elegant complexity of the nanoarrays, nanomachines, and networks of interacting proteins that mediate such processes. Studies employing an array of biophysical, genetic, cell biology, and mathematical methods are providing an increasingly detailed understanding of the mechanisms of these systems within well-studied bacteria. Furthermore, comparisons of these processes in diverse bacterial species are providing insight into novel regulatory and functional mechanisms. This review summarizes research presented at the BLAST meeting on these fundamental mechanisms and diverse adaptations, including findings of importance for applications involving bacteria of medical or agricultural relevance.
INTRODUCTION
For 25 years, the vibrant Bacterial Locomotion and Signal Transduction (BLAST) biennial meetings have characterized the elegant complexity of the mechanisms bacteria use to sense and respond to their environment. A hallmark of this behavior is bacterial chemotaxis, a specialized signal transduction system used to couple appropriate locomotion to continuous sensing of local environmental changes (Fig. 1). Modern chemotaxis research was started some 50 years ago by Julius Adler with the development of assays that would allow him to characterize bacterial motility (1) (for a review of the history of bacterial chemotaxis, see reference 2). Much of the early work focused on model organisms such as Escherichia coli, Salmonella enterica, or Bacillus subtilis. Approaches involved the visualization of bacterial movement, or “tracking” (3), and the identification of the vast number of proteins involved in the motor and flagella (4, 5). Proteins involved in signal transduction in chemotaxis are part of a large family of histidine aspartic acid phosphorelay proteins (aka “two-component signaling” [TCS] systems) that regulate behaviors other than motility, including gene transcription and global regulator molecules such as cyclic di-GMP (see Fig. 3). Indeed, much of the pioneering work in the mechanisms of signal transduction was first presented in meetings of BLAST researchers. Thus, from the initial motility tracking assays, BLAST meetings have witnessed the expansion of the focus on locomotion and signal transduction to a wide array of processes investigated with highly sophisticated methodologies. Furthermore, comparisons of these processes in diverse bacterial species are providing insight into novel regulatory and functional mechanisms, as well as key information for applications involving bacteria of medical or agricultural importance.
FIG 1.

Overview of bacterial chemotaxis systems. Polar arrays of chemoreceptors (gray), CheW (cyan), and CheA (blue) activate autophosphorylation of CheA. The phosphoryl group is transferred to the CheY response regulator, which binds to the switch on the flagellar motor to cause clockwise rotation, disruption of the bundle of flagella, and tumbling of the cell (bottom inset). Binding of attractant ligands from the environment (orange triangles) to chemoreceptors inhibits phosphorylation of CheA and clockwise rotation of the motor, thus suppressing tumbling, so the cell swims longer toward favorable environments (top inset). Note that the switch of one motor to clockwise rotation is sufficient for the bacterium to tumble. Adaptation to ongoing stimuli is mediated by covalent modification of the receptors (see Fig. 2): specific Glu/Gln residues in the receptor cytoplasmic domain are methylated by CheR and demethylated/deamidated by CheB.
FIG 3.

Intracellular signaling variations. (A) Control of cyclic di-GMP (cdG) levels in a cell. Diguanylate cyclase (DGC) proteins synthesize cdG (red hexagons) from two GTP molecules, while phosphodiesterase (PDE) proteins facilitate its degradation to GMP. (B to E) cdG targets discussed in BLAST XIII; see the text for details. (B) A novel CheY-like protein in C. crescentus directly binds to cdG, causing its localization to FliM. (C) FlgZ bound to cdG sequesters MotC (green ovals), preventing its incorporation into the stator in P. aeruginosa. (D) cdG directly binds to the type IV pilus extension ATPase, MshE, to stimulate pilus production in V. cholerae. (E) cdG may cause the DGC YfiN to alter its conformation, inducing its interaction with FtsZ (maroon circles) in order to prevent Z-ring constriction. (F) A paradigm two-component signal (TCS) transduction system. Signal transmission occurs via His-Asp phosphorelay between conserved His kinase (HK; green) and receiver (REC; blue) modules connected by polypeptide linker regions (gray lines). In response to activation by the sensor domain, the HK module autophosphorylates on an invariant His (H) residue after hydrolysis of ATP by the catalytic (CA) domain. The phosphoryl group (P) is donated to an invariant Asp (D) in the REC module that activates the associated DNA binding effector domain to modulate gene transcription. Effector domains may instead encode enzymatic domains, such as diguanylate cyclases. (G to I) Variations of the paradigm TCS system discussed in BLAST XIII; see the text for details. (G) The S. enterica SsrA/B system in which the phosphorylated form of the RR SsrB (SsrB-P) regulates Salmonella pathogenicity island 2 (SPI-2). Unphosphorylated SsrB antagonizes the H-NS nucleoid protein to derepress csgD. (H) The multistep CckA-ChpT-CtrA-CpdR His-Asp phosphorelay system of Brucella abortus, in which a phosphoryl group is transmitted via a REC module in a hybrid histidine kinase (Hy-HK) to a histidine phosphotransferase (HPT) protein. The phosphoryl group is then transferred to either of two response regulators to mediate outputs. CA*, degenerate CA domain. (I) The S. aureus BraSR system in which the HK BraS is activated upon direct interaction with the BraE permease (pink rectangle) when its ligand bacitracin (star) is bound. BraR-P induces expression of a second permease system (yellow rectangle and circle) which then exports the bacitracin. CM, cytoplasmic membrane; PG, peptidoglycan; OM, outer membrane.
Here we summarize the research presented at BLAST XIII to illustrate the exciting blend of an array of methods revealing both fundamental mechanisms and diverse adaptations. To frame the discussion, Fig. 1 summarizes the process of chemotaxis from the binding of chemoattractant to the switching of the flagellar motor. The process is illustrated for E. coli, but many variations are now recognized. For example, instead of (or in addition to) the rotary motor shown in Fig. 1 that propels the flagella of numerous bacteria, including E. coli, S. enterica, and B. subtilis (6), other bacteria employ diverse gliding motility machineries. Pseudomonas aeruginosa and Myxococcus xanthus have motility machineries consisting of retractable type IV pili (7) (referred to as twitching motility and social motility, respectively). M. xanthus also contains a second “adventurous” gliding motility machinery involving focal adhesion (FA) complexes, and Flavobacterium johnsoniae (8) contains a completely distinct gliding motility motor. Detailed studies of the rotary motor have revealed diversity among species as well (see Fig. 4). As one example, E. coli uses a ring of some 10 to 12 MotA and MotB stator proteins to drive the flagellar motor (9), while Shewanella oneidensis can switch between a proton-driven motor that uses MotA/MotB and a sodium-driven motor that uses PomA/PomB, to adapt to changes in the environment (10). Likewise, the signal transduction pathways that lead from the actual chemical stimulus, usually outside the cell or in the periplasm, to a change in motility behavior differ among species.
FIG 4.

Common features and diverse elements of flagellar motors from different bacterial species. (A) A slide from a 3D reconstruction of a cell tip of the Lyme disease spirochete (84) reveals multiple motors embedded in the cytoplasmic membrane (CM). Noticeably, the spirochetal flagella reside within the periplasmic space between the CM and the outer membrane (OM). Flagellar motors that rotate external flagella of E. coli, Campylobacter jejuni, and Hylemonella gracilis (A to C) (85) and flagellar motors that rotate periplasmic flagella of Treponema primitia (86), B. burgdorferi (84), and Leptospira interrogans (87) (E to G) are shown along with the schematic model for each flagellar motor in the bottom panel (88). The rotor is colored in yellow or orange, the putative stator is in blue, and the export apparatus is in purple. (Reproduced from references 84 and 88 with permission of the publishers.)
BLAST XIII also highlighted the ways in which the field of bacterial motility, chemotaxis, and signal transduction extends beyond these processes and beyond microbiology. For instance, the field has long included analysis of the physics of motility and chemoreception (11), the complexity of the signal transduction systems has attracted bioinformatic analyses (12, 13), and the resulting insights into modularity of these systems have inspired protein engineering (14). It has become clear that motility and its related signal transduction pathways are never isolated processes in the bacterial cell but are part of an integrated network of regulatory and metabolic pathways. As specific examples, motility is coupled to cell division (15, 16) and two-component signaling contributes to virulence in numerous organisms (17–19). Furthermore, complex regulatory pathways are also involved in the transition from motile to sessile states, and the interplay of motility and biofilm formation is complex (20).
This review highlights the research presented in talks at BLAST XIII held in Tucson, AZ, from 19 to 22 January 2015. We had a total of 133 attendees from 15 countries across the world, and a total of 52 oral presentations and 59 poster presentations. In keeping with BLAST tradition, this meeting provided an excellent forum for young scientists to present and discuss their research, with 29% of the talks given by postdocs and 32% given by graduate students, as well as one undergraduate giving a poster that won one of the two poster awards. This review article highlights the research presented in most (80%) of the oral presentations, grouped into the following sequence of topics: (i) sensing fundamentals, (ii) sensing variations and applications, (iii) intracellular signaling variations, (iv) motor fundamentals, (v) motor variations, and (vi) niche adaptations. The variations and adaptations provide examples of the importance of this research area with respect to harnessing and controlling bacteria for agricultural and medical applications.
