

Abstract
We found a novel spontaneous mouse mutant with depigmentation in the ventral body, which we called White Spotting (WS) mouse. Genetic investigation revealed deletion of a > 1.2-Mb genomic region containing nine genes (Kit, Kdr, Srd5a3, Tmeme165, Clock, Pdcl2, Nmu, Exoc1, and Cep135). We designated this mutant allele KitWS. Interestingly, homozygous mutants (KitWS/WS) showed a peri-implantation lethal phenotype. Expression analyses of these nine genes in blastocysts suggested that Exoc1 was a prime candidate for this phenotype. We produced Exoc1 knockout mice, and the same peri-implantation lethal phenotype was seen in Exoc1−/− embryos. In addition, the polygenic effect without Exoc1 was investigated in genome-edited KitWE mice carrying the Mb-scale deletion induced by the CRISPR/Cas9 system. As KitWE/WE embryos did not exhibit the abnormal phenotype, which was seen in KitWS/WS. We concluded that peri-implantation lethality in KitWS/WS was caused by a monogenic defect of Exoc1.
There have been many reports of spontaneous mouse mutants with depigmentation phenotypes1, among which Kit gene mutant mouse strains have been investigated in detail. More than 150 Kit mutant strains, including 72 spontaneous mutants, are listed in the Mouse Genome Informatics (MGI) database (http://www.informatics.jax.org). The Kit protein consists of an intracellular tyrosine kinase domain and an extracellular domain containing five immunoglobulin-like repeats2. The KitW gene, with a spontaneous point mutation resulting in the skipping of a critical exon, encodes a non-functional Kit protein lacking the extracellular domain3. Mice heterozygous for the KitW mutation (Kit+/W) show depigmentation in the ventral body, and KitW/W homozygotes die around the first week of life4. Several large-scale mutations have been found around the Kit gene region. The KitW–57J allele shows a deletion of approximately 80 kb in the upstream region of the Kit gene5. Moreover, megabase (Mb)-scale mutations were found in KitW–bd (2.8 Mb inversion)5 and KitW–19H (>3.5 Mb deletion)6. More than 20 genes, including Kit, are completely deleted in the KitW–19H allele. Interestingly, KitW–19H/W–19H mice die at the peri-implantation stage. Therefore, one or more of these 20 genes is/are thought to be the gene(s) responsible for this phenotype. However, the causative gene has not yet been conclusively identified.
The exocyst complex plays a critical role in exocytosis, which is a vesicle transportation event involved in provision of plasma membrane proteins and lipids to the plasma membrane and secretion of soluble vesicle components, such as hormones and cytokines. The exocyst complex is composed of eight proteins: Exoc 1, 2, 3, 4, 5, 6, 7, and 87. As several exocyst complex-related proteins, such as Arp3 and desmosomal proteins (Dsp, Dsc3, and Dsg2), are essential for early embryo development8,9,10,11, exocyst components are predicted to play important roles at the early embryonic stage. The in vivo functions of only two of eight exocyst components have been investigated in gene-modified mouse models. Exoc4 null mutant mice showed embryonic lethality at embryonic day 10.5 due to abnormal mesoderm formation12. In addition, Exoc6 spontaneous mutant mice exhibit severe microcytic anemia13. However, the in vivo functions of the other exocyst components are unclear.
Here, we report a de novo spontaneous Mb-scale mutant allele, KitWS, with a genomic deletion of at least 1.2 Mb containing nine genes (Kit, Kdr, Srd5a3, Tmeme165, Clock, Pdcl2, Nmu, Exoc1, and Cep135). As KitWS/WS showed a peri-implantation lethal phenotype, we attempted to identify the causative gene for this phenotype. Gene expression analysis in blastocysts suggested Exoc1 as the first candidate gene. We produced and analyzed an Exoc1 knockout mouse strain to determine the in vivo function of this gene. As expected, the abnormal phenotype of Exoc1 null embryos was the same as that of KitWS/WS. Finally, we introduced the Mb-scale deletion allele (the region from Kit to Nmu) in mice using the CRISPR/Cas9 system. This introduced mutation did not affect early embryonic development. These data indicate that Exoc1, which is located in the vicinity of the Kit gene, is the monogenic causative gene for peri-implantation lethality.
Results
Appearance of the novel natural mutant with depigmentation
A male mouse generated from a pair of C57BL/6J (B6J) obtained from Charles River Laboratories Japan exhibited depigmentation in the ventral body (Fig. 1A) and the tip of the tail and foot (Fig. 1B,C). We called this White Spotting (WS) mouse. These abnormalities were not observed in its male or female siblings. To examine whether the abnormal pigmentation was heritable, we carried out test crosses between the WS male and wild-type females. Of 20 progeny, nine mice (3 females and 6 males) showed depigmentation, which suggested that the abnormal phenotype is inherited in an autosomal dominant manner.
Figure 1. Depigmentation in WS mouse.