SENSING FUNDAMENTALS
Many of the fundamentals of signal transduction by two-component signaling systems have been deduced through investigations of the well-studied chemotaxis system of E. coli. A wide range of bacteria use this system to sense and respond to attractant and repellent molecules in their environment. Sensing begins with binding of environmental signals (such as aspartate, serine, maltose, and ribose) to a remarkable membrane-bound array of chemoreceptors that form a hexagonal lattice (Fig. 2B) with the cytoplasmic proteins CheA and CheW (21–24). It is widely accepted that attractant binding to the periplasmic domain of the chemoreceptor causes an approximately 2-Å piston movement of transmembrane helix 2 (TM2) toward the cytoplasm (25, 26), but it is not yet clear how the signal is transmitted 200 Å through the cytoplasmic domain to control the kinase activity of CheA bound to the membrane-distal cytoplasmic tip of the receptor.
FIG 2.
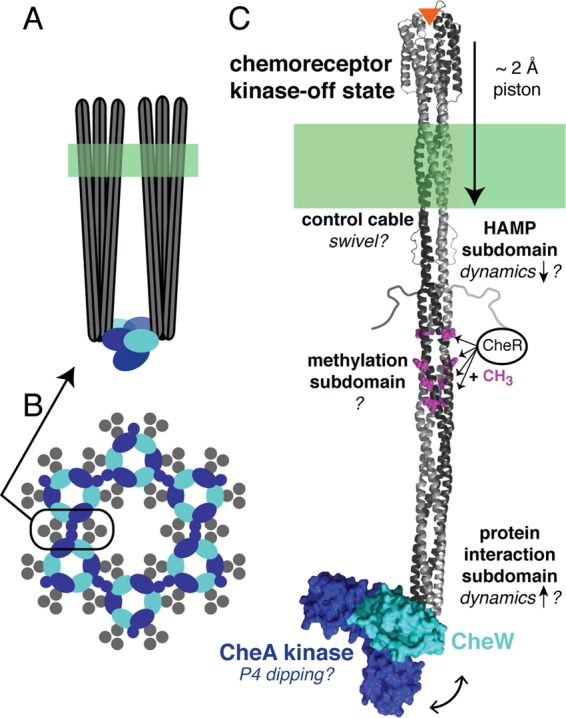
Hexagonal array of chemoreceptors. A cartoon model of an array is presented, showing a side view of two receptor trimers of dimers with CheA and CheW bound at the membrane-distal tip of the receptor (A) and a view at the membrane-distal tip of the hexagonal lattice, which can extend ∼200 nm (B). (C) Structural model of one receptor dimer in complex with CheW and CheA (P4 and P5 domains). It is widely accepted that binding of an attractant ligand (orange triangle) causes an approximately 2-Å piston movement of transmembrane helix 2, but it is not yet clear what changes occur in the cytoplasm to turn off the kinase. Evidence was presented for various proposed changes in the receptor (indicated by question marks and discussed in the text), including a swivel of the control cable, a decrease in the dynamics of the HAMP subdomain, possible changes in dynamics of the methylation subdomain (presently not well characterized), and an increase in the dynamics of the protein interaction subdomain, as well as a dipping (double-headed arrow) of the P4 domain of CheA.
Protein domains can propagate information via changes in structure or changes in dynamics. Recently, various groups have proposed that connections between successive subdomains of the cytoplasmic domain cause inverse stabilization of these subdomains in the kinase-on and kinase-off signaling states, and experimental methods, including mutagenesis, disulfide cross-linking, hydrogen exchange, and electron paramagnetic resonance (EPR), are being used to develop and test these proposals (27–31). Peter Ames from Sandy Parkinson's group (University of Utah) presented results of extensive mutagenesis of the 5-residue “control cable” that connects the cytoplasmic end of transmembrane helix 2 (TM2) to the HAMP subdomain in the serine receptor (32), which he interpreted in the context of a model in which the helix packing interfaces of TM2 and HAMP are out of register and in which this structural conflict destabilizes the HAMP subdomain to create the kinase-on state. He reported that shortened control cables lock the receptor in the kinase-on state and suggested that this is due to a further destabilization of the HAMP subdomain. Ames proposed that the ligand-induced 2-Å piston movement causes a swivel of the control cable at the critical initial two residues, to relieve the TM2/HAMP structural conflict and stabilize HAMP to yield the kinase-off state (Fig. 2C).
Speakers from two different research groups presented site-directed spin labeling EPR studies providing new evidence that dynamics in different chemoreceptor subdomains change with the signaling state. Both groups measured EPR spectra of spin labels incorporated at unique Cys residues introduced throughout the cytoplasmic domain of the E. coli aspartate receptor, using different constructs, conditions, and methods. Nicholas Bartelli from Jerry Hazelbauer's group (University of Missouri—Columbia) spin-labeled the intact receptor and then reconstituted individual receptor dimers into ∼10-nm-diameter lipid bilayer “nanodiscs.” In such bilayers, isolated dimers couple ligand binding and adaptational modification to receptor conformation as demonstrated by ligand-enhanced methylation rates, but multiple receptor dimers in the same nanodisc (presumably forming trimers of dimers) are needed to activate and control the kinase (33, 34). Dipanjan Samanta from Brian Crane's group (Cornell University) spin-labeled two soluble cytoplasmic-domain variants that incorporated different HAMP domains to mimic the kinase-on and kinase-off states. Results reported by both groups for the methylation subdomain did not show a clear signaling-related pattern: dynamics did not change significantly upon ligand addition (Bartelli) or in the kinase-on versus kinase-off variants (Samanta), but both groups observed increased dynamics for the 4E unmethylated variant relative to the 4Q methylated-mimic variant. This may reflect the fact that full coupling of ligand and methylation control of receptor function is observed only for receptors in complex with CheA and CheW (35). Based on both EPR line shapes and pulsed EPR measurements of the distribution of distances between spin labels, Samanta concluded that the protein interaction subdomain (cytoplasmic tip) of the receptor has increased dynamics in the kinase-off state (Fig. 2C) (36), irrespective of whether inactivation is achieved by HAMP state or demethylation. This finding is consistent with a recent hydrogen exchange study of receptor constructs in functional complexes with CheA and CheW (27).
A molecular dynamics (MD) study presented by Keith Cassidy from Klaus Schulten's group (University of Illinois at Champaign-Urbana) revealed unexpected large-scale motions in CheA within arrays. Building on previous models of the array's extended organization (21, 23), Cassidy utilized computational modeling and MD simulations to generate an atomically resolved model of the array unit cell from existing crystal structures and new electron cryotomography data obtained by Peijun Zhang's group (University of Pittsburgh) using arrays of cytoplasmic receptor fragments, CheA, and CheW bound to lipid monolayers. All-atom MD simulations of the 1.2 million-atom array model revealed a periodic “dipping” of the CheA P4 domain (Fig. 2C) below the CheA/W baseplate and allowed the prediction of key residues affecting the conformational dynamics of CheA. Data from other laboratories have suggested signaling-related changes in the mobility of CheA linkers and domains (37, 38); it remains to be seen how such changes are affected by the receptor and how they modulate kinase activity.
Results of research on chemoreceptors from other organisms revealed both the universality of the features observed in E. coli and new features characteristic of more-complex chemotaxis systems found in other bacteria. Davi Ortega from Grant Jensen's group (California Institute of Technology) discussed the evolution of the chemotaxis system in archaea. Ortega presented evidence that the system was acquired by lateral gene transfer from bacteria to archaea and presented electron cryotomography images of the membrane-bound and cytoplasmic receptor arrays of archaea, both of which have the same hexagonal array architecture as that observed in bacteria (39). Tino Krell (Estación Experimental del Zaidín, Spain) presented results on the chemotaxis system of the human pathogen P. aeruginosa, a model for bacteria with greater diversity and complexity of chemotaxis proteins than are found in the single chemosensory pathway of E. coli. Krell and coworkers used isothermal titration calorimetry to investigate binding of this organism's four different CheR methyltransferases to three C-terminal pentapeptides of its chemoreceptors. Since only CheR2 bound the McpB pentapeptide and methylated the McpB receptor, Krell suggested that the role of pentapeptide binding is to mediate receptor-specific methylation and adaptation in separate chemosensory systems responding to different stimuli. Furthermore, Krell's group identified a common feature in the sequences of CheR methyltransferases that bind to receptor pentapeptides, which may facilitate prediction of receptor/CheR interaction partners in other bacteria (40).
SENSING VARIATIONS AND APPLICATIONS
Chemotaxis research is also focused on responses by organisms of medical or agricultural importance. For example, several groups study the chemoreceptors of Helicobacter pylori, the causative agent of some human cancers and ulcers. Emily Sweeney, of the groups of James Remington and Karen Guillemin (University of Oregon), presented the crystal structure of the TlpA ligand binding domain, a tandem-PAS structure, and suggested that it may serve as a heme-binding receptor to sense levels of O2, NO, or CO and enable H. pylori to perform chemotaxis toward a site of damage in the stomach. Findings from the Manson group revealed important insights into the mechanism by which norepinephrine mediates chemotaxis responses of pathogenic E. coli. Mike Manson (Texas A&M University) reported (i) that norepinephrine induces expression of two enzymes that can convert it to 3,4-dihydroxymandelic acid (DHMA), (ii) that nonpathogenic E. coli bacteria that express these enzymes exhibit an attractant response to norepinephrine, and (iii) that this response is mediated by the serine receptor binding DHMA. Interestingly, standard chemotaxis assays might not have detected the response, because it peaks between 5 and 50 nM DHMA and then disappears at higher concentrations. The microflow assay used in this study was likely critical for detecting the DHMA response and may enable discovery of other responses to clarify the role of chemotaxis in pathogenesis by enteric bacteria (41).
Novel methods for identifying ligands and receptors mediating chemotaxis are also playing a role in studies of agriculturally important bacterial systems. Monica Gerth (University of Otago) described a high-throughput fluorescence thermal shift assay developed in her group to identify the ligand specificity of the 43 chemoreceptors found in Pseudomonas syringae pv. actinidiae, a pathogen of kiwifruit. The strategy involved expression and purification of each receptor ligand-binding domain for screening with this assay against a commercially available collection of over 900 ligands. Hits were confirmed with isothermal titration calorimetry to measure binding and with capillary assays to measure chemotaxis. The successful identification of a number of ligands and the lack of false positives suggested that this assay provides an efficient means to characterize complex chemotaxis networks present in many bacteria (42). Benjamin Webb from Birgit Scharf's group (Virginia Tech) won the award for Outstanding Talk by a Graduate Student. He described a modified agarose capillary assay method for automated data acquisition enabling simultaneous time course measurements for up to 10 strains. Webb demonstrated the power of this assay in a study of the role of chemotaxis in symbiosis between a soil bacterium and alfalfa (43). Capillary assays on Sinorhizobium meliloti and various knockout strains were used to identify the receptors involved in chemotaxis to the seed exudate of the host plant Medicago sativa.
INTRACELLULAR SIGNALING VARIATIONS
The BLAST XIII meeting also introduced exciting variations in intracellular signaling mechanisms. One theme which emerged from this meeting involved identification of new targets of the second messenger cyclic di-GMP (cdG). cdG is generally involved in regulation of the transition between the motile and sessile states in bacteria (44), often by regulation of the motility apparatus, but the mechanisms are not always understood (Fig. 3A). While the enzymes involved in the production (diguanylate cyclases [DGC]) and degradation (phosphodiesterases [PDE]) of cdG are readily identifiable by the presence of conserved enzymatic domains, less is known about proteins involved in directly detecting and responding to cdG.
To identify targets for cdG in Caulobacter crescentus, Urs Jenal (University of Basel) and his group used the cdG-specific capture compound mass spectrometry technique (45). Jenal presented the identification of a novel set of proteins containing a CheY-like receiver domain followed by an arginine-rich domain (Fig. 3B). Biochemical and mutant analyses indicated that the Arg-rich domain directly binds cdG and is necessary for localization of these proteins to FliM in the polar flagellar apparatus of swimming cells. Interestingly, phosphorylation of the CheY-like domain seems to play no role in localization. Functional analyses of these novel CheY-like proteins suggested that their role is complex and that they likely impact not only chemotaxis but also other aspects of the flagellar motor. A different mechanism by which cdG affects the flagellar motor in P. aeruginosa was presented by Amy Baker from George O'Toole's group (Geisel School of Medicine at Dartmouth) (Fig. 3C). Her data suggest that cdG may control swarming in this organism by altering the ratio of two different stator complexes (MotAB and MotCD) incorporated into the motor of its single polar flagellum. Higher levels of MotCD promote swarming, while higher levels of MotAB prevent swarming (20). Insight as to how cdG may control this behavior came from an observation that FlgZ (containing a cdG-responsive PilZ domain) interacts with MotC. This observation suggests a model in which FlgZ-cdG interacts with membrane pools of MotC, which prevents MotC incorporation into the stator. As a consequence, the ratio of MotAB to MotCD would be increased, repressing swarming when cdG levels are high. Yet another novel mechanism by which cdG influences motility was presented by Christopher Jones from Fitnat Yildiz's group (University of California, Santa Cruz) (Fig. 3D). Jones showed that cdG influences motility in Vibrio cholerae via regulation of the assembly of its type IV pilus, MshA. The mshA locus was identified as a suppressor of the nonmotile phenotype in the PDE cdgJ mutant background. Jones and his colleagues then used high-speed surface imaging to determine that cdgJ mutants have an enhanced surface interaction phenotype and that the Msh pili increased V. cholerae surface interactions and adhesion. The researchers also demonstrated that surface pilus production correlated with cdG levels and that cdG directly binds to the extension ATPase MshE. Together, the data suggest a model in which cdG levels stimulate MshE-mediated pilus assembly, leading to an increase in surface attachment and a decrease in motility. In a final exciting development, identification of a cdG target which does not involve motility was presented by Hyo Kyung Kim from Rasika Harshey's group (University of Texas at Austin) (Fig. 3E). Kim showed that cdG regulates E. coli and Salmonella cell division in response to cell envelope stress. Starting from the observation that in some cells the DGC YfiN localizes to the midcell in a pattern characteristic of the FtsZ Z-ring, Kim then demonstrated that YfiN is mislocalized under restrictive temperature in ftsZts but not ftsAts background strains. Furthermore, Kim observed that YfiN interacts directly with FtsZ by bacterial two-hybrid assay. YfiN midcell localization is facilitated under conditions of osmotic stress or outer membrane permeabilization. Kim presented a model in which cell envelope stress stimulates a conformational change in YfiN, causing it to interact with FtsZ to prevent constriction of the Z-ring, thereby blocking cell division.
A second theme that emerged from the meeting involved new roles for His-Asp phosphorelay proteins (a.k.a. two-component signal transduction proteins). In the paradigmatic system, a histidine kinase (HK) autophosphorylates on an invariant histidine residue in response to an external stimulus (Fig. 3F) (46). The cognate response regulator (RR) protein mediates transfer of the HK phosphoryl group to itself on an invariant aspartic acid. For the vast majority of the response regulators, phosphorylation modulates transcription of specific gene targets (47). BLAST XIII addressed activity of response regulators that can be uncoupled from the cognate kinase. Stuti Desai from Linda Kenney's group (National University of Singapore) demonstrated that there are distinct roles for the unphosphorylated and phosphorylated forms of the S. enterica RR, SsrB, which could coordinate the virulence state and the carrier state of this organism (Fig. 3G). The SsrA and SsrB (SsrA/B) HK/RR pair is necessary for transcriptional activation of genes required for vacuolar survival (virulence state). SsrB displaces the nucleoid protein H-NS on certain genes (48). Desai demonstrated that it is the nonphosphorylatable mutant of SsrB that can activate biofilm formation (carrier state) via transcriptional activation of the csgD gene, which encodes a master regulator of biofilm formation. Desai used genetic experiments to demonstrate that SsrB antagonizes H-NS in vivo and used atomic force microscopy to demonstrate that SsrB alters the condensation of H-NS on a DNA fragment containing the csgD promoter. McKenzie Lehman from Ken Bayles' group (University of Nebraska Medical Center) presented data on the LytSR two-component system, which plays a role in Staphylococcus aureus biofilm production. LytSR regulates the lrgAB operon, which is important for release of extracellular DNA. Lehman used a transcriptional reporter to show that localization of lrg expression to biofilm towers depends on the phosphorylation state of the LytR RR, which is controlled by both the kinase activity and the phosphatase activity of the HK LytS in response to membrane potential (18). In planktonic cells, however, LytR also accepts phosphoryl groups from acetyl phosphate pools (49), which influences lrg expression. The significance of this effect in planktonic cells is not yet known but hints at possible novel roles for LytR in planktonic cells.
Insight into the mechanics of signal transmission by more-complex His-Asp phosphorelay systems was also covered in BLAST XIII. One complex system is the multistep phosphorelay signaling pathway used by many alphaproteobacteria to regulate the complex cell cycles in this group of bacteria (50) (Fig. 3H). This pathway consists of a HK (CckA), a histidine phosphotransferase protein (ChpT), and two RR (CtrA and CpdR). Jonathan Willett from Sean Crosson's group (University of Chicago) examined the role of this signaling pathway in the pathogen Brucella abortus. Using phosphoprofiling (51), Willett demonstrated that CtrA and CpdR are the only RRs phosphorylated by CckA/ChpT and that these proteins are necessary for appropriate polar cell division exhibited by B. abortus. Importantly, Willett obtained crystal structures of ChpT (1.6 Å) and a ChpT-CtrA complex (2.7 Å). These structures revealed that ChpT is an obligate dimer with a histidine kinase fold; it includes the catalytic domain (CA) responsible for ATP hydrolysis in autophosphorylating HKs. Consistent with ChpT's role as a phosphotransferase protein, the ATP binding residues in the CA domain are not conserved and the protein does not bind ATP. The ChpT-CtrA complex revealed asymmetry in the ChpT dimer, with the phosphoaccepting histidine residue aligned with the CtrA phosphoaccepting aspartic acid residue in one of the dimer subunits. In the second dimer subunit, however, the phosphoaccepting histidine residue was flipped outward and not competent for phosphotransfer to the respective CtrA. Molecular dynamics simulations suggest that the ChpT phosphoaccepting histidine has distinct structural and dynamic characteristics across the dimer in the ChpT and ChpT-CtrA structures.
A second multicomponent His-Asp phosphorelay system addressed in BLAST XIII involved HK/RR pairs that are activated by interaction with distinct sensor proteins. Tarek Msadek (Pasteur Institute) reported on a novel mechanism by which intramembrane sensor HKs function with transport proteins to detect their respective ligands (Fig. 3I). The S. aureus BraSR HK-RR pair functions with two ABC transporter complexes (BraDE and VraDE) to mediate resistance to the antibiotics nisin and bacitracin. These antibiotics elicit BraSR-dependent upregulation of the two ABC transporters. The BraS HK sensing domain contains two transmembrane helices connected by a three-residue periplasmic loop. Msadek demonstrated that the periplasmic loop is not necessary for BraS sensing of the antibiotics. Rather, the antibiotics are bound by the BraE permease, and that information is relayed to BraS through interaction between the BraS and BraE transmembrane segments. Interestingly, the BraDE ABC transporter seems to be a dedicated sensor, whereas antibiotic resistance is provided exclusively by VraDE. In the closely related cationic antimicrobial peptide (CAMP) resistance system, the intramembrane sensing kinase GraS is likewise activated by CAMP binding via the VraG permease (52).