Depigmentation phenotypes in the ventral body (A) and the tips of the feet (B) and tail (C) were seen in WS mice.
Identification of deleted genome region
As Kit gene mutant mouse strains were reported to show depigmentation with an autosomal dominant inheritance pattern14,15, we postulated that the abnormal pigmentation in WS mice is caused by a de novo Kit gene mutation. Kit protein expression in WS was confirmed by western blotting. As Kit is expressed in the brain16, we examined Kit expression in the cerebrum and cerebellum from wild-type and WS adults. As expected, very weak Kit signals were observed in WS mice (Fig. 2A). Next, we performed fluorescence in situ hybridization (FISH) to investigate genomic mutation on the Kit gene locus (5qc3.3). Bacterial artificial chromosome (BAC) DNA vectors were used as probes. The BAC RP23-142L11 (derived from Chr. 5: 75,533,600–75,737,738), which carries the full-length Kit gene region (Chr. 5: 75,574,987–75,656,721), was labelled with Cy-3. As a marker for chromosome 5, BAC RP23-153I24 (derived from Chr. 5: 98,182,487–98,393,147), which carries the full-length Fgf5 gene region (Chr. 5: 98,254,184–98,277,033), was labelled with FITC. These two probes were hybridized to WS chromosomal spreads. Although FITC (green) signals were observed on both copies of chromosome 5, Cy-3 (red) signals were observed on only one copy of chromosome 5. These results suggested that the depigmentation phenotype in WS mice is caused by heterozygous chromosome breakage of 5qc3.3, which includes the Kit gene.
Figure 2. Kit gene expression and location.
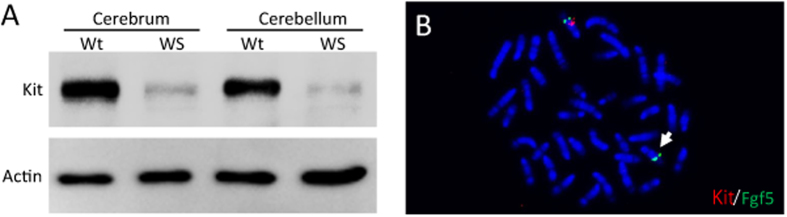
(A) Kit protein was detected on Western blotting of the cerebrum and cerebellum. The signals of Kit in WS mice were weaker than in wild-type. (B) FISH to metaphase chromosome spreads of WS mice. FITC (green) signals, which reflected the Fgf5 gene locus, were observed on both copies of chromosome 5. In contrast, Cy3 (red) signals, reflecting the Kit gene locus, were only seen on one copy (white arrow).
To identify the deletion region in WS by polymorphism analysis, F1 progeny with normal pigmentation (C3B6F1-WT) and depigmentation (C3B6F1-WS) were produced by crossing WS with C3H/HeJ (C3H) mice (Fig. 3A). In C3B6F1-WS mice, there are no B6J allelic polymorphisms in the deletion region derived from the WS genome. On the other hand, both B6 and C3H allelic polymorphisms were detected in all regions of the genome in C3B6F1-WT. The presence of the B6J genomic region in C3B6F1-WS mice was examined by single nucleated polymorphism (SNPs) analyses. At the rs29824030 SNP which is located in the Kit gene region, we found the C3H genotype only in C3B6F1-WS mice (Fig. 3B). Moreover, we then analyzed SNPs in 5qc3.3. Surprisingly, SNPs from B6J were not detected on rs33458703, rs31548445, rs31561146, or rs33566798 (Table 1 and Fig. 3C). These results indicated that a genomic region of at least 1.2 Mb containing nine genes (Kit, Kdr, Srd5a3, Tmeme165, Clock, Pdcl2, Nmu, Exoc1, and Cep135) is deleted in WS mice (Fig. 3C & D). This de novo large mutant allele was designated as KitWS.
Figure 3. Identification of deletion region in WS.

(A) C3B6F1-WT and C3B6F1-WS mice were obtained by mating C3H wild-type and WS (C57BL/6 genetic background). The depigmentation phenotype was only seen in C3B6F1-WS. (B) Waveform of SNP rs29824030. Double peaks (Guanine (G) from C57BL/6J and Adenine (A) from C3H) were detected in C3B6F1-WT. The single A peak was detected in C3B6F1-WS. (C) Illustration of the sites of SNPs analyzed in the present study. The arrows indicate the SNP sites detected by double peaks in C3B6F1-WS. The arrowhead indicates the single peak. (D) Physical maps of the KitW–19H, KitWS, and KitWE alleles. The solid line, dashed line, and blank indicate undeleted, unknown, and deleted genomic regions, respectively. *The map of KitW–H19 is referenced from the report of Nagle41.