MOTOR FUNDAMENTALS
The bacterial flagellar motor is arguably one of the best-studied molecular machines in any living organism (Fig. 4). Despite over 50 years of research, many fundamental aspects of these motors, including protein export and assembly, rotation switching, and torque generation, remain to be understood at the molecular level. BLAST XIII featured extensive discussions on the flagellar ATPase complex (FliH and FliI) and its role in flagellar assembly (Fig. 4 and 5), on CheY-P binding and switching, and on stator-rotor interactions.
FIG 5.

Overview of the flagellar motor architecture and its central interactions. (A) An intact T. primitia motor map (gray) is overlaid with a Shigella injectisome map (cyan) (89). They provide a framework for docking atomic models of the export gate complex (enlarged in panel B) and the ATPase complex (panel C) (90, 91). Together with recent stator-rotor interaction studies (D) (92) and a switch complex model (E) (93), the results presented in BLAST XIII provide new insights into structure-function relationships of this fascinating molecular machine. (Adapted from images published in references 89, 92, 93, and 94 with permission of the publishers.)
Tao Lin from Steven Norris's group (University of Texas Medical School at Houston) used the Lyme disease spirochete, Borrelia burgdorferi, as a model system to investigate the structure and function of the flagellar ATPase complex (Fig. 4 and 5). They found that inactivation of either fliH or fliI has profound effects on flagellar structure, morphology, motility, and cell division but does not completely block protein export and flagellar assembly (53). Electron cryotomography results indicated that disruption of either fliH or fliI resulted in the loss of the ATPase complex from the otherwise intact flagellar motor. Furthermore, fliH and fliI mutants administered by either needle injection or use of infected Ixodes scapularis ticks were nonvirulent in mice. Genetic complementation of fliH and fliI mutants in trans restored their wild-type morphology, motility, and flagellar motor structure. However, both full-length flagella and infectivity were not recovered in the complemented stains, supporting the idea of the critical roles of flagella in the infection cycle of this bacterium.
It is generally believed that both ATP hydrolysis and proton motive force (PMF) are required to energize different steps of flagellar protein export and assembly (54, 55). Marc Erhardt (Helmholtz Center for Infection Research) and his colleagues were able to form functional full-length flagella in the absence of the ATPase complex by mutations that increased the PMF and substrate levels (56). Additionally, they provided evidence that protein secretion in a catalytically inactive ATPase mutant of injectisomes was rescued by increased PMF. Their results support the idea of the dispensability of the ATPase activity in both flagella and injectisomes.
Another highly investigated structural component of the flagellar motor is the C-ring (Fig. 5), which is also known as the switch complex. This complex is located at the cytoplasmic base of the bacterial flagellum, and it is essential for flagellar assembly, torque generation, and rotation switching in response to chemotaxis signaling (57). However, the domain arrangements of three proteins (FliG, FliN, and FliM) within the switch complex remain uncertain (58–61). To understand the FliG conformation in solution and its in vivo assembly, Matthew Baker from Lawrence Lee's group (Victor Chang Cardiac Research Institute) used small-angle X-ray scattering to study purified FliG. His results suggested that FliG exists in solution predominantly as a compact conformation. However, targeted disulfide cross-linking studies indicated that FliG undergoes large conformational changes in vivo and self-assembles with adjacent molecules through intermolecular interactions between the middle and C-terminal domains of FliG.
A sophisticated signaling system allows B. burgdorferi to sense chemical stimuli. Ultimately, this system regulates the direction of flagellar rotation and the switch between two motility phenotypes: “run” and “flex,” a nontranslational mode in spirochete (62). The nonchemotactic mutants ΔcheA2 and ΔcheY3 constantly run, while the ΔcheX mutant constantly flexes (62). Xiaowei Zhao from Jun Liu's group (University of Texas Medical School at Houston) used electron cryotomography to study B. burgdorferi mutants involved in running and flexing behavior. Two distinctive motor conformations are directly associated with the two swimming behaviors in ΔcheX and ΔcheY3 mutants. This study provided direct structural evidence that CheY-P interacts with the switch complex protein FliM, triggering a large conformational change in the switch complex, which might be essential for flagellar switching in this spirochete.
The amino-terminal domain of FliM is important for CheY-P binding and phosphorylation-dependent switching in E. coli (63). Afanzar Oshri from Michael Eisenbach's group (Weizmann Institute of Science) reported on the unique properties of switching and chemotactic responsiveness in the absence of the amino-terminal domain of FliM. These were explained in the context of CheY acetylation and by a model in which CheY levels alter the number of exchangeable FliM and thus the responsiveness of the motor.
Finally, for the stator-rotor interaction (Fig. 5), several conserved charged residues in the rotor and stator are known to play critical roles (64). These charged residues have been extensively characterized in the default rotation (counterclockwise [CCW]). However, their specific role in the context of clockwise (CW) rotation remains to be determined. Yasuhiro Onoue from Michio Homma's group (Nagoya University) systematically constructed charge-neutralizing mutations in a chimeric rotor or stator expressed in E. coli. A tethered-cell assay was used to measure rotational speeds in both directions for each cell. They found that conserved charged residues in the stator contributed equally to rotations in the two directions. In contrast, some of the conserved charged residues in the rotor appeared to impact rotation differently in the two directions.
Studies of CheY in vivo are crucial to understand its specific roles in bacterial chemotaxis and motility. One common approach to study the localization and dynamics of proteins involves fusing the protein of interest to fluorescent proteins. Diana di Paolo from Richard Berry's group (University of Oxford) employed a novel technique for protein internalization in live bacteria based on electroporation. The organic fluorophores used to label the chemotaxis proteins of interest were smaller, brighter, and more photostable than the fluorescent protein counterparts. Di Paolo was able to exploit this technique for better tracking of a single CheY protein between the sensory cluster at the cell pole and the flagellar motors. This novel approach can be extended to track other functionally important proteins during flagellar rotation and switching.
Tracking individual cells in three dimensions (3D) is essential for understanding bacterial motility behavior. A typical 3D tracker requires sophisticated engineering and is a low-throughput assay that tracks one cell at a time. Katja M. Taute from Tom Shimizu's group (AMOLF Institute, Amsterdam, The Netherlands), who won the award for Outstanding Talk by a Postdoc, demonstrated a simple, label-free 3D bacterial tracking technique that extracts z-axis information from the diffraction patterns observed by examination of out-of-focus objects in phase-contrast microscopy. Impressively, dozens of bacteria could be followed simultaneously with micron-scale spatial resolution and video rate temporal resolution, over a z range of ∼200 μm and lateral range of ∼300 μm by 350 μm. This simple and elegant technique can be used to study a wide range of enteric, marine, and soil bacteria.
Most uniflagellated bacteria switch the direction of motor rotation between CW and CCW to alternate between forward and backward swimming. The intervals between motor switches determine a cell's swimming trajectory, and altering these times can lead to directed motility. The signaling protein CheY-P regulates the level of chemotactic response by affecting the intervals of the CW motor rotation. Michael Morse from Jay Tang's group (Brown University) measured the distribution of motor rotation intervals and found the clear presence of a peak switching time for both CW and CCW rotation in C. crescentus. The first-passage time theory for a biased random walker fits well with their data, suggesting that the motor switching behavior of C. crescentus is governed by a stochastic process. Junhua Yuan (University of Science and Technology) monitored motor rotation by attaching nanogold particles to the flagellar hooks of E. coli cells lacking filaments. Their data support the model that motor switching near zero load occurs at random times and can be treated as a two-state Poisson process (65).
Ismaël Duchesne from Simon Rainville's group (Université Laval) showed how the medium can influence the motility behavior of bacteria. Duchesne examined motility in liquid crystals. Liquid crystals are considered anisotropic, meaning that the physical properties are not identical in all directions. Instead of the commonly known “random walk,” bacteria in an anisotropic medium run back and forth in one dimension. In the transition zone between isotropic and anisotropic media, bacteria become sticky as their propulsive force and the viscosity of the medium increase. The authors concluded that (i) the swimming behavior of bacteria is drastically different in anisotropic environments, (ii) “artificial” liquid crystals such as solutions of disodium chromoglycate (cromolyn) can be used to probe that phenomenon, and (iii) motile microorganisms could be used as a probe to determine the physical properties of a medium.
Yuhai Tu (Thomas J. Watson Research Center) asked a fundamental question: how efficient are bacterial flagellar motors? It was originally thought that the efficiency of the flagellar motor at high load should be close to 100%. However, recent experimental results suggested that the maximum energy efficiency is 71% (66). To better understand the mechanisms, a mathematical model for the rotary motor was developed. The surprising result is that there is maximal efficiency at intermediate loads which depends on the PMF. This new model can explain the lower-than-expected energy efficiency observed in recent experiments.
MOTOR VARIATIONS
In addition to the thorough dissection of flagellar motor structure and function, BLAST meetings also highlight the functionality of nonflagellar motors, such as those used for movement over solid surfaces (gliding motility) by M. xanthus, F. johnsoniae, and Cellulophaga algicola.
M. xanthus gliding motors contain PMF-harnessing AglQRS (Mot/Tol/Exb family) motor proteins together with numerous cell envelope proteins arranged in focal adhesion (FA) complexes. FA complexes move through the length of the cell but remain stationary with respect to the substratum as the cells are propelled forward (67). Beiyan Nan from David Zusman's group (University of California, Berkeley) reported that the AglS motor protein directly interacts with the bacterial actin homolog MreB. Using single-particle-tracking photoactivated localization, Nan observed that MreB-mCherry (mCh) consisted of a mixture of polymerized and unpolymerized molecules in moving cells and that a subpopulation of unpolymerized MreB-mCh moved helically. The MreB-mCh movements were dependent on moving motors, while motor movements were dependent on the presence of MreB filaments. Nan proposed that in M. xanthus, MreB provides a scaffold for gliding motors, which in turn drive the movement of MreB filaments (68).
In F. johnsoniae, gliding involves the PMF-dependent rapid movement of the motility adhesion, SprB, along the cell surface. SprB is secreted by the type IX secretion system (69), which appears to be intertwined with the motility system. Yongtao Zhu from Mark McBride's group (University of Wisconsin—Milwaukee) reported on efforts to distinguish the motor components from the secretion system components by comparative analysis of F. johnsoniae and C. algicola motility mutants. Abhishek Shrivastava from Howard Berg's group (Harvard University) used F. johnsoniae cells tethered by SprB antibodies to demonstrate that SprB is linked to a rotary motor which operates at constant speed rather than constant torque (70).
PHENOTYPIC DIVERSITY OF MOTILITY FOR NICHE ADAPTATION
A number of presentations at the BLAST XIII meeting described the phenotypic diversity of motility behavior in adaptation to specific niches and environments. Three presentations focused on enteric bacteria, including S. enterica, a pathogen that colonizes the human intestine and causes foodborne illness. Motile bacteria are more virulent due to coregulation of motility with genes of pathogenicity island I (71). However, flagellin stimulates the host immune response. As a compromise, subpopulations of motile bacteria can coexist beside subpopulations of nonmotile bacteria. The selective advantage of this heterogeneity is a division of labor, where motile cells are pathogenic and nonmotile cells serve as a reservoir to continue the infection (72). The molecular mechanisms that underlie this phenomenon feed into the flagellar transcriptional hierarchy. In S. enterica and E. coli, this hierarchy begins with flhDC, encoding the FlhD4 FlhC2 (FlhDC) transcriptional activator (73). A second-level gene is fliA, encoding the FliA flagellum-specific sigma factor (74). FliA increases expression of level III genes, including fliT, which encodes an anti-FlhDC factor (75), and fliZ, which encodes a posttranslational activator of FlhDC (76). A third gene discussed in this context is ydiV, encoding the YdiV anti-FlhDC factor (77). The impact of FliZ and YdiV on the bistability of flagellar gene expression in S. enterica was recently described by different groups (78, 79).
Phil Aldridge (Newcastle University) reported a regulatory circuit in S. enterica that controls flagella in response to the nutritional situation and is quite different from the E. coli regulatory circuit. While E. coli responds to high-nutrient conditions with a shutdown of motility, S. enterica increases motility. The Aldridge study discovered that the number of flagella per bacterium correlated with the growth rate such that rapidly growing bacteria produced more flagella than slowly growing ones. The conclusion from this group was that the flagellar systems of E. coli and S. enterica share a large degree of genetic similarity but process nutritional signals differently.
The second presentation on this topic was by Santosh Koirala from Chris Rao's group (University of Illinois at Urbana-Champaign), who explored the effect of nutrients on motility heterogeneity in S. enterica. Bacteria were grown in minimal medium supplemented with glucose and various amounts of yeast extract. Gene expression was monitored by fluorescent transcriptional fusions from the respective promoters. The response to an increase in FlhDC concentration exhibited hysteresis, which is indicative of a truly bistable system. Since the effect of hysteresis was lost in mutants of either ydiV or fliZ, the authors concluded that the competitive interaction of YdiV and FliZ was responsible for the bistability of flagellar gene expression.
The final presentation on this topic was by Mary Stewart from Brad Cookson's group (University of Washington). She presented a single-cell study, where flow cytometry and fluorescence reporter fusions were used to determine expression of the flagellin gene, fliC. A distinction was made between a fliC-off (nonmotile) subpopulation and a fliC-on (motile) subpopulation. The presence of YdiV was sufficient to maintain the fliC-off subpopulation. A functional FliZ was required to activate FlhDC and maintain the fliC-on subpopulation at late time points during growth. Since bistability could still be observed in mutants lacking fliZ, it was concluded that the entire YdiV-FliZ regulatory circuit was not necessary to achieve bistability.
A single-cell approach was also used by Y. S. Dufour (Yale University) to study phenotypic diversity in E. coli swimming behavior. In a recent publication (80), diversity was predicted to be beneficial when cells are swimming in diverse environments. Dufour used epifluorescence microscopy to count the tagged CheR and CheB proteins in individual cells immobilized in a hydrogel after observing their swimming behavior. The tumble bias of single cells is partially explained by a nonlinear ratio of CheR and CheB numbers. This is an interesting new technique that will facilitate the investigation of the molecular origins of phenotypic diversity and chemotactic performance in controlled environments.
An intriguing example of motor adaptation was presented by Shannon Au (The Chinese University of Hong Kong) for H. pylori, a bacterium that colonizes a rather extreme ecological niche, the human stomach. While E. coli uses phospho-CheY to bind to the switch protein FliM and change the direction of flagellar rotation, H. pylori seems to adopt an additional regulation mechanism mediated through the binding of spermidine synthase SpeE to FliM and switch. SpeE was identified as a binding partner of FliM in a glutathione S-transferase (GST) pulldown assay, followed by mass spectrometry. The crystal structure of the SpeE-FliM complex revealed the interacting surface, where one SpeE dimer interacted with FliM. No large structural change in FliM was observed upon SpeE binding. However, binding of SpeE suppressed the interaction of FliG with FliM. This is a good example of how different bacterial species use different mechanisms to control chemotaxis systems.
Continuing with motor adaptation as a mechanism to survive diverse environments, the presentation by Susanne Brenzinger from Kai Thormann's group (Justus Liebig University Giessen) focused on S. oneidensis, a bacterium that is used in many bioindustrial applications. In addition to the sodium-dependent PomAB stator, S. oneidensis also possesses the proton-dependent MotAB stator present in many bacteria (e.g., E. coli) (81). By fluorescence recovery after photobleaching (FRAP) and continuous bleaching analysis, the exchange rate and number of stator components in the motor were determined. PomAB/MotAB ratios and PomAB turnover respond to NaCl availability. Spontaneous mutations in the plug domain of MotB led to stator turnover that was about three times higher.
Soil is another environment that is very specific and highly variable and requires adaptation. John Kirby (University of Iowa) and his research group studied the predatory behavior of M. xanthus toward the soil bacterium B. subtilis, which is capable of forming fruiting bodies in response to starvation. Previously published research established that M. xanthus consumed a variety of laboratory strains of B. subtilis, while an ancestral strain escaped predation by using production of the secondary metabolite bacillaene as a defense (82). Kirby showed that the resistant B. subtilis strain produced multicellular broccoli-shaped megastructures that formed on or near the fruiting bodies of M. xanthus. These structures were up to 500 μm in diameter and rose far above the agar surface. The authors hypothesized that the so-called “militarized zone” between the two bacterial cultures may contain multiple yet-to-be identified secondary metabolites that protect B. subtilis from predation (83).
Among all the environments bacteria can find themselves in, a human, animal, or plant host is certainly a challenging one. Our final summary illustrates the importance of motility and chemotaxis in Lyme disease, a zoonotic disease which can be transmitted from a variety of animal hosts to humans by means of ticks and is caused by B. burgdorferi. Mark Wooten (University of Toledo College of Medicine) used a mouse model to study the effect of CheY3 on dissemination of B. burgdorferi throughout the host tissue. Using fluorescently labeled bacterial strains, transgenic mice with fluorescent immune cells, and fluorescent confocal intravital microscopy, the researchers found that wild-type B. burgdorferi can travel through the host tissue at an amazing speed, over 40 times faster than any immune cell of the skin tissue. This enables the bacteria to spread from the skin inoculation site throughout the tissue, where they can persist for two or more years. The nonchemotactic strain ΔcheY3 moved through the tissue more slowly, was unable to reverse direction, and was consequently cleared from the host by 72 to 96 h postinfection.
Taking the data together, signaling and motility systems are an important contributor to a bacterium's ability to adapt to various environments. Phenotypic diversity and niche adaptation of these systems are critical to interactions between bacteria and their physical environment, between different bacterial species, and between bacteria and host organisms. An understanding of such interactions is critical to our coexistence with beneficial bacteria and to control of harmful bacteria.
CONCLUSION
BLAST XIII continued in its tradition of characterizing canonical motility and sensory systems with unprecedented detail using an impressive combination of novel and traditional biophysical, genetic, cell biology, and mathematical methods. By highlighting alternate motility machines and emerging model systems, BLAST meetings always provide a unique view of common biological themes and their exploitation by bacteria.
ACKNOWLEDGMENTS
We thank BLAST XIII Chair Karen Ottemann and Vice Chair Alan Wolfe, as well as all organizers, presenters, and attendees of the meeting. Special recognition goes to Conference Coordinators Tarra Bollinger and Peggy O'Neill.