Table 1. SNP analysis with C3B6F1-WS.
| gene | SNP | Locus | C57BL/6J | C3H/HeJ | C3B6F1-WT | C3B6F1-WS |
|---|---|---|---|---|---|---|
| Pdgfra | rs13478329 | 75,192,475 | G | A | G/A | G/A |
| Pdgfra-Kit | rs33555487 | 75,205,382 | C | A | C/A | C/A |
| Kit | rs29824030 | 75,581,667 | G | A | G/A | A |
| Kdr | rs33458703 | 75,943,660 | A | G | A/G | G |
| Clock | rs31548445 | 76,216,518 | C | T | C/T | T |
| Cep135 | rs31561146 | 76,627,052 | G | A | G/A | A |
| Cep135-Aasdh I | rs33566798 | 76,802,546 | T | C | T/C | C |
| Cep135-Aasdh II | rs33892425 | 76,820,488 | G | C | G/C | G/C |
| Aasdh | rs6257272 | 76,887,200 | C | T | C/T | C/T |
Peri-implantation lethality involving Kit WS
King et al.6 reported a genomic deletion of more than 3.5 Mb (Chr. 5: 73.2 − 76.9 Mb) in the KitW–19H mutant allele (Fig. 3D). Interestingly, homozygous KitW–19H mutants (KitW–19H/W–19H) died around the peri-implantation stage, although Kit homozygous null mutant mice (KitW/W) survived until birth4. These reports suggest that the causative gene(s) for peri-implantation lethality is/are located around the Kit gene. To understand the characteristics of KitWS/WS during embryonic development, Kit+/WS mice were intercrossed. We first analyzed the Kit genotypes in pre-implantation embryos on embryonic day (E) 3.5. Five of 33 blastocysts were KitWS/WS and showed no morphological abnormalities. In contrast, no KitWS/WS E7.5 embryos (0/24) were found (Table 2). To confirm the embryonic lethality from the 2-cell to blastocyst stages, we collected 2-cell stage embryos from Kit+/WS intercrosses. Forty-four 2-cell stage embryos were collected from oviducts and cultured in KSOM medium. All embryos were developed to blastocyst. These results indicated that homozygous KitWS mutants die at the peri-implantation stage.
Table 2. Embryos from Kit+/WS intercross.
| Stage | KitWS/WS | Kit+/WSor Kit+/+ | Total |
|---|---|---|---|
| E3.5 | 5 (15%) | 28 (85%) | 33 |
| E7.5 | 0 (0%) | 24 (100%) | 24 |
Genetic abnormalities in blastocysts are sometimes obscured by maternal factors8,17. To analyze the KitWS/WS phenotype in detail without the effects of maternal factors, we performed in vitro cultivation of blastocysts for 1 week. In cultivated blastocysts, embryonic stem (ES) cells and trophoblast giant (TG) cells arise from the inner cell mass (ICM) and trophectoderm (TE), respectively18,19. Thirty-three blastocysts derived from Kit+/WS intercrosses were cultured on gelatine-coated dishes. In wild-type or Kit+/WS blastocysts, both ES cell-like dome-shaped colonies and TG-like cells with an enlarged nucleus were observed (Fig. 4A). Interestingly, no ES cell-like colonies developed from KitWS/WS even though TG-like cells were observed (Fig. 4B and Table 3). These results suggest that the early embryonic death in KitW/WS is caused by defects in ICM proliferation.
Figure 4. Cultivated KitWS/WS blastocysts.
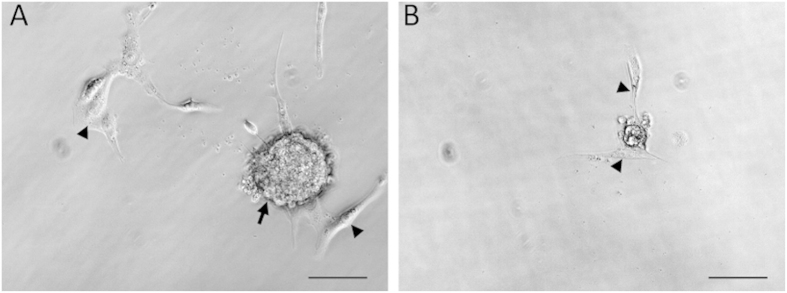
(A) Dome-shaped ES cell-like colonies (arrow) and TG-like cells (arrowheads) were observed in cultured blastocysts derived from Kit+/+ or Kit+/WS. (B) In contrast, although TG cells were seen (arrowheads), no ES cell-like colonies were seen in KitWS/WS. Scale bar = 100 μm.
Table 3. Cultured blastocyst derived from Kit+/WS intercross.
| Genotype | ICMoutgrowth | No ICMoutgrowth | Total |
|---|---|---|---|
| Kit+/WS or Kit+/+ | 23 (82%) | 5 (18%) | 28 |
| KitWS/WS | 0 (0%) | 5 (100%) | 5 |
Association between peri-implantation lethality and Exoc1 mutation
To narrow down the number of candidate causative genes for peri-implantation lethality, we examined the blastocyst expression of nine genes located in the region of the deletion in KitWS (Fig. 3C). On RT-PCR analysis with wild-type blastocysts, signals were detected for Kit, Srd5a3, Tmem165, and Exoc1, but not Kdr, Clock, Nmu, Pdcl2, or Cep135 (Fig. 5), suggesting that one or more of the former four genes (Kit, Srd5a3, Tmem165, and Exoc1) may be related to peri-implantation lethality.
Figure 5. RT-PCR of blastocyst.
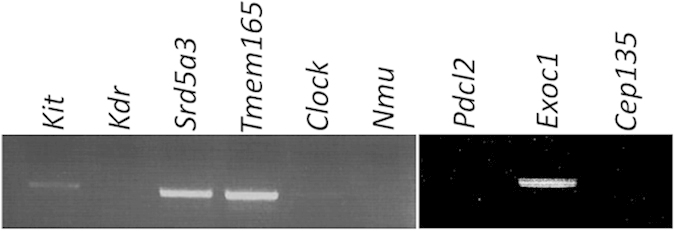
Kit, Srd5a3, Tmeme165, and Exoc1 signals were detected on RT-PCR analysis of nine candidate genes using RNAs from wild-type blastocysts.
It has been reported that neither Kit, Srd5a3, nor Tmem165 knockout mice died before E7.54,20,21. We first examined Exoc1 as the candidate gene. To produce Exoc1 null mutant mice, an Exoc1 gene-targeted ES cell line (#78575) was obtained from the European Conditional Mouse Mutagenesis Program (EUCOMM), and heterozygous Exoc1 mutants (Exoc1−/+) were successfully generated from the ES cells. In Exoc1−/+, the proper targeting allele was confirmed by Southern blotting analysis (Fig. 6A) and Exoc1 protein expression level was shown to be decreased (Fig. 6B).
Figure 6. Exoc1 knockout mouse.

(A) Southern blotting of Exoc1−/+ and Exoc1+/+ mice. MfeI-digested DNA fragments were detected by 5′ outer and LacZ inner probes, and the BamHI-digested DNA fragments were detected by 3′ outer probes. (B) Western blotting showed that Exoc1 protein expression was reduced in Exoc1−/+ mice. (C–E) Cultivated blastocysts from Exoc1−/+ intercrosses. TG-like cells with an enlarged nucleus (arrowheads) were seen in cultivated embryos of either genotype. Dome-shaped colonies (arrows) were not seen in Exoc1−/−. M, MfeI; Ba, BamHI; Scale bar = 100 μm.
As expected, no Exoc1−/− embryos (0/30) were found at the E7.5 stage (Table 4). To investigate whether Exoc1 plays a critical role in ICM growth of post-blastocyst embryos, we performed in vitro cultivation of blastocysts derived from Exoc1−/+ intercrosses. Interestingly, no ES cell-like dome-shaped colonies were observed in any Exoc1−/− blastocysts (9/9) despite the development of TG-like cells (Fig. 6E and Table 5). In contrast, both ES and TG-like cells were observed in Exoc1−/+ and Exoc1+/+ embryos. To investigate whether Exoc1 is only expressed cells from ICM, we then performed X-gal staining. As expected, the blue color signals were detected in ES cell-like dome-shaped colonies and ES cells but not in TG-like cells (Supplemental Fig. 1). These results indicated that Exoc1 is essential for peri-implantation development, especially for ICM outgrowth in mouse embryos.
Table 4. Embryos from Exoc1−/+ intercross.
| Stage | Exoc1−/− | Exoc1−/+ | Exoc1+/+ | Total |
|---|---|---|---|---|
| E7.5 | 0 (0%) | 24 (80%) | 6 (20%) | 30 |
Table 5. Cultured blastocyst derived from Exoc1−/+ intercross.
| Genotype | ICM outgrowth | No ICMoutgrowth | Total |
|---|---|---|---|
| Exoc1+/+ | 12 (63%) | 7 (37%) | 19 |
| Exoc1−/+ | 10 (48%) | 11 (52%) | 21 |
| Exoc1−/− | 0 (0%) | 9 (100%) | 9 |
Lack of association between peri-implantation lethality and Mb-scale deletion (from Kit to Nmu)
The detection of not only Exoc1 but also Kit, Srd5a3, and Tmem165 mRNAs in blastocysts (Fig. 5) suggested that the peri-implantation lethality may also be caused by polygenic effects involving these genes. Therefore, we attempted to produce new gene-modified mice carrying a large deletion mutation extending from Kit to Nmu, but not Exoc1, by CRISPR/Cas9 system.