Research on signaling, flagella, and motility in our laboratories is supported by grants from the National Institutes of Health (R01-GM085288 to L.K.T. and R01AI087946 and R01GM107629 to J.L.) and from the Welch Foundation (AU-1714 to J.L.) and by the North Dakota Agricultural Experiment Station (B.M.P.) and Wayne State University (P.I.H.).
Biographies

Birgit M. Prüß received her Ph.D. from Ruhr-Universitaet Bochum in Germany. She was a postdoctoral fellow with Alan J. Wolfe at Loyola University Chicago and then with Philip Matsumura at the University of Illinois at Chicago. Since 2004, she has been a faculty member at North Dakota State University, currently at the associate professor level. Her research interest is global gene regulation in E. coli biofilms, with a particular emphasis on using this information to influence biofilm amounts by deliberately changing the bacteria's environment. She teaches bacterial physiology.

Jun Liu, associate professor, got his Ph.D. training in physics at the Institute of Physics of the Chinese Academy of Sciences. He made a career transition from a physicist to a biophysicist by working with Ken Taylor at Florida State University as a postdoctoral fellow. After further training with Sriram Subramanian as a staff scientist at the National Institutes of Health, he started his laboratory as an assistant professor at the UT Medical School at Houston. His laboratory is interested in 3D structure/function of molecular machines in bacterial pathogens. In collaboration with Steven Norris and William Margolin, he developed two model systems (a Lyme disease spirochete system and a bacterial minicell system) to study bacterial motility, signal transduction, protein secretion, and DNA translocation in native cellular environments. In particular, he has been dedicated to developing a high-throughput cryo-electron tomography system to effectively visualize molecular machines that orchestrate those fundamental processes in cells.

Penelope I. Higgs did her Ph.D. thesis with Kathleen Postle in molecular biosciences at Washington State University. She then did a postdoctoral fellowship in the laboratory of David Zusman at the University of California, Berkeley, where she became interested in the highly complex signaling mechanisms that control the Myxococcus xanthus developmental program. She continued this work as a group leader at the Max Planck Institute for Terrestrial Microbiology in Marburg, Germany, where she also expanded her interests to mechanisms of spore differentiation and cell fate segregation. Her group recently moved to Wayne State University, where she is currently an assistant professor.

Lynmarie K. Thompson earned a B.S. at California Institute of Technology and a Ph.D. at Yale University, working on photosystem II with Gary Brudvig. She then learned solid-state nuclear magnetic resonance (NMR) methods for membrane protein studies as a postdoctoral fellow with Robert Griffin at Massachusetts Institute of Technology. She joined the faculty at University of Massachusetts Amherst in 1991, where she is currently a professor of chemistry and in the graduate program in molecular and cellular biology, and she also directs the Chemistry-Biology Interface NIH Training Program. Her research program investigates structures and mechanisms of membrane proteins, with an emphasis on biophysical studies of the mechanism of transmembrane signaling by bacterial chemotaxis receptors.
REFERENCES
- 1.Adler J, Templeton B. 1967. The effect of environmental conditions on the motility of Escherichia coli. J Gen Microbiol 46:175–184. doi: 10.1099/00221287-46-2-175. [DOI] [PubMed] [Google Scholar]
- 2.Adler J. 2011. My life with nature. Annu Rev Biochem 80:42–70. doi: 10.1146/annurev-biochem-121609-100316. [DOI] [PubMed] [Google Scholar]
- 3.Berg HC. 1971. How to track bacteria. Rev Sci Instrum 42:868–871. doi: 10.1063/1.1685246. [DOI] [PubMed] [Google Scholar]
- 4.Homma M, Ohnishi K, Iino T, Macnab RM. 1987. Identification of flagellar hook and basal body gene products (FlaFV, FlaFVI, FlaFVII and FlaFVIII) in Salmonella typhimurium. J Bacteriol 169:3617–3624. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Dimmitt K, Simon MI. 1971. Purification and partial characterization of Bacillus subtilis flagellar hooks. J Bacteriol 108:282–286. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Berg HC. 2003. The rotary motor of bacterial flagella. Annu Rev Biochem 72:19–54. doi: 10.1146/annurev.biochem.72.121801.161737. [DOI] [PubMed] [Google Scholar]
- 7.Burrows LL. 2012. Pseudomonas aeruginosa twitching motility: type IV pili in action. Annu Rev Microbiol 66:493–520. doi: 10.1146/annurev-micro-092611-150055. [DOI] [PubMed] [Google Scholar]
- 8.Nan B, McBride MJ, Chen J, Zusman DR, Oster G. 2014. Bacteria that glide with helical tracks. Curr Biol 24:R169–R173. doi: 10.1016/j.cub.2013.12.034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Blair DF, Berg HC. 1988. Restoration of torque in defective flagellar motors. Science 242:1678–1681. doi: 10.1126/science.2849208. [DOI] [PubMed] [Google Scholar]
- 10.Paulick A, Koerdt A, Lassak J, Huntley S, Wilms I, Narberhaus F, Thormann KM. 2009. Two different stator systems drive a single polar flagellum in Shewanella oneidensis MR-1. Mol Microbiol 71:836–850. doi: 10.1111/j.1365-2958.2008.06570.x. [DOI] [PubMed] [Google Scholar]
- 11.Berg HC, Purcell EM. 1977. Physics of chemoreception. Biophys J 20:193–219. doi: 10.1016/S0006-3495(77)85544-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Szurmant H, Hoch JA. 2010. Interaction fidelity in two-component signaling. Curr Opin Microbiol 13:190–197. doi: 10.1016/j.mib.2010.01.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Wuichet K, Alexander RP, Zhulin IB. 2007. Comparative genomic and protein sequence analyses of a complex system controlling bacterial chemotaxis. Methods Enzymol 422:1–31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Skerker JM, Perchuk BS, Siryaporn A, Lubin EA, Ashenberg O, Goulian M, Laub MT. 2008. Rewiring the specificity of two-component signal transduction systems. Cell 133:1043–1054. doi: 10.1016/j.cell.2008.04.040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Davis NJ, Cohen Y, Sanselicio S, Fumeaux C, Ozaki S, Luciano J, Guerrero-Ferreira RC, Wright ER, Jenal U, Viollier PH. 2013. De- and repolarization mechanism of flagellar morphogenesis during a bacterial cell cycle. Genes Dev 27:2049–2062. doi: 10.1101/gad.222679.113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Prüss BM, Matsumura P. 1996. A regulator of the flagellar regulon of Escherichia coli, flhD, also affects cell division. J Bacteriol 178:668–674. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Kim HS, Caswell CC, Foreman R, Roop RM II, Crosson S. 2013. The Brucella abortus general stress response system regulates chronic mammalian infection and is controlled by phosphorylation and proteolysis. J Biol Chem 288:13906–13916. doi: 10.1074/jbc.M113.459305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Lehman MK, Bose JL, Sharma-Kuinkel BK, Moormeier DE, Endres JL, Sadykov MR, Biswas I, Bayles KW. 2015. Identification of the amino acids essential for LytSR-mediated signal transduction in Staphylococcus aureus and their roles in biofilm-specific gene expression. Mol Microbiol 95:723–737. doi: 10.1111/mmi.12902. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Walthers D, Carroll RK, Navarre WW, Libby SJ, Fang FC, Kenney LJ. 2007. The response regulator SsrB activates expression of diverse Salmonella pathogenicity island 2 promoters and counters silencing by the nucleoid-associated protein H-NS. Mol Microbiol 65:477–493. doi: 10.1111/j.1365-2958.2007.05800.x. [DOI] [PubMed] [Google Scholar]
- 20.Kuchma SL, Delalez NJ, Filkins LM, Snavely EA, Armitage JP, O'Toole GA. 2015. Cyclic Di-GMP-mediated repression of swarming motility by Pseudomonas aeruginosa PA14 requires the MotAB stator. J Bacteriol 197:420–430. doi: 10.1128/JB.02130-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Briegel A, Li X, Bilwes AM, Hughes KT, Jensen GJ, Crane BR. 2012. Bacterial chemoreceptor arrays are hexagonally packed trimers of receptor dimers networked by rings of kinase and coupling proteins. Proc Natl Acad Sci U S A 109:3766–3771. doi: 10.1073/pnas.1115719109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Briegel A, Ortega DR, Tocheva EI, Wuichet K, Li Z, Chen S, Muller A, Iancu CV, Murphy GE, Dobro MJ, Zhulin IB, Jensen GJ. 2009. Universal architecture of bacterial chemoreceptor arrays. Proc Natl Acad Sci U S A 106:17181–17186. doi: 10.1073/pnas.0905181106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Liu J, Hu B, Morado DR, Jani S, Manson MD, Margolin W. 2012. Molecular architecture of chemoreceptor arrays revealed by cryoelectron tomography of Escherichia coli minicells. Proc Natl Acad Sci U S A 109:E1481–E1488. doi: 10.1073/pnas.1200781109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Zhang P, Khursigara CM, Hartnell LM, Subramaniam S. 2007. Direct visualization of Escherichia coli chemotaxis receptor arrays using cryo-electron microscopy. Proc Natl Acad Sci U S A 104:3777–3781. doi: 10.1073/pnas.0610106104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Chervitz SA, Falke JJ. 1996. Molecular mechanism of transmembrane signaling by the aspartate receptor: a model. Proc Natl Acad Sci U S A 93:2545–2550. doi: 10.1073/pnas.93.6.2545. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Falke JJ, Hazelbauer GL. 2001. Transmembrane signaling in bacterial chemoreceptors. Trends Biochem Sci 26:257–265. doi: 10.1016/S0968-0004(00)01770-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Koshy SS, Li X, Eyles SJ, Weis RM, Thompson LK. 2014. Hydrogen exchange differences between chemoreceptor signaling complexes localize to functionally important subdomains. Biochemistry 53:7755–7764. doi: 10.1021/bi500657v. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Orban-Glaß I, Voskoboynikova N, Busch KB, Klose D, Rickert C, Mosslehy W, Roder F, Wilkens V, Piehler J, Engelhard M, Steinhoff HJ, Klare JP. 2015. Clustering and dynamics of phototransducer signaling domains revealed by site-directed spin labeling electron paramagnetic resonance on SRII/HtrII in membranes and nanodiscs. Biochemistry 54:349–362. doi: 10.1021/bi501160q. [DOI] [PubMed] [Google Scholar]