Two CRISPR targets located 240 kb upstream of Kit and 81 kb downstream of the Nmu gene region were designed separated by a distance of approximately 1.1 Mb (Fig. 7A). Both targeted CRISPR expression vectors were co-microinjected into the pronuclei of 54 fertilized B6J oocytes, and nine neonates were obtained. As expected, one founder mouse exhibited abnormal pigmentation similar to that in KitWS/+ mice (Fig. 7B). We then confirmed the genomic sequence around the target sites in this mouse. Sequence analysis revealed deletion of a 1.07-Mb genomic region (Chr. 5: 75,804,497–76,875,127) in the founder mouse (Fig. 7C). We named this induced large mutant allele KitWE (Fig. 3D). The phenotype of KitWE/WE was examined by in vitro cultivation of blastocysts derived from Exoc1WE/+ intercrosses. In contrast to KitWS/WS and Exoc1−/−, ES cell-like colonies developed from KitWE/WE blastocytes (6/8) (Fig. 7D and Table 6). These results indicated that the loss of three genes (Kit, Srd5a3, and Tmem165) expressed in the blastocyst was not associated with the failure of embryonic development at the peri-implantation stage. Therefore, we concluded that Exoc1 is the causative gene for peri-implantation lethality in KitWS/WS.
Figure 7. KitWE genome edited mice.
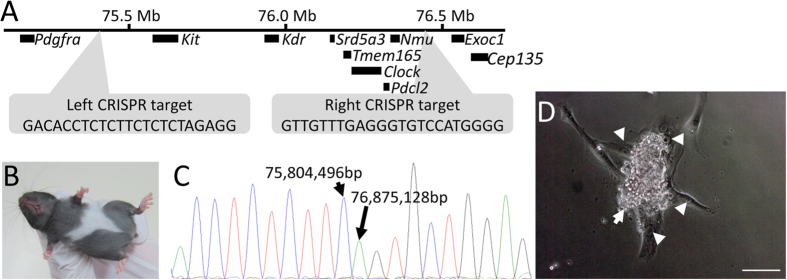
(A) Illustration of CRISPR/Cas9 target sites and genetic regions of candidate genes. The left CRISPR target site was located on Chr. 5: 75,408,459–75,408,481 and the right target site was located on Chr. 5: 76,444,361–76,444,383. The genomic region between left and right targets contained the seven genes, Kit, Kdr, Srd5a3, Tmeme165, Clock, Pdcl2, and Nmu. (B) The depigmentation phenotype of KitWE/+ mice was the same as KitWS/+. (C) Genomic sequencing analysis exhibited that the genetic region between 75,804,497 and 76,875,127 on Chr. 5 was deleted. (D) Cultivated KitWE/WE blastocyst. Both ES (arrow) and TS (arrowheads) like-cells were seen. Sale Bar = 100 μm.
Table 6. Cultured blastocyst derived from Kit+/WE intercross.
| Genotype | ICM outgrowth | No ICMoutgrowth | Total |
|---|---|---|---|
| Kit+/+ | 11 (92%) | 1 (8%) | 12 |
| Kit+/WE | 12 (55%) | 10 (45%) | 22 |
| KitWE/WE | 6 (75%) | 2 (25%) | 8 |
Discussion
In this study, we found a novel spontaneous mutant mouse with ventral body depigmentation. The results of polymorphism analyses indicated deletion of a genomic region of more than 1.2 Mb containing nine genes (from Kit to Cep135) in the KitWS mutant allele. In blastocyst cultivation experiments, inhibition of ICM outgrowth was seen in both KitWS/WS and Exoc1−/−, but not KitWE/WE. These results suggested that peri-implantation lethality in KitWS/WS is caused by monogenic Exoc1 gene deletion.
In polymorphism analyses with C3B6F1-WS (Table 1 and Fig. 3), only C3H alleles were detected in five SNPs located between Kit and Cep135. These data clearly indicated that the nine genes are completely deleted in KitWS, although the deletion ends were not determined. As depigmentation phenotypes in laboratory mice can be found easily, a great variety of genetic mutations in and the around Kit gene region have been reported22. However, the KitWS differs from other Kit mutant alleles. Therefore, the KitWS is considered to be a de novo spontaneous mutation allele.
Of the nine candidate genes (Kit, Kdr, Srd5a3, Tmeme165, Clock, Pdcl2, Nmu, Exoc1, and Cep135), the biological functions of six (Kit, Kdr, Srd5a3, Tmeme165, Clock, and, Nmu) have been analyzed using knockout mice. As described above, Kit null mutant mice (KitW/W) die around the first week of life4. Kdr knockout mice die between E8.5 and E9.5 due to failure of blood island formation and vasculogenesis23. Srd5a3 knockout mice die between E11.5 and E13.5 with open neural tubes20. No embryonic lethal phenotypes were seen in knockout mice for each of the three genes, Tmem16521, Clock24, and Nmu25. Although there have been no previous reports regarding Cep135 null mutant mice, a 1-bp deletion mutation of CEP135 was found in humans. This c.970delC mutation generates a non-functional truncated CEP135 protein. Patients homozygous for the c.970delC mutation can survive26. Lopez et al. reported that Pdcl2 was only expressed in germline cells27. Pdcl2 mRNA was not detected by RT-PCR in wild-type blastocysts in the present study (Fig. 5). Based on these previous reports and our observations, Exoc1 emerged as the most likely candidate responsible for the observed peri-implantation embryonic lethal phenotype. In accordance with this hypothesis, the present study exhibited that the abnormal phenotype of Exoc1−/− embryos was the same as that of KitWS/WS.
Exoc1, which was initially found in the yeast secretory pathway28, is a component of the exocyst complex29. The exocyst complex plays an important role in the tethering step in exocytosis30. As cell membrane proteins and lipids are transported to the cell membrane by exocytosis, the exocyst complex is involved in not only vesicle secretion but also cell adhesion and migration7. In fact, Andersen et al. reported that the Exoc1-containing exocyst complex was required for assembly of the desmosome, which is one of the key structures in cell–cell adhesion31. Furthermore, the exocyst complex is involved in dynamic remodelling of the actin cytoskeleton by regulating Arp2/3-mediated actin polymerization32. The peri-implantation lethal phenotypes of mutations in desmosome-related genes (Dsp, Dsc3, and Dsg2) and Arp3 knockout mice are similar to that of Exoc1−/−, suggesting that Exoc1 may function in desmosome assembly and/or remodelling of the actin cytoskeleton in early embryo development8,9,10,11. In addition, 15% KitWS/WS (Table 2) and 11% Exoc1−/− (Table 5) blastocysts were obtained from Kit+/WS and Exoc1−/− intercross, respectively, indicating lower Mendelian ratio than expected. However, all 2-cell embryos from Kit+/WS intercross showed no developmental failure to the blastocyst stage in vitro (data not shown). Therefore, we might also need to investigate the function of Exoc1 in early embryo development prior to the 2-cell stage or in gamete development in future.
We examined the polygenic effects in mice carrying the KitWE large deletion induced by the CRISPR/Cas9 system. While the seven genes (from Kit to Nmu) were completely deleted in KitWE/WE blastocysts (Fig. 3D), ICM outgrowth was normal (Fig. 7D). These observations indicated that the seven genes, including Kit, Srd5a3, and Tmem165, expressed in the blastocyst were not involved in monogenic or polygenic defects during peri-implantation embryonic development. In addition, we clearly showed that the CRISPR/Cas9 system was able to induce Mb-scale deletion by zygote microinjection. This technology is expected to produce mice with precise large deletions for investigation of various human diseases associated with chromosome breakage.
In conclusion, we discovered that Exoc1 null mutation is the cause of peri-implantation lethality. Although the molecular mechanisms of action of Exoc1 are unclear, our findings will contribute to understanding of early embryonic development.
Materials and Methods
Animals
Laboratory mice were purchased from Charles River Laboratories Japan. Animal experiments were carried out as described previously33,34. All animal experiments were carried out humanely in accordance with the Regulations for Animal Experiments of the University of Tsukuba and Fundamental Guidelines for Proper Conduct of Animal Experiment and Related Activities in Academic Research Institutions under the jurisdiction of the Ministry of Education, Culture, Sports, Science, and Technology of Japan and with approval from the Institutional Animal Experiment Committee of our university.
Western blotting
Western blotting analyses were performed according to standard protocols with antibodies to Kit (3074: CST), Actin (P68133: Millipore), Exoc1 (11690-1-AP: Protein Technologies Ltd.), and GAPDH (FL335: Santa Cruz).
Genomic PCR & sequencing
Investigation of SNPs and detection of the deletion region in the KitWE allele were performed with PrimeSTAR® GXL DNA Polymerase (Takara) and the primers listed in Supplemental Table 1. An Applied Biosystems® 3500 Genetic Analyzer and a BigDye® Terminator v3.1 Cycle Sequencing Kit (Thermo Fisher Scientific) were used to confirm DNA sequences of PCR products, which were purified with a Fast Gene Gel/PCR Extraction Kit (Nippon Genetics).
FISH
Generation FITC- and Cy3-labelled BAC DNA probes, preparation of chromosome spreads from KitWS/+ mice, and probe–chromosome spread hybridization were conducted as described35.
Embryo manipulation & blastocyst cultivation
Superovulation was induced by injection of pregnant mare serum gonadotropin (PMSG) and human chorionic gonadotropin (hCG) with an interval of 48 hours, followed by mating with male mice. The 2-cell embryos were collected by oviduct flushing and cultured in KSOM medium. The zona pellucida of the blastocyst was removed with acid Tyrode’s solution. Then, blastocysts were cultured in medium as described previously36 on gelatine-coated dishes.
RT-PCR
RNA was collected from approximately 100 blastocysts with Sepasol® (Nakalai Tesque) according to the manufacturer’s protocol. cDNA synthesis was performed as described33. PCR was performed with AmpliTaq Gold PCR Master Mix (Thermo Fisher Scientific) and various primers (Supplemental Table 2).