- 29.Starrett DJ, Falke JJ. 2005. Adaptation mechanism of the aspartate receptor: electrostatics of the adaptation subdomain play a key role in modulating kinase activity. Biochemistry 44:1550–1560. doi: 10.1021/bi048089z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Swain KE, Gonzalez MA, Falke JJ. 2009. Engineered socket study of signaling through a four-helix bundle: evidence for a yin-yang mechanism in the kinase control module of the aspartate receptor. Biochemistry 48:9266–9277. doi: 10.1021/bi901020d. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Zhou Q, Ames P, Parkinson JS. 2009. Mutational analyses of HAMP helices suggest a dynamic bundle model of input-output signalling in chemoreceptors. Mol Microbiol 73:801–814. doi: 10.1111/j.1365-2958.2009.06819.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Kitanovic S, Ames P, Parkinson JS. 2011. Mutational analysis of the control cable that mediates transmembrane signaling in the Escherichia coli serine chemoreceptor. J Bacteriol 193:5062–5072. doi: 10.1128/JB.05683-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Li M, Khursigara CM, Subramaniam S, Hazelbauer GL. 2011. Chemotaxis kinase CheA is activated by three neighbouring chemoreceptor dimers as effectively as by receptor clusters. Mol Microbiol 79:677–685. doi: 10.1111/j.1365-2958.2010.07478.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Amin DN, Hazelbauer GL. 2010. The chemoreceptor dimer is the unit of conformational coupling and transmembrane signaling. J Bacteriol 192:1193–1200. doi: 10.1128/JB.01391-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Li G, Weis RM. 2000. Covalent modification regulates ligand binding to receptor complexes in the chemosensory system of Escherichia coli. Cell 100:357–365. doi: 10.1016/S0092-8674(00)80671-6. [DOI] [PubMed] [Google Scholar]
- 36.Samanta D, Borbat PP, Dzikovski B, Freed JH, Crane BR. 2015. Bacterial chemoreceptor dynamics correlate with activity state and are coupled over long distances. Proc Natl Acad Sci U S A 112:2455–2460. doi: 10.1073/pnas.1414155112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Briegel A, Ames P, Gumbart JC, Oikonomou CM, Parkinson JS, Jensen GJ. 2013. The mobility of two kinase domains in the Escherichia coli chemoreceptor array varies with signalling state. Mol Microbiol 89:831–841. doi: 10.1111/mmi.12309. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Wang X, Vallurupalli P, Vu A, Lee K, Sun S, Bai WJ, Wu C, Zhou H, Shea JE, Kay LE, Dahlquist FW. 2014. The linker between the dimerization and catalytic domains of the CheA histidine kinase propagates changes in structure and dynamics that are important for enzymatic activity. Biochemistry 53:855–861. doi: 10.1021/bi4012379. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Briegel A, Ortega DR, Huang AN, Oikonomou CM, Gunsalus RP, Jensen GJ. 2015. Structural conservation of chemotaxis machinery across Archaea and Bacteria. Environ Microbiol Rep 7:414–419. doi: 10.1111/1758-2229.12265. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.García-Fontana C, Corral Lugo A, Krell T. 2014. Specificity of the CheR2 methyltransferase in Pseudomonas aeruginosa is directed by a C-terminal pentapeptide in the McpB chemoreceptor. Sci Signal 7:ra34. doi: 10.1126/scisignal.2004849. [DOI] [PubMed] [Google Scholar]
- 41.Pasupuleti S, Sule N, Cohn WB, MacKenzie DS, Jayaraman A, Manson MD. 2014. Chemotaxis of Escherichia coli to norepinephrine (NE) requires conversion of NE to 3,4-dihydroxymandelic acid. J Bacteriol 196:3992–4000. doi: 10.1128/JB.02065-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.McKellar JL, Minnell JJ, Gerth ML. 2015. A high-throughput screen for ligand binding reveals the specificities of three amino acid chemoreceptors from Pseudomonas syringae pv. actinidiae. Mol Microbiol 96:694–707. doi: 10.1111/mmi.12964. [DOI] [PubMed] [Google Scholar]
- 43.Webb BA, Hildreth S, Helm RF, Scharf BE. 2014. Sinorhizobium meliloti chemoreceptor McpU mediates chemotaxis toward host plant exudates through direct proline sensing. Appl Environ Microbiol 80:3404–3415. doi: 10.1128/AEM.00115-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Römling U, Galperin MY, Gomelsky M. 2013. Cyclic di-GMP: the first 25 years of a universal bacterial second messenger. Microbiol Mol Biol Rev 77:1–52. doi: 10.1128/MMBR.00043-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Nesper J, Reinders A, Glatter T, Schmidt A, Jenal U. 2012. A novel capture compound for the identification and analysis of cyclic di-GMP binding proteins. J Proteomics 75:4874–4878. doi: 10.1016/j.jprot.2012.05.033. [DOI] [PubMed] [Google Scholar]
- 46.Salazar ME, Laub MT. 2015. Temporal and evolutionary dynamics of two-component signaling pathways. Curr Opin Microbiol 24:7–14. doi: 10.1016/j.mib.2014.12.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Galperin MY. 2006. Structural classification of bacterial response regulators: diversity of output domains and domain combinations. J Bacteriol 188:4169–4182. doi: 10.1128/JB.01887-05. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Walthers D, Li Y, Liu Y, Anand G, Yan J, Kenney LJ. 2011. Salmonella enterica response regulator SsrB relieves H-NS silencing by displacing H-NS bound in polymerization mode and directly activates transcription. J Biol Chem 286:1895–1902. doi: 10.1074/jbc.M110.164962. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Wolfe AJ. 2010. Physiologically relevant small phosphodonors link metabolism to signal transduction. Curr Opin Microbiol 13:204–209. doi: 10.1016/j.mib.2010.01.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Brilli M, Fondi M, Fani R, Mengoni A, Ferri L, Bazzicalupo M, Biondi EG. 2010. The diversity and evolution of cell cycle regulation in alpha-proteobacteria: a comparative genomic analysis. BMC Syst Biol 4:52. doi: 10.1186/1752-0509-4-52. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Skerker JM, Prasol MS, Perchuk BS, Biondi EG, Laub MT. 2005. Two-component signal transduction pathways regulating growth and cell cycle progression in a bacterium: a system-level analysis. PLoS Biol 3:e334. doi: 10.1371/journal.pbio.0030334. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Falord M, Karimova G, Hiron A, Msadek T. 2012. GraXSR proteins interact with the VraFG ABC transporter to form a five-component system required for cationic antimicrobial peptide sensing and resistance in Staphylococcus aureus. Antimicrob Agents Chemother 56:1047–1058. doi: 10.1128/AAC.05054-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Lin T, Gao L, Zhao X, Liu J, Norris SJ. 2015. Mutations in the Borrelia burgdorferi flagellar type III secretion system genes fliH and fliI profoundly affect spirochete flagellar assembly, morphology, motility, structure, and cell division. mBio 6:e00579-15. doi: 10.1128/mBio.00579-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Chevance FF, Hughes KT. 2008. Coordinating assembly of a bacterial macromolecular machine. Nat Rev Microbiol 6:455–465. doi: 10.1038/nrmicro1887. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Macnab RM. 2003. How bacteria assemble flagella. Annu Rev Microbiol 57:77–100. doi: 10.1146/annurev.micro.57.030502.090832. [DOI] [PubMed] [Google Scholar]
- 56.Erhardt M, Mertens ME, Fabiani FD, Hughes KT. 2014. ATPase-independent type-III protein secretion in Salmonella enterica. PLoS Genet 10:e1004800. doi: 10.1371/journal.pgen.1004800. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Sowa Y, Berry RM. 2008. Bacterial flagellar motor. Q Rev Biophys 41:103–132. [DOI] [PubMed] [Google Scholar]