Production of Exoc1 KO mice
The Exoc1 gene-targeted mouse ES cells were provided by EUCOMM, International Mouse Phenotyping Consortium (IMPC). The chimeric mice were developed by aggregation of ES cells with morula embryos derived from ICR mice. Southern blotting was conducted as described35. X-gal staining of cultivated blastocysts and ES cells were conducted as described37.
CRISPR/Cas9
The px330 plasmid (Addgene #42230), used as a CRISPR expression vector, was a gift from Dr. Feng Zhang38. The left and right 20-nt target sequences (Supplemental Table 3) were inserted into px330. The EGxxFP assay39 with pCAG-EGxxFP (Addgene #50716) to check cleavage activities of these two vectors was performed (Supplemental Fig. 2) as decribed34. These vectors (5 ng/μl each, circled) were co-microinjected into the pronuclei of one-cell-stage embryos derived from C57BL/6J, and the injected embryos were transferred into pseudopregnant ICR mice according to standard protocols40.
Additional Information
How to cite this article: Mizuno, S. et al. Peri-implantation lethality in mice carrying megabase-scale deletion on 5qc3.3 is caused by Exoc1 null mutation. Sci. Rep. 5, 13632; doi: 10.1038/srep13632 (2015).
Supplementary Material
Acknowledgments
This work was supported by Grants-in-Aid for Scientific Research (B) (to K.Y., S.M. and F.S.) and Grant-in-Aid for Young Scientists (B) (to S.M.) from the Ministry of Education, Culture, Sports, Science, and Technology, Japan. We thank the members of the Yagami Laboratory for helpful discussions and encouragement.
Footnotes
Author Contributions F.S. designed the study and managed the project. S.M. and T.K. performed most experiments. D.Y., T.Y., T.T.H.D., S.M.-I., Y.H. and S.T. generated mutant mice. S.M., F.S. and K.Y. wrote the manuscript and all authors discussed the results and commented on the manuscript.
References
- Bennett D. C. & Lamoreux M. L. The color loci of mice–a genetic century. Pigment Cell Res. 16, 333–344 (2003). [DOI] [PubMed] [Google Scholar]
- Yarden Y. et al. Human proto-oncogene c-kit: a new cell surface receptor tyrosine kinase for an unidentified ligand. EMBO J. 6, 3341–3351 (1987). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hayashi S. et al. Exon skipping by mutation of an authentic splice site of c-kit gene in W/W mouse. Nucleic Acids Res. 25, 1267–1271 (1991). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Russell E. S. & Lawson F. A. Selection and inbreeding for longevity of a lethal type. J. Hered. 50, 19–25 (1959). [Google Scholar]
- Klüppel M. et al. Long-range genomic rearrangements upstream of Kit dysregulate the developmental pattern of Kit expression in W57 and Wbanded mice and interfere with distinct steps in melanocyte development. Development. 124, 65–77 (1997). [DOI] [PubMed] [Google Scholar]
- King D. P. et al. The mouse Clock mutation behaves as an antimorph and maps within the W19H deletion, distal of Kit. Genetics. 146, 1049–60 (1997). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu J. & Guo W. The exocyst complex in exocytosis and cell migration. Protoplasma. 249, 587–597 (2012). [DOI] [PubMed] [Google Scholar]
- Vauti F. et al. Arp3 is required during preimplantation development of the mouse embryo. FEBS Lett. 581, 5691–5697 (2007). [DOI] [PubMed] [Google Scholar]
- Gallicano G. I. et al. Desmoplakin is required early in development for assembly of desmosomes and cytoskeletal linkage. J Cell Biol. 143, 2009–2022 (1998). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Den Z. et al. Desmocollin 3 is required for pre-implantation development of the mouse embryo. J Cell Sci. 119, 482–489 (2006). [DOI] [PubMed] [Google Scholar]
- Eshkind L. et al. Loss of desmoglein 2 suggests essential functions for early embryonic development and proliferation of embryonal stem cells. Eur J Cell Biol. 81, 592–598 (2002). [DOI] [PubMed] [Google Scholar]
- Friedrich G. A. et al. The secretory protein Sec8 is required for paraxial mesoderm formation in the mouse. Dev Biol. 192, 364–374 (1997). [DOI] [PubMed] [Google Scholar]
- White R. A. et al. Iron metabolism mutant hbd mice have a deletion in Sec15l1, which has homology to a yeast gene for vesicle docking. Genomics. 86, 668–73 (2005). [DOI] [PubMed] [Google Scholar]
- Hou L. et al. Signaling and transcriptional regulation in the neural crest-derived melanocyte lineage: interactions between KIT and MITF. Development. 127, 5379–5389 (2000). [DOI] [PubMed] [Google Scholar]
- Takabayashi S. et al. A novel Kit gene mutation in CF1 mice involved in the extracellular domain of the KIT protein. Exp Anim. 61, 435–444 (2012). [DOI] [PubMed] [Google Scholar]
- Goldstein B. J. et al. Adult c-Kit(+) progenitor cells are necessary for maintenance and regeneration of olfactory neurons. J Comp Neurol. 523, 15–31 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sengupta J. & Ghosh D. Multi-level and multi-scale integrative approach to the understanding of human blastocyst implantation. Prog Biophys Mol Biol. 114, 49–60 (2014). [DOI] [PubMed] [Google Scholar]
- Evans M. J. & Kaufman M. H. Establishment in culture of pluripotential cells from mouse embryos. Nature. 292, 154–156 (1981). [DOI] [PubMed] [Google Scholar]
- Tanaka S. et al. Promotion of trophoblast stem cell proliferation by FGF4. Science. 282, 2072–2075 (1998). [DOI] [PubMed] [Google Scholar]
- Cantagrel V. et al. SRD5A3 is required for converting polyprenol to dolichol and is mutated in a congenital glycosylation disorder. Cell. 142, 203–217 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- International Mouse Phenotype Consortium, Phenotype association of Tmem169. Available at: https://www.mousephenotype.org/data/genes/MGI:894407, (Accessed: 14th July, 2015).