- 58.Lee LK, Ginsburg MA, Crovace C, Donohoe M, Stock D. 2010. Structure of the torque ring of the flagellar motor and the molecular basis for rotational switching. Nature 466:996–1000. doi: 10.1038/nature09300. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Minamino T, Imada K, Kinoshita M, Nakamura S, Morimoto YV, Namba K. 2011. Structural insight into the rotational switching mechanism of the bacterial flagellar motor. PLoS Biol 9:e1000616. doi: 10.1371/journal.pbio.1000616. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Paul K, Gonzalez-Bonet G, Bilwes AM, Crane BR, Blair D. 2011. Architecture of the flagellar rotor. EMBO J 30:2962–2971. doi: 10.1038/emboj.2011.188. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Vartanian AS, Paz A, Fortgang EA, Abramson J, Dahlquist FW. 2012. Structure of flagellar motor proteins in complex allows for insights into motor structure and switching. J Biol Chem 287:35779–35783. doi: 10.1074/jbc.C112.378380. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Charon NW, Cockburn A, Li C, Liu J, Miller KA, Miller MR, Motaleb MA, Wolgemuth CW. 2012. The unique paradigm of spirochete motility and chemotaxis. Annu Rev Microbiol 66:349–370. doi: 10.1146/annurev-micro-092611-150145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Bren A, Eisenbach M. 1998. The N terminus of the flagellar switch protein, FliM, is the binding domain for the chemotactic response regulator, CheY. J Mol Biol 278:507–514. doi: 10.1006/jmbi.1998.1730. [DOI] [PubMed] [Google Scholar]
- 64.Zhou J, Lloyd SA, Blair DF. 1998. Electrostatic interactions between rotor and stator in the bacterial flagellar motor. Proc Natl Acad Sci U S A 95:6436–6441. doi: 10.1073/pnas.95.11.6436. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Wang F, Yuan J, Berg HC. 2014. Switching dynamics of the bacterial flagellar motor near zero load. Proc Natl Acad Sci U S A 111:15752–15755. doi: 10.1073/pnas.1418548111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Lo CJ, Sowa Y, Pilizota T, Berry RM. 2013. Mechanism and kinetics of a sodium-driven bacterial flagellar motor. Proc Natl Acad Sci U S A 110:E2544–E2551. doi: 10.1073/pnas.1301664110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Zhang Y, Ducret A, Shaevitz J, Mignot T. 2012. From individual cell motility to collective behaviors: insights from a prokaryote, Myxococcus xanthus. FEMS Microbiol Rev 36:149–164. doi: 10.1111/j.1574-6976.2011.00307.x. [DOI] [PubMed] [Google Scholar]
- 68.Nan B, Bandaria JN, Guo KY, Fan X, Moghtaderi A, Yildiz A, Zusman DR. 2015. The polarity of myxobacterial gliding is regulated by direct interactions between the gliding motors and the Ras homolog MglA. Proc Natl Acad Sci U S A 112:E186–E193. doi: 10.1073/pnas.1421073112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Shrivastava A, Johnston JJ, van Baaren JM, McBride MJ. 2013. Flavobacterium johnsoniae GldK, GldL, GldM, and SprA are required for secretion of the cell surface gliding motility adhesins SprB and RemA. J Bacteriol 195:3201–3212. doi: 10.1128/JB.00333-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Shrivastava A, Lele PP, Berg HC. 2015. A rotary motor drives flavobacterium gliding. Curr Biol 25:338–341. doi: 10.1016/j.cub.2014.11.045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Chubiz JE, Golubeva YA, Lin D, Miller LD, Slauch JM. 2010. FliZ regulates expression of the Salmonella pathogenicity island 1 invasion locus by controlling HilD protein activity in Salmonella enterica serovar Typhimurium. J Bacteriol 192:6261–6270. doi: 10.1128/JB.00635-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Stewart MK, Cookson BT. 2012. Non-genetic diversity shapes infectious capacity and host resistance. Trends Microbiol 20:461–466. doi: 10.1016/j.tim.2012.07.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Yanagihara S, Iyoda S, Ohnishi K, Iino T, Kutsukake K. 1999. Structure and transcriptional control of the flagellar master operon of Salmonella typhimurium. Genes Genet Syst 74:105–111. doi: 10.1266/ggs.74.105. [DOI] [PubMed] [Google Scholar]
- 74.Kutsukake K, Iino T. 1994. Role of the FliA-FlgM regulatory system on the transcriptional control of the flagellar regulon and flagellar formation in Salmonella typhimurium. J Bacteriol 176:3598–3605. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Yamamoto S, Kutsukake K. 2006. FliT acts as an anti-FlhD2C2 factor in the transcriptional control of the flagellar regulon in Salmonella enterica serovar Typhimurium. J Bacteriol 188:6703–6708. doi: 10.1128/JB.00799-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Saini S, Brown JD, Aldridge PD, Rao CV. 2008. FliZ Is a posttranslational activator of FlhD4C2-dependent flagellar gene expression. J Bacteriol 190:4979–4988. doi: 10.1128/JB.01996-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Wada T, Hatamoto Y, Kutsukake K. 2012. Functional and expressional analyses of the anti-FlhD4C2 factor gene ydiV in Escherichia coli. Microbiology 158:1533–1542. doi: 10.1099/mic.0.056036-0. [DOI] [PubMed] [Google Scholar]
- 78.Koirala S, Mears P, Sim M, Golding I, Chemla YR, Aldridge PD, Rao CV. 2014. A nutrient-tunable bistable switch controls motility in Salmonella enterica serovar Typhimurium. mBio 5:e01611-14. doi: 10.1128/mBio.01611-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Stewart MK, Cookson BT. 2014. Mutually repressing repressor functions and multi-layered cellular heterogeneity regulate the bistable Salmonella fliC census. Mol Microbiol 94:1272–1284. doi: 10.1111/mmi.12828. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Frankel NW, Pontius W, Dufour YS, Long J, Hernandez-Nunez L, Emonet T. 2014. Adaptability of non-genetic diversity in bacterial chemotaxis. eLife 3:e03526. doi: 10.7554/eLife.03526. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Paulick A, Delalez NJ, Brenzinger S, Steel BC, Berry RM, Armitage JP, Thormann KM. 2015. Dual stator dynamics in the Shewanella oneidensis MR-1 flagellar motor. Mol Microbiol 96:993–1001. doi: 10.1111/mmi.12984. [DOI] [PubMed] [Google Scholar]
- 82.Müller S, Strack SN, Hoefler BC, Straight PD, Kearns DB, Kirby JR. 2014. Bacillaene and sporulation protect Bacillus subtilis from predation by Myxococcus xanthus. Appl Environ Microbiol 80:5603–5610. doi: 10.1128/AEM.01621-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Müller S, Strack SN, Ryan SE, Kearns DB, Kirby JR. 2015. Predation by Myxococcus xanthus induces Bacillus subtilis to form spore-filled megastructures. Appl Environ Microbiol 81:203–210. doi: 10.1128/AEM.02448-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Zhao X, Zhang K, Boquoi T, Hu B, Motaleb MA, Miller KA, James ME, Charon NW, Manson MD, Norris SJ, Li C, Liu J. 2013. Cryoelectron tomography reveals the sequential assembly of bacterial flagella in Borrelia burgdorferi. Proc Natl Acad Sci U S A 110:14390–14395. doi: 10.1073/pnas.1308306110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Chen S, Beeby M, Murphy GE, Leadbetter JR, Hendrixson DR, Briegel A, Li Z, Shi J, Tocheva EI, Muller A, Dobro MJ, Jensen GJ. 2011. Structural diversity of bacterial flagellar motors. EMBO J 30:2972–2981. doi: 10.1038/emboj.2011.186. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Murphy GE, Leadbetter JR, Jensen GJ. 2006. In situ structure of the complete Treponema primitia flagellar motor. Nature 442:1062–1064. doi: 10.1038/nature05015. [DOI] [PubMed] [Google Scholar]
- 87.Raddi G, Morado DR, Yan J, Haake DA, Yang XF, Liu J. 2012. Three-dimensional structures of pathogenic and saprophytic Leptospira species revealed by cryo-electron tomography. J Bacteriol 194:1299–1306. doi: 10.1128/JB.06474-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Zhao X, Norris SJ, Liu J. 2014. Molecular architecture of the bacterial flagellar motor in cells. Biochemistry 53:4323–4333. doi: 10.1021/bi500059y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Abrusci P, Vergara-Irigaray M, Johnson S, Beeby MD, Hendrixson DR, Roversi P, Friede ME, Deane JE, Jensen GJ, Tang CM, Lea SM. 2013. Architecture of the major component of the type III secretion system export apparatus. Nat Struct Mol Biol 20:99–104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Ibuki T, Imada K, Minamino T, Kato T, Miyata T, Namba K. 2011. Common architecture of the flagellar type III protein export apparatus and F- and V-type ATPases. Nat Struct Mol Biol 18:277–282. doi: 10.1038/nsmb.1977. [DOI] [PubMed] [Google Scholar]
- 91.Imada K, Minamino T, Tahara A, Namba K. 2007. Structural similarity between the flagellar type III ATPase FliI and F1-ATPase subunits. Proc Natl Acad Sci U S A 104:485–490. doi: 10.1073/pnas.0608090104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Takekawa N, Kojima S, Homma M. 2014. Contribution of many charged residues at the stator-rotor interface of the Na+-driven flagellar motor to torque generation in Vibrio alginolyticus. J Bacteriol 196:1377–1385. doi: 10.1128/JB.01392-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Sircar R, Borbat PP, Lynch MJ, Bhatnagar J, Beyersdorf MS, Halkides CJ, Freed JH, Crane BR. 2015. Assembly states of FliM and FliG within the flagellar switch complex. J Mol Biol 427:867–886. doi: 10.1016/j.jmb.2014.12.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Minamino T. 2014. Protein export through the bacterial flagellar type III export pathway. Biochim Biophys Acta 1843:1642–1648. doi: 10.1016/j.bbamcr.2013.09.005. [DOI] [PubMed] [Google Scholar]