- Mouse Genome Informatics, Phenotype, Alleles & Disease Models Search for Kit. Available at: http://www.informatics.jax.org/allele/summary?markerId=MGI:96677, (Accessed: 14th July, 2015).
- Shalaby F. et al. Failure of blood-island formation and vasculogenesis in Flk-1-deficient mice. Nature. 376, 62–66 (1995). [DOI] [PubMed] [Google Scholar]
- Vitaterna M. H. et al. Mutagenesis and mapping of a mouse gene, Clock, essential for circadian behavior. Science. 264, 719–725 (1994). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hanada R. et al. Neuromedin U has a novel anorexigenic effect independent of the leptin signaling pathway. Nat Med. 10, 1067–1073 (2004). [DOI] [PubMed] [Google Scholar]
- Hussain M. S. et al. A truncating mutation of CEP135 causes primary microcephaly and disturbed centrosomal function. Am J Hum Genet. 90, 871–878 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lopez P. et al. A novel germ line-specific gene of the phosducin-like protein (PhLP) family. A meiotic function conserved from yeast to mice. J Biol Chem. 278, 1751–1757 (2003). [DOI] [PubMed] [Google Scholar]
- Novick P. et al. Identification of 23 complementation groups required for post-translational events in the yeast secretory pathway. Cell. 21, 205–215 (1980). [DOI] [PubMed] [Google Scholar]
- TerBush D. R. et al. The Exocyst is a multiprotein complex required for exocytosis in Saccharomyces cerevisiae. EMBO J. 15, 6483–6494 (1996). [PMC free article] [PubMed] [Google Scholar]
- Guo W. et al. Protein complexes in transport vesicle targeting. Trends Cell Biol. 10, 251–255 (2000). [DOI] [PubMed] [Google Scholar]
- Andersen N. J. & Yeaman C. Sec3-containing exocyst complex is required for desmosome assembly in mammalian epithelial cells. Mol Biol Cell. 21, 152–164 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu J. et al. The role of the exocyst in matrix metalloproteinase secretion and actin dynamics during tumor cell invadopodia formation. Mol Biol Cell. 20, 3763–3771 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mizuno S. et al. A novel locus on proximal chromosome 18 associated with agenesis of the corpus callosum in mice. Mamm Genome. 21, 525–533 (2010). [DOI] [PubMed] [Google Scholar]
- Mizuno S. et al. Simple generation of albino C57BL/6J mice with G291T mutation in the tyrosinase gene by the CRISPR/Cas9 system. Mamm Genome. 25, 327–334 (2014). [DOI] [PubMed] [Google Scholar]
- Mizuno S. et al. Truncated Cables1 causes agenesis of the corpus callosum in mice. Lab Invest. 94, 321–330 (2014). [DOI] [PubMed] [Google Scholar]
- Tanimoto Y. et al. Embryonic stem cells derived from C57BL/6J and C57BL/6N mice. Comp Med. 58, 347–52 (2008). [PMC free article] [PubMed] [Google Scholar]
- Hasegawa Y. et al. Generation and Characterization of Ins1-cre-driver C57BL/6N for Exclusive Pancreatic Beta Cell-specific Cre-loxP Recombination. Exp Anim. 63, 183–191 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cong L. et al. Multiplex genome engineering using CRISPR/Cas systems. Science. 339, 819–23 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mashiko D. et al. Generation of mutant mice by pronuclear injection of circular plasmid expressing Cas9 and single guided RNA. Sci Rep. 3, 3355 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gordon J. W. & Ruddle F. H. Integration and stable germ line transmission of genes injected into mouse pronuclei. Science. 214, 1244–1246 (1981). [DOI] [PubMed] [Google Scholar]
- Nagle D. L. Structural analysis of chromosomal rearrangements associated with the developmental mutations Ph, W19H, and Rw on mouse chromosome 5. Proc Natl Acad Sci USA. 91, 7237–7241 (1994). [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